미토콘드리아
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
미토콘드리아는 세포 내에서 에너지를 생산하는 세포 소기관으로, 이중막 구조를 가지며 내막과 외막, 막 사이 공간, 크리스타, 기질로 구성된다. 주요 기능은 ATP를 생산하는 것이며, 세포 사멸, 칼슘 저장, 다양한 대사 과정에도 관여한다. 미토콘드리아 DNA는 모계 유전되며, 유전 암호는 세포핵과 다르다. 미토콘드리아는 α 프로테오박테리아에서 기원했으며, 미토콘드리아 기능 부전은 다양한 질병, 특히 신경 질환, 대사 질환, 암, 노화와 관련이 있다.
더 읽어볼만한 페이지
- 미토콘드리아 - 미토콘드리아 내막
미토콘드리아 내막은 크리스타 구조로 표면적을 넓혀 산화적 인산화 효율을 높이고 미토콘드리아 기질을 분리하며 물질대사에 중요한 역할을 하는 다양한 단백질과 수송체를 포함하고 특정 분자에 대해 조절된 투과성을 갖는 미토콘드리아의 안쪽 막이다. - 미토콘드리아 - 고동물
고동물은 미토콘드리아가 없는 원시 진핵생물이 존재했다는 가설에서 파생된 분류군이었으나, 이후 연구에서 아르케조아로 여겨졌던 생물들이 미토콘드리아를 잃어버린 생물로 밝혀지면서 가설과 분류군 모두 쇠퇴하고 폐기되었다. - 세포소기관 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포소기관 - 세포막
세포막은 세포질을 둘러싸고 세포 내외부 환경을 구분하는 선택적 투과성 막으로, 인지질 이중층과 단백질, 탄수화물로 구성되어 물질 수송, 세포 형태 유지, 세포 간 신호 전달, 세포 접합 등의 기능을 수행하며, 유동 모자이크 모델로 설명된다. - 세포 호흡 - 유비퀴논
유비퀴논(CoQ10)은 미토콘드리아 전자 전달계의 주요 구성 요소인 지용성 유기 화합물로, 세포 호흡과 에너지 생산에 필수적이며 항산화 작용을 통해 세포를 보호하고 면역 반응에도 관여한다. - 세포 호흡 - 광합성
광합성은 생물이 빛에너지를 이용하여 이산화탄소와 물로부터 유기물을 합성하고 산소를 방출하는 과정으로, 엽록체 내 틸라코이드 막에서 일어나는 명반응과 스트로마에서 일어나는 암반응으로 구성되며, 환경에 따라 탄소 농축 메커니즘을 통해 효율을 높이기도 하고, 지구 대기의 산소를 생성하는 주요 원천이다.
| 미토콘드리아 | |
|---|---|
| 기본 정보 | |
 | |
| 라틴어 | organella |
| 세포의 일부 | 세포 |
| 복수형 | mitochondria |
| 생물학적 역할 | |
| 주요 기능 | 세포 호흡 |
| 관련 질병 | 신경퇴행성 질환, 심장 질환, 자폐증 |
| 특징 | |
| 형태 | 구형 또는 막대 모양 |
| 크기 | 길이: 0.5 ~ 1.0 μm 직경: 0.5 μm |
| 구조 | |
| 막 구조 | 이중막 (내막, 외막) |
| 주요 구성 요소 | DNA, 리보솜, 단백질, 지질 |
| 진화적 기원 | |
| 기원 | 세포내 공생설에 따른 알파프로테오박테리아 |
| 추가 정보 | |
| 발견 | 1890년 리하르트 알트만 |
| 기능 | ATP 생성 칼슘 이온 조절 세포 사멸 조절 열 발생 |
| 관련 용어 | 미토좀 하이드로제노좀 |
| 예외 | 일부 생물은 미토콘드리아가 없음 (예: Henneguya salminicola) |
2. 구조
전자 현미경 연구를 통해 미토콘드리아는 다양한 형태를 가진다는 것이 밝혀졌지만, 광학 현미경으로는 작은 과립으로만 관찰된다. 세포마다 미토콘드리아 수는 다르지만, 보통 많은 수가 존재한다. 전자 현미경 관찰에 따르면, 미토콘드리아는 바깥쪽 막과 내부로 구성된다. 막은 외막과 내막의 이중막 구조이며, 내막 일부는 안쪽으로 돌출하여 여러 겹으로 접힌 크리스타 구조를 형성한다. 미토콘드리아에는 TCA 회로 및 전자 전달계에 관여하는 모든 효소가 있어, 세포 호흡을 통해 유기물의 화학 에너지를 ATP로 전환하는 중요한 역할을 한다.[279]
2. 막 사이 공간
5. 라멜라 구조
6. 기질
7. 크리스테 구조
8. 내막
11. Mitochondrial DNA
13. 리보솜
14. ATP 합성 효소
15. Porin
]]
미토콘드리아는 세포막과 유사한 이중막 구조로, 단백질을 포함하는 인지질 이중층으로 구성된 내막과 외막을 가진다. 외막과 내막은 서로 다른 기능을 수행하며, 내막 내부는 기질, 내막과 외막 사이는 막간 공간이라고 한다.[280] 미토콘드리아의 길이는 약 2~6 마이크로미터(μm), 직경은 약 0.5μm이다.
미토콘드리아는 다양한 형태를 가질 수 있으며,[20] 인지질 이중층과 단백질로 구성된 외막과 내막을 포함한다.[16] 이중 막 구조로 인해 미토콘드리아는 외막, 막 사이 공간, 내막, 크리스타 공간, 기질의 다섯 부분으로 구성된다. 미토콘드리아는 표면적을 넓혀 ATP 생산을 증가시키는 접힌 구조를 가지고 있다. 외막이 제거된 미토콘드리아는 미토플라스트라고 불린다.
미토콘드리아는 Oxymonad의 ''Monocercomonoides''를 제외한 모든 진핵생물에서 발견된다.[82] 콩 모양으로 묘사되지만, 대부분 세포에서 역동적인 네트워크를 형성하며 분열과 융합을 겪는다. 세포의 모든 미토콘드리아 집단을 콘드리오라고 한다.[83] 미토콘드리아는 세포 유형에 따라 수와 위치가 다르다. 단세포 생물은 단일 미토콘드리아를 가지는 경우가 많지만, 인간 간세포는 세포당 약 1000~2000개의 미토콘드리아를 가지며, 이는 세포 부피의 1/5을 차지한다.[16] 세포의 미토콘드리아 함량은 크기와 막 전위에서 차이를 보이며,[84] 세포 분열 시 불균등한 분할 등으로 인해 ATP 수준 및 세포 과정에서 외적 차이가 발생한다.[85] 미토콘드리아는 근원섬유 사이, 정자 편모 주위에 위치할 수 있다.[16] 세포 골격과 함께 세포 내에서 복잡한 3D 분지 네트워크를 형성하며, 세포 골격과의 연관성은 미토콘드리아 모양을 결정하고 기능에 영향을 미칠 수 있다.[86] 미토콘드리아 네트워크의 구조는 집단에 다양한 물리적, 화학적, 신호 전달상의 이점 또는 불이점을 제공할 수 있다.[87] 세포 내 미토콘드리아는 항상 미세소관을 따라 분포하며, 소포체와도 관련이 있다.[88] 비멘틴 또한 세포 골격과의 연관성에 중요하다.[89]
미토콘드리아 연관 소포체 막(MAM)은 세포 생리 및 항상성에 중요한 역할을 하는 구조적 요소이다. 미토콘드리아와 소포체 사이 계면에서 유래된 막 구조로, 전자 현미경과 형광 현미경으로 관찰되었다.[90] 연구에 따르면 MAM에서 소포체와 미토콘드리아는 10–25 nm 떨어져 있으며 단백질 연결 복합체에 의해 유지된다.[90][91][92]
정제된 MAM은 Ca 신호 전달 관련 채널과 인지질 교환 관여 효소가 풍부하다.[90][92] MAM은 세포 내 지질 저장 및 신호 전달 조절에 중요한 역할을 하며, 미토콘드리아 관련 세포 현상에 영향을 미친다. MAM은 세포자멸사 및 칼슘 신호 전달 메커니즘에 대한 통찰력을 제공하며, 미토콘드리아를 세포 생리에 통합된 요소로 강조한다.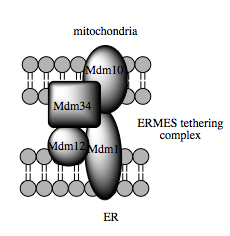
MAM은 ER 및 미토콘드리아 생리 통합을 가능하게 하는 신호 전달, 대사 및 이동 허브이다. 이들 소기관 간 결합은 구조적, 기능적 연결을 포함하며, 세포 생리 및 항상성에 중요하다. MAM은 미토콘드리아를 정적이고 고립된 단위가 아닌, 다양한 세포 과정에 통합된 요소로 제시한다.[107] 최근 뉴런의 미토콘드리아와 MAM이 특수화된 세포 간 통신 부위(체성 접합부)에 고정되어 미세 아교 세포 과정이 뉴런 기능을 보호하며, MAM은 세포 품질 관리 역할을 하는 것으로 추정된다.[108]
1. 내막 2. 외막 3. 크리스테 (판 모양) 4. 기질
미토콘드리아 직경은 0.5 μm 정도이지만, 형태는 생물 종이나 세포 조건에 따라 구형, 원통형, 끈 모양, 그물 모양 등 다양하며, 길이가 10 μm에 달하는 경우도 있다. 1 세포당 수는 1개부터 수천 개까지 다양하며, 분열과 융합을 반복하기도 한다.
