남극빙어과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
남극빙어과는 남극해에 서식하는 어류로, 헤모글로빈과 적혈구가 없어 혈액이 무색인 특징을 갖는다. 어식성이며 매복형 포식자이고, 몸길이는 25~50cm이다. 헤모글로빈과 미오글로빈의 손실은 산소 운반 능력 저하를 가져왔지만, 큰 혈관, 많은 혈액량, 큰 심장, 높은 심박출량 등의 생리적 적응을 통해 이를 보완한다. 남극빙어는 단일 계통군으로, 저서성 조상으로부터 진화했으며, 남극 전선대와 남극 순환 해류의 형성 이후 종 분화가 일어났다.
더 읽어볼만한 페이지
| 남극빙어과 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Channichthyidae |
| 명명자 | T. N. Gill, 1861 |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 조기어강 |
| 상목 | 극기류 |
| 목 | 농어목 |
| 아목 | 노토테니아아목 |
| 과 | 남극빙어과 |
| 속 | |
| 속 | 카에노케팔루스 카에노드라코 캄프소케팔루스 찬니크티스 키오노바티스쿠스 키오노드라코 크리오드라코 다코드라코 네오파게톱시스 파게톱시스 프세우도카에니크티스 |
| 특징 | |
| 서식 환경 | 남극 해역 |
| 수온 | (약 -1.8°C에서 2°C) |
| 연구 | 2022년, 남극해에서 거대한 번식지가 발견됨. |
2. 하위 속
| 속명 | 명명자 및 명명 연도 |
|---|---|
| 차노케팔루스 | 리처드슨, 1844 |
| 차노드라코 | 리건, 1914 |
| 샴소케팔루스 | 길, 1861 |
| 찬니크티스 | 리처드슨, 1844 |
| 키오노바티스쿠스 | 안드리야셰프 & 네옐로프, 1978 |
| 키오노드라코 | 뢴베르그, 1905 |
| 크리오드라코 | 돌로, 1900 |
| 다코드라코 | 웨이트, 1916 |
| 네오파제토프시스 | 니벨린, 1947 |
| 파제토프시스 | 리건, 1913 |
| 슈도차니크티스 | 노먼, 1937 |
[7][8]
얼음물고기는 어식동물(piscivore)로 알려져 있지만, 크릴새우도 섭취한다[33]。 얼음물고기는 매복형 포식자(Ambush predator)로, 몸길이의 50%에 달하는 물고기를 먹을 수 있지만, 오랜 기간 먹지 않고도 생존할 수 있다. 몸길이는 25cm 정도이다.[34]。
3. 특징
2022년, 독일 연구팀은 남극해 심해에서 약 6,000만 개에 달하는 검은얼음물고기 둥지가 모인 번식지를 발견했다. 이 번식지는 바다코끼리의 서식지 및 활동 영역과 일치하며, 바다코끼리에게 포식당하는 것으로 추정된다.[35]。
3. 1. 식성 및 몸 크기
모든 남극빙어는 어식성 어류이지만, 크릴을 먹기도 한다.[9] 남극빙어는 전형적인 매복 포식자이다. 따라서 먹이를 먹지 않고도 오랫동안 생존할 수 있으며, 종종 자기 몸길이의 최대 50%에 달하는 물고기를 섭취한다. 남극빙어의 최대 몸길이는 25cm로 기록되었다.[10]
3. 2. 순환계 및 호흡계
남극빙어의 혈액은 무색 투명한데, 이는 혈액 내 산소 결합 단백질인 헤모글로빈이 없기 때문이다.[11][12] 적혈구 또한 거의 존재하지 않거나, 있더라도 극히 드물고 기능이 정지된 상태이다.[15] 대신 산소는 혈장에 용해되어 운반된다. 낮은 온도에서는 기체의 용해도가 증가하는 경향이 있어, 물의 낮은 온도에서 산소의 높은 용해도와 낮은 대사율을 통해 남극빙어는 헤모글로빈 없이 생존 가능하다.[11]
하지만 헤모글로빈을 가진 다른 어류에 비해 산소 운반 능력은 10% 미만이다.[16] 이를 보완하기 위해 남극빙어는 일반적인 어류보다 더 큰 혈관(모세 혈관 포함), 더 많은 혈액량(4배), 더 큰 심장, 더 큰 심박출량(5배)을 가진다.[11]
심장에는 관상 동맥이 없지만, 심실 근육이 스펀지처럼 되어 있어 혈액에서 직접 산소를 흡수할 수 있다.[19] 또한, 비늘이 없는 피부를 통해 산소를 흡수한다고 알려졌었지만, 현재는 피부보다 아가미를 통해 흡수하는 산소의 양이 훨씬 더 많다고 밝혀졌다.[19] 피부를 통해 흡수되는 소량의 산소는 심장에 산소를 보충하는 역할을 할 수 있다.[19]
남극빙어는 적혈구를 가진 남극어류에 비해 더 큰 심장 미토콘드리아와 증가된 미토콘드리아 생성을 가지고 있다.[21][22] 이는 미토콘드리아 표면적을 증가시키고 세포외 영역과 미토콘드리아 사이의 거리를 줄여 산소 공급을 원활하게 한다.
