키노사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
키노사우루스는 '개'를 뜻하는 그리스어 'kyon'과 '도마뱀'을 뜻하는 'sauros'에서 유래된 이름으로, 남아프리카 공화국의 카루 누층군에서 화석이 발견된 멸종된 키노돈트의 속이다. 1876년 리처드 오언에 의해 처음 기술되었으며, 콧구멍의 위치와 어금니의 특징을 통해 키노캠사와 유사점이 묘사되었다. 키노사우루스는 넓은 주둥이, 튼튼한 아래턱, 그리고 두개골의 특징을 가지며, 두정공의 유무와 치아 형태에 따라 다른 초기 키노돈트와 구별된다. 남아프리카 공화국 발푸어 층의 키스테케팔루스 화석대와 답토케팔루스 화석대에서 발견되며, 충적평원과 호수 환경에서 서식했을 것으로 추정된다.
더 읽어볼만한 페이지
- 페름기 아프리카의 단궁류 - 리스트로사우루스
리스트로사우루스는 후기 페름기부터 초기 트라이아스기에 번성한 멧돼지 크기의 디키노돈트 수궁류 속으로, 페름기-트라이아스기 대멸종 이후 판게아 대륙에 빠르게 퍼져나가 초기 트라이아스기 생태계를 지배했으며, 이는 대륙 이동설의 중요한 증거가 된다. - 1927년 기재된 화석 분류군 - 베이징 원인
베이징 원인은 중국에서 발견된 호모 에렉투스 화석으로, 석기 사용, 불 사용, 직립 보행 등의 특징을 보이며, 현생 인류의 아시아 기원설을 뒷받침하는 증거로 여겨졌으나 현재는 아프리카 기원설이 정설로 받아들여진다. - 생흔 화석 - 네오사우로이데스
- 생흔 화석 - 위석
위석은 동물, 특히 악어, 조류, 공룡 등이 음식 분쇄, 부력 조절, 칼슘 저장을 위해 위 내에 보관하는 돌을 의미하며, 척추동물, 멸종된 공룡 등 다양한 종에서 발견된다.
| 키노사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
| 모식종 | Cynosaurus suppostus |
| 모식종 명명자 | Owen, 1859 |
| 이명 | Baurocynodon Brink, 1951 Cynosuchoides Broom, 1931 Cynosuchus Owen, 1876 Mygalesuchus Broom, 1942 |
| 생물학적 분류 | |
| 속 | †키노사우루스속 |
| 종 | †키노사우루스 서포스투스 |
| 계 | 진핵생물역 |
| 문 | 척삭동물문 |
| 아문 | 척추동물아문 |
| 상강 | 사지상강 |
| 강 | 단궁강 |
| 목 | 수궁목 |
| 아목 | 키노돈아목 |
| 미분류 | 상키노돈트 |
| 시간 척도 | |
| 화석 범위 | 페름기 후기, 에서 |
2. 어원
속명 '''키노사우루스'''의 Cyno-는 그리스어로 '개'를 뜻하는 'kyon'과 그리스어로 '도마뱀'을 뜻하는 'sauros'에서 유래되었다.
