체관부
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
체관부는 식물의 유기 양분 수송을 담당하는 조직으로, 체관 요소, 체관부 섬유, 체관부 유세포 등으로 구성된다. 체관 요소는 당을 수송하는 세포이며, 사관 요소와 사세포로 나뉜다. 체관부 섬유는 기계적 지지를 담당하며, 체관부 유세포는 양분 저장 등에 관여한다. 체관부는 일차 체관부와 이차 체관부로 구분되며, 일차 체관부는 정단 분열 조직에서, 이차 체관부는 관다발 형성층에서 유래한다. 체관부는 광합성 산물의 수송 통로이며, 압력 흐름설에 따라 원천에서 저장소로 물질이 이동한다. 한국에서는 소나무 속껍질이 구황식물로 사용되었으며, 닥나무 등의 인피섬유는 한지 등의 원료로 사용된다. 임업 및 농업에서는 환상박피를 통해 식물의 생장을 조절하거나, 영양번식에 활용하기도 한다.
더 읽어볼만한 페이지
- 조직 (생물학) - 상피 조직
상피 조직은 세포 간 결합, 극성, 다양한 세포 형태를 특징으로 하며, 신체 내부 표면을 구성하고, 세포 층의 수와 기능에 따라 분류되며, 보호, 흡수, 분비, 수송 등의 역할을 수행하며, 외배엽, 중배엽, 내배엽에서 유래하고, 암종과 관련된다. - 조직 (생물학) - 근육
근육은 수축과 이완을 통해 몸의 움직임을 담당하는 조직으로, 민무늬근, 가로무늬근, 심근의 세 가지 유형으로 나뉘며, 골격근, 심장근, 평활근으로도 분류되고, 근섬유 다발로 구성되어 액틴과 미오신의 상호작용으로 수축이 일어나며, 발생은 배아의 축방 중배엽에서 시작되고 성별에 따른 차이와 노화에 따른 퇴화 현상을 보인다. - 식물해부학 - 꽃자루
- 식물해부학 - 표피 (식물학)
표피는 식물의 잎, 줄기, 뿌리 등을 덮는 투명한 세포층으로 수분 손실을 막고 보호 기능을 수행하며, 기공을 통해 가스 교환을 조절하고, 동물의 경우 상피 조직으로 구성되어 피부를 형성한다. - 식물생리학 - 광합성
광합성은 생물이 빛에너지를 이용하여 이산화탄소와 물로부터 유기물을 합성하고 산소를 방출하는 과정으로, 엽록체 내 틸라코이드 막에서 일어나는 명반응과 스트로마에서 일어나는 암반응으로 구성되며, 환경에 따라 탄소 농축 메커니즘을 통해 효율을 높이기도 하고, 지구 대기의 산소를 생성하는 주요 원천이다. - 식물생리학 - 저분자
저분자는 분자량이 900달톤 이하인 작은 분자로, 세포막 투과가 용이하여 의약품으로 많이 사용되지만, 표적 적용에 제한이 있고 내성 문제가 발생할 수 있으며, 의약품 외에도 이차 대사산물, 연구 도구, 항게놈 치료제 등으로 활용된다.
| 체관부 | |
|---|---|
| 개요 | |
| 명칭 | 체관부 |
| 로마자 표기 | phloem |
| 그리스어 어원 | φλοιός(φλοιός, 껍질) |
| 설명 | 관다발식물에서 광합성 산물을 운반하는 조직 |
| 구조 및 기능 | |
| 주요 기능 | 광합성 산물 (유기물) 운반 및 식물체 지지 |
| 구성 요소 | 사관 반세포 사부 섬유 유세포 |
| 유기물의 이동 | 잎에서 생성된 광합성 산물(주로 설탕)을 식물의 다른 부분으로 운반 |
| 이동 방향 | 원천 세포(잎)에서 수용 세포(뿌리, 열매, 생장점 등)로 양방향 수송 중력 방향에 영향을 받지 않음 |
| 기타 기능 | 양분 저장 식물체 지지 |
| 발달 | |
| 1차 사부 | 식물 정단 분열 조직으로부터 발달 원사부와 후생사부로 나뉨 |
| 2차 사부 | 관다발 형성층으로부터 발달 목본 식물에서 줄기나 뿌리의 두께 생장에 기여 |
| 추가 정보 | |
| 최초 발견 | 카를 나겔리(Carl Nägeli)에 의해 1858년 발견 |
| 참고 자료 | Annu Rev Plant Biol (2004), "Transport mechanisms for organic forms of carbon and nitrogen between source and sink" Ontogeny, Cell Differentiation, and Structure of Vascular Plants (1989), "Phloem" Development (2021), "Metaphloem development in the Arabidopsis root tip" Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body (2006), "Phloem" The Arabidopsis Book (2015), "Vascular Cambium Development" Merriam-Webster.com Dictionary, "Phloem" |
| 이미지 및 기타 | |
 | |
| |
| |
2. 구성
체관부는 체관 요소, 체관부 섬유, 체관부 유세포 등으로 구성된다.[26]
- 체관 요소: 광합성으로 만들어진 유기물이 이동하는 통로 역할을 하는 살아있는 세포이다. 세포핵이나 액포 등이 없어 물질 이동이 쉽다. 속씨식물의 경우 사관 요소를 가지며, 사관 요소는 세로로 연결되어 사관을 형성한다. 사관 요소 사이의 격벽은 사판이라고 불리며, 대형 사공이 모여 체와 같은 사역을 형성한다.