3. 2. 1. 헤모글로빈 결핍
남극빙어의 혈액은 혈액 내 산소 결합 단백질인 헤모글로빈이 없기 때문에 무색이다.[11][12] 남극빙어과(Channichthyidae)는 성체가 되었을 때 헤모글로빈이 없는 것으로 알려진 유일한 척추동물이다. 헤모글로빈을 생성하지 않지만, 헤모글로빈 유전자의 잔재는 그들의 게놈에서 발견될 수 있다. 헤모글로빈 단백질은 두 개의 하위 단위(알파와 베타)로 구성된다. 16종의 남극빙어 중 15종에서 베타 하위 단위 유전자가 완전히 삭제되었고 알파 하위 단위 유전자는 부분적으로 삭제되었다.[13] 한 종의 남극빙어, ''Neopagetopsis ionah''는 더 완전하지만 여전히 기능하지 않는 헤모글로빈 유전자를 가지고 있다.[14]
적혈구 (RBC)는 일반적으로 존재하지 않으며, 존재하더라도 드물고 기능이 정지된 상태이다.[15] 산소는 혈장에 용해되어 헤모글로빈 단백질 없이 온몸으로 운반된다. 물의 낮은 온도에서 산소의 높은 용해도와 낮은 대사율을 통해 남극빙어는 헤모글로빈 없이 살 수 있다(기체의 용해도는 온도가 감소함에 따라 증가하는 경향이 있다).[11] 그러나, 남극빙어 혈액의 산소 운반 능력은 헤모글로빈을 가진 친척의 10% 미만이다.[16]
근육에서 사용되는 산소 결합 단백질인 미오글로빈은 모든 남극빙어 골격근에서 존재하지 않는다. 10종의 경우 미오글로빈은 심장 근육, 특히 심실에서 발견된다.[17] 남극빙어 심장 심실에서 미오글로빈 유전자 발현의 손실은 최소 4번 이상 발생했다.[11][18]
헤모글로빈의 부재를 보상하기 위해 남극빙어는 다른 물고기에 비해 더 큰 혈관(모세 혈관 포함), 더 많은 혈액량(다른 물고기의 4배), 더 큰 심장, 더 큰 심장 박출량(5배 더 큼)을 가지고 있다.[11] 그들의 심장에는 관상 동맥이 없고 심실 근육은 매우 스펀지 같아서 펌프질하는 혈액에서 직접 산소를 흡수할 수 있다.[19] 그들의 심장, 큰 혈관 및 낮은 점성(RBC가 없는) 혈액은 저압에서 매우 높은 유량을 수행하도록 특화되었다.[20] 이것은 헤모글로빈의 부족으로 인한 문제를 줄이는 데 도움이 된다. 과거에는 그들의 비늘이 없는 피부가 산소 흡수에 도움이 된다고 널리 생각되었다. 그러나 현재 분석에 따르면 피부에 의해 흡수되는 산소의 양은 아가미를 통해 흡수되는 양보다 훨씬 적다.[19] 피부에 의해 흡수되는 약간의 추가 산소는 아가미로 펌프질하기 전에 피부와 신체에서 정맥혈을 받는 심장에 산소 공급을 보충하는 데 역할을 할 수 있다.[19] 또한 남극빙어는 적혈구를 가진 남극어류에 비해 더 큰 심장 미토콘드리아와 증가된 미토콘드리아 생성을 가지고 있다.[21][22] 이러한 적응은 미토콘드리아 표면적을 증가시키고 세포외 영역과 미토콘드리아 사이의 거리를 줄임으로써 향상된 산소 공급을 용이하게 한다.