미토콘드리아는 외막과 내막의 지질 막을 가지며, 내막 안쪽은 '''기질'''[224], 내막과 외막 사이는 '''막간 공간'''이다. 내막은 기질 쪽으로 함입된 크리스테 구조를 형성한다. "cristae"는 "빗"을 의미한다.[225]
살아있는 세포를 위상차 현미경으로 관찰하면 미토콘드리아를 명확하게 확인할 수 있으며,[225] 세포 내에서 움직이는 모습도 관찰 가능하다.[225]
2. 1. 외막
미토콘드리아 외막은 미토콘드리아 전체를 둘러싸고 있는 막으로, 두께는 60~75 옹스트롬 (Å)이다. 세포막과 비슷하게 단백질과 인지질의 무게 비율이 약 1:1이다. 외막에는 포린이라는 적분 막 단백질이 많이 존재하는데, 이 포린은 분자량 5000 달톤 이하의 분자가 통과할 수 있는 통로(channel)를 형성한다.[281] 이보다 큰 분자는 능동 수송을 통해서만 외막을 통과할 수 있다.[281]주요 수송 단백질은 기공 형성 전압 의존성 음이온 채널(VDAC)이다. VDAC는 세포질과 막 사이 공간 사이의 뉴클레오타이드, 이온, 대사 산물의 주요 수송체이다.[21][22] 이는 그람 음성 세균 외막에서와 유사하게 외막을 가로지르는 베타 배럴로 형성된다.[23]
외막은 또한 지방산 합성 및 신장, 에피네프린 (아드레날린) 산화, 트립토판 분해 등의 기능에 관련된 다양한 효소를 포함한다. 이러한 효소에는 모노아민 산화 효소, 로테논 비의존적 NADH-시토크롬 c 환원 효소, 키누레닌 수산화 효소 및 지방산 Co-A 리가아제가 있다.[25]
미토콘드리아 외막은 소포체 막과 결합하여 MAM (미토콘드리아 연관 ER 막)이라는 구조를 형성할 수 있는데, 이는 ER-미토콘드리아 칼슘 신호 전달에서 중요하며 ER과 미토콘드리아 사이의 지질 이동에 관여한다.[91]
외막의 진화적 기원은 진핵생물의 세포내막계로 여겨지며, 현재에도 소포체 막과 물리적으로 관련되어 있어 칼슘 신호 전달 및 지질 교환을 수행한다.[226]
외막이 파괴되면 막 사이 공간에 존재하는 단백질(시토크롬 c 등)이 세포질로 유출되어 세포의 세포자멸사를 유발할 수 있다.[228]
2. 2. 내막
미토콘드리아 내막은 다음과 같은 세 가지 주요 기능을 수행하는 단백질들을 포함하고 있다.[16]내막은 151개 이상의 서로 다른 폴리펩타이드를 포함하며, 단백질 대 인지질 비율이 매우 높다(무게 기준으로 3:1 이상, 즉 인지질 15개당 단백질 약 1개). 미토콘드리아 전체 단백질의 약 1/5을 차지한다.[26] 또한, 내막에는 특이한 인지질인 카디오리핀이 풍부하다. 이 인지질은 1942년 소 심장에서 처음 발견되었으며, 일반적으로 미토콘드리아 및 박테리아 세포막의 특징이다.[27] 카디오리핀은 두 개의 지방산이 아닌 네 개의 지방산을 포함하며, 내막을 불투과성으로 만드는 데 도움이 될 수 있다.[16] 카디오리핀의 파괴는 신경 질환 및 암을 포함한 여러 임상 질환으로 이어질 수 있다.[28]
외막과 달리 내막에는 포린이 포함되어 있지 않으며, 모든 분자에 대해 투과성이 매우 낮다. 거의 모든 이온과 분자는 기질에 들어가거나 나가기 위해 특수 막 수송체를 필요로 한다. 단백질은 내막 전위 이동 복합체(TIM) 또는 OXA1L을 통해 기질로 운반된다.[24] 또한, 전자 전달계의 효소 작용에 의해 형성된 내막을 가로지르는 막 전위가 존재한다. 내막 미토콘드리아 융합은 내막 단백질 OPA1에 의해 매개된다.[29]
내막은 기질과 막 사이 공간을 분리하며, 미토콘드리아의 기능적 정체성을 담당한다. 산화적 인산화와 관련된 호흡 사슬 복합체 등의 효소군이 내막에 규칙적으로 배열되어 있다. 외막과는 대조적으로, 기본적으로 내막은 불투과성이며, 어떤 물질을 내막을 가로질러 수송하기 위해서는 각 물질에 특이적인 수송체가 필요하다.
호흡 사슬 복합체는 내막을 사이에 두고 기질로부터 양성자를 막 사이 공간으로 펌핑하여 막 사이 공간 쪽의 프로톤 농도를 높여 농도 기울기를 형성한다. 이 농도 기울기는 물질 수송 및 ATP 합성에 관여한다.
또한, 기질로의 단백질 수송 장치나 미토콘드리아의 분열·융합과 관련된 단백질군 등이 존재하며, 미토콘드리아를 구성하는 전체 단백질의 약 20% (150개 이상)가 포함되어 있다. 단백질과 인지질의 중량비는 3:1 정도이다. 내막의 진화적 기원은 공생 세균의 세포막에서 유래되었으며, 내막에 특징적인 인지질 카르디올리핀의 존재가 그 증거로 여겨진다.
일반적으로 내막은 안쪽으로 함입되어 크리스테라고 불리는 구조를 형성한다. 이를 통해 내막의 표면적을 증가시켜 ATP 합성 능력을 높인다. 외막과 내막의 표면적 비는 세포의 ATP 요구량과 상관관계가 있으며, 간에서는 5배 정도, 골격근에서는 더 큰 값이다.
2. 3. 미토콘드리아 기질
기질(matrix)은 미토콘드리아 내막으로 둘러싸인 공간이다. 미토콘드리아 기질에는 미토콘드리아, 리보솜, tRNA, DNA, 게놈 복사본, 그리고 수백 가지의 효소가 존재한다.[283] 효소는 지방산과 피루브산의 산화, TCA 회로를 포함한 반응을 촉매한다.[283]미토콘드리아는 자체적인 유전 물질을 가지며, RNA와 단백질을 스스로 만들 수 있다. 미토콘드리아 DNA는 내막을 구성하는 펩티드(사람에서는 13개) 정보를 저장하고 있다. 이렇게 자체 생성된 펩티드는 숙주 세포의 핵에서 유래한 폴리펩티드와 함께 미토콘드리아 막을 구성한다.
미토콘드리아 기질에는 미토콘드리아 전체 단백질의 약 2/3가 들어있다.[16] 기질은 내막에 있는 ATP 합성 효소를 통해 ATP를 만드는 데 중요한 역할을 한다. 기질에는 수백 가지 효소, 미토콘드리아 리보솜, tRNA, 미토콘드리아 DNA 유전체 사본 등이 고농도로 섞여 있다. 효소의 주요 기능은 피루브산과 지방산의 산화, 시트르산 회로 반응을 일으키는 것이다.[16] DNA 분자는 TFAM를 포함한 단백질에 의해 뉴클레오이드 형태로 포장된다.[32]

미토콘드리아 기질에서 TCA 회로와 β 산화 등을 통해 생성된 NADH는 전자 전달계의 복합체 I에 전자를 전달하여 NAD+를 만든다. TCA 회로 중간 물질인 숙신산(succinate)은 복합체 II(숙신산 탈수소효소)에 전자를 전달하여 푸마르산(fumarate)으로 바뀐다. 복합체 I, III, IV는 전자를 받아 막 사이 공간으로 양성자(H+)를 내보낸다. ATP 합성 효소는 이렇게 만들어진 양성자 농도 기울기를 이용하여 ADP와 무기 인산(Pi)을 결합시켜 ATP를 합성한다.
알데히드 탈수소 효소는 미토콘드리아 기질에 존재하며,[249] 에탄올 섭취 시 생성되는 아세트알데히드를 아세트산으로 분해한다.[249] 동양인 중에는 이 효소의 활성이 유전적으로 낮은 사람이 있어, 아세트알데히드의 독성이 강하게 나타나기 쉽다.[249]
3. 기능
미토콘드리아는 전자 현미경으로 관찰했을 때 다양한 형태를 가지고 있지만, 광학 현미경으로는 작은 과립 형태로만 보인다. 세포마다 미토콘드리아의 수는 다르지만, 일반적으로 많은 수가 존재한다. 전자 현미경으로 보면 미토콘드리아는 외막과 내막의 이중막 구조를 가지고 있으며, 내막은 안쪽으로 돌출되어 여러 겹으로 접힌 '크리스테' 구조를 형성한다. 미토콘드리아에는 TCA 회로와 전자 전달계에 관여하는 모든 효소가 들어 있어, 세포 호흡을 통해 유기물의 화학 에너지를 ATP로 전환하는 중요한 역할을 한다.[279]
미토콘드리아의 기본적인 기능은 유기 물질을 세포가 사용하는 에너지 형태인 ATP로 전환하는 것이다. 그러나 ATP 생산 외에도 미토콘드리아는 다양한 기능을 수행한다.
미토콘드리아의 일부 기능은 특정 세포에서만 수행된다. 예를 들어, 간 세포의 미토콘드리아는 단백질 대사의 부산물인 암모니아를 해독하는 효소를 가지고 있다. 이러한 기능을 조절하는 유전자의 변이는 여러 가지 미토콘드리아 질환을 일으킬 수 있다.
미토콘드리아는 세포 호흡을 통해 ATP를 생성하고 세포 대사를 조절한다.[17] 시트르산 회로와 산화적 인산화는 ATP 생산의 핵심 반응이다.
미토콘드리아 지방산 생합성(mtFASII)은 세포 호흡과 미토콘드리아 생합성에 필수적이다.[45] 또한 리소포스포지질 및 스핑고지질과 같은 생리 활성 지질의 수준에 영향을 미쳐 세포 내 신호 전달에 관여한다.[46]
옥타노일-ACP (C8)는 mtFASII의 최종 생성물로, 리포산 생합성의 시작 기질이다.[47] 리포산은 피루브산 탈수소 효소 복합체, α-케토글루타르산 탈수소 효소 복합체, 분지쇄 α-케토산 탈수소 효소 복합체, 글리신 분해 시스템과 같은 중요한 미토콘드리아 효소 복합체의 조효소이므로 mtFASII는 에너지 대사에 영향을 미친다.[48] mtFASII의 다른 생성물은 미토콘드리아 번역, FeS 클러스터 생합성 및 산화적 인산화 복합체의 조립 조절에 역할을 한다.[47] 또한 mtFASII와 아실화된 ACP는 미토콘드리아에서 아세틸-CoA 소비를 조절한다.[47]
미토콘드리아는 다음과 같은 여러 다른 대사 활동에서 중추적인 역할을 한다.
- 미토콘드리아 활성 산소를 통한 신호 전달[69]
- 막 전위 조절[17]
- 칼슘 신호 전달 (칼슘 유발 세포 자멸사 포함)[70]
- 세포 대사 조절[71]
- 특정 헴 합성 반응[72] (참고: ''포르피린'')
- 스테로이드 합성[73]
- 호르몬 신호 전달[74] – 미토콘드리아는 호르몬에 민감하고 반응하며, 부분적으로 미토콘드리아 에스트로겐 수용체(mtER)의 작용에 의해 영향을 받는다. 이러한 수용체는 뇌[75]와 심장[76]을 포함한 다양한 조직과 세포 유형에서 발견되었다.