키노사우루스의 화석은 남아공의 카루 누층군과 관련된 '시스테세팔루스'(Cistecephalus)와 '답토세팔루스 군집지대'(Daptocephalus Assemblage Zone)에서 발견되었다.[12] ''키노사우루스''(Cynosaurus)는 1876년 리처드 오언에 의해 처음으로 기술되었다.[2] 오언은 1876년에 "영국 박물관 소장 남아프리카 화석 파충류의 기술 및 삽화 카탈로그"라는 제목의 논문을 작성했다. 오언은 화석의 이름을 ''키노수쿠스 서포스투스''(Cynosuchus suppostus) 오언, 1876으로 명명했는데, 이는 1927년 K. 슈미트에 의해 나중에 ''키노사우루스''로 이름이 변경되었다.[2] 오언은 ''키노수쿠스 서포스투스''를 앞니와 송곳니의 위치가 키노캠사와 유사하다고 묘사했다. 차이점은 ''키노수쿠스''가 콧구멍의 위치가 더 작고 위쪽을 향하고 있다는 것이다. ''키노수쿠스 서포스투스''의 바깥쪽 콧구멍은 위턱과 아래턱 앞부분과 위치가 가까웠고 콧구멍은 거의 수평이었다. 오언은 어금니가 상대적으로 크다고 묘사했다. 오언은 또한 위턱이 뒤로 물러나면서 수축되고 큰 어금니와 결합되어 ''키노수쿠스 서포스투스''가 더 넓고 짧은 두개골을 가지고 있음을 보여준다고 언급했다. 비골은 넓고 두꺼우며 상악골에 의해 겹쳐져 있었다.[3]
3. 발견 및 역사
3. 1. 남아프리카에서의 발견
키노사우루스의 화석은 남아공의 카루 누층군과 관련된 '시스테세팔루스'(Cistecephalus)와 '답토세팔루스 군집지대'(Daptocephalus Assemblage Zone)에서 발견되었다.[12] ''키노사우루스''(Cynosaurus)는 1876년 리처드 오언에 의해 처음으로 기술되었다.[2] 오언은 ''키노수쿠스 서포스투스''(Cynosuchus suppostus)를 앞니와 송곳니의 위치가 키노캠사와 유사하다고 묘사했다. 차이점은 ''키노수쿠스''가 콧구멍의 위치가 더 작고 위쪽을 향하고 있다는 것이다. ''키노수쿠스 서포스투스''의 바깥쪽 콧구멍은 위턱과 아래턱 앞부분과 위치가 가까웠고 콧구멍은 거의 수평이었다. 오언은 어금니가 상대적으로 크다고 묘사했다. 오언은 또한 위턱이 뒤로 물러나면서 수축되고 큰 어금니와 결합되어 ''키노수쿠스 서포스투스''가 더 넓고 짧은 두개골을 가지고 있음을 보여준다고 언급했다. 비골은 넓고 두꺼우며 상악골에 의해 겹쳐져 있었다.[3]
4. 특징