3. 2. 2. 미오글로빈 결핍
남극빙어의 혈액은 혈액 내 산소 결합 단백질인 헤모글로빈이 없기 때문에 무색이다.[11][12] 남극빙어과(Channichthyidae)는 성체가 되었을 때 헤모글로빈이 없는 것으로 알려진 유일한 척추동물이다. 헤모글로빈을 생성하지 않지만, 헤모글로빈 유전자의 잔재는 그들의 게놈에서 발견될 수 있다. 헤모글로빈 단백질은 두 개의 하위 단위(알파와 베타)로 구성된다. 16종의 남극빙어 중 15종에서 베타 하위 단위 유전자가 완전히 삭제되었고 알파 하위 단위 유전자는 부분적으로 삭제되었다.[13] 한 종의 남극빙어, ''Neopagetopsis ionah''는 더 완전하지만 여전히 기능하지 않는 헤모글로빈 유전자를 가지고 있다.[14]
적혈구 (RBC)는 일반적으로 존재하지 않으며, 존재하더라도 드물고 기능이 정지된 상태이다.[15] 산소는 혈장에서 용해되어 헤모글로빈 단백질 없이 온몸으로 운반된다. 물의 낮은 온도에서 산소의 높은 용해도와 낮은 대사율을 통해 남극빙어는 헤모글로빈 없이 살 수 있다(기체의 용해도는 온도가 감소함에 따라 증가하는 경향이 있다).[11] 그러나, 남극빙어 혈액의 산소 운반 능력은 헤모글로빈을 가진 친척의 10% 미만이다.[16]
근육에서 사용되는 산소 결합 단백질인 미오글로빈은 모든 남극빙어 골격근에서 존재하지 않는다. 10종의 경우 미오글로빈은 심장 근육, 특히 심실에서 발견된다.[17] 남극빙어 심장 심실에서 미오글로빈 유전자 발현의 손실은 최소 4번 이상 발생했다.[11][18]
헤모글로빈의 부재를 보상하기 위해 남극빙어는 다른 물고기에 비해 더 큰 혈관(모세 혈관 포함), 더 많은 혈액량(다른 물고기의 4배), 더 큰 심장, 더 큰 심장 박출량(5배 더 큼)을 가지고 있다.[11] 그들의 심장에는 관상 동맥이 없고 심실 근육은 매우 스펀지 같아서 펌프질하는 혈액에서 직접 산소를 흡수할 수 있다.[19] 그들의 심장, 큰 혈관 및 낮은 점성(RBC가 없는) 혈액은 저압에서 매우 높은 유량을 수행하도록 특화되었다.[20] 이것은 헤모글로빈의 부족으로 인한 문제를 줄이는 데 도움이 된다. 과거에는 그들의 비늘이 없는 피부가 산소 흡수에 도움이 된다고 널리 생각되었다. 그러나 현재 분석에 따르면 피부에 의해 흡수되는 산소의 양은 아가미를 통해 흡수되는 양보다 훨씬 적다.[19] 피부에 의해 흡수되는 약간의 추가 산소는 아가미로 펌프질하기 전에 피부와 신체에서 정맥혈을 받는 심장에 산소 공급을 보충하는 데 역할을 할 수 있다.[19] 또한 남극빙어는 적혈구를 가진 남극어류에 비해 더 큰 심장 미토콘드리아와 증가된 미토콘드리아 생성을 가지고 있다.[21][22] 이러한 적응은 미토콘드리아 표면적을 증가시키고 세포외 영역과 미토콘드리아 사이의 거리를 줄임으로써 향상된 산소 공급을 용이하게 한다.
산소를 근육 속에 저장하는 단백질인 미오글로빈은 모든 남극빙어과의 골격근에 존재하지 않는다. 다만, 남극빙어과 10종에서는 심근의 특히 심장의 심실에서 미오글로빈이 발견된다.[42] 남극빙어 심실의 심근에서의 미오글로빈의 유전자 발현은 적어도 4개의 서로 다른 시기에 걸쳐 손실된 것으로 여겨진다.[36][43]
3. 2. 3. 적응
남극빙어의 혈액은 혈액 내 산소 결합 단백질인 헤모글로빈이 없기 때문에 무색이다.[11][12] 남극빙어과(Channichthyidae)는 성체가 되었을 때 헤모글로빈이 없는 것으로 알려진 유일한 척추동물이다. 헤모글로빈을 생성하지 않지만, 헤모글로빈 유전자의 잔재는 그들의 게놈에서 발견될 수 있다. 헤모글로빈 단백질은 두 개의 하위 단위(알파와 베타)로 구성된다. 16종의 남극빙어 중 15종에서 베타 하위 단위 유전자가 완전히 삭제되었고 알파 하위 단위 유전자는 부분적으로 삭제되었다.[13] 한 종의 남극빙어, ''Neopagetopsis ionah''는 더 완전하지만 여전히 기능하지 않는 헤모글로빈 유전자를 가지고 있다.[14]