- 체관부 섬유: 세포벽이 발달한 가늘고 긴 세포로, 보통 원형질이 없는 죽은 세포이다. 목부 섬유 등과 함께 식물체를 지지하는 역할을 한다. 목부 섬유에 비해 일반적으로 길며, 보통 벽공이 불명료하다.
- 체관부 유세포: 유기 양분을 저장하는 역할을 하는 유세포이다. 수지(樹脂)나 타닌 등을 저장하거나 분비 조직을 가진 경우도 있다.
체관부 조직은 체관 요소, 특수한 동반세포 또는 알부민 세포와 비특수화된 세포들을 포함한 유조직 세포들, 그리고 섬유와 후각세포와 같은 지지 세포들로 구성된다.
2. 1. 체관 요소
체관 요소(sieve element)는 광합성 산물 등 유기물의 통로 역할을 하는 살아있는 세포이다.[1] 세포핵이나 액포 등의 세포소기관이 퇴화되어 있어 물질 이동이 용이하다.[1][2] 세포 간 물질 수송은 원형질 연락이 확대된 사공(篩孔, sieve pore)을 통해 이루어진다.[1] 사공에서 원형질 연락 주위에는 칼로스(callose)가 침착되어 있다.[3]속씨식물은 사관 요소를 가지며, 사관 요소는 세로로 연결되어 사관(篩管, sieve tube)을 형성한다.[1] 사관 요소 사이의 격벽은 사판(篩板, sieve plate)이라고 불리며, 대형 사공이 모여 체와 같은 사역을 형성한다.[1] 사판에서 사역의 배치 양식은 다양하며, 복합 사판(compound sieve plate)과 단사판(simple sieve plate)으로 구분된다.[4] 사관에서의 물질 수송은 주로 사판을 통해 이루어지지만, 측면에도 사역이 존재한다(측사역).[5] 사관 요소에는 반세포(伴細胞, companion cell)가 부수하며, 반세포는 원형질 연락으로 사관 요소와 연결되어 당 등의 물질 이동 및 대사 조절에 관여한다.[1][2]
속씨식물 이외의 관다발 식물(양치식물, 겉씨식물)은 사세포(篩細胞, sieve cell)를 가진다.[4] 사세포는 사판을 갖지 않고, 세포 측면에 산재하는 사역을 통해 물질 수송이 이루어진다.[3] 겉씨식물에는 단백 세포(albuminous cell)가 사세포에 부수하며, 반세포와 유사한 기능을 수행한다.[1]
2. 2. 체관부 섬유
체관부 섬유(사부 섬유, phloem fiber)는 발달된 세포벽을 가진 가늘고 긴 세포이며, 보통 원형질이 없는 죽은 세포이다.[26] 체관부 섬유는 모여서 체관부 섬유 조직(phloem fiber tissue)을 형성한다.[26] 목부 섬유 등과 함께 식물체의 기계적 지지에 기여한다. 목부 섬유에 비해 일반적으로 길며, 보통 벽공이 불명료하다.[26] 길이는 삼(Cannabis sativa)(삼과)에서 1 cm, 아마(Linum usitatissimum)(아마과)에서 2–4 cm, 닥나무(Boehmeria nivea)(쐐기풀과)에서는 22 cm에 달한다. 유관속 주변에 후벽 세포가 발달하는 종에서는 체관부 섬유가 발달하지 않는다(예: 호박). 속씨식물 이외에서는 보통 체관부 섬유가 보이지 않는다.[26]섬유는 길고 가는 지지 세포로서 유연성을 제한하지 않고 장력 강도를 제공한다.[10]
체관부 섬유는 그 바깥쪽에 형성되는 피층 섬유와 함께 '''인피섬유'''(bast fiber)라고 불리며, 천연섬유로서 다양하게 활용된다. 아마(린넨, 아마과), 삼(삼, 삼과), 닥나무(저마, 쐐기풀과), 황마(주트, 아욱과) 등의 인피섬유는 베나 밧줄의 원료로 쓰이며(그림 8a), 닥나무(뽕나무과)나 꾸지뽕나무, 가마(팥꽃나무과)의 인피섬유는 한지나 지폐의 원료로 사용된다.[33][34] (그림 8b).