키노사우루스에서 파생된 특징은 다음과 같다. 하악 앞부분의 수직에 가까운 멘텀, 상대적으로 높은 수평 가지가 있는 튼튼한 아래턱, 두개골 길이의 최대 32%에 달하는 넓은 주둥이를 가졌다. 그리고 성체 키노사우루스는 솔방울 구멍이 없다.[4] 초기 키노돈트에서는 두정골이 뇌 상자의 측벽까지 복측으로 뻗어 있다.[5] 또한, 에피프테리고이드는 전두골과 새로운 접촉을 하도록 확장되었으며, 두정 능선은 솔방울 구멍을 통합하도록 길어졌다.[5]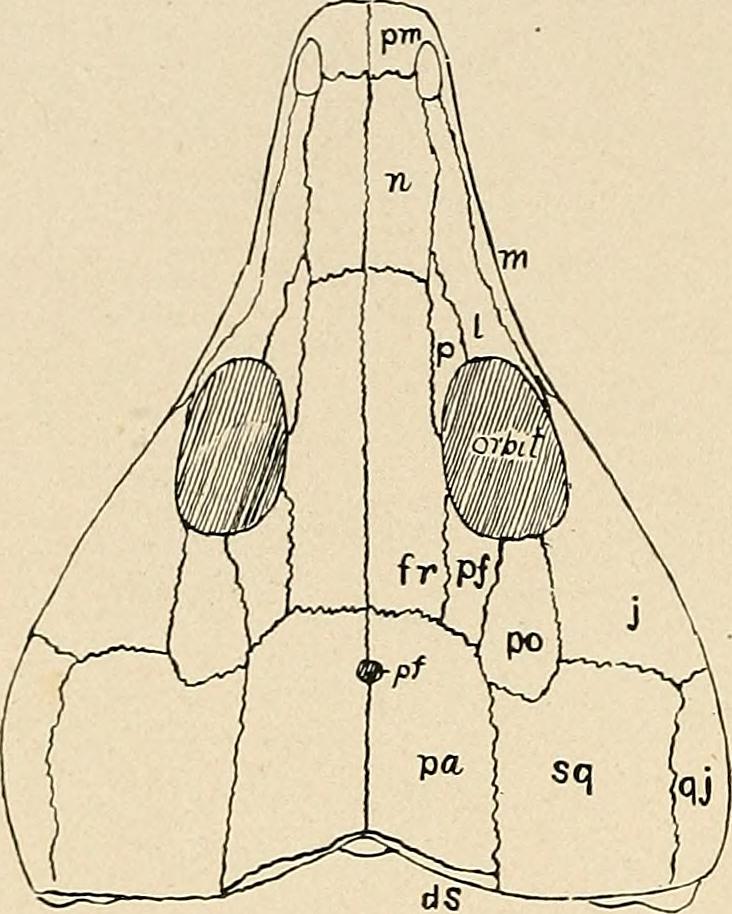
두개골셉토맥실라는 코뼈를 상하로 나누는 평평한 다리 역할을 한다.[4] 코뼈는 앞쪽보다 뒤쪽이 더 넓다.[4] 상악골 표면에는 수평으로 나란한 두 줄을 이루는 작은 영양공이 많이 있다.[4] 전상악골의 경우 전상악골과 구개 돌기 사이에 정중선을 따라 틈이 있다.[4] 서골은 한 쌍이 아니며 뾰족해지면서 날카로운 점에 도달한다.[4] 서골은 뒤쪽의 익상골에도 닿지 않는다.[4] 미세 CT 스캔을 통해 화석 두개골의 내부 구조를 관찰할 수 있다.[6] 미세 CT 스캔 결과, ''키노사우루스''의 4개 표본에서 한 쌍의 골화 궤도접형골이 관찰되었다.[6] 궤도접형골은 두 개의 얇은 판 모양 구조로 구성되어 있고, 복내측으로 연결되어 있으며, 단면에서 U자 모양으로 보인다.[6]
교근 부착 부위의 빠른 진화는 초기 키노돈트의 초기 다양화를 보여줄 수 있다.[7] ''프로키노수쿠스''(Procynosuchus)와 ''드비니아''(Dvinia)에서는 교근 와의 위치가 관상돌기 상부에 위치하는 것이 교근의 분화 초기 단계로 보인다.[7] ''키노사우루스''와 ''나니크토사우루스''(Nanictosaurus)에서는 교근 와가 치골 기저부까지 확장되어 있다.[7]
두정공''키노사우루스''에는 두정공의 위치 근처에서 납작해진 날카로운 시상 능선이 있다.[8] ''키노사우루스'' 두개골의 CT 스캔에서는 두정 튜브가 존재하지 않았지만 대신 뇌내강이 위쪽으로 밀려 올라갔다.[8] 표본인 ''키노사우루스 와이츠''에서는 두정공이 없는 것으로 나타났다.[8] 또 다른 ''키노사우루스'' 두개골 표본에서 두정공의 부재는 성체에서 두정공이 닫히는 ''마세토그나투스''와 마찬가지로 발생학적 변화 때문이었다.[8] 현존하는 도마뱀인 아놀리스 카롤리넨시스에서는 송과선 개구부의 크기가 감소하지만 사라지지는 않는다.[8] 또 다른 표본은 두정 튜브의 증거를 보여주었지만, 부재는 발생학이 아닌 종내 변이에서 비롯되었다.[8]
많은 도마뱀은 머리 꼭대기에 두정안을 가지고 있다.[9] 적도 근처에 사는 현존하는 외온동물은 환경의 안정성으로 인해 셋째 눈이 쓸모없게 되어 송과선 개구부를 갖는 빈도가 덜하다.[8] 도마뱀의 위도 분포와 두정안 발생 사이에는 명확한 관계가 있다.[9] 두정안이 없는 도마뱀은 저위도로, 이는 적도 특성을 시사한다.[9]