적혈구 (RBC)는 일반적으로 존재하지 않으며, 존재하더라도 드물고 기능이 정지된 상태이다.[15] 산소는 혈장에서 용해되어 헤모글로빈 단백질 없이 온몸으로 운반된다. 물의 낮은 온도에서 산소의 높은 용해도와 낮은 대사율을 통해 남극빙어는 헤모글로빈 없이 살 수 있다(기체의 용해도는 온도가 감소함에 따라 증가하는 경향이 있다).[11] 그러나, 남극빙어 혈액의 산소 운반 능력은 헤모글로빈을 가진 친척의 10% 미만이다.[16]
근육에서 사용되는 산소 결합 단백질인 미오글로빈은 모든 남극빙어 골격근에서 존재하지 않는다. 10종의 경우 미오글로빈은 심장 근육, 특히 심실에서 발견된다.[17] 남극빙어 심장 심실에서 미오글로빈 유전자 발현의 손실은 최소 4번 이상 발생했다.[11][18]
헤모글로빈의 부재를 보상하기 위해 남극빙어는 다른 물고기에 비해 더 큰 혈관(모세 혈관 포함), 더 많은 혈액량(다른 물고기의 4배), 더 큰 심장, 더 큰 심장 박출량(5배 더 큼)을 가지고 있다.[11] 그들의 심장에는 관상 동맥이 없고 심실 근육은 매우 스펀지 같아서 펌프질하는 혈액에서 직접 산소를 흡수할 수 있다.[19] 그들의 심장, 큰 혈관 및 낮은 점성(RBC가 없는) 혈액은 저압에서 매우 높은 유량을 수행하도록 특화되었다.[20] 이것은 헤모글로빈의 부족으로 인한 문제를 줄이는 데 도움이 된다. 과거에는 그들의 비늘이 없는 피부가 산소 흡수에 도움이 된다고 널리 생각되었다. 그러나 현재 분석에 따르면 피부에 의해 흡수되는 산소의 양은 아가미를 통해 흡수되는 양보다 훨씬 적다.[19] 피부에 의해 흡수되는 약간의 추가 산소는 아가미로 펌프질하기 전에 피부와 신체에서 정맥혈을 받는 심장에 산소 공급을 보충하는 데 역할을 할 수 있다.[19] 또한 남극빙어는 적혈구를 가진 남극어류에 비해 더 큰 심장 미토콘드리아와 증가된 미토콘드리아 생성을 가지고 있다.[21][22] 이러한 적응은 미토콘드리아 표면적을 증가시키고 세포외 영역과 미토콘드리아 사이의 거리를 줄임으로써 향상된 산소 공급을 용이하게 한다.
4. 진화

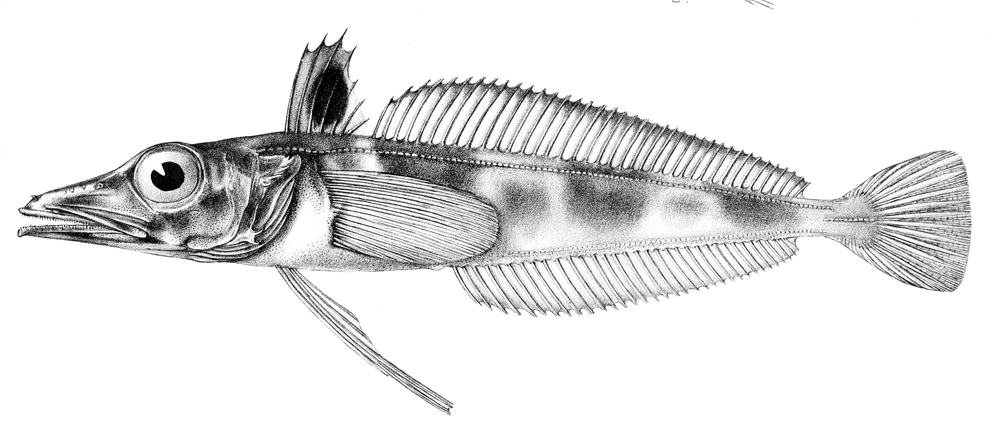
남극빙어의 진화는 남극 전선대와 남극 순환 해류의 형성으로 시작된 것으로 알려져 있다.[23] 남극 순환 해류는 2,500만~2,200만 년 전에 형성되어 남극해를 열적으로 고립시켰다.
제3기 중반, 남극해의 종 감소는 새로운 생태적 지위를 만들었다. 헤모글로빈이 없는 돌연변이체는 경쟁이 적은 환경에서 생존하여 돌연변이를 보상하는 후손을 남길 수 있었다. 이후, 피오르의 주기적인 개방은 미오글로빈 손실을 유발했을 수 있다.[11]
최근 연구에 따르면 헤모글로빈 결핍은 극한 환경에 대한 적응이 아니며, 남극빙어는 혈액 순환에 더 많은 에너지를 소비하여 이를 보완한다.[36]
4. 1. 진화 과정
남극빙어과는 단일계통군으로 간주되며, 느린 저서성 조상으로부터 진화한 것으로 보인다.[2] 차갑고 잘 혼합된 산소가 풍부한 남극해는 헤모글로빈 없이도 낮은 신진대사율을 가진 물고기가 생존할 수 있는 환경을 제공했지만, 효율은 떨어졌다.남극빙어가 언제 진화했는지는 알려져 있지 않다. 두 가지 주요 가설이 제시되었다. 첫 번째는 남극해가 크게 냉각된 후 약 600만 년 전에 나타났다는 것이다. 두 번째는 훨씬 더 오래되어 1,500만~2,000만 년 전이라는 것이다.[2]
남극빙어의 진화는 여전히 논쟁 중이지만, 남극 전선대 (APFZ)와 남극 순환 해류 (ACC)의 형성은 남극어류 진화의 시작을 알리는 것으로 널리 여겨진다.[23] ACC는 시계 방향으로 북동쪽으로 이동하며 너비가 최대 10000km에 달한다. 이 해류는 2,500만~2,200만 년 전에 형성되었으며, 북쪽의 따뜻한 아열대 순환류로부터 분리되어 남극해를 열적으로 고립시켰다.