2. 3. 체관부 유세포
체관부에 존재하는 유세포는 '''체관부유세포'''(사부유세포, phloem parenchymatous cell)라고 불리며, 모여서 '''체관부유조직'''(사부유조직, phloem parenchyma)을 형성한다.[26] 유기 양분의 저장 등에 관여한다.[26] 수지(樹脂)나 타닌 등을 저장하거나 분비 조직을 가진 경우도 있다.[26][27] 일차 체관부에서는 체관부 유세포가 체관요소에 부수적으로 존재한다.이와사|쿠라타니|사이토|츠카야|2013|p=605|loc=「체관부」일본어 이차 체관부의 경우에는 이차 물관부의 유조직과 마찬가지로, 세로로 배열된 세포로 이루어진 축방향 유조직(axial parenchyma)과 방사 방향으로 배열된 세포로 이루어진 방사유조직(ray parenchyma)이 있다.[26]3. 체관부의 종류
체관부는 형성 위치에 따라 일차 체관부와 이차 체관부로 나뉜다.[26] 일차 체관부는 정단 분열 조직에서 유래하며 모든 관다발 식물에 존재한다. 반면 이차 체관부는 관다발 형성층에서 생성되며, 목본식물과 같이 이차 생장을 하는 식물에서 수피 안쪽에 위치한다.
3. 1. 일차 체관부
