엘라스모테리움
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
엘라스모테리움은 멸종된 코뿔소의 한 속으로, 1809년에 처음 명명되었다. 이 속은 유라시아 지역에서 발견되었으며, 특히 두개골 파편과 치아가 많이 발견되었다. 엘라스모테리움은 현존하는 코뿔소와는 다른 엘라스모테리움아과에 속하며, 마이오세 이후 멸종된 유일한 종이다. 가장 큰 종인 *E. caucasicum*은 5m에 달하는 몸길이와 이마에 뿔이 특징이며, 치아 구조와 골격 형태를 통해 초원 환경에 적응했음을 알 수 있다. 엘라스모테리움은 약 3만 6천 년 전에 멸종된 것으로 알려져 있으며, 플라이스토세 멸종 시기와 현생 인류의 이동과 관련이 있을 수 있다.
더 읽어볼만한 페이지
- 고대 코뿔소 - 털코뿔소
털코뿔소는 코엘로돈타속에 속하는 멸종된 코뿔소 종으로, 플라이스토세 동안 북부 유라시아 전역에 분포했으며, 두 개의 뿔과 두꺼운 털을 가지고 최대 2톤에 달하는 대형 초식 동물이었고, 마지막 빙하기 이후 멸종했다. - 유럽의 고대 포유류 - 털코뿔소
털코뿔소는 코엘로돈타속에 속하는 멸종된 코뿔소 종으로, 플라이스토세 동안 북부 유라시아 전역에 분포했으며, 두 개의 뿔과 두꺼운 털을 가지고 최대 2톤에 달하는 대형 초식 동물이었고, 마지막 빙하기 이후 멸종했다. - 유럽의 고대 포유류 - 털매머드
털매머드는 플라이스토세 후기부터 홀로세에 걸쳐 유라시아와 북아메리카에 서식했으며 털로 덮인 몸과 긴 상아를 가진 빙하기 대표 동물로, 아시아코끼리와 가장 가까운 친척 관계이며, 구석기 시대 인류와 공존하여 고생물학적 연구와 복원 시도가 이루어지고 있다.
| 엘라스모테리움 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Elasmotherium |
| 명명자 | J. Fischer, 1808년 |
| 모식종 | †Elasmotherium sibiricum |
| 분포 | Elasmotherium의 대략적인 분포도 |
| 생물학적 분류 | |
| 과 | 코뿔소과 |
| 속 | 엘라스모테리움속 (Elasmotherium) |
| 종 | †E. caucasicum †E. chaprovicum †E. peii †E. primigenium |
| 시대 | |
| 생존 시기 | 후기 마이오세 ~ 후기 플라이스토세 (약 700만 년 전 ~ 3만 9천 년 전) |
| 이명 | |
| 이명 | Stereoceros Enigmatherium E. fischeri = E. sibiricum E. inexpectatum = E. caucasicum |
2. 명칭