제3기 중반 동안, 남극해의 종 감소는 광범위한 비어있는 생태적 지위를 식민지화할 기회를 열었다. 헤모글로빈이 없는 돌연변이체가 덜 적합했음에도 불구하고, 경쟁의 부재는 돌연변이체조차 비어있는 서식지를 식민지화하고 돌연변이를 보상하는 후손을 남길 수 있게 했다. 이후, 주기적인 피오르의 개방은 소수의 개체에 의해 식민지화된 서식지를 만들었다. 이러한 조건은 또한 미오글로빈의 손실을 허용했을 수 있다.[11]
헤모글로빈 결핍은 극한 환경에의 적응(산소 용해도가 커 헤모글로빈 의존도가 낮아지고, 적혈구 결핍은 혈액 점성을 낮춤)으로 여겨져 왔다. 하지만, 최신 연구에 따르면 헤모글로빈 결핍은 적응적이지 않다고 여겨진다.[36] 사실, 남극빙어는 혈액 순환에 일반 어류의 2배에 달하는 에너지를 소비하는 등 생리학적으로 헤모글로빈 결핍을 과감하게 보완하고 있다.[36]
남극빙어는 저서 어류를 조상으로 둔다. 차갑고, 영양 풍부하며, 산소가 풍부한 남극해 해수는 비록 효율이 떨어지는 헤모글로빈이 없는 저대사 종이라도 생존할 수 있는 환경을 제공했다.
제3기(6,430만 년 전부터 260만 년 전) 중반에 남극해에서 종이 폭발적으로 증가하며, 다양한 서식지 분화가 발생했다. 경쟁이 적은 남극해에서는 헤모글로빈을 가지지 못한 적응력이 부족한 돌연변이라도, 분화된 환경 속에서 자손을 남기고, 그 돌연변이의 단점을 보완하기 위한 진화를 하는 것이 허용되었다. 또한, 후에 피오르에 몇몇 종이 서식했다. 이러한 환경이 미오글로빈 결핍도 허용한 것으로 생각된다.[36]
4. 2. 헤모글로빈과 미오글로빈 손실
남극빙어과 어류는 헤모글로빈이 없어 혈액이 무색 투명하다.[36][37] 이는 혈액 속 산소 운반 단백질인 헤모글로빈이 없기 때문이다. 남극빙어는 성체가 되어서도 헤모글로빈을 가지지 않는 유일한 척추동물이다.[36][37] 남극빙어에는 헤모글로빈 유전자 흔적이 남아있지만, 헤모글로빈을 생산하지 않는다. 헤모글로빈은 α 서브 유닛과 β 서브 유닛의 두 종류로 구성되는데, 남극빙어과 16종 중 15종에서 β 관련 유전자가 완전히 소실되었고, α 유전자도 부분적으로 결손되었다.[38] 나머지 1종인 ''Neopagetopsis ionah''는 다른 종에 비해 유전자를 많이 남겼지만, 기능하지 않는다.[39]거의 모든 종류에서 적혈구가 없으며, 있더라도 소량의 잔해뿐이다.[40] 산소는 헤모글로빈 없이 혈장에 용해되어 운반된다. 낮은 신진대사와 저온으로 산소가 혈장에 용해되기 쉬운 환경이기에 헤모글로빈 없이도 생존 가능하다.[36] 그러나 산소 운반 능력은 헤모글로빈의 10% 이하이다.[41]
근육 속 산소 저장 단백질인 미오글로빈은 모든 남극빙어과의 골격근에 존재하지 않는다. 다만, 남극빙어과 10종에서는 심근, 특히 심장의 심실에서 미오글로빈이 발견된다.[42] 남극빙어 심실 심근에서의 미오글로빈 유전자 발현은 적어도 4번의 서로 다른 시기에 걸쳐 손실된 것으로 여겨진다.[36][43]
헤모글로빈 결핍은 극한 환경에의 적응(산소 용해도가 커 헤모글로빈 의존도가 낮아지고, 적혈구 결핍은 혈액 점성을 낮춤)으로 여겨져 왔다. 하지만, 최신 연구에 따르면 헤모글로빈 결핍은 적응적이지 않다.[36] 실제로 남극빙어는 혈액 순환에 일반 어류의 2배에 달하는 에너지를 소비하는 등 생리학적으로 헤모글로빈 결핍을 과감하게 보완하고 있다.[36]
최근 연구(2019)는 헤모글로빈 손실이 적응적 가치를 갖는다고 주장한다.[24] 철은 얼음고기가 서식하는 환경에서 제한적인 영양소이다.[25] 얼음고기는 더 이상 헤모글로빈을 합성하지 않음으로써 내인성 철 사용을 최소화하고 있다. ''Champsocephalus gunnari'' 망막 샘플에서 헤모글로빈 알파 3'f 발현이 발견되었는데, 이는 얼음고기 내 헤모글로빈 유전자 조각의 제한적 전사 및 번역이 있음을 보여준다. 이 헤모글로빈 조각은 철 결합 부위를 포함하지 않아, 철 보존을 위한 선택적 작용을 시사한다.