정단 분열 조직(분열조직)에서 유래하는 전형성층에서 형성되는 체관부를 '''일차 체관부'''라고 한다. 모든 관다발 식물은 일차 체관부를 가지지만, 이차 생장을 하는 목본식물에서는 이차 체관부의 발달과 함께 일차 체관부는 파괴된다.[19]일차 체관부는 보통 일차 목부의 사이(그림 5a)나 바깥쪽(그림 5b)에 위치하지만, 체관부가 목부를 둘러싸고 있는 경우도 있다.(그림 5c)[19]
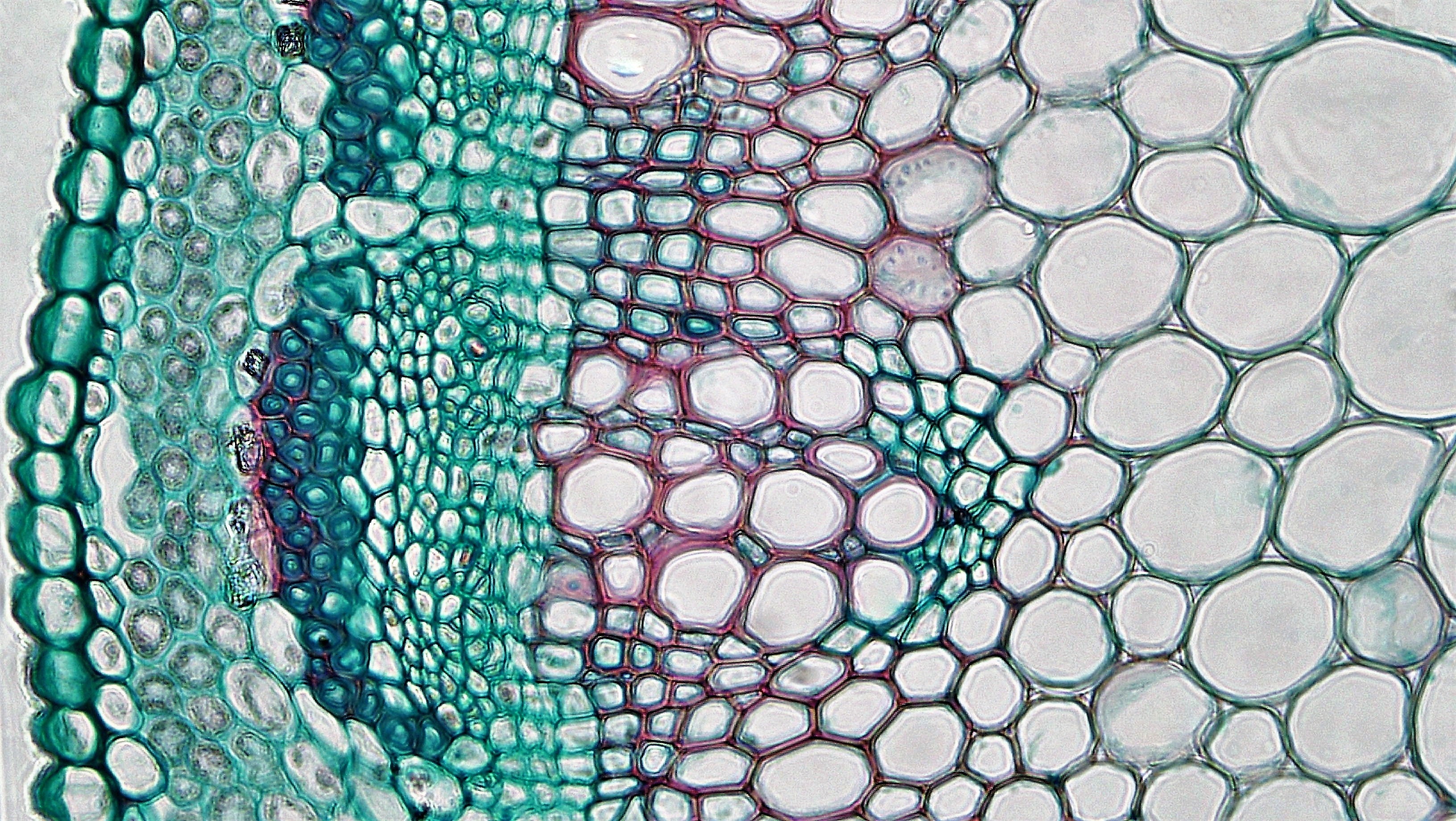
일차 체관부 중에서 가장 먼저 분화하는 부분을 '''원시 체관부''', 그 후에 분화하는 부분을 '''후생 체관부'''라고 한다.[26] 원시 체관부·후생 체관부의 형태적 차이는 원시 목부·후생 목부만큼 명료하지 않지만, 일반적으로 후생 체관부가 더 굵고 길다.[19] 또 원시 체관부의 사관에는 반세포가 없는 경우가 많다.[19]
식물이 배(胚)일 때, 관다발 조직은 배 중앙에 있는 원형층(procambium) 조직에서 발생한다. 원시 체관부 자체는 속씨식물에서 처음으로 잎이 나타나는 곳인 자엽절(cotyledonary node)까지 확장되는 중앙맥(mid-vein)에 나타나며, 연속적인 가닥을 형성한다. 