- 면역 세포의 발달 및 기능[77]
- 신경 미토콘드리아는 특수화된 체세포 연결을 통해 미세 아교 세포에 대한 신경 상태를 보고함으로써 세포 품질 관리에 기여한다.[108]
- 발달 중인 뉴런의 미토콘드리아는 미세 아교 세포로의 세포 간 신호 전달에 기여하며, 이러한 소통은 뇌 발달의 적절한 조절에 필수적이다.[78]
미토콘드리아 단백질(미토콘드리아 DNA에서 전사된 단백질)은 조직과 종에 따라 다르다. 사람의 경우, 심장 미토콘드리아에서 615가지의 서로 다른 유형의 단백질이 확인되었으며,[79] 쥐의 경우 940개의 단백질이 보고되었다.[80] 미토콘드리아 프로테옴은 동적으로 조절되는 것으로 생각된다.[81]
3. 1. 에너지 전환
미토콘드리아의 주 기능은 ATP 생산이며, 이는 해당과정의 주 생산물인 피루브산과 NADH 대사, 또는 지방산의 베타 산화를 통해 이루어진다. 세포 유형이나 산소 유무에 따라 ATP 생성에는 두 가지 방법이 있다.해당과정에서 생성된 피루브산은 능동 수송을 통해 미토콘드리아 내막을 거쳐 기질로 들어간다. 기질에서 조효소 A와 결합해 아세틸-CoA를 생성하고, 이는 TCA 회로(구연산 회로 또는 크레브스 회로)로 들어간다. 1개의 피루브산은 3개의 NADH와 1개의 FADH2를 생성하여 전자 전달계에 관여한다. 숙신산 탈수소 효소를 제외한 TCA 회로에 관여하는 모든 효소는 미토콘드리아 기질에 존재한다. 지방산은 베타 산화를 통해 에너지를 획득할 수 있다.
TCA 회로에서 생성된 NADH와 FADH는 미토콘드리아 기질에서 내막의 전자 전달계 단백질(ComplexⅠ~Ⅳ)로 전자를 전달한다. NADH는 complexⅠ에, FADH는 ComplexⅡ에 전자를 전달한다. 전자는 전자 친화력이 높은 구조로 전달되며, 이때 손실된 에너지로 전자 전달계 복합체들은 수소 이온을 기질에서 막간 공간으로 수송한다. Complex Ⅰ, Ⅲ, Ⅳ는 양성자 (H)를 막 사이 공간으로 펌핑하여 전기화학적 기울기를 형성한다.
Coenzyme-Q는 ComplexⅠ과 Ⅲ, ComplexⅡ와 Ⅲ 사이에서 전자를 전달한다. Complex Ⅲ에서 Cytochrome-C로 전달된 전자는 Complex Ⅳ로 전달된다. Complex Ⅳ는 시토크롬 c에서 받은 전자를 산소와 수소 이온에 전달하여 물(HO)을 만든다.
막간 공간과 기질의 수소 이온 농도 기울기로 인해 ATP 합성 효소가 막간 공간의 수소 이온을 기질로 통과시키면서 ADP와 P를 ATP로 합성한다.

미토콘드리아는 세포 호흡을 통해 ATP를 생성하고 세포 대사를 조절한다.[17] 시트르산 회로와 산화적 인산화는 ATP 생산의 핵심 반응이다. 미토콘드리아는 ATP 생산 외에도 다양한 기능을 수행한다.
유산소 호흡은 산소 존재에 의존하며, 혐기성 발효보다 ATP 생산 효율이 약 13배 높다.[33] 식물 미토콘드리아는 아질산염을 사용하여 산소 없이 ATP를 생산할 수 있다.[34] ATP는 특정 단백질을 통해 내막을, 포린을 통해 외막을 통과한다.[35]
특정 조건에서 양성자는 ATP 합성 대신 미토콘드리아 탈공역을 통해 기질로 다시 들어가 열을 발생시킨다.[17] 이 과정은 서모제닌(UCP1)이라는 양성자 채널에 의해 매개되며, 갈색 지방 조직에서 비-떨림성 열 발생에 관여한다.[44]
미토콘드리아 지방산 생합성(mtFASII)은 세포 호흡과 미토콘드리아 생합성에 필수적이다.[45] 리소포스포지질 및 스핑고지질과 같은 생리 활성 지질 수준에 영향을 미쳐 세포 내 신호 전달에 관여한다.[46]
옥타노일-ACP (C8)는 mtFASII의 최종 생성물로, 리포산 생합성의 시작 기질이다.[47] 리포산은 피루브산 탈수소 효소 복합체, α-케토글루타르산 탈수소 효소 복합체, 분지쇄 α-케토산 탈수소 효소 복합체, 글리신 분해 시스템과 같은 미토콘드리아 효소 복합체의 조효소이므로 mtFASII는 에너지 대사에 영향을 미친다.[48]
mtFASII와 아실화된 ACP는 미토콘드리아에서 아세틸-CoA 소비를 조절한다.[47]
내막은 기질과 막 사이 공간을 분리하며, 산화적 인산화 관련 효소군이 규칙적으로 배열되어 있다. 내막은 불투과성이며, 물질 수송을 위해 특정 수송체가 필요하다. 호흡 사슬 복합체는 기질에서 막 사이 공간으로 프로톤을 펌핑하여 농도 구배를 형성하고, 이는 물질 수송 및 ATP 합성에 관여한다.
내막에는 단백질 수송 장치, 미토콘드리아 분열·융합 관련 단백질군 등이 존재하며, 전체 단백질의 약 20%를 차지한다. 단백질과 인지질의 중량비는 3:1 정도이다. 내막은 공생 세균의 세포막에서 유래했으며, 카르디올리핀이 그 증거로 여겨진다.
내막은 크리스테라는 구조를 형성하여 표면적을 증가시키고 ATP 합성 능력을 높인다. 외막과 내막의 표면적 비는 세포의 ATP 요구량과 상관관계가 있다. 다세포 동물이나 육상 식물에서는 판상 크리스테가 일반적이지만, 관상 크리스테도 흔하다. 조직에 따라 크리스테 형태가 다를 수 있다.
기질(matrix)은 내막으로 둘러싸인 공간으로, TCA 회로나 β 산화 등 미토콘드리아 대사 기능 관련 효소군이 존재한다. mtDNA, 리보솜, tRNA, 전사 인자, 번역 인자 등이 존재하며, 단백질 농도가 매우 높다.
미토콘드리아는 해당 작용과 TCA 회로에서 생성된 산물을 이용하여 전자 전달계에 전자를 전달하고, 프로톤 농도 기울기로 ATP 합성 효소를 구동하여 ADP를 산화적 인산화로 ATP로 변환한다.[234]
미토콘드리아는 ATP, GTP 합성 외에도 스테로이드, 헴 합성, 칼슘, 철 농도 조절, 세포 주기, 세포 자멸사 조절 등 다양한 기능을 수행한다. 이러한 기능은 특정 세포에서만 작동하며, 관련 유전자 변이는 미토콘드리아 질환을 유발할 수 있다.
3. 1. 1. 젖산 대사
해당과정에서 생성된 피루브산은 주로 속근 섬유에서 젖산 탈수소 효소에 의해 젖산으로 변환된다. 속근 섬유는 미토콘드리아가 적고 글리코겐이 많아 혐기적 당 분해를 통해 빠르게 ATP를 생성할 수 있지만, 장시간 운동에는 불리하다. 젖산성 역치보다 높은 운동 강도에서는 속근 섬유가 많이 사용된다.[238]반면, 지근 섬유나 심근은 미오글로빈이 많아 산소를 이용하기 쉬운 환경을 갖추고 있다. 붉은 근육에서는 해당과정의 산물인 피루브산을 미토콘드리아의 TCA 회로로 보내고, NADH도 미토콘드리아로 전달하여 ATP를 합성한다. 이 방식은 젖산 축적 없이 낮은 운동 강도에서 주로 작용한다.
속근 섬유에서 발생한 젖산은 혈액을 통해 간으로 운반되어 코리 회로에서 ATP를 소비하여 포도당 재생에 사용된다. 젖산 탈수소 효소는 젖산을 피루브산으로 되돌리는 역반응도 촉매할 수 있다. 지근 섬유나 심근에서는 외부에서 받아들인 젖산을 피루브산으로 되돌려 미토콘드리아의 TCA 회로에 투입하기도 한다.
젖산 대사에는 세포막을 통한 젖산 수송이 필요하며, 이는 단일 카르복실산 수송체(Monocarboxylate Transporter|모노카복실레이트 수송체영어 (MCT))에 의해 이루어진다.[250]
3. 2. 열 생산
특정 조건에서 양성자는 ATP 합성 과정에 기여하지 않고 미토콘드리아 기질로 다시 들어갈 수 있다. 이 과정을 ''양성자 누출'' 또는 미토콘드리아 탈공역이라고 하며, 양성자가 기질로 촉진 확산되기 때문에 발생한다. 이 과정은 양성자 전기화학 기울기의 억제되지 않은 잠재 에너지를 열로 방출하는 결과를 낳는다.[17] 이 과정은 서모제닌 또는 UCP1이라고 불리는 양성자 채널에 의해 매개된다.[44] 서모제닌은 주로 갈색 지방 조직 또는 갈색 지방에서 발견되며, 비-떨림성 열 발생을 담당한다. 갈색 지방 조직은 포유류에서 발견되며, 초기 생애와 동면하는 동물에서 가장 높은 수준을 보인다. 인간의 경우, 갈색 지방 조직은 출생 시 존재하며 나이가 들면서 감소한다.[44]막 사이 공간의 프로톤은 ATP 합성에 관여하지 않고 촉진 확산에 의해 매트릭스로 되돌아가는 경우가 있는데, 이를 "프로톤 누출" 또는 "미토콘드리아 탈공역"이라고 한다. 이로 인해 축적되었던 전기화학적 포텐셜은 열로서 방출된다. 탈공역 단백질이 이 과정을 매개하며, 근육의 떨림을 동반하지 않는 열 생산에 관여한다.[251] 서모제닌은 어린 개체나 동면 중인 포유류에서 발견되는 갈색 지방 조직의 미토콘드리아에 많이 존재한다.
3. 3. 세포 사멸 (Apoptosis)
프로그램 세포 죽음(PCD)은 기관 발달 및 세포 항상성에 매우 중요하며, 악성 형질 전환을 예방하고 면역에 근본적인 역할을 한다.[65]미토콘드리아는 세포자멸사의 내인성 경로에서 핵심적인 역할을 하며,[66] 선천 면역의 신호 허브로도 확인되었다.[67] 미토콘드리아 구성 요소가 세포질에 노출되면 세포자멸사, 자가포식 또는 전염증 유전자의 유도를 유발할 수 있다.[68][67]
미토콘드리아는 시토크롬 'c'를 방출하여 아폽토솜 형성을 유도하고 세포자멸사에 기여한다. 또한 다양한 손상 관련 분자 패턴(DAMP)의 공급원이며, 이는 병원체 관련 분자 패턴(PAMP)에 반응하는 패턴 인식 수용체(PRR)에 의해 인식된다. 예를 들어, 미토콘드리아 mtDNA는 CpG 부위 메틸화가 부족하여 세균 DNA와 유사하며, Toll-like receptor 9 및 cGAS에 의해 감지될 수 있다. 이중 가닥 RNA(dsRNA)는 양방향 미토콘드리아 전사로 생성되며, RIG-I 유사 수용체를 통해 바이러스 감지 경로를 활성화할 수 있다. 미토콘드리아 단백질의 ''N''-포름화는 포르밀 펩타이드 수용체에 의해 인식될 수 있다.[68]
세포에 DNA 손상 등의 스트레스가 발생하면 아폽토시스 유도 분자 p53나 아폽토시스를 조절하는 Bcl-2 패밀리 단백질을 통해 미토콘드리아의 막전위가 변화되고 외막의 전위 의존성 음이온 채널이 닫힌다.[252] 그러면 미토콘드리아 기능이 저하되고, 시토크롬 c가 유출되어 아폽토솜이 형성된다. 활성화된 카스파제-9는 하류 이펙터를 활성화하여 DNA를 절단하고 세포 자살을 유도한다.