계통 발생 관계는 심장 조직에서 미오글로빈 비발현이 적어도 네 번의 개별적인 시기에 진화했음을 나타낸다.[17] 이는 심장 미오글로빈이 흔적 기관이거나, 빙어에게 유해할 수도 있음을 시사한다. 연구 결과, 빙어 미오글로빈은 다른 분류군보다 기능이 뛰어나거나 덜하지 않다.[11] 이는 미오글로빈이 선택적으로 억제되었을 가능성이 낮다는 것을 의미한다. 심장 미오글로빈이 자연적으로 없는 빙어 종이 심장 미오글로빈을 자연적으로 발현하는 물고기보다 미오글로빈 없이 더 나은 성능을 보였다.[11] 이는 심장 미오글로빈이 없는 물고기가 보상 적응을 겪었음을 시사한다.
남극빙어는 저서 어류를 조상으로 둔다. 차갑고, 영양 풍부하며, 산소가 풍부한 남극해 해수는 효율이 떨어지는 헤모글로빈이 없는 저대사 종이라도 생존할 수 있는 환경을 제공했다.
제3기 중반 남극해에서 종이 폭발적으로 증가하며 다양한 서식지 분화가 발생했다. 경쟁이 적은 남극해에서는 헤모글로빈을 가지지 못한 적응력이 부족한 돌연변이라도 분화된 환경 속에서 자손을 남기고, 그 돌연변이의 단점을 보완하기 위한 진화를 하는 것이 허용되었다. 또한, 후에 피오르에 몇몇 종이 서식했다. 이러한 환경이 미오글로빈 결핍도 허용한 것으로 생각된다.[36]
4. 3. 형질 고정 이유
남극빙어과는 단일계통군으로, 느린 저서성 조상으로부터 진화한 것으로 보인다.[2] 차갑고 산소가 풍부한 남극해는 헤모글로빈 없이도 낮은 신진대사율을 가진 물고기가 생존할 수 있는 환경을 제공했지만, 효율은 떨어졌다.남극빙어가 언제 진화했는지는 정확히 알려져 있지 않지만, 두 가지 주요 가설이 있다. 첫 번째는 남극해가 크게 냉각된 후 약 600만 년 전에 나타났다는 것이고, 두 번째는 이보다 훨씬 오래되어 1,500만~2,000만 년 전이라는 것이다.[2]
남극빙어의 진화는 여전히 논쟁 중이지만, 남극 전선대 (APFZ)와 남극 순환 해류 (ACC)의 형성은 남극 어류 진화의 시작을 알리는 것으로 널리 알려져 있다.[23] 남극 순환 해류는 시계 방향으로 북동쪽으로 이동하며 너비가 최대 10000km에 달한다. 이 해류는 2,500만~2,200만 년 전에 형성되었으며, 북쪽의 따뜻한 아열대 순환류로부터 분리되어 남극해를 열적으로 고립시켰다.
제3기 중반, 남극해의 종 감소는 광범위한 비어있는 생태적 지위를 식민지화할 기회를 열었다. 헤모글로빈이 없는 돌연변이체가 덜 적합했음에도 불구하고, 경쟁의 부재는 이들조차 비어있는 서식지를 식민지화하고 돌연변이를 보상하는 후손을 남길 수 있게 했다. 이후, 주기적인 피오르의 개방은 소수의 개체에 의해 식민지화된 서식지를 만들었다. 이러한 조건은 또한 미오글로빈의 손실을 허용했을 수 있다.[11]
남극해는 지난 1,000만~1,400만 년 동안 매우 춥지만 안정적인 온도로 특징지어져 왔다.[26] 낮은 온도는 높은 수중 산소 함량을 가능하게 하며, 이는 높은 수준의 수직 혼합과 결합되어 남극 해역의 산소 가용성이 유난히 높다는 것을 의미한다. 헤모글로빈과 미오글로빈의 손실은 더 따뜻한 환경에서는 부정적인 결과를 초래할 수 있다.[12] 또한 온도 안정성은 "운이 좋은"데, 온도 변동이 심하면 유해한 돌연변이가 있는 개체를 제거할 가능성이 높은 더 스트레스가 많은 환경이 조성될 것이다.