단백질 PIN1에 의해 수송되는 호르몬 옥신(auxin)은 이러한 원시 체관부 가닥의 성장을 담당하며, 이 조직의 최종적 동일성을 알리는 신호를 보낸다. SHORTROOT(SHR)과 166도 그 과정에 참여하는 반면, 캘로스 합성효소 3(Callose Synthase 3)은 SHR과 마이크로RNA165가 갈 수 있는 위치를 억제한다. 또한 체관부 분화 과정에서 NAC45/86 유전자의 발현은 식물에서 특정 세포의 핵을 제거하여 사부 요소(sieve element)를 생성하는 기능을 한다.[19]
배에서 뿌리 체관부는 배아 뿌리와 자엽 사이에 있는 상위하배축(upper hypocotyl)에서 독립적으로 발달한다.[20]
3. 2. 이차 체관부
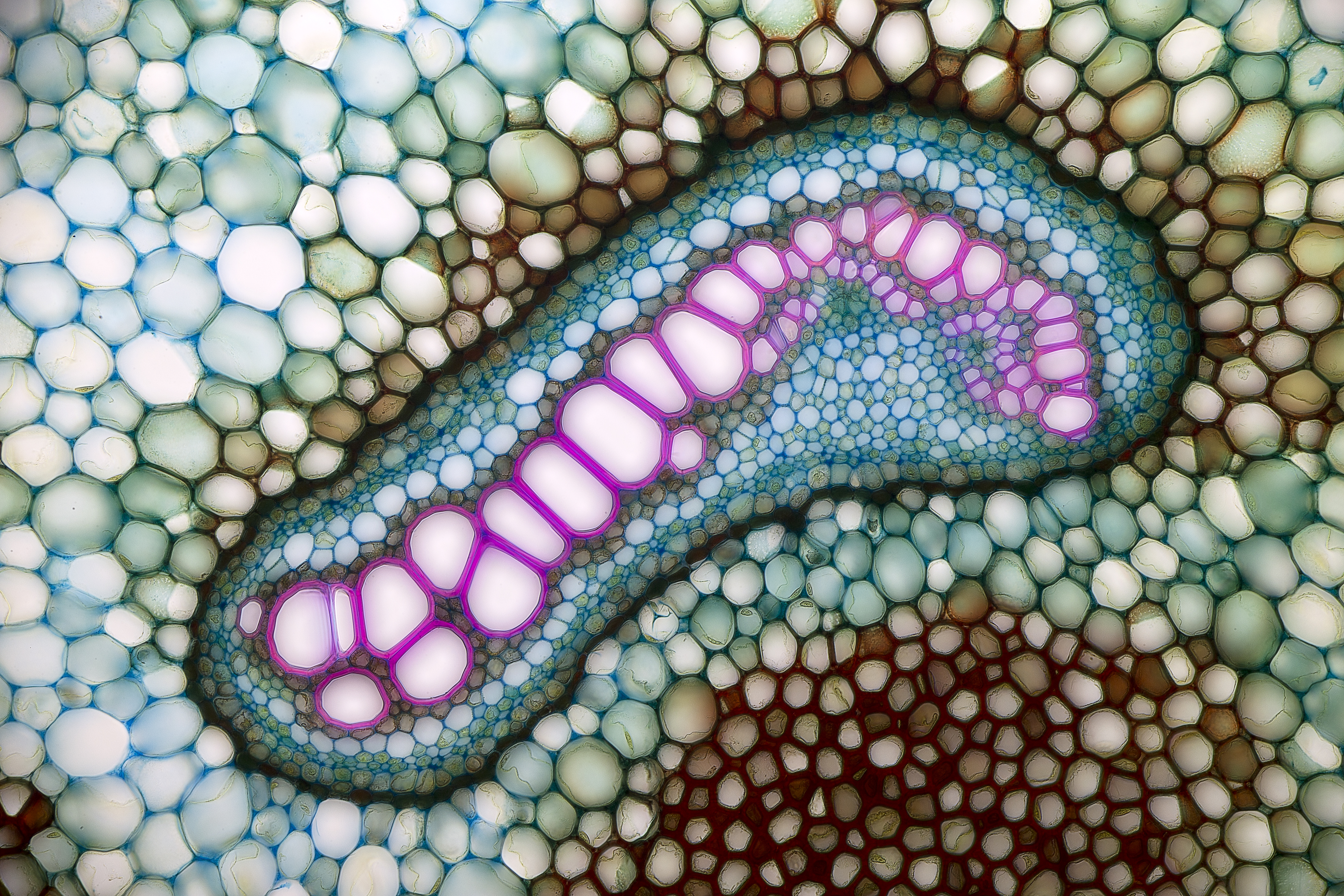
줄기와 뿌리에서 일차 사부와 일차 물관부 사이에 형성된 분열 조직인 관다발 형성층(간단히 형성층이라고도 함)에서 형성된 사부는 '''이차 사부'''(차생 사부, secondary phloem)라고 한다.[26] 이차 사부는 관다발 형성층의 바깥쪽에 추가된다. 목본식물처럼 관다발 형성층에 의한 이차 성장을 하는 식물은 이차 사부를 가지며, 이차 사부는 수피의 내층에 위치한다. 생물학에서 '''사부'''(靭皮, bast)는 이차 사부를 의미한다.[26] 이차 사부에서는 일차 사부에 비해 구성 요소가 방사 방향으로 질서정연하게 배열되는 경향이 있으며, 사부 요소가 짧고 밀도가 높다.