엘라스모테리움은 1809년 독일/러시아 고생물학자 고트헬프 피셔 폰 발트하임(Gotthelf Fischer von Waldheim)에 의해 처음 기술되었다. 그는 모스크바 대학교에 기증된 왼쪽 아래턱뼈, 네 개의 어금니, 세 번째 소구치의 치근을 근거로 1808년 모스크바 자연사 학회에서 발표했다.[4] 속명은 고대 그리스어 ''elasmos''("층상의")와 ''therion''("짐승")에서 유래하며, 법랑질의 층상 구조를 가리킨다. 종명 ''sibericus''는 소장품이 주로 시베리아에서 온 것에 대한 언급일 가능성이 높지만, 표본의 정확한 기원은 알려지지 않았다. 1877년, 독일의 자연주의자 요한 프리드리히 폰 브란트(Johann Friedrich von Brandt)는 이를 새롭게 설립된 엘라스모테리이네아(Elasmotheriinae)에 배치했다.[20]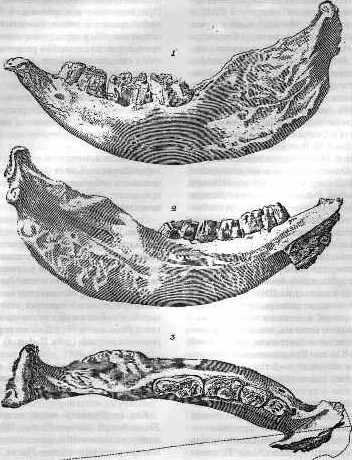
엘라스모테리움(Elasmotherium)은 1809년 독일/러시아 고생물학자 고트헬프 피셔 폰 발트하임(Gotthelf Fischer von Waldheim)에 의해 처음 기술되었다. 그 근거는 왼쪽 아래턱뼈, 네 개의 어금니, 그리고 세 번째 소구치의 치근으로, 1807년 예카테리나 다시코바(Ekaterina Dashkova) 공주가 모스크바 대학교에 기증한 것이었다. 그는 1808년 모스크바 자연사 학회에서 이를 처음 발표했다.[4] 속명은 고대 그리스어의 ''elasmos''("층상의")와 ''therion''("짐승")에서 유래하며, 법랑질의 층상 구조를 가리킨다. 종명 ''sibericus''는 다시코바 공주의 소장품이 주로 시베리아에서 온 것에 대한 언급일 가능성이 높다. 하지만 표본의 정확한 기원은 알려지지 않았다. 1877년, 독일의 자연주의자 요한 프리드리히 폰 브란트(Johann Friedrich von Brandt)는 이를 새롭게 설립된 아과 엘라스모테리이네아(Elasmotheriinae)에 배치했는데, 현생 코뿔소와는 별개였다.[20]
3. 분류
이 속은 수백 개의 발견지에서 알려져 있으며, 주로 두개골 파편과 치아가 발견되었지만, 경우에 따라서는 유라시아 동유럽에서 중국에 이르는 지역에 흩어져 있는 후두골의 거의 완전한 골격도 발견되었다.[5] 종으로의 분류는 주로 치아와 턱의 미세한 차이점과 두개골의 모양에 기초한다.[6]
3. 1. 종
엘라스모테리움아과는 현존하는 모든 코뿔소를 포함하는 코뿔소아과와 구분되며, 이 둘의 분기 시점은 논란이 있지만 대략 올리고세인 약 3500만 년 전으로 추정된다.[7] 엘라스모테리움은 마이오세 이후 엘라스모테리움아과에서 알려진 유일한 종이다.[8]