치아''키노사우루스''는 띠가 없는 타원형의 단순한 송곳니를 가지고 있다.[10] 뒤쪽 송곳니는 뒤쪽 부속 돌기이며, ''키노사우루스''는 가장 뒤쪽 치아에 두 번째 뒤쪽 부속 돌기를 가지고 있다.[10] ''키노사우루스''의 앞쪽 부속 돌기는 보이지 않는다.[10] 대부분의 초기 키노돈트는 옆면에서 삼첨치아형 뒷 송곳니를 보인다.[10]
''프로키노수쿠스 델라하르페아이''와 ''드비니아''는 ''키노사우루스''보다 기저에 있으며, 대부분의 키노돈트가 위쪽 4개, 아래쪽 3개의 앞니를 가지는 반면, 5개 이상의 위쪽 앞니와 4개 이상의 아래쪽 앞니를 가지고 있다.[10] ''프로갈레사우루스'' 또한 ''키노사우루스''보다 기저에 있으며, 송곳니에 강한 세로 홈 또는 줄무늬가 있다.[4] ''키노사우루스''보다 파생된 ''갈레사우루스''는 뒤쪽에 불완전한 뼈로 된 두 번째 구개 돌기를 가지고 있다.[4]
4. 1. 두개골 (Cranium)
셉토맥실라는 코뼈를 상하로 나누는 평평한 다리 역할을 한다.[4] 코뼈는 앞쪽보다 뒤쪽이 더 넓다.[4] 상악골 표면에는 수평으로 나란한 두 줄을 이루는 작은 영양공이 많이 있다.[4] 전상악골의 경우 전상악골과 구개 돌기 사이에 정중선을 따라 틈이 있다.[4] 서골은 한 쌍이 아니며 뾰족해지면서 날카로운 점에 도달한다.[4] 서골은 뒤쪽의 익상골에도 닿지 않는다.[4] 미세 CT 스캔을 통해 화석 두개골의 내부 구조를 관찰할 수 있다.[6] 미세 CT 스캔 결과, ''키노사우루스''의 4개 표본에서 한 쌍의 골화 궤도접형골이 관찰되었다.[6] 궤도접형골은 두 개의 얇은 판 모양 구조로 구성되어 있고, 복내측으로 연결되어 있으며, 단면에서 U자 모양으로 보인다.[6]교근 부착 부위의 빠른 진화는 초기 키노돈트의 초기 다양화를 보여줄 수 있다.[7] ''프로키노수쿠스''(Procynosuchus)와 ''드비니아''(Dvinia)에서는 교근 와의 위치가 관상돌기 상부에 위치하는 것이 교근의 분화 초기 단계로 보인다.[7] ''키노사우루스''와 ''나니크토사우루스''(Nanictosaurus)에서는 교근 와가 치골 기저부까지 확장되어 있다.[7]
4. 2. 두정공 (Parietal foramen)
''키노사우루스''에는 두정공의 위치 근처에서 납작해진 날카로운 시상 능선이 있다.[8] ''키노사우루스'' 두개골의 CT 스캔에서는 두정 튜브가 존재하지 않았지만 대신 뇌내강이 위쪽으로 밀려 올라갔다.[8] 표본인 ''키노사우루스 와이츠''에서는 두정공이 없는 것으로 나타났다.[8] 또 다른 ''키노사우루스'' 두개골 표본에서 두정공의 부재는 성체에서 두정공이 닫히는 ''마세토그나투스''와 마찬가지로 발생학적 변화 때문이었다.