이차 사부의 각 사부 요소는 단기간만 기능하며, 안쪽에서 추가되는 새로운 조직에 밀려 붕괴된다. 따라서 이차 물관부의 두께가 매년 증가하는 것과는 달리, 이차 사부의 두께는 어느 정도 이상 변하지 않는다.[26][27]
이차 물관부와 마찬가지로 이차 사부에도 축 방향 요소(사부 요소, 사부 섬유, 축 방향 유조직)와 방사 방향 요소(방사 유조직)가 존재한다. 이들의 기원 세포는 관다발 형성층에 존재하며 이차 물관부와 공유한다. 따라서 이차 사부의 방사 조직은 관다발 형성층을 사이에 두고 이차 물관부의 방사 조직과 연속되어 있다.[27](그림 6c). 또한 이차 성장하는 줄기나 뿌리는 둘레가 증가하므로, 사부 방사 조직은 그것을 메우듯 세포 분열에 의해 바깥쪽으로 넓어지는 경우가 있다(그림 6c). 이러한 방사 조직은 확장 방사 조직(dilated ray)이라고 한다.
4. 체관부 수송 (체관부 전류)
광합성으로 생성된 당은 체관부의 체관 요소(체관)나 사부 세포 조직을 통해 식물체 전체로 수송된다(그림 7). 이 수송은 '''체관부 수송'''(사부 수송) 또는 '''체관부 이동'''(사부 이동)이라고 한다.[32] 체관부 수송으로 운반되는 수용액(체관액)에는 다량의 당(주로 수크로스, 때때로 올리고당 또는 당알코올)과 아미노산, 무기 양분, 식물 호르몬, 전령 RNA(mRNA) 등이 포함되어 식물체 전체의 정보 교환 통로가 되기도 한다. 체관부 수송은 체관 단면적 1 cm2당 1시간에 수 그램의 물질을 수송하며, 수송 속도는 시속 1m에 달하기도 한다.
물관부 수송이 뿌리에서 기공으로 향하는 일방 통행인 것과 달리, 체관부 수송 방향은 시기와 식물 상태에 따라 변한다. 체관액 공급 쪽을 '''원천'''(source), 받는 쪽을 '''저장소'''(sink)라고 한다. 저장 기관은 광합성이 활발할 때는 저장소, 초봄처럼 활발한 활동 전에는 원천이 되는 등, 같은 기관이 시기에 따라 원천/저장소 역할을 바꾼다. 원천과 저장소 사이 체관부 수송은 기본적으로 저장소보다 원천 쪽 삼투압이 높아 발생하는 물 흐름 때문이다. 이 가설은 '''압력 흐름설'''(pressure flow theory)로, 엘른스트 뮌히 (1930)가 제창했다. 단, 속씨식물 이외의 체관부 수송(사부 세포 조직 이용)이 압력 흐름설로 설명되는지는 불명확하다.
원천에서 체관부로 물질이 이동하는 것을 적재(체관부 적재, 로딩, phloem loading), 체관부에서 저장소로 이동하는 것을 하역(체관부 하역, 언로딩, phloem unloading)이라고 한다.[32] 심플라스트 경로(원형질 연락 통과)와 아포플라스트 경로(세포벽 등 세포막 외부 통과)가 있다. 예를 들어 원형질 연락으로 운반된 수크로스가 중계 세포(intermediate cell)에서 올리고당으로 변환, 올리고당은 커서 엽육 세포로 못 돌아가 체관 방향 수송이 촉진되기도 한다(폴리머 트래핑). 적재 시 당 농도가 낮은 곳에서 높은 곳으로 수송될 때는 능동 수송이 사용된다. 수크로스가 능동 수송으로 수송 세포(transfer cell)에 흡수되어 체관으로 수송되기도 한다. 하역 시에는 저장소에서 당이 소비되거나 녹말 등 불용성 물질로 변환되어 보통 저장소 쪽 당 농도가 낮다.
체관부를 통한 전기 신호 전달도 알려져 있다. 식물체 일부에서 발생한 전기 자극이 체관부로 전달되어 유전자 발현, 호흡, 광합성, 식물 호르몬 양, 체관부 하역 등에 변화를 일으킨다.