엘라스모테리움 속(genus)은 시노테리움(''Sinotherium'')에서 진화했을 가능성이 높으며,[9] 가장 오래된 종은 ''Elasmotherium primigenium''으로 중국 산시성 딩볜현의 후기 마이오세 지층에서 발견되었다.[9] 엘라스모테리움은 플라이스토세 초기에 동유럽에 도착했다.[10]
엘라스모테리움아과는 높은 치관과 치아 법랑질이 잇몸 아래로 뻗어 있는 고치성 치열 패턴을 가지는데,[11] 이는 강가 하안습지의 거친 사료에 대한 적응으로 해석된다.[12]
| 종 | 명명 | 발견 지역 및 시기 |
|---|---|---|
| *E. chaprovicum* | Shvyreva, 2004 | 북부 코카서스, 몰도바, 아시아 (260만 년 전~220만 년 전, 후기 플라이오세~초기 플라이스토세) [14] |
| *E. peii* | Chow, 1958 | 중국 산시성, 러시아 (220만 년 전~160만 년 전) [15][2] |
| *E. caucasicum* | 알렉세이 보리샤크(Aleksei Borissiak), 1914 | 흑해 지역, 중국 북부 니허완(Nihewan) (110만 년 전~80만 년 전, 초기 플라이스토세) [17][18][12] |
| *E. sibiricum* | 요한 피셔 폰 발트하임(Johann Fischer von Waldheim), 1808 | 러시아 남서부, 서시베리아, 우크라이나, 몰도바 (중기 플라이스토세) [19] |
4. 특징
엘라스모테리움은 일반적으로 매머드나 털코뿔소와 같은 당대 거대동물처럼 털이 많은 동물로 묘사되지만, 때로는 현대 코뿔소처럼 털이 없는 모습으로 그려지기도 한다. 1948년 러시아 고생물학자 발렌틴 테랴예프는 엘라스모테리움이 돔 모양 뿔을 가진 반수생 동물이며, 다른 코뿔소와 달리 습지 맥처럼 네 발가락을 가졌고 하마와 유사하다고 제안했다. 그러나 엘라스모테리움은 세 개의 발가락만으로 기능을 수행했다는 사실이 밝혀지면서[41] 테랴예프의 주장은 과학계에서 큰 주목을 받지 못했다.[42]

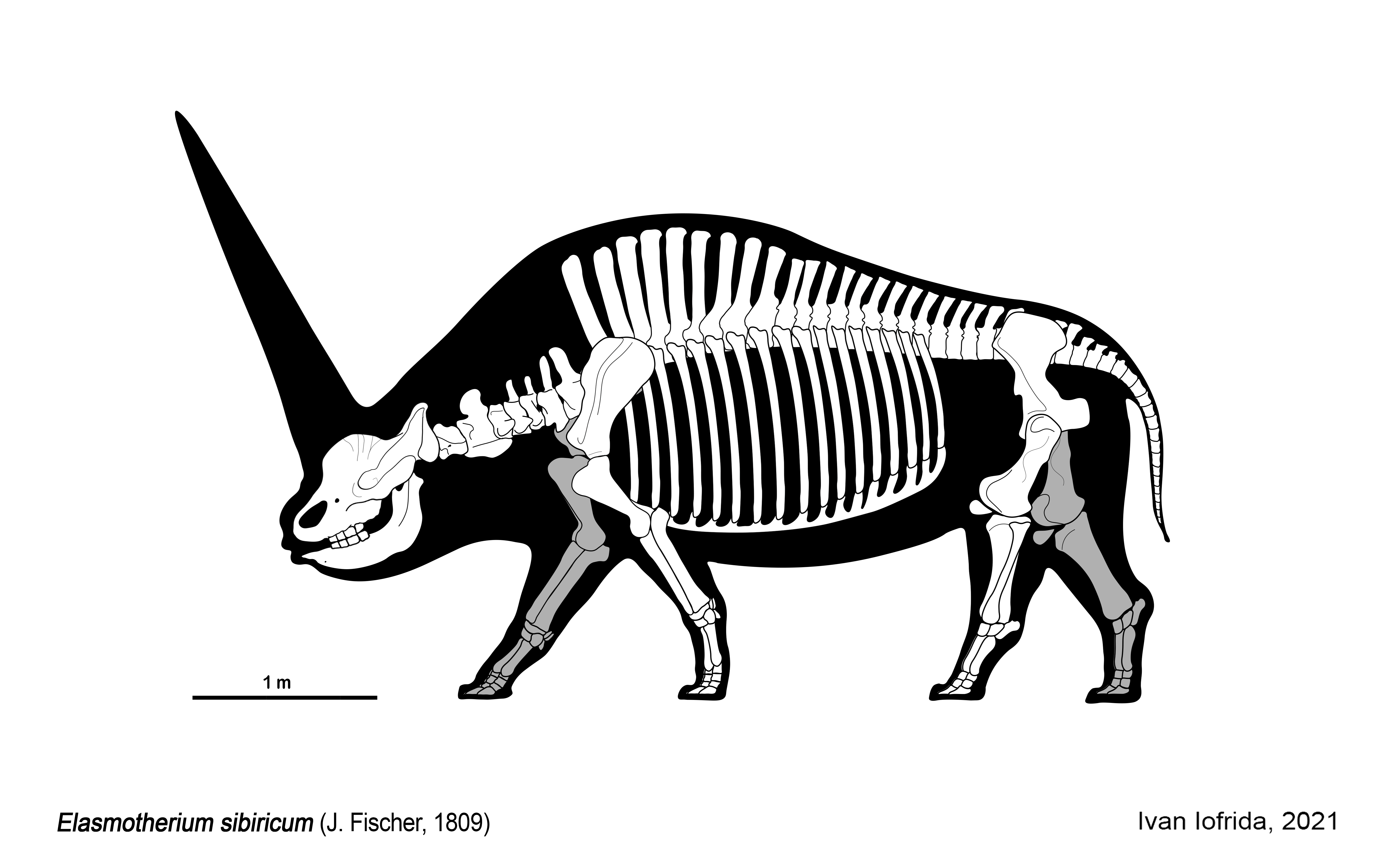
''E. sibiricum'' 표본은 최대 길이 4.5m, 어깨 높이 2.5m에 달했으며, ''E. caucasicum''은 최소 길이 5m, 추정 무게 3.5ton~5ton에 달했다.[20][21] 이는 엘라스모테리움을 제4기의 가장 큰 코뿔소로 만들었으며,[38] 두 종 모두 털매머드와 크기가 비슷하고 당시 털코뿔소보다는 컸다.[22][8] 발은 굽있는 형태였고 앞발이 뒷발보다 컸으며, 앞뒤 모두 세 개의 발가락과 흔적 기관인 다섯 번째 중수골이 있었다.[23]
엘라스모테리움은 튼튼한 몸과 긴 다리를 가져 질주에 적합했으며, 초원 생활에 적응한 것으로 보인다.