[8] 현존하는 도마뱀인 아놀리스 카롤리넨시스에서는 송과선 개구부의 크기가 감소하지만 사라지지는 않는다.[8] 또 다른 표본은 두정 튜브의 증거를 보여주었지만, 부재는 발생학이 아닌 종내 변이에서 비롯되었다.[8]많은 도마뱀은 머리 꼭대기에 두정안을 가지고 있다.[9] 적도 근처에 사는 현존하는 외온동물은 환경의 안정성으로 인해 셋째 눈이 쓸모없게 되어 송과선 개구부를 갖는 빈도가 덜하다.[8] 도마뱀의 위도 분포와 두정안 발생 사이에는 명확한 관계가 있다.[9] 두정안이 없는 도마뱀은 저위도로, 이는 적도 특성을 시사한다.[9]
4. 3. 치아 (Tooth)
''키노사우루스''는 띠가 없는 타원형의 단순한 송곳니를 가지고 있다.[10] 뒤쪽 송곳니는 뒤쪽 부속 돌기이며, ''키노사우루스''는 가장 뒤쪽 치아에 두 번째 뒤쪽 부속 돌기를 가지고 있다.[10] ''키노사우루스''의 앞쪽 부속 돌기는 보이지 않는다.[10] 대부분의 초기 키노돈트는 옆면에서 삼첨치아형 뒷 송곳니를 보인다.[10]''프로키노수쿠스 델라하르페아이''와 ''드비니아''는 ''키노사우루스''보다 기저에 있으며, 대부분의 키노돈트가 위쪽 4개, 아래쪽 3개의 앞니를 가지는 반면, 5개 이상의 위쪽 앞니와 4개 이상의 아래쪽 앞니를 가지고 있다.[10] ''프로갈레사우루스'' 또한 ''키노사우루스''보다 기저에 있으며, 송곳니에 강한 세로 홈 또는 줄무늬가 있다.[4] ''키노사우루스''보다 파생된 ''갈레사우루스''는 뒤쪽에 불완전한 뼈로 된 두 번째 구개 돌기를 가지고 있다.[4]
5. 고생태 (Paleoenvironment)
키노사우루스의 화석은 남아프리카 공화국의 카루 초군에 속하는 보퍼트 그룹의 발푸어 층에 있는 키스테케팔루스 화석대와 답토케팔루스 화석대에서 발견되었다.[4] 남아프리카 공화국의 카루 분지에서는 강둑이 범람하여 토양 축적이 시작될 수 있는 충적평원을 만들었을 것이다.[11] 발푸어 층 하부의 토양 퇴적물은 잎 자국이 풍부한 호수 환경을 시사한다.[11] 이는 해안 습지와 늪이 있었음을 시사한다.[11] 또한 이 지층에서는 수생 생물의 흔적 화석이 발견되었다.[11]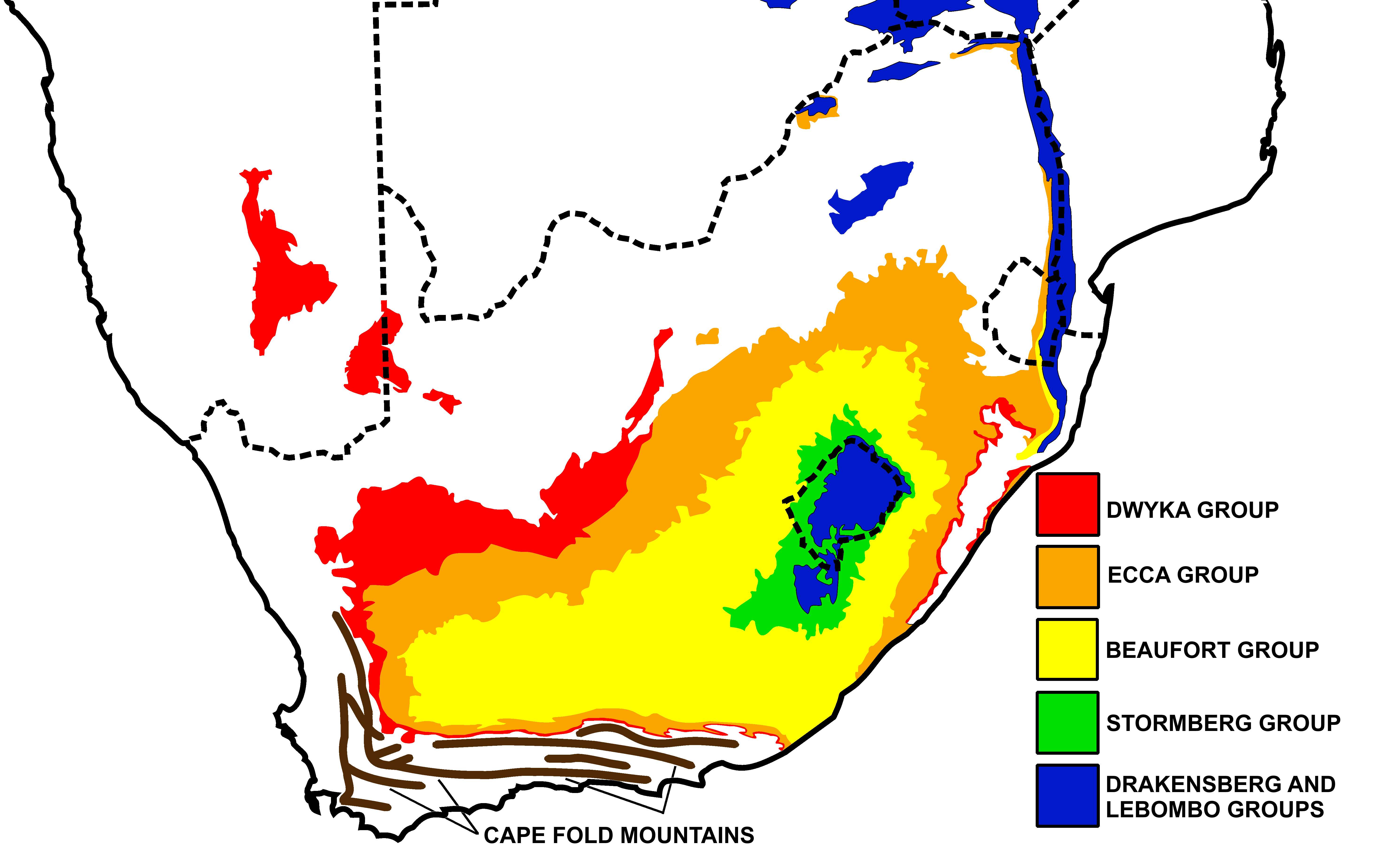
참조
[1]
서적
The Origin and Evolution of Mammals
Oxford University Press
2005
[2]
논문
New reptilian generic names
1927
[3]
서적
Descriptive and illustrated catalog of the fossil reptilia of South Africa in the collection of the British Museum