5. 한국과 체관부
소나무 속껍질(체관부 포함)은 흉년에 구황식물로 섭취되었다.[39] 송피떡은 소나무 인피를 이용하며, 구황식물로 만들어졌다는 이야기가 전해진다.[40] 
닥나무(뽕나무과)나 꾸지뽕나무의 체관부는 한지 원료로 사용된다.[33][34]
5. 1. 전통적 이용
소나무의 속껍질(체관부 포함)은 흉년 때 구황식물로 섭취되었다.[39] 송피떡은 소나무의 인피를 이용하며, 구황식물로 만들어졌다는 이야기가 전해진다.[40]닥나무(뽕나무과)나 꾸지뽕나무의 체관부는 한지의 원료로 사용된다.[33][34]
5. 2. 임업 및 농업에서의 활용
목본식물에서 체관부는 수피 내층에 존재하기 때문에, 수피를 환상으로 박리하면 체관부를 절단할 수 있으며, 이 기법은 '''환상박피'''(環狀剝皮, girdling, ring barking)라고 한다. 환상박피는 임업, 농업, 원예 등 다양한 분야에서 이용된다.예를 들어, 나무줄기에서 환상박피를 하면 그 식물을 고사시킬 수 있다(감아말리기). 비버는 종종 환상박피를 하며, 도입된 아르헨티나에서 나무를 대규모로 고사시킨 사례가 보고되었다.[44] 임업에서는 간벌 작업의 노력 절감 및 안전화를 위해 환상박피가 이용되고 있다.[45][46] 그러나 이 작업은 임내에 다수의 고사목을 발생시키게 되어, 고사목을 이용하는 해충 증가 등의 위험이 우려되고 있다. 또한 외래 수종 제거에도 유사한 방법이 이용된다.[47]
가지 등에 환상박피를 하면 그 부분 위쪽에 당 등 동화산물이 남는다. 다양한 수종에서 환상박피가 수세 억제, 개화 촉진, 생리낙과 억제, 과실 품질 향상 등에 효과적이라는 보고가 있으며, 포도, 감, 감귤류 등에서는 실용화되고 있다.[48][49][50][51] 환상박피된 부분 위에서 부정근 형성이 촉진되는 경우가 있으므로, 임업, 농업, 원예에서 이것을 이용하여 영양번식시키는 경우도 있다(휘묻이라고 한다).[52][53]
5. 3. 현대적 연구 및 이용
최근에는 체관부의 기능 및 수송 메커니즘에 대한 연구가 활발하게 진행되고 있으며, 이를 통해 식물 생장 조절 및 작물 생산성 향상에 기여할 수 있을 것으로 기대된다. 체관 수액은 관다발 식물 전체에 정보 신호를 전달하는 역할을 하는 것으로 알려져 있다. 적재 및 하역 패턴은 주로 전도도와 원형질 연락사의 수, 그리고 용질 특이적 세포막 수송 단백질의 위치 의존적 기능에 의해 결정된다. 최근 연구에 따르면 이동성 단백질과 RNA는 식물의 장거리 통신 신호 시스템의 일부이다. 원형질 연락사를 통과하는 거대분자의 지향적 수송 및 분류에 대한 증거도 존재한다.[17]당, 아미노산, 특정 식물 호르몬, 그리고 심지어 메신저 RNA와 같은 유기 분자는 체관 요소를 통해 체관부에서 수송된다.[17]
체관부는 또한 초파리 ''Drosophila montana''를 포함한 파리목 곤충의 산란과 번식을 위한 인기 있는 장소로 사용된다.[18]
참조
[1]
논문
Transport mechanisms for organic forms of carbon and nitrogen between source and sink
[2]
서적
Collins Edexcel International GCSE Biology, Student Book
[3]
어원사전
phloem
[4]
사전
φλοιός
[5]
논문
Das Wachstum des Stammes und der Wurzel bei den Gefäßpflanzen und die Anordnung der Gefäßstränge im Stengel
https://www.biodiver[...]
1858
[6]
서적
Ontogeny, Cell Differentiation, and Structure of Vascular Plants
[7]
논문
Metaphloem development in the Arabidopsis root tip
https://journals.bio[...]
2021
[8]
서적
Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development
https://onlinelibrar[...]
Wiley
2006-08-25
[9]
논문
Vascular Cambium Development
2015-05-00
[10]
서적
Biology of Plants
Worth Publishers
[11]
논문
Structural and functional heterogeneity in phloem loading and transport
2013-07-05
[12]
서적
Plant physiology, development and metabolism
https://www.worldcat[...]