대부분의 연구는 얼음 물고기에서 헤모글로빈 손실이 무작위적인 진화적 사건으로 인해 발생한 중립적이거나 부적응적인 특징이라고 시사하지만,[27] 일부 연구자들은 헤모글로빈 손실이 얼음 물고기에게 필요한 적응과 관련이 있을 수 있다고 제안하기도 했다.[27] 대부분의 동물은 헤모글로빈 생산에 철이 필요하며, 철은 종종 해양 환경에서 제한적이다.[28] 헤모글로빈 손실을 통해 얼음 물고기는 철 요구량을 최소화할 수 있다. 이러한 최소화는 850만 년 전 북극의 다양성이 극적으로 감소했을 때 얼음 물고기의 생존에 도움이 되었을 수 있다.[27]
최근 연구에 따르면 헤모글로빈과 미오글로빈은 산소 전달 및 사용뿐만 아니라 산화 질소를 분해하는 과정에도 관여한다.[29] 남극빙어가 헤모글로빈과 미오글로빈을 잃었을 때 산소 운반 능력이 감소했을 뿐만 아니라 전체 산화 질소 수치가 상승했다.[11] 산화 질소는 남극빙어에서 아가미 혈관 확장, 심박출량 및 동력 출력을 포함한 다양한 심혈관 과정을 조절하는 역할을 한다.[30] 산화 질소의 존재는 또한 혈관 신생, 미토콘드리아 생성을 증가시키고 근육 비대를 유발할 수 있으며, 이 모든 특징은 남극빙어의 특징이다. 산화 질소 매개 형질 발현과 남극빙어의 특이한 심혈관 특성 사이의 유사성은 이러한 비정상적인 특성이 시간이 지남에 따라 진화했지만, 이 특성의 대부분은 단순히 증가된 산화 질소 수치에 대한 즉각적인 생리적 반응이었으며, 이는 차례로 항상성 진화 과정을 초래했을 수 있음을 시사한다.[11]
헤모글로빈 결핍은 극한 환경에의 적응(산소 용해도가 커 헤모글로빈 의존도가 낮아지고, 적혈구 결핍은 혈액 점성을 낮춤)으로 여겨져 왔지만, 최신 연구에 따르면 헤모글로빈 결핍은 적응적이지 않다고 여겨진다.[36] 남극빙어는 혈액 순환에 일반 어류의 2배에 달하는 에너지를 소비하는 등 생리학적으로 헤모글로빈 결핍을 과감하게 보완하고 있다.[36]
남극빙어는 저서 어류를 조상으로 둔다. 차갑고, 영양 풍부하며, 산소가 풍부한 남극해 해수는 비록 효율이 떨어지는 헤모글로빈이 없는 저대사 종이라도 생존할 수 있는 환경을 제공했다. 제3기 중반에 남극해에서 종이 폭발적으로 증가하며, 다양한 서식지 분화가 발생했다. 경쟁이 적은 남극해에서는 헤모글로빈을 가지지 못한 적응력이 부족한 돌연변이라도, 분화된 환경 속에서 자손을 남기고, 그 돌연변이의 단점을 보완하기 위한 진화를 하는 것이 허용되었다. 또한, 후에 피오르에 몇몇 종이 서식했다. 이러한 환경이 미오글로빈 결핍도 허용한 것으로 생각된다.[36]
참조
[1]
논문
Family-group names of Recent fishes
https://www.research[...]
2014
[2]
논문
Antarctic icefishes (Channichthyidae): a unique family of fishes. A review, Part I
2005
[3]
서적
Antarctic Ecosystems
1990
[4]
논문
Early life history of two Channichthys species from the Kerguelen Islands, Antarctica (Pisces: Notothenioidei: Channichthyidae)
https://www.research[...]
2002
[5]
뉴스
'Major Discovery' Beneath Antarctic Seas: A Giant Icefish Breeding Colony
https://www.nytimes.[...]
2022-01-13
[6]
논문
A vast icefish breeding colony discovered in the Antarctic
[7]
FishBase family
[8]
BioRef
2021-10-12
[9]
논문
The role of notothenioid fish in the food web of the Ross Sea shelf waters: a review
2004
[10]
논문
Fish length-weight relationships in the Weddell Sea and Bransfield Strait
2003
[11]
논문
When Bad Things Happen to Good Fish: The Loss of Hemoglobin and Myoglobin Expression in Antarctic Icefishes
2006-05-15
[12]
논문
Vertebrates without Erythrocytes and Blood Pigment
1954-05-08
[13]
논문
Do the hemoglobinless icefishes have globin genes?