3. 4. 칼슘 저장
세포 내 자유 칼슘 농도는 다양한 반응을 조절하고 세포 내 신호 전달에 중요하며, 미토콘드리아는 일시적으로 칼슘을 저장하여 세포의 칼슘 항상성에 기여한다.[49][50] 미토콘드리아는 칼슘을 빠르게 흡수하여 나중에 방출함으로써 칼슘에 대한 "세포질 완충제" 역할을 수행한다.[73][51][52]
소포체는 칼슘의 가장 중요한 저장 부위이며,[99] 미토콘드리아와 소포체 사이에는 칼슘과 관련하여 상당한 상호 작용이 존재한다.[53] 칼슘은 기질로 내막 미토콘드리아 칼슘 단일 수송체를 통해 흡수되며, [54] 이는 주로 미토콘드리아의 막 전위에 의해 구동된다.[50] 저장된 칼슘은 나트륨-칼슘 교환 단백질이나 "칼슘 유도 칼슘 방출" 경로를 통해 다시 세포 내부로 방출된다.[54]
이러한 칼슘의 흡수와 방출 과정은 막 전위의 큰 변화와 함께 칼슘 스파이크 또는 칼슘 파동을 일으켜, 신경 세포에서 시냅스 소포의 신경 전달 물질 방출 및 내분비 세포에서 호르몬 방출과 같은 과정을 조율하는 2차 전달자 단백질을 활성화한다.[55]
미토콘드리아 기질로의 Ca2+ 유입은 산화 스트레스 감소를 촉진하여 호흡 생체 에너지학을 조절하는 메커니즘으로 작용한다. 세포질 및 미토콘드리아 칼슘의 동시 증가는 신경 활동을 미토콘드리아 에너지 대사와 동기화시킨다. 미토콘드리아 기질 칼슘 수치는 수십 마이크로 몰 수준에 도달하여 크렙스 회로의 주요 조절 효소인 이소시트르산 탈수소 효소를 활성화한다.[57]
미토콘드리아의 칼슘 신호 전달에서 소포체의 역할은 미토콘드리아의 역할이 널리 받아들여지기 전에 인정되었다. 미토콘드리아 외막에 위치한 Ca2+ 채널의 낮은 친화도가 이 소기관의 세포 내 Ca2+ 흐름 변화에 대한 반응성과 모순되는 것처럼 보였으나,[90][99] 미토콘드리아와 소포체 사이의 긴밀한 물리적 연관성은 접촉 지점에서 Ca2+ 마이크로 도메인을 생성하여 소포체에서 미토콘드리아로 효율적인 Ca2+ 전달을 촉진한다.[90]
칼슘 전달은 소포체 막 Ca2+ 채널인 IP3R의 자발적인 클러스터링 및 활성화에 의해 생성된 "Ca2+ 펌프"에 반응하여 발생하며,[90][91] 이 펌프의 강도는 소포체에 의한 Ca2+ 재흡수에 의해 조절된다. 미토콘드리아는 세포질로 방출된 자유 이온을 유입하는 싱크 역할을 하여 Ca2+ 펌프를 완충한다.[90][100][101]
Ca2+ 터널링은 낮은 친화성 Ca2+ 수용체인 VDAC1을 통해 발생하며, 이는 소포체 막의 IP3R 클러스터에 물리적으로 테더되어 미토콘드리아와 소포체 접촉 부위(MAM)에서 농축된다.[90][91][102] 미토콘드리아가 Ca2+ 싱크 역할을 하는 능력은 산화적 인산화 동안 생성된 전기화학적 기울기의 결과이다.[102] 정상적인 칼슘 유입은 일시적인 탈분극을 유발하며, 이는 양성자를 펌핑하여 수정된다.
칼슘의 전송은 양방향으로, 소포체에 존재하는 Ca2+ 펌프 SERCA와 채널 IP3R의 특성은 미토콘드리아와 소포체 접촉 부위(MAM) 기능에 의해 조정된 피드백 조절을 촉진한다. MAM에 의한 Ca2+ 제거는 Ca2+가 IP3R 활성을 변화시키기 때문에 Ca2+ 신호 전달의 시공간 패턴을 허용한다.[90] SERCA 역시 미토콘드리아의 피드백에 영향을 받으며, MAM에 의한 Ca2+ 흡수는 ATP 생산을 자극하여 SERCA가 MAM에서 지속적인 Ca2+ 유출을 위해 소포체를 Ca2+로 다시 채울 수 있도록 에너지를 제공한다.[100][102]
미토콘드리아와 소포체 접촉 부위(MAM)에서 소포체의 Ca2+ 방출을 조절하는 것은 중요하며, Ca2+ 흡수의 특정 창(window)만이 미토콘드리아와 세포를 항상성 상태로 유지한다. 시트르산 회로를 통해 유동에 중요한 탈수소 효소를 활성화하여 신진대사를 자극하기 위해서는 충분한 세포내 Ca2+ 신호 전달이 필요하다.[103][104] 그러나 미토콘드리아 내 Ca2+ 신호 전달이 특정 임계값을 초과하면 미토콘드리아 막 전위를 붕괴시켜 세포 사멸의 내인성 경로를 자극한다.[90]
4. 미토콘드리아 게놈
미토콘드리아는 세포의 에너지 요구에 따라 DNA를 복제하고 분열한다. 세포 에너지 요구량이 높으면 미토콘드리아는 늘어나 분열하고, 낮으면 파괴되거나 비활성화된다. 세포 분열 시 미토콘드리아 분배는 불균등하게 일어날 수 있다. 미토콘드리아는 세균처럼 이분법으로 복제하지만, 서로 융합하기도 한다. 때로는 단백질과 폴리리보솜이 많은 곳에서 새로운 미토콘드리아가 생성되기도 한다.
미토콘드리아 지방산 생합성(mtFASII)은 세포 호흡과 미토콘드리아 생합성에 필수적이다.[45] 또한 세포 내 신호 전달에 영향을 미쳐 생리 활성 지질 수준에 영향을 미치는 매개체 역할을 한다.[46] 옥타노일-ACP (C8)는 mtFASII의 가장 중요한 최종 생성물로, 리포산 생합성의 시작 기질을 형성한다.[47] 리포산은 주요 미토콘드리아 효소 복합체의 조효소이므로 mtFASII는 에너지 대사에 영향을 미친다.[48] mtFASII의 다른 생성물은 미토콘드리아 번역, FeS 클러스터 생합성 및 산화적 인산화 복합체의 조립 조절에 역할을 한다.[47] 또한 mtFASII와 아실화된 ACP의 도움으로 아세틸-CoA는 미토콘드리아에서의 소비를 조절한다.[47]
미토콘드리아 안에는 세포핵과는 별도로 고유한 DNA, 즉 미토콘드리아 DNA(mtDNA)가 존재한다. mtDNA는 세포핵과는 다른 고유한 유전 정보를 가지고 있으며, DNA 분자 크기, 형태, 유전자 수와 종류는 생물 종에 따라 매우 다양하다. 일반적으로 mtDNA는 GC 함량이 낮고(20-40%), 염기쌍이 수십 kb 정도이다. mtDNA에는 전자 전달계 관련 단백질, 리보솜 RNA, tRNA 등 수십 종류의 유전자가 암호화되어 있다.
사람을 포함한 척추동물의 mtDNA는 진핵생물 중에서는 상당히 특수한 성질을 많이 가지고 있어 연구가 많이 진행되었다. mtDNA와 이를 기반으로 합성되는 산물의 일부는 미토콘드리아뿐만 아니라 세포 표면에도 존재하며, mtDNA에 돌연변이가 발생한 경우에는 자연 면역계가 특이적으로 세포 자체를 파괴하여 제거한다.[260] mtDNA에 돌연변이가 발생하면 미토콘드리아 질환을 일으킬 가능성도 있다.
가장 작은 mtDNA를 가진 생물은 아피콤플렉스문 원생생물로, 크기가 6 kb에 불과한 선형 게놈이다. 전자 전달계 관련 3개의 단백질 유전자와 단편화된 리보솜 RNA 유전자군만 존재한다. 반대로 가장 큰 mtDNA는 머스크멜론이 가진 2400 kb의 거대한 게놈이다. 유전자 수는 비교적 많지만 100개 미만이며, 다량의 반복 서열이나 그룹 2 인트론 등의 비유전자 영역이 대부분을 차지한다.
사람을 포함한 다세포 동물의 mtDNA는 길이 16 kb 전후의 단일 환상 DNA로 구성되어 있으며, 37개의 유전자(호흡 사슬 복합체와 ATP 합성 효소의 서브 유닛 13개, tRNA 22개, rRNA 2개)를 포함한다. 유전자 지도 등에서 mtDNA가 환상으로 표현되는 경우가 많지만, 물리적으로 환상의 mtDNA를 가진 생물은 극히 일부이다. 많은 생물에서는 mtDNA의 대부분이 기본 단위가 여러 번 반복되는 선형 반복 구조를 가지고 있다. 소수이지만 항상 선형 mtDNA를 가진 생물도 존재한다.
미토콘드리아 게놈은 알파프로테오박테리아로부터 물려받았으며, 그 유전자 발현은 세균과 공통된 특징을 가지고 있어 진핵생물의 세포핵 DNA와는 다르다. 예를 들어, 여러 유전자가 함께 전사되고, 그것이 유전자별로 절단되어 폴리아데닐화되어 성숙 mRNA가 되는 점, 번역 시작에 포르밀화 메티오닌이 이용되는 점, 세포핵에 존재하는 스플라이소솜형 인트론이 존재하지 않는 점 등이 있다.