4. 1. 뿔
엘라스모테리움은 전통적으로 이마에 둥근 돔 모양의 케라틴성 뿔을 가지고 있었다고 여겨진다. 뿔은 깊이 13cm, 주름진 표면, 둘레 90cm 정도였다. 이 주름은 뿔을 생성하는 조직을 위한 혈관 자리로 해석된다.[43][52]코뿔소의 뿔은 뼈에 붙어 있지 않고 밀도가 높은 피부 조직 표면에서 자라 뼈의 불규칙성과 융기를 만들어 스스로 고정된다.[44] 가장 바깥쪽 층은 각질화된다.[45] 층이 노화됨에 따라 자외선으로 인한 각질 분해, 건조, 지속적인 마모로 인해 뿔의 직경이 감소한다.[46] 그러나 중앙의 멜라닌과 칼슘 침전물은 각질을 단단하게 만들어 뿔의 독특한 모양을 만든다.[47]
등에는 일반적으로 무거운 뿔을 지탱하는 것으로 여겨지는 큰 근육 혹이 있었을 가능성이 높다.[48]
2021년 연구에 따르면, 두개골 돔은 매우 약하고 큰 뿔에는 적합하지 않았으며, 오히려 작은 뿔을 나타내는 것으로 보인다. 이 돔은 '''루싱고릭스'''와 하드로사우루스의 볏과 유사하게 일종의 공명실로 기능했을 수 있다.[30]
가장 큰 종인 *E. caucasicum*는 전체 길이 5m, 전체 높이 2.2m, 체중은 3.6ton에서 4.5ton에 달했던 것으로 추정된다. 두개골에는 안와(眼窩) 상부, 전두골(前頭骨) 끝(이마)에 받침대 모양의 큰 돌출부가 있다. 이 상부는 거칠거칠한 조면(粗面)으로 되어 있으며, 여기에 길이 2m에 달하는 것으로 추정되는 체모(體毛)가 굳어진 뿔이 있었던 것으로 보인다. 다른 코뿔소의 경우 뿔은 비골(鼻骨) 위에 있으며, 이는 가장 큰 차이점이다.
4. 2. 치아
다른 코뿔소들과 마찬가지로 엘라스모테리움(*Elasmotherium*)은 씹는 데 사용되는 두 개의 소구치와 세 개의 어금니를 가지고 있었고, 절치(앞니)와 견치(송곳니)는 없었다. 대신 잡는 입술을 이용하여 먹이를 뜯어 먹었다.[41] 엘라스모테리움은 큰 치관과 잇몸선 아래까지 뻗어 있는 법랑질을 가졌고, 치아가 계속 자라는 진정한 고치였다. 엘라스모테리움 화석에서는 치근의 흔적이 거의 발견되지 않는다.
절치(切齒)는 없어지고, 입술로 식물을 뜯어 먹었다. 구치(臼齒)의 치관(齒冠)은 높아지고, 에나멜질의 능선(畝)이 복잡한 무늬를 형성하고 있었다. 아마도 단단한 벼과 식물을 먹기 위한 적응이었을 것이다.