2018
[13]
논문
Ripening-related changes in the cell walls of Spanish pear (Pyrus communis)
https://onlinelibrar[...]
1994
[14]
논문
Phloème, transport interorgane et signalisation à longue distance
https://www.scienced[...]
2008-05-01
[15]
논문
Modelling the Fate of Chemicals in Plants
[16]
서적
Phloem Translocation
[17]
논문
Phloem Transport: Cellular Pathways and Molecular Trafficking
[18]
논문
Larval niche differences between the sibling species, Drosophila montana and D. littoralis(Diptera) in Northern Finland
https://www.research[...]
1996-01-01
[19]
논문
Dynamics of long-distance signaling via plant vascular tissues
2015
[20]
논문
"The Plant Vascular System " Evolution, Development and Functions
[21]
논문
Phloem development
2023
[22]
서적
Esau's Plant Anatomy
John Wiley & Sons, Inc
[23]
논문
Reconstructing Food Ways: Role of Skolt Sami Cultural Revitalization Programs in Local Plant Use
http://www.bioone.or[...]
2016
[24]
웹사이트
Using Tree Bark Flours in Cooking
https://waldenlabs.c[...]
2016-09-29
[25]
백과사전
Phloem
https://www.merriam-[...]
Merriam-Webster
2020-04-20
[26]
서적
図説 植物用語事典
八坂書房
2001
[27]
서적
Plant Structure: A Colour Guide
Jones & Bartlett Learning
2008
[28]
서적
維管束植物の形態と進化
文一総合出版
2002
[29]
서적
An Introduction to Plant Structure and Development
Cambridge University Press
2005
[30]
서적
植物の系統
文一総合出版
1999
[31]
웹사이트
師管について
https://jspp.org/hir[...]
日本植物生理学会
2006-01-23
[32]
서적
テイツ/ザイガー 植物生理学・発生学
講談社
2017
[33]
논문
繊維作物の種類および利用と植物分類上の区分との関係
1980
[34]
논문
非木材繊維利用の現状と将来
1997
[35]
논문
サトウヤシとヤシ砂糖,ヤシ酒
1959
[36]
논문
ヤシ科植物のバイオテクノロジー: スリランカとインドネシアの事例
http://www.sagopalm.[...]
2003
[37]
서적
Transport in Plants I. Phloem Transport
Springer, Berlin
1975
[38]
논문
アガベから造られるテキーラ、イヌリンなどの製品およびイヌリンの研究経過報告
http://www.tervis.co[...]
2007
[39]
논문
Death and disease during the great Finnish famine 1695-1697
2014
[40]
논문
松皮に含まれる食物繊維の加熱・貯蔵中の変化
2010
[41]
논문
ニホンジカによる人工林剥皮害と下層植生との関連性
日本森林学会
2007
[42]
서적
新常用和漢薬集
http://www.tokyo-sho[...]
東京生薬協会
2020-03-28
[43]
논문
キハダ優良品種の増殖と徳島県下におけるキハダ造林に関する調査研究
http://www.pref.toku[...]
2001
[44]
논문
Tierra del Fuego: the beavers must die
2008
[45]
논문
ヒノキの巻き枯らし間伐実証試験
https://agriknowledg[...]
群馬県林業試験場
2009
[46]
논문
ヒノキ巻枯らし間伐林における森林害虫の発生
https://agriknowledg[...]
2009
[47]
논문
特集「外来種と植生管理」 巻き枯らしによるハリエンジュの管理
2015
[48]
논문
環状はく皮処理が主幹形モモの生育と果実品質に及ぼす影響
https://agriknowledg[...]
2007
[49]
논문
キウイフルーツの生育期の環状はく皮による果実肥大効果、果実品質及び樹体への影響
https://agriknowledg[...]
愛媛県立果樹試験場
2002
[50]
논문
ブドウ‘安芸クイーン’の着色実態および環状はく皮と着果量の軽減による着色改善
2007
[51]
논문
カキ‘西村早生’の雄花と雌花の着生に及ぼす側枝の環状はく皮ならびに結縛の影響
https://kochi.repo.n[...]
1995
[52]
논문
環状はく皮処理による亜熱帯系桑古条さし木の時期別露地実験
日本蚕糸学会
1986
[53]
논문
各都道府県の林業・林産業と遺伝育種の関わり(24)石川県
森林遺伝育種学会
2019
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com