미토콘드리아 기능에 관련된 모든 유전자가 mtDNA에 존재하는 것은 아니다. 미토콘드리아 게놈은 세균 게놈에 비해 유전자 수가 극단적으로 감소했다. 미토콘드리아가 필요로 하는 대다수 유전자는 세포핵에 암호화되어 있으며, 세포질에서 생합성된 유전자 산물이 미토콘드리아로 수송된다. 이는 진화 과정에서 유전자가 세포핵으로 이동했기 때문으로 추정된다. 마이토솜과 같이 모든 DNA를 완전히 잃은 미토콘드리아도 존재한다. 원생생물 레크리노모나스(Reclinomonas americana)는 다른 생물에서는 세포핵에서 수송되는 단백질 유전자가 mtDNA상에 존재하며, 비교적 원시적인 미토콘드리아 게놈을 보유하고 있다고 여겨진다. 미토콘드리아에는 호흡 기능에 관여하는 소수성 단백질이 존재하며, 소수성이기 때문에 수송이 어려워 미토콘드리아 내부에서 만들어야 하므로 미토콘드리아에 유전자가 남아 있는 이유 중 하나로 여겨진다. 하나의 미토콘드리아에는 2~10개의 DNA 분자가 존재한다.[264] 이들 모두가 완전히 같은 정보를 갖는 것은 아니며, 복수의 이질적인 DNA 분자를 포함하고 있는 것으로 확인되었다.[230]
4. 1. mtDNA의 유전
미토콘드리아 DNA의 모계 유전(maternal inheritance)은 동물을 포함한 대부분의 진핵생물에서 볼 수 있다. 드물게 부계 유전을 하는 경우도 있는데 침엽수(소나무와 주목 제외)에서 흔하게 나타나며[284], 인간의 경우 아주 낮은 확률로 나타난다고 한다[285].단일 계통 유전은 서로 다른 미토콘드리아 계통 간의 유전자 재조합 가능성이 낮고 미토콘드리아 DNA가 대체로 클론임을 의미한다. 만약 유전자 재조합이 없다면 모든 미토콘드리아 DNA는 일배체형(haplotype, 홑가닥)으로서 동일하게 유전되기 때문에 집단의 진화적 역사를 연구하는 데 유용하게 사용될 수 있다. 하지만 종종 인간의 미토콘드리아 DNA 상호간에는 재조합이 일어나기도 한다.
미토콘드리아는 미토콘드리아가 유래되었을 것으로 추정되는 진정세균 무리보다 무척 적은 규모의 게놈을 가지고 있다. 일부는 소실된 것으로 보이지만 상당수가 세포핵으로 이전한 것으로 보인다. 이러한 현상은 진화적으로 볼 때 비교적 일반적으로 보인다.
미토콘드리아의 단일 계통 유전은 일부 효모종 미토콘드리아의 Petite 돌연변이(자신의 DNA 일부 또는 모두를 잃어 산화적 인산화를 못하는 비정상적인 미토콘드리아 돌연변이. petite와 정상적인 효모 사이의 융합이 일어나게 되면 딸세포의 일정 비율은 petite가 됨.)에서 볼 수 있듯이 유전자 간 분쟁(intragenomic conflict)을 야기할 것으로 보인다. 이러한 세포 소기관의 분쟁은 남성과 여성의 분화로 해결할 수 있으며 유전자 간 분쟁은 성의 분화가 일어난 원인 중 하나로 생각된다.

미토콘드리아는 자체 게놈을 가지고 있다. 인간 미토콘드리아 게놈은 약 16 킬로베이스의 원형 이중 가닥 DNA 분자이다.[133] 이 유전자는 37개의 유전자를 암호화한다. 즉, 호흡 복합체 I, III, IV 및 V의 단백질 소단위체에 대한 13개, 미토콘드리아 tRNA에 대한 22개(20개의 표준 아미노산과 류신 및 세린에 대한 추가 유전자), rRNA에 대한 2개(12S 및 16S rRNA)이다.[133] 하나의 미토콘드리아는 DNA 사본을 2개에서 10개까지 포함할 수 있다.[134]
원핵생물과 마찬가지로, 부호화 DNA의 비율이 매우 높고 반복 서열은 없다. 미토콘드리아 유전자는 다유전자 전사체로 전사되며, 분해 및 폴리아데닐화되어 성숙한 mRNA를 생성한다. 미토콘드리아 기능에 필요한 대부분의 단백질은 세포핵의 유전자에 의해 암호화되며, 해당 단백질은 미토콘드리아로 수입된다.[136] 핵과 미토콘드리아 게놈에 의해 암호화된 유전자의 정확한 수는 종에 따라 다르다. 대부분의 미토콘드리아 게놈은 원형이다.[137] 일반적으로, 인간 미토콘드리아 게놈의 경우와 같이 미토콘드리아 DNA에는 인트론이 없다.[136] 그러나 효모[139] 및 ''Dictyostelium discoideum''을 포함한 원생생물[140]과 같은 일부 진핵생물 미토콘드리아 DNA에서 인트론이 관찰되었다.[138] 단백질 부호화 영역 사이에는 tRNA가 존재한다. 미토콘드리아 tRNA 유전자는 핵 tRNA와 다른 서열을 가지고 있지만, 높은 서열 유사성을 가진 미토콘드리아 tRNA와 유사한 것이 핵 염색체에서 발견되었다.[142]
미토콘드리아 DNA에서 유전자 재조합이 거의 일어나지 않기 때문에, 이는 집단 유전학 및 진화 생물학 연구에 유용한 정보원이 된다.[144] 모든 미토콘드리아 DNA가 단일 단위, 즉 단일배형으로 유전되기 때문에, 서로 다른 개인의 미토콘드리아 DNA 간의 관계는 유전자 나무로 표현될 수 있다. 이러한 유전자 나무의 패턴은 집단의 진화 역사를 추론하는 데 사용될 수 있다. 이에 대한 고전적인 예는 인류 진화 유전학에서 찾아볼 수 있는데, 여기에서 분자 시계는 미토콘드리아 이브의 비교적 최근 시기를 제공하는 데 사용될 수 있다.[145][146] 이는 종종 최근의 현대 인류의 아프리카 기원 확장을 강력하게 뒷받침하는 것으로 해석된다.[147] 또 다른 인간 관련 예로는 네안데르탈인 뼈에서 추출한 미토콘드리아 DNA의 염기 서열 분석이 있다. 네안데르탈인과 현생 인류의 미토콘드리아 DNA 염기 서열 간의 상대적으로 큰 진화적 거리는 네안데르탈인과 현생 인류 간의 교배가 없었다는 증거로 해석되어 왔다.[148]
하지만, 미토콘드리아 DNA는 집단 내 여성의 역사만을 반영한다. 이는 재조합되지 않는 Y 염색체 부위와 같은 부계 유전 염기 서열을 사용하여 부분적으로 극복할 수 있다.[147]
최근의 분자 시계 측정 결과[149] 인간과 유인원의 가장 최근 공통 조상 시점까지 거슬러 올라가, 7,884년에 1번의 돌연변이가 발생하는 값이 보고되었으며, 이는 상염색체 DNA의 돌연변이율 추정치(세대당 염기당 10-8)와 일치한다.[150]
개인의 미토콘드리아 유전자는 드문 경우를 제외하고는 모계로부터만 상속된다.[160] 인간의 경우, 난자가 정자에 의해 수정될 때 미토콘드리아와 그에 따른 미토콘드리아 DNA는 대개 난자에서만 유래한다. 정자의 미토콘드리아는 난자에 들어가지만, 배아에 유전 정보를 제공하지는 않는다.[161] 대신 부계 미토콘드리아는 유비퀴틴으로 표시되어 배아 내부에서 파괴되도록 선택된다.[162] 난자는 비교적 소수의 미토콘드리아를 포함하지만, 이 미토콘드리아는 분열하여 성체 유기체의 세포를 채운다.
단일 부모 유전은 미토콘드리아의 서로 다른 계통 간의 유전자 재조합 기회를 거의 제공하지 않지만, 단일 미토콘드리아는 2~10개의 DNA 사본을 포함할 수 있다.[134] 재조합이 발생하는 경우 다양성을 유지하기보다는 유전적 완전성을 유지한다. 그러나 미토콘드리아 DNA에서 재조합의 증거를 보여주는 연구가 있다. 재조합에 필요한 효소가 포유류 세포에 존재한다는 것은 분명하다.[168] 또한 동물 미토콘드리아가 재조합을 겪을 수 있음을 시사하는 증거가 있다.[169] 인간의 경우 데이터가 더 논쟁적이지만, 재조합의 간접 증거가 존재한다.[170][171]
단일 부모 유전을 겪고 재조합이 거의 또는 전혀 없는 개체는 기능이 손실될 때까지 유해한 돌연변이가 축적되는 뮬러의 래칫의 영향을 받을 것으로 예상할 수 있다. 미토콘드리아의 동물 개체군은 mtDNA 병목 현상이라고 하는 발달 과정을 통해 이러한 축적을 피한다. 병목 현상은 세포 내 확률적 과정을 이용하여 유기체가 발달함에 따라 돌연변이 부하의 세포 간 변동성을 증가시킨다.
4. 2. 유전 암호
미토콘드리아는 자체 게놈을 가지고 있는데, 인간 미토콘드리아 게놈은 약 16 킬로베이스의 원형 이중 가닥 DNA 분자이다.[133] 이 유전자는 37개의 유전자를 암호화한다. 즉, 호흡 복합체 I, III, IV 및 V의 단백질 소단위체에 대한 13개, 미토콘드리아 tRNA에 대한 22개(20개의 표준 아미노산과 류신 및 세린에 대한 추가 유전자), rRNA에 대한 2개(12S 및 16S rRNA)이다.[133]미토콘드리아의 유전 암호표는 세포핵이나 일반적인 원핵생물에서 이용되는 보편 암호표와 비교하여 약간의 차이를 보인다. 예를 들어, 세포핵에서는 종결 코돈이어야 할 UGA가, 미토콘드리아에서는 트립토판을 코딩하는 경우가 많다.
| 생물 | 코돈 | 표준 | 미토콘드리아 |
|---|---|---|---|
| 포유류 | AGA, AGG | 아르기닌 | 종결 코돈 |
| 무척추동물 | AGA, AGG | 아르기닌 | 세린 |
| 균류 | CUA | 류신 | 트레오닌 |
| 위에 언급된 모든 것 | AUA | 이소류신 | 메티오닌 |
| UGA | 종결 코돈 | 트립토판 |
표준 유전 암호의 약간의 변형은 1979년에 인간 미토콘드리아 유전학을 연구하던 과학자들이 대체 암호를 사용한다는 것을 밝혀내면서 발견되었다.[152] 그러나 대부분의 식물을 포함한 다른 많은 진핵생물의 미토콘드리아는 표준 암호를 사용한다.[153]
미토콘드리아에서는 종종 RNA 편집이 이루어지기도 한다. 예를 들어 고등 식물의 미토콘드리아에서는 DNA 서열상의 CGG가 mRNA의 경우, UGG로 편집되어 트립토판을 코딩한다는 예가 알려져 있다.