4. 3. 골격 및 이동
현대의 긴 이빨을 가진 발굽 포유류는 일반적으로 개방된 환경에서 풀을 뜯어 먹는 동물이며[49], 긴 이빨은 질기고 섬유질인 풀을 씹는 데 적응한 것일 수 있다.[50] 엘라스모테리움의 이빨 마모는 풀을 뜯는 흰코뿔소와 비슷하며[51], 두 동물 모두 머리가 아래쪽을 향하고 있어 생활 방식이 비슷하고 낮은 곳의 식물에만 도달할 수 있었다. 실제로 엘라스모테리움의 머리는 모든 코뿔소 중 가장 둔각을 가지고 있었으며, 가장 낮은 수준에 도달할 수 있었기 때문에 습관적으로 풀을 뜯어 먹었을 것이다.[52] 엘라스모테리움은 또한 설치류에서 흔히 볼 수 있는 자라는 이빨을 가지고 있었으며, 치아 생리학은 습하고[53] 입자가 많은 토양에서 먹이를 끌어올리는 데 영향을 받았을 수 있다. 따라서 이들은 현대의 매머드와 유사하게 매머드 스텝과 강가에 서식했을 수 있다.[42]엘라스모테리움은 최고 속도가 40kph에서 45kph인, 30kph로 달리는 흰코뿔소와 비슷한 달리기 사지를 가지고 있었다. 그러나 엘라스모테리움은 흰코뿔소보다 두 배인 약 5MT의 무게를 가지고 있어 보행과 이동이 더 제한되어 훨씬 느린 속도를 달성했을 가능성이 높다. 2.5MT에서 11MT의 무게를 가진 코끼리는 20kph를 넘는 속도로 걸을 수 없다.[54] 발은 굽있는 형태였고, 앞발이 뒷발보다 컸으며, 앞뒤 모두 세 개의 발가락이 있었고, 흔적 기관인 다섯 번째 중수골이 있었다.[23]
5. 고생물학
엘라스모테리움은 현존하는 모든 코뿔소를 포함하는 코뿔소아과(Rhinocerotinae)와 구분되는 엘라스모테리움아과(Elasmotheriinae)에 속한다. 엘라스모테리움아과와 코뿔소아과의 분기 시점은 논란이 있는데, 기존 연구는 에오세(Eocene)인 약 4700만 년 전으로 추정했지만,[38] 최근 연구는 올리고세(Oligocene)인 약 3500만 년 전으로 추정한다.[7]
엘라스모테리움은 마이오세(Miocene) 이후 엘라스모테리움아과에서 알려진 유일한 종으로, 다른 종들은 사바나의 확장과 함께 멸종되었다.[8] 가장 오래된 엘라스모테리움 종인 ''Elasmotherium primigenium''은 중국 산시성(Shaanxi) 딩볜현(Dingbian County)의 후기 마이오세 지층에서 발견되었다. 엘라스모테리움은 중국에서도 발견되는 엘라스모테리움 속(genus)인 시노테리움( ''Sinotherium'')으로부터 진화했을 가능성이 높다.[9] 엘라스모테리움은 플라이스토세(Pleistocene) 초기인 약 250만 년 전에 동유럽에 도착했다.[10]
높은 치관(tooth crown)과 치아 법랑질(tooth enamel)이 잇몸 아래로 뻗어 있는 고치성(Hypsodonty) 치열 패턴은 엘라스모테리움아과의 특징으로 여겨지며,[11] 아마도 강가의 하안습지(riparian zone)에 있는 더 거친 사료에 대한 적응으로 해석된다.[12]
- E. primigenium*을 제외하고는 네 종의 연대종 *엘라스모테리움*이 있다. 가장 오래된 종부터 가장 최근 종까지 순서대로 *E. chaprovicum*, *E. peii*, *E. caucasicum*, 그리고 *E. sibiricum*이며, 이들은 후기 플라이오세부터 후기 플라이스토세까지 걸쳐 서식했다.[2]

- E. chaprovicum*은 원래 *E. caucasicum*으로 여겨졌으나,[13] 치아를 기반으로 새로운 종으로 재정의되었으며, 카프로브 동물군 복합체의 이름을 따서 명명되었다.[6] 카프로브 동물군 복합체는 중기 빌라프랑키안(MN17)에 속하며, 후기 플라이오세의 피아첸시안(Piacenzian)과 초기 플라이스토세의 젤라시안(Gelasian)에 걸쳐 북캅카스, 몰도바 및 아시아에 분포했으며, 260만 년 전~220만 년 전으로 추정된다.[14]
- E. peii*는 (Chow, 1958)에 의해 처음으로 중국 산시성에서 발견된 유해를 바탕으로 기술되었다.[15] 2018년에 산시성에서 추가 유해가 발견되어 기술되었다.[16] 이 종은 *엘라스모테리움*의 고전적인 서식 범위에서도 많은 유해가 발견되었으며, 일부 자료에서는 이 종을 *E. caucasicum*의 동의어로 간주했지만, 현재는 별개의 종으로 간주된다.[2] 220만 년 전~160만 년 전의 프세쿠프스(Psekups) 동물군 복합체에서 발견된다.[2]