1997
[14]
논문
A genomic fossil reveals key steps in hemoglobin loss by the antarctic icefishes
[15]
논문
The blood cells of the Antarctic icefish Chaenocephalus aceratus Lönnberg: light and electron microscopic observations
1981-07-01
[16]
논문
Oxygen uptake and circulation by a hemoglobinless Antarctic fish (Chaenocephalus aceratus Lonnberg) compared with three red-blooded Antarctic fish
2015-10-15
[17]
논문
Variable expression of myoglobin among the hemoglobinless antarctic icefishes
[18]
논문
Two species of Antarctic icefishes (Genus Champsocephalus) share a common genetic lesion leading to the loss of myoglobin expression
2004
[19]
논문
Gills of Antarctic Fish
1998-01
[20]
논문
Mechanical Performance of the Isolated and Perfused Heart of the Haemoglobinless Antarctic Icefish Chionodraco Hamatus (Lonnberg): Effects of Loading Conditions and Temperature
1991-06-29
[21]
논문
High mitochondrial densities in the hearts of Antarctic icefishes are maintained by an increase in mitochondrial size rather than mitochondrial biogenesis
2008-08-15
[22]
논문
Draft genome assembly and transcriptome data of the icefish Chionodraco myersi reveal the key role of mitochondria for a life without hemoglobin at subzero temperatures
2019-12
[23]
서적
Antarctic Fish Biology: Evolution in a Unique Environment
Academic Press, Inc.
1993
[24]
논문
Vascular Expression of Hemoglobin Alpha in Antarctic Icefish Supports Iron Limitation as Novel Evolutionary Driver
2019-11-12
[25]
논문
Early season depletion of dissolved iron in the Ross Sea polynya: Implications for iron dynamics on the Antarctic continental shelf
2011-12-15
[26]
논문
Cenozoic evolution of Antarctic glaciation, the circus-Antarctic Ocean and their impact on global paleooceanography.
1977
[27]
논문
Vascular Expression of Hemoglobin Alpha in Antarctic Icefish Supports Iron Limitation as Novel Evolutionary Driver
2019-11-12
[28]
논문
Growth Limitation of Marine Fish by Low Iron Availability in the Open Ocean
2019
[29]
논문
Nitric oxide dioxygenase function and mechanism of flavohemoglobin, hemoglobin, myoglobin, and their associated reductases.
2004
[30]
논문
Control of cardiovascular function in the icefish Chionodraco hamatus: involvement of serotonin and nitric oxide.
2003
[31]
FishBase family
[32]
논문
Temperature and evolution: Southern Ocean cooling and the Antarctic marine fauna
1990
[33]
논문
The role of notothenioid fish in the food web of the Ross Sea shelf waters: a review
2015-10-15
[34]
논문
Fish length-weight relationships in the Weddell Sea and Bransfield Strait
2003
[35]
웹사이트
南極海底に「世界最大」の繁殖地 コオリウオの巣、ずらり6千万個
https://www.asahi.co[...]
朝日新聞DIGITAL
2022-02-23
[36]
논문
When Bad Things Happen to Good Fish: The Loss of Hemoglobin and Myoglobin Expression in Antarctic Icefishes
http://jeb.biologist[...]
2012-04-07
[37]
논문
Vertebrates without Erythrocytes and Blood Pigment
https://www.nature.c[...]
2012-04-07
[38]
논문
Do the hemoglobinless icefishes have globin genes?
2015-10-15
[39]
논문
A genomic fossil reveals key steps in hemoglobin loss by the antarctic icefishes
[40]
논문
The blood cells of the Antarctic icefish Chaenocephalus aceratus Lönnberg: light and electron microscopic observations
1981-07-01
[41]
논문
Oxygen uptake and circulation by a hemoglobinless Antarctic fish (Chaenocephalus aceratus Lonnberg compared with three red-blooded Antarctic fish
2015-10-15
[42]
논문
Variable expression of myoglobin among the hemoglobinless antarctic icefishes
[43]
논문
Two species of Antarctic icefishes (Genus Champsocephalus) share a common genetic lesion leading to the loss of myoglobin expression
2004
[44]
논문
Gills of Antarctic Fish
http://www.sciencedi[...]
2012-04-09
[45]
논문
Mechanical Performance of the Isolated and Perfused Heart of the Haemoglobinless Antarctic Icefish Chionodraco Hamatus (Lonnberg): Effects of Loading Conditions and Temperature
http://rstb.royalsoc[...]
2012-05-18
[46]
간행물
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com