5. 미토콘드리아 기원
미토콘드리아는 리케차와 유사한 호기성 세균인 α 프로테오박테리아가 진핵 세포에 들어간 결과로 획득된 것으로 추정된다.[230] 린 마굴리스의 세포 내 공생설에서는 호기성 박테리아가 기원으로 여겨졌으나, 1970년대에 이미 미토콘드리아의 기원이 α 프로테오박테리아라는 의견이 제시되었다. 탈질 세균인 ''Paracoccus denitrificans''와 암소 호기 조건에서 배양한 홍색 광합성 세균인 ''Rhodobacter sphaeroides''는 호흡 사슬 구성과 저해제 반응이 미토콘드리아와 유사하고, 특히 시토크롬 c가 미토콘드리아와 호환성을 갖는다는 점이 주목받았다.[265]
세포 핵 DNA에 코딩된 시토크롬 c뿐만 아니라, mtDNA에 코딩된 리보솜 RNA 서열을 이용한 계통 분석에서도 α 프로테오박테리아 기원임이 밝혀졌고, 1980년대에는 미토콘드리아의 α 프로테오박테리아 기원이 널리 받아들여졌다.[266]
초기 분석에서는 고등 식물 미토콘드리아의 리보솜 RNA 서열이 다른 미토콘드리아 서열과 비교하여 진화적 거리가 매우 작아, 미토콘드리아 기원이 여러 번 일어났다는 의견도 있었으나, 현재는 부정되었고 진핵생물의 미토콘드리아 기원은 단일 기원이라고 여겨진다.[267]
α 프로테오박테리아는 매우 다양한 세균을 포함하는 분류군이며, 그중 어떤 세균이 미토콘드리아의 기원인지에 대해서는 오랫동안 논의가 있었다. 초기에는 탈질 세균이나 광합성 세균이 기원이라고 생각되었지만, 샤페로닌 Hsp60(GroEL)을 사용한 계통 분석을 통해 리케차가 가장 가깝다는 것이 밝혀져 유력한 설이 되었다. 리케차는 세포 내 기생 생물이며, TCA 사이클을 가지고 호기 호흡을 할 수 있지만 해당 과정을 갖지 않고, 세포막에 ADP/ATP 수송체를 가지며, 게놈이 작고 AT 함량이 높은 점 등 미토콘드리아와 공통된 특징이 많다.[267]
1998년 발진티푸스를 일으키는 리케차의 일종인 ''Rickettsia prowazekii''의 게놈이 해독되었고, 조상으로 여겨지는 ''Reclinomonas americana''의 미토콘드리아 게놈과 공통 유전자 및 배치 순서가 보존된 유전자군 등이 발견되어 분석되었다. 대부분의 결과는 미토콘드리아가 리케차에 가깝다는 가설을 지지했지만, ADP/ATP 수송체는 기원이 다른 것으로 밝혀졌다.[268]
20세기 말부터 21세기 초, 전 세계 해양에는 자유 생활성 및 부유성 세균인 펠라지박터(잠정적으로 ''Candidatus'' Pelagibacter ubique로 명명)가 존재한다는 것이 밝혀졌다.[269] 펠라지박터는 리케차 목 중에서 가장 조상적인 위치에서 파생되었다고 추정되며, 미토콘드리아의 기원을 펠라지박터와 기타 일반적인 리케차 사이에서 찾을 수 있다.[270]
한편, 아메바와 비슷한 원생 생물인 펠로미사나 미포자충 등, 원생 생물 중에는 미토콘드리아가 없는 것도 있다. 이를 미토콘드리아 공생 이전의 진핵생물 생존으로 보는 설(아케조아 가설)이 있었으나, 이후 부정되었다.
6. 미토콘드리아와 질병
미토콘드리아의 에너지 대사 기능은 당뇨병(DM)과 같은 질병의 이해에 중요한 단서를 제공한다. 특히, 운동이 세포 내 미토콘드리아 발생 기작과 밀접한 관련이 있다는 연구 결과는 주목할 만하다.
세포 증식과 미토콘드리아의 관계에 대한 연구도 활발히 진행 중이다. 종양 세포는 빠른 증식을 위해 지질, 단백질, 뉴클레오타이드 같은 생체 활성 화합물을 합성하는 데 많은 ATP를 필요로 한다.[58] 종양 세포의 ATP는 주로 산화적 인산화(OxPhos) 경로를 통해 생성된다.[59] 이 경로를 방해하면 세포 주기가 정지되는데, 이는 미토콘드리아가 세포 증식에 중요한 역할을 한다는 것을 보여준다.[59] 미토콘드리아 ATP 생산은 세포 부피, 용질 농도, 세포 구조 조절뿐만 아니라 감염 시 세포 분열 및 분화에도 필수적이다.[60][61][62][63] 세포 주기의 각 단계마다 ATP 수준이 다르다는 사실은 ATP 양과 세포의 새로운 세포 주기 진입 능력 사이에 관계가 있음을 시사한다.[64] 세포 주기는 미토콘드리아에서 생성되는 ATP의 양에 민감하게 반응하며,[64] 이는 미토콘드리아가 세포 주기 조절에 중요한 역할을 한다는 가설을 뒷받침한다.[64] 구체적인 메커니즘은 아직 명확히 밝혀지지 않았지만, 낮은 에너지 세포 주기 체크포인트가 다음 세포 분열을 시작하기 전에 에너지 상태를 감시한다는 연구 결과가 있다.[71]
프로그램 세포 죽음(PCD)은 기관 발달, 세포 항상성 유지, 악성 형질 전환 예방, 항바이러스 방어, 병원체 제거, 염증 및 면역 세포 모집 등 다양한 생리학적 기능에 중요한 역할을 한다.[65]
미토콘드리아는 PCD의 한 형태인 세포자멸사의 내인성 경로에서 핵심적인 역할을 한다.[66] 최근에는 선천 면역의 신호 허브로도 확인되었다.[67] 미토콘드리아의 공생 기원은 다른 세포 구성 요소와 구별되며, 미토콘드리아 구성 요소가 세포질에 노출되면 감염 표지와 동일한 경로를 유발하여 세포자멸사, 자가포식 또는 전염증 유전자를 유도한다.[68][67]
미토콘드리아는 시토크롬 'c'를 방출하여 세포자멸체 형성을 유도하고, 다양한 손상 관련 분자 패턴(DAMP)의 공급원이다. DAMP는 감염 중 병원체 관련 분자 패턴(PAMP)에 반응하는 패턴 인식 수용체(PRR)에 의해 인식된다. 미토콘드리아 mtDNA는 CpG 부위 메틸화가 부족하여 세균 DNA와 유사하며, Toll-like receptor 9 및 cGAS에 의해 감지될 수 있다.[68] 이중 가닥 RNA(dsRNA)는 양방향 미토콘드리아 전사로 생성되며, RIG-I 유사 수용체를 통해 바이러스 감지 경로를 활성화한다.[68] 미토콘드리아 단백질의 ''N''-포름화는 포르밀 펩타이드 수용체에 의해 인식될 수 있다.[68]
일반적으로 미토콘드리아 구성 요소는 세포 내에서 격리되어 있지만, 세포자멸사 동안 미토콘드리아 막 투과성이 발생하거나 미토콘드리아 손상 후 수동적으로 방출된다. 미토콘드리아는 대사 신호에 반응하여 mtDNA를 방출하며 선천 면역에 적극적으로 관여한다.[67] 미토콘드리아는 세포 내 위치로, BAX, MAVS (바깥 막에 위치), NLRX1 (기질에서 발견)과 같은 면역 및 세포자멸사 조절 단백질을 위한 위치이며, 이러한 단백질은 미토콘드리아 대사 상태 및 역학에 의해 조절된다.[67]
미토콘드리아는 다음과 같은 여러 대사 활동에서 중요한 역할을 한다.
- 미토콘드리아 활성 산소를 통한 신호 전달[69]
- 막 전위 조절[17]
- 칼슘 신호 전달 (칼슘 유발 세포 자멸사 포함)[70]
- 세포 대사 조절[71]
- 헴 합성[72] (참고: ''포르피린'')
- 스테로이드 합성[73]
- 호르몬 신호 전달[74] (미토콘드리아 에스트로겐 수용체(mtER)는 뇌[75]와 심장[76] 등 다양한 조직과 세포에서 발견된다.)
- 면역 세포 발달 및 기능[77]
- 신경 미토콘드리아는 미세 아교 세포에 대한 신경 상태 보고를 통해 세포 품질 관리에 기여[108]
- 발달 중인 뉴런의 미토콘드리아는 미세 아교 세포로의 세포 간 신호 전달에 기여하며 뇌 발달 조절에 필수적[78]
간 세포의 미토콘드리아는 단백질 대사 부산물인 암모니아를 해독하는 효소를 가지고 있다. 이러한 기능을 조절하는 유전자의 돌연변이는 미토콘드리아 질환을 유발할 수 있다.