- E. caucasicum*은 러시아 고생물학자 알렉세이 보리샤크(Aleksei Borissiak)에 의해 1914년에 처음으로 기술되었는데, 그는 이 종이 흑해 지역에서 초기 플라이스토세 타마니안(Tamanian) 동물군 단위(110만 년 전~80만 년 전, 타만 반도)의 구성원으로 번성했던 것으로 보인다고 말했다. 이는 이 동물군에서 가장 흔하게 발견되는 포유류이다. *E. caucasicum*은 *E. sibiricum*보다 더 원시적인 것으로 여겨지며, 아마도 조상 종을 나타내는 것으로 생각된다.[17][18] 또한 중국 북부의 초기 플라이스토세 니허완(Nihewan) 동물군 집합체에서도 알려져 있으며, 약 160만 년 전에 멸종한 것으로 추정된다. 이는 *엘라스모테리움*이 러시아와 중국에서 별도로 진화했음을 시사한다.[12]
요한 피셔 폰 발트하임(Johann Fischer von Waldheim)에 의해 1808년에 기술된 *E. sibiricum*은 연대순으로 가장 최근의 종이며, 중기 플라이스토세에 나타나 러시아 남서부에서 서시베리아까지, 그리고 남쪽으로 우크라이나와 몰도바까지 분포했다.[19]

현대의 고치(高齒) 발굽 포유류는 일반적으로 열린 환경에서 초식(草食)을 하는데,[31] 고치는 질기고 섬유질이 많은 풀을 씹는 데 대한 적응일 수 있다.[32] ''엘라스모테리움''의 치아 마모는 초식성인 흰코뿔소와 유사하며,[33] 두 종 모두 머리가 아래쪽으로 향해 있어 유사한 생활 방식과 낮은 위치의 식물만 먹을 수 있는 능력을 나타낸다. 사실, ''엘라스모테리움''의 머리는 다른 어떤 코뿔소보다 각도가 가장 둔하여 가장 낮은 수준의 식물만 닿을 수 있었고, 따라서 습관적으로 초식을 했을 것이다.[34] ''엘라스모테리움''은 또한 계속 자라는 치아인 진고지(眞高齒)를 보이는데,[35] 이는 일반적으로 설치류에서 볼 수 있으며, 치아 생리학은 습하고 알갱이가 있는 토양에서 음식을 뽑아 올리는 것에 영향을 받았을 수 있다. 따라서, 현대의 매머드와 유사하게 맘모스 스텝과 하천변 모두에 서식했을 가능성이 있다.[12]
6. 멸종
엘라스모테리움 속(屬)에는 *E. primigenium*을 제외하고 네 종의 연대종이 있었다. 연대순으로 가장 오래된 종부터 나열하면, *E. chaprovicum*, *E. peii*, *E. caucasicum*, *E. sibiricum*이며, 이들은 후기 플라이오세부터 후기 플라이스토세까지 서식했다.[2]
| 종 | 명명 시기 및 학자 | 서식 시기 (추정) | 발견 지역 및 특징 |
|---|---|---|---|
| *E. chaprovicum* | 2004년, Shvyreva | 260만 년 전 ~ 220만 년 전 | 북부 코카서스, 몰도바, 아시아, 카프로브 동물군 복합체(Khaprov Faunal Complex) |
| *E. peii* | 1958년, Chow | 220만 년 전 ~ 160만 년 전 | 중국 산시성, 프세쿠프스(Psekups) 동물군 복합체 |
| *E. caucasicum* | 1914년, 알렉세이 보리샤크(Aleksei Borissiak) | 110만 년 전 ~ 80만 년 전 (멸종: 약 160만 년 전) | 타마니 반도 (타마니안(Tamanian) 동물군), 중국 북부 니허완(Nihewan) 동물군, *E. sibiricum*보다 원시적인 조상 종 |
| *E. sibiricum* | 1808년, 요한 피셔 폰 발트하임(Johann Fischer von Waldheim) | 중기 플라이스토세 ~ 3만 6천 년 ~ 3만 5천 년 전 | 러시아 남서부, 서시베리아, 우크라이나, 몰도바, 카자흐스탄 파블로다르주 |
엘라스모테리움은 이전에는 약 20만 년 전에 멸종된 것으로 알려졌으나, 카자흐스탄 파블로다르 주에서 발견된 *엘라스모테리움 시비리쿰*(E. sibiricum) 두개골은 약 3만 6천 년~3만 5천 년 전 서시베리아 평원에서도 서식했음을 보여준다.[38] 시베리아 스멜로프스카야 동굴과 바트팍 동굴에서는 5만 년 전 유해가 발견되기도 했다.[37]
이 시기는 플라이스토세 멸종 시기와 거의 일치하며, 기후 변화와 현생 인류의 이동이 겹치는 시기이다.[38]
7. 대중 문화
- 프리히스토릭 파크: 2화에 등장한다. 100만 년 전 시베리아에서 수컷 한 마리가 보호되어, 파크에서 사육된다.
- 극장판 가면라이더 555 파라다이스 로스트: 이 종을 모티브로 한 엘라스모테리움 올페녹이 등장한다.
- ZOIDS: 이 종을 모티브로 한 사이클롭스 일식이 등장한다.
참조
[1]