1876
[4]
간행물
Cranial morphology and phylogenetic analyses of Cynosaurus suppostus (Therapsida, Cynodontia) from the upper Permian of the Karoo Basin, South Africa
2018
[5]
간행물
Evolutionary patterns among Permo-Triassic therapsids
[6]
간행물
The mystery of a missing bone: revealing the orbitosphenoid in basal Epicynodontia (Cynodontia, Therapsida) through computed tomography
2017
[7]
간행물
The oldest cynodont: new clues on the origin and early diversification of the Cynodontia
2007
[8]
간행물
Physiological implications of the abnormal absence of the parietal foramen in a late Permian cynodont (Therapsida)
2015
[9]
간행물
The pineal gland and geographical distribution of animals
1975
[10]
간행물
A new cynodont record from the Tropidostoma Assemblage Zone of the Beaufort Group: implications for the early evolution of cynodonts in South Africa
2007
[11]
간행물
Changing palaeoenvironments and tetrapod populations in the Daptocephalus Assemblage Zone (Karoo Basin, South Africa) indicate early onset of the Permo-Triassic mass extinction
2018
[12]
저널
Cranial morphology and phylogenetic analysis of Cynosaurus suppostus (Therapsida, Cynodontia) from the Upper Permian of the Karoo Basin, South Africa.
https://ri.conicet.g[...]
2018-03
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com