미토콘드리아 단백질은 조직과 종에 따라 다르다. 사람 심장 미토콘드리아에는 615가지, 쥐에서는 940가지 단백질이 확인되었다.[79][80] 미토콘드리아 프로테옴은 동적으로 조절된다.[81]
미토콘드리아 손상과 기능 부전은 세포 대사에 영향을 미쳐 다양한 질환의 원인이 된다. 미토콘드리아 질환은 자폐증을 포함한 신경 질환,[14] 근육병증, 당뇨병, 다발성 내분비병증 등으로 나타날 수 있다.[185] mtDNA 돌연변이로 인한 질환에는 Kearns–Sayre 증후군, MELAS 증후군, 레베르 유전성 시신경병증이 있다.[186] 이러한 질환은 대부분 난자를 통해 유전된다. Kearns-Sayre 증후군, Pearson 증후군, 진행성 외안 안근마비는 mtDNA 재배열, MELAS 증후군, 레베르 유전성 시신경병증, MERRF 증후군 등은 mtDNA 점 돌연변이에 기인한다.[185]
약물 내성 암세포는 미토콘드리아 생성 증가를 보이며,[187] 암세포가 면역 세포로부터 미토콘드리아를 빼앗을 수 있다는 연구 결과도 있다.[188]
핵 유전자 결함은 미토콘드리아 단백질 기능 부전을 유발하며, 프리드리히 운동실조증, 유전성 경직성 하반신 마비, 윌슨병 등이 이에 해당한다.[189] 이러한 질환은 우성 관계로 유전된다. 코엔자임 Q10 결핍, 바스 증후군 등은 산화적 인산화 효소 핵 돌연변이로 발생한다.[185] 살충제 노출과 파킨슨병 발병 간 연관성 등 환경적 요인도 미토콘드리아 질환을 유발할 수 있다.[190][191] 미토콘드리아 기능 부전은 정신분열증, 양극성 장애, 치매, 알츠하이머병,[192][193] 파킨슨병, 간질, 뇌졸중, 심혈관 질환, 근육통성 뇌척수염/만성 피로 증후군(ME/CFS), 망막색소변성증, 당뇨병과 관련된다.[194][195]
2형 당뇨병 환자의 심근증에는 미토콘드리아 매개 산화 스트레스가 관여한다. 지방산 공급 증가는 심근 세포의 지방산 섭취 및 산화를 증가시켜 활성 산소종(ROS) 생성을 늘린다. ROS는 탈공역 단백질(UCP) 증가, 아데닌 뉴클레오티드 수송체(ANT)를 통한 양성자 누출 증폭을 유발하여 미토콘드리아를 탈공역시킨다. 탈공역은 산소 소비를 늘려 지방산 산화 증가를 악화시키는 악순환을 초래한다. 미토콘드리아 탈공역으로 ATP 합성 효율이 감소하고, 손상된 근소포체 칼슘 방출과 미토콘드리아 재흡수 감소는 세포질 칼슘 농도를 제한한다. 미토콘드리아 내 칼슘 농도 감소는 탈수소 효소 활성화 및 ATP 합성을 저해하여 당뇨병 환자의 심장 문제를 유발한다.[196]
미토콘드리아는 고환 체세포 발달, 정원 세포 줄기 세포 분화, 내강 산성화, 테스토스테론 생성 등을 조절하며, 정자 미토콘드리아 기능 부전은 불임의 원인이 될 수 있다.[197]
미토콘드리아 대체 요법(MRT)은 기증자 미토콘드리아를 사용하여 mtDNA 돌연변이 질환 전파를 방지하는 체외 수정 방법이지만,[198] 연구가 더 필요하며 유전자 변형 및 안전성 우려가 있다. 이러한 질환은 드물지만 쇠약하고 진행성 질환일 수 있어 윤리적 문제를 야기한다.[199]
노화의 징후#미토콘드리아 기능 부전도 참조
전자 누출로 활성 산소가 형성되어 미토콘드리아 내 산화 스트레스를 유발하고 mtDNA 돌연변이율을 높인다.[200] 1956년 제안된 산화 스트레스와 노화의 연관성은[201] 미토콘드리아 자유 라디칼 노화설로 구체화되었다.[202] 산화 스트레스는 mtDNA 돌연변이를 유발하고, 이는 효소 이상 및 추가적인 산화 스트레스를 유발하는 악순환을 초래한다.
노화 과정에서 미토콘드리아는 호흡 사슬 단백질 효소 활성 감소 등 여러 변화를 겪는다.[203][204] 돌연변이 mtDNA는 매우 오래된 세포의 약 0.2%에서만 발견된다.[205] 미토콘드리아 유전체 결실은 산화 스트레스와 파킨슨병 신경 세포 사멸을 유발한다.[206] 미토콘드리아 기능 부전은 근위축성 측삭 경화증에서도 발생한다.[207][208]
미토콘드리아는 난소 기능에 중요한 ATP를 제공하며, 기능 감소는 염증을 유발하여 조기 난소 부전 및 가속화된 난소 노화를 초래한다. 기능 부전은 정량적(mtDNA 복제 수 및 결실), 질적(돌연변이, 가닥 절단), 산화적 손상(ROS)으로 나타나며, 난소 노화, 난자-황체 세포 상호 작용 방해, 유전 질환(취약 X 증후군), 배아 선택 방해와 관련된다.[209]
미토콘드리아는 해당계나 TCA 회로 산물을 이용하여 전자 전달계에 고에너지 전자를 전달하고, 프로톤 농도 기울기로 ATP 합성효소를 구동하여 ADP를 산화적 인산화로 ATP로 변환한다.[234]
해당계 등 미토콘드리아가 관여하지 않는 ATP 생성계도 있지만, 진핵생물 세포 활동에 필요한 ATP 대부분은 미토콘드리아에서 공급된다. 미토콘드리아 TCA 회로에서는 ATP와 등가인 GTP도 생성되어 "진핵 세포의 에너지를 만들어내는 장소"로 비유된다.[235]
미토콘드리아는 ATP, GTP 합성 외에도 스테로이드, 헴 합성, 칼슘, 철 농도 조절, 세포 주기 및 세포 자멸사 조절 등 다양한 기능을 수행하며, 기능에 따라 특정 세포에서만 작동한다.
이러한 다양한 기능에는 다수 유전자가 관여하며, 유전자 변이 세포가 자연 면역에서 제거되지 않으면 미토콘드리아 기능 저하로 미토콘드리아 질환을 유발한다.
1955년 오토 바르부르크는 체세포가 장기간 저산소 상태에 노출되면 호흡 장애를 일으키고, 정상 산소 농도로 되돌려도 변성, 괴사되며, 일부 산소 호흡 대신 에너지 생성 경로를 앙진시켜 생존하는 세포가 암세포가 된다는 설을 발표했다. 이 설에서는 산소 호흡보다 해당 작용에 의한 에너지 생산에 의존하는 세포는 하등 동물, 태생기 미숙 세포가 일반적이며, 체세포가 ATP 생산을 산소 호흡 대신 해당 작용에 의존하여 세포가 퇴화해 암세포가 된다고 했다.[271]
암세포 발생과 mtDNA 돌연변이 관여는 오래전부터 지적되었다. 발암성 화학 물질이 DNA보다 mtDNA에 결합하기 쉽고, 암 조직 mtDNA가 정상 조직보다 높은 비율로 돌연변이 축적이 관찰되었기 때문이다. 그러나 모성 유전 암 미확인, mtDNA 단독 영향 검증 방법 미확립 등이 가설 증명 장애였다.
2008년 쓰쿠바 대학의 하야시 준이치 등은 암 전이능 획득이라는 암세포 악성화에 mtDNA 변이가 관여함을 지적했다.[272] 마우스 폐암세포 세포질 이식을 통한 세포 잡종 비교로 mtDNA 특수 병원성 돌연변이가 암세포 전이능 획득 원인임을 발견하고, 사람 암세포주에서도 mtDNA 돌연변이가 암세포 전이능을 유도할 수 있음을 밝혀 mtDNA가 ATP 합성 외 생명 현상에도 관여함을 밝혔다. 하야시 등에 따르면 mtDNA 돌연변이에는 활성 산소종(ROS) 매개가 중요하며,[272] ROS 제거로 전이능 억제가 가능할 것이라고 추측했다. 암세포 전이능 획득 메커니즘은 복잡하며 다양한 요인이 고려될 수 있어, 이는 그 요인 중 하나일 뿐이다.
교토 대학의 이가키 타츠리는 ① Ras 유전자 활성화와 미토콘드리아 기능 장애 세포는 세포 노화를 일으켜 세포 노화 관련 분비 인자(SASP 인자)를 방출하고 주변 조직 암화를 촉진하며, ② 세포 분열 정지와 JNK 유전자 활성화 증폭으로 세포 내 JNK 활성이 증대되어 SASP 인자 생산을 유도함을 밝혔다.[273][274][275]
7. 대한민국과 미토콘드리아
이 섹션은 주어진 원본 소스에 존재하지 않으므로, 작성할 수 없습니다.
참조
[1]
백과사전
mitochondrion
https://web.archive.[...]
Oxford University Press
[2]
서적
Biology: Exploring Life
http://www.phschool.[...]
Pearson/Prentice Hall
2009-01-06
[3]
웹사이트
Mighty Mitochondria and Neurodegenerative Diseases
https://sitn.hms.har[...]
2012-02-01
[4]
뉴스
Animal that doesn't need oxygen to survive discovered
https://www.newscien[...]
[5]
논문
A cnidarian parasite of salmon (Myxozoa: ''Henneguya'') lacks a mitochondrial genome
2020-03
[6]
논문
Evolutionary biology: essence of mitochondria
2003-11
[7]
논문
Organelles that illuminate the origins of ''Trichomonas'' hydrogenosomes and ''Giardia'' mitosomes
2017-04
[8]
논문
Quantification of mitochondrial morphology in neurites of dopaminergic neurons using multiple parameters
2016-03
[9]
논문
Mitochondrial biogenesis: pharmacological approaches
[10]
논문
Mitochondrial biogenesis in health and disease. Molecular and therapeutic approaches
[11]
논문
Is a 'Mitochondrial Psychiatry' in the Future? A Review
[12]
논문
Mitochondrial dysfunction in cardiac disease: ischemia--reperfusion, aging, and heart failure
2001-06
[13]
논문
Mitochondrial biogenesis and dynamics in the developing and diseased heart
2015-10
[14]
논문
Evidence of Mitochondrial Dysfunction in Autism: Biochemical Links, Genetic-Based Associations, and Non-Energy-Related Mechanisms
2017
[15]
논문
Normal and disordered reticulocyte maturation
2011-05
[16]
서적
Molecular Biology of the Cell
Garland Publishing Inc.
[17]
서적
Fundamentals of Biochemistry
https://archive.org/[...]
John Wiley and Sons, Inc.
[18]
논문
On the origin of mitochondria: a genomics perspective
2003-01
[19]
논문
Origin and Early Evolution of the Eukaryotic Cell
2021-10
[20]
웹사이트
Mitochondrion – much more than an energy converter
http://www.bscb.org/[...]
British Society for Cell Biology
[21]
논문
VDAC channels
2001-09
[22]
논문
The supramolecular assemblies of voltage-dependent anion channels in the native membrane
2007-07
[23]
논문
Structure and evolution of mitochondrial outer membrane proteins of beta-barrel topology
2010-06
[24]
논문
Protein transport into mitochondria
https://epub.ub.uni-[...]
2000-04
[25]
논문
Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario
2006-08
[26]
논문
Formation and regulation of mitochondrial membranes
2014-01
[27]
논문
Cardiolipin and apoptosis
2002-12
[28]
논문
Advances in methods to analyse cardiolipin and their clinical applications
2022-12
[29]
논문
Mitochondrial fission, fusion, and stress
2012-08
[30]
논문
Mitochondrial Ultrastructure Is Coupled to Synaptic Performance at Axonal Release Sites
2018
[31]
논문
Structure and dynamics of the mitochondrial inner membrane cristae
[32]
논문
Mitochondrial DNA nucleoid structure
2012-09
[33]
논문
The molecular machinery of Keilin's respiratory chain
2003-12
[34]
논문
Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria
2007-07
[35]
논문
Protein import into mitochondria
[36]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[37]
학술지
Succinate dehydrogenase and fumarate hydratase: linking mitochondrial dysfunction and cancer
2006-08-00
[38]
서적
Biochemistry
Wiley
2004
[39]
서적
Physical Chemistry
Freeman
2006
[40]
학술지
The role of oxidative damage in mitochondria during aging: a review
http://pdfs.semantic[...]