웹사이트
Elasmotherium
https://paleobiodb.o[...]
National Center for Ecological Analysis and Synthesis
2000-03-23
[2]
논문
On the importance of the representatives of the genus Elasmotherium (Rhinocerotidae, Mammalia) in the biochronology of the Pleistocene of Eastern Europe
2015-08-01
[3]
논문
Evolution and extinction of the giant rhinoceros Elasmotherium sibiricum sheds light on late Quaternary megafaunal extinctions
https://www.nature.c[...]
2019-01-01
[4]
논문
Sur L'Elasmotherium et le Trogontothérium
https://books.google[...]
Imprimerie de l'Université Impériale
1809-01-01
[5]
서적
Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia: Abstracts of the International INQUA-SEQS Conference (Rostov-on-Don, June 21–26, 2010)
https://web.archive.[...]
Russian Academy of Science
2010-03-26
[6]
서적
Late Pliocene large mammals from Northeastern Sea of Azov Region
http://paleorostov.n[...]
SSC RAS Publishing
2008-01-01
[7]
논문
Ancient and modern genomes unravel the evolutionary history of the rhinoceros family
2021-08-01
[8]
논문
Diversity and evolutionary trends of the Family Rhinocerotidae (Perissodactyla)
http://www.rhinoreso[...]
1998-01-01
[9]
논문
The most primitive Elasmotherium (Perissodactyla, Rhinocerotidae) from the Late Miocene of northern China
https://www.tandfonl[...]
2021-04-26
[10]
논문
On the importance of the representatives of the genus Elasmotherium (Rhinocerotidae, Mammalia) in the biochronology of the Pleistocene of Eastern Europe
https://linkinghub.e[...]
2015-08-27
[11]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2003-01-01
[12]
서적
The World of Elephants
http://www.sovrainte[...]
Consiglio Nazionale delle Ricerche
2001-01-01
[13]
웹사이트
The Development of the Late Pleistocene to Early Middle Pleistocene Large Mammal Fauna of Ukraine
http://www.senckenbe[...]
Senckenberg Gesellschaft für Naturforschung (SGN)
2024-05-01
[14]
웹사이트
Results of the Khapry Faunal Unit revision
http://www.senckenbe[...]
Senckenberg Gesellschaft für Naturforschung (SGN)
2024-05-01
[15]
논문
New elasmotherine Rhinoceroses from Shansi
1958-01-01
[16]
논문
New postcranial bones of elasmotherium peii from shanshenmiaozui in Nihewan basin, Northern China
http://journals.open[...]
2018-09-01
[17]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2005-01-01
[18]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2005-01-01
[19]
서적
Pleistocene Large Mammal Associations of the Sea of Azov and Adjacent Regions
https://web.archive.[...]
Russian Academy of Science
2010-03-26
[20]
논문
On the fossil rhinoceros ''Elasmotherium'' (including the collections of the Russian Academy of Sciences)
http://www.rhinoreso[...]
2005-01-01
[21]
서적
Эласмотерии плейстоцена Евразии
https://www.geokniga[...]
2016-01-01
[22]
논문
Changes in Western European Rhinocerotidae related to climatic variations
http://www.rhinoreso[...]
1995-01-01
[23]
논문
About the Hyroideum, Sternum and Metacarpale V Bones of ''Elasmotherium sibiricum'' Fischer (Rhinocerotidae)
http://palaeontologi[...]
1977-01-01
[24]
논문
The Elasmotherium
1878-08-08
[25]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2009-01-01
[26]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2009-01-01
[27]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2009-01-01
[28]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2009-01-01
[29]
문서
(저자와 출판년도 정보가 부족하여 제목만 표시)
2009-01-01
[30]
논문
The experience in reconstructing of the head of Elasmotherium (Rhinocerotidae)
https://zmmu.msu.ru/[...]
2021-11-16
[31]
논문
Hypsodonty in ungulates: an adaptation for grass consumption or for foraging in open habitat?
2008-02-01
[32]
서적
Origin and evolution of the grazing guild in Cenozoic New World terrestrial mammals
https://archive.org/[...]
Cambridge University Press
2000-01-01
[33]
논문
Causal Factors Shortening Tooth Series with Age
1938-01-01
[34]
서적
The rhinoceroses from Neumark-Nord and their nutrition
http://www.rhinoreso[...]
Halle/Saale
2010-01-01
[35]
논문
Hypsodonty and enamel microstructure in the Paleocene gondwanatherian mammal ''Sudamerica amerghinoi''
http://www.app.pan.p[...]
1999-01-01
[36]
저널
Limb Designs, Function and Running Performance in Ostrich-mimics and Tyrannosaurs
http://www.mnhn.ul.p[...]
1998-12-00
[37]
서적
Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia: Abstracts of the International INQUA-SEQS Conference (Rostov-on-Don, June 21–26, 2010)
http://seqs2010-rost[...]
Russian Academy of Science
2011-03-26
[38]
저널
Evolution and extinction of the giant rhinoceros ''Elasmotherium sibiricum'' sheds light on late Quaternary megafaunal extinctions
https://ora.ox.ac.uk[...]
[39]
웹사이트
Elasmotherium
https://paleobiodb.o[...]
National Center for Ecological Analysis and Synthesis
2011-03-23
[40]
저널
Evolution and extinction of the giant rhinoceros ''Elasmotherium sibiricum'' sheds light on late Quaternary megafaunal extinctions
https://ora.ox.ac.uk[...]
[41]
저널
On the fossil rhinoceros ''Elasmotherium'' (including the collections of the Russian Academy of Sciences)
http://www.rhinoreso[...]
[42]
서적
The World of Elephants
http://www.sovrainte[...]
Consiglio Nazionale delle Ricerche
[43]
저널
The Elasmotherium
1878-08-08
[44]
harvnb
[45]
harvnb
[46]
harvnb
[47]
harvnb
[48]
harvnb
[49]
저널
Hypsodonty in ungulates: an adaptation for grass consumption or for foraging in open habitat?
2008-02-00
[50]
서적
Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record
https://archive.org/[...]
Cambridge University Press
[51]
저널
Causal Factors Shortening Tooth Series with Age
[52]
서적
Elefantenreich – Eine Fossilwelt in Europa
http://www.rhinoreso[...]
Halle/Saale
[53]
저널
Hypsodonty and enamel microstructure in the Paleocene gondwanatherian mammal ''Sudamerica amerghinoi''
http://www.app.pan.p[...]
[54]
저널
Limb Designs, Function and Running Performance in Ostrich-mimics and Tyrannosaurs
http://www.mnhn.ul.p[...]
1998-12-00
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com