2004-05-00
[41]
학술지
Chemiosmotic hypothesis of oxidative phosphorylation
1967-01-00
[42]
학술지
Proton current flow in mitochondrial systems
1967-06-00
[43]
웹사이트
Chemistry 1997
http://nobelprize.or[...]
2007-12-16
[44]
학술지
Thermoregulation: what role for UCPs in mammals and birds?
2005-11-00
[45]
학술지
Mitochondrial fatty acid synthesis, fatty acids and mitochondrial physiology
2017-01-00
[46]
학술지
Altering the Mitochondrial Fatty Acid Synthesis (mtFASII) Pathway Modulates Cellular Metabolic States and Bioactive Lipid Profiles as Revealed by Metabolomic Profiling
2016-03-00
[47]
학술지
Impact of Mitochondrial Fatty Acid Synthesis on Mitochondrial Biogenesis
2018-10-00
[48]
학술지
The emerging role of the mitochondrial fatty-acid synthase (mtFASII) in the regulation of energy metabolism
2019-11-00
[49]
학술지
Mitochondrial calcium overload is a key determinant in heart failure
2015-09-00
[50]
서적
Basic Neurochemistry
Lippincott Williams & Wilkins
[51]
학술지
Mitochondrial calcium and its role in calcification. Histochemical localization of calcium in electron micrographs of the epiphyseal growth plate with K-pyroantimonate
1974-05-00
[52]
학술지
The role of mitochondria in growth plate calcification as demonstrated in a rachitic model
1978-07-00
[53]
학술지
Mitochondria-endoplasmic reticulum choreography: structure and signaling dynamics
2007-10-00
[54]
학술지
Mitochondria - the Kraken wakes!
1998-03-00
[55]
학술지
Calcium release channel RyR2 regulates insulin release and glucose homeostasis
2015-05-00
[56]
학술지
Pulsing of membrane potential in individual mitochondria: a stress-induced mechanism to regulate respiratory bioenergetics in Arabidopsis
2012-03-00
[57]
학술지
Mitochondrial free Ca²⁺ levels and their effects on energy metabolism in Drosophila motor nerve terminals
2013-06-00
[58]
학술지
Mitochondrial metabolism and cancer
2009-10-00
[59]
학술지
Energy metabolism in tumor cells
2007-03-00
[60]
학술지
ROS-mediated PI3K activation drives mitochondrial transfer from stromal cells to hematopoietic stem cells in response to infection
2019-12-00
[61]
학술지
ATP synthase. The machine that makes ATP
1994-12-00
[62]
학술지
The metabolism of human mesenchymal stem cells during proliferation and differentiation
2011-10-00
[63]
학술지
A role for anions in ATP synthesis and its molecular mechanistic interpretation
2011-06-00
[64]
학술지
Changes in mitochondrial mass, membrane potential, and cellular adenosine triphosphate content during the cell cycle of human leukemic (HL-60) cells
1999-07-00
[65]
학술지
Cell death as part of innate immunity: Cause or consequence?
2021-08-00
[66]
학술지
Apoptotic pathways: the roads to ruin
1998-09-00
[67]
학술지
Metabolism and Innate Immunity Meet at the Mitochondria
2021-07-27
[68]
학술지
A break in mitochondrial endosymbiosis as a basis for inflammatory diseases
2024-02-00
[69]
학술지
Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers
2013-02-00
[70]
학술지
Mitochondrial calcium signalling and cell death: approaches for assessing the role of mitochondrial Ca2+ uptake in apoptosis
[71]
논문
Mitochondria: more than just a powerhouse
2006-07-00
[72]
논문
Evolutionary consideration on 5-aminolevulinate synthase in nature
1997-08-00
[73]
논문
T channels and steroid biosynthesis: in search of a link with mitochondria
2006-08-00
[74]
논문
Estrogenic control of mitochondrial function and biogenesis
2008-12-00
[75]
논문
Different expression of alpha and beta mitochondrial estrogen receptors in the aging rat brain: interaction with respiratory complex V
2010-08-00
[76]
논문
Sexual hormones: effects on cardiac and mitochondrial activity after ischemia-reperfusion in adult rats. Gender difference
2012-10-00
[77]
논문
Mitochondria as central hub of the immune system
2019-09-00
[78]
논문
Microglial control of neuronal development via somatic purinergic junctions
2022-09-00
[79]
논문
Characterization of the human heart mitochondrial proteome
2003-03-00
[80]
논문
Systematic characterization of the murine mitochondrial proteome using functionally validated cardiac mitochondria
2008-04-00
[81]
논문
Altered proteome biology of cardiac mitochondria under stress conditions
2008-06-00
[82]
논문
A Eukaryote without a Mitochondrial Organelle
2016-05-00
[83]
논문
Mitochondrial fusion, division and positioning in plants
2010-06-00
[84]
논문
Connecting variability in global transcription rate to mitochondrial variability
2010-12-00
[85]
논문
Mitochondrial variability as a source of extrinsic cellular noise
[86]
논문
Cytoskeleton and mitochondrial morphology and function
1998-07-00
[87]
논문
What is the function of mitochondrial networks? A theoretical assessment of hypotheses and proposal for future research
2015-06-00
[88]
논문
Interrelationships of endoplasmic reticulum, mitochondria, intermediate filaments, and microtubules--a quadruple fluorescence labeling study
[89]
논문
Vimentin supports mitochondrial morphology and organization
2008-02-00
[90]
논문
Ca(2+) transfer from the ER to mitochondria: when, how and why
2009-11-00
[91]
논문
MAM: more than just a housekeeper
2009-02-00
[92]
논문
An intimate liaison: spatial organization of the endoplasmic reticulum-mitochondria relationship
2010-08-00
[93]
논문
Intracellular trafficking of phospholipids: import of phosphatidylserine into mitochondria
[94]
논문
Interactions between the endoplasmic reticulum, mitochondria, plasma membrane and other subcellular organelles
2009-10-00
[95]
논문
Fission and selective fusion govern mitochondrial segregation and elimination by autophagy
2008-01-00
[96]
논문
Making heads or tails of phospholipids in mitochondria
2011-01-00
[97]
논문
An ER-mitochondria tethering complex revealed by a synthetic biology screen
2009-07-00
[98]
논문
A unique mitochondria-associated membrane fraction from rat liver has a high capacity for lipid synthesis and contains pre-Golgi secretory proteins including nascent lipoproteins
1994-11-00
[99]
논문
Essential Roles of Intracellular Calcium Release Channels in Muscle, Brain, Metabolism, and Aging
2015-00-00
[100]
논문
Functional coupling between ryanodine receptors, mitochondria and Ca(2+) ATPases in rat submandibular acinar cells
2008-05-00
[101]
논문
Sorting of calcium signals at the junctions of endoplasmic reticulum and mitochondria
2001-04-00
[102]
논문
The IP(3) receptor-mitochondria connection in apoptosis and autophagy
2011-05-00
[103]
논문
Mag-Fluo4 in T cells: Imaging of intra-organelle free Ca2+ concentrations
2017-06-00
[104]
논문
Old players in a new role: mitochondria-associated membranes, VDAC, and ryanodine receptors as contributors to calcium signal propagation from endoplasmic reticulum to the mitochondria
[105]
논문
σ-1 receptor at the mitochondrial-associated endoplasmic reticulum membrane is responsible for mitochondrial metabolic regulation
2012-12-00
[106]
논문
Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca(2+) signaling and cell survival
2007-11-00
[107]
논문
Endoplasmic Reticulum-Mitochondrial Contactology: Structure and Signaling Functions
2018-07-00
[108]
논문
Microglia monitor and protect neuronal function through specialized somatic purinergic junctions
https://epub.ub.uni-[...]
2022-02-10
[109]
논문
The Genomics and Cell Biology of Host-Beneficial Intracellular Infections
2021-10-00
[110]
논문
Mitochondria and the origin of eukaryotes
https://knowablemaga[...]
2022-06-08
[111]
서적
Origins of Sex: Three billion years of genetic recombination
https://archive.org/[...]
Yale University Press
[112]
서적
Origin of mitochondria and hydrogenosomes
Springer Verlag
[113]
논문
Mitochondrial connection to the origin of the eukaryotic cell
2003-04-00
[114]
논문
The genome of Rickettsia prowazekii and some thoughts on the origin of mitochondria and hydrogenosomes
1999-05-00
[115]
논문
Mitochondrial evolution
1999-03-00
[116]
논문
Phylogenomic evidence for a common ancestor of mitochondria and the SAR11 clade
2011-06-14
[117]
논문
Deep mitochondrial origin outside the sampled alphaproteobacteria
2018-05-00
[118]
논문
Phylogenetic analyses with systematic taxon sampling show that mitochondria branch within Alphaproteobacteria
Springer Science and Business Media LLC
2020-09-00
[119]
논문
Dating Alphaproteobacteria evolution with eukaryotic fossils
2021-06-00
[120]
논문
On the bacterial ancestry of mitochondria: New insights with triangulated approaches
2022-05-16
[121]
논문
Site-and-branch-heterogeneous analyses of an expanded dataset favour mitochondria as sister to known Alphaproteobacteria
https://www.research[...]
2022-03-00
[122]
논문
The evolutionary origin of host association in the Rickettsiales
2022-08-00
[123]
논문
Properties of human mitochondrial ribosomes
2003-09-00
[124]
논문
On the origin of mitosing cells
1967-03-00
[125]
논문
Rickettsiaceae, rickettsia-like endosymbionts, and the origin of mitochondria
2001-02-00
[126]
논문
Determining divergence times with a protein clock: update and reevaluation
1997-11-00
[127]
논문
Archamoebae: the ancestral eukaryotes?
[128]
논문
The first metazoa living in permanently anoxic conditions
2010-04-00
[129]
잡지
The mud creature that lives without oxygen
https://www.newscien[...]
2010-04-07
[130]
논문
Mitochondrion-related organelles in eukaryotic protists
[131]
논문
The Oxymonad Genome Displays Canonical Eukaryotic Complexity in the Absence of a Mitochondrion
https://academic.oup[...]
2019-10-00
[132]
논문
Genomics of Preaxostyla Flagellates Illuminates the Path Towards the Loss of Mitochondria
2023-12-00
[133]
논문
Mitochondria: dynamic organelles in disease, aging, and development
2006-06-00
[134]
논문
Counting target molecules by exponential polymerase chain reaction: copy number of mitochondrial DNA in rat tissues
1992-03-00
[135]
논문
Mitochondrial DNA replication in mammalian cells: overview of the pathway
2018-07-00
[136]
논문
Sequence and organization of the human mitochondrial genome
1981-04-00
[137]
논문
Linear mitochondrial DNAs of yeasts: frequency of occurrence and general features
1993-04-00
[138]
논문
Intervening sequences in the mitochondrial genome
1978-12-00
[139]
논문
A maturase-encoding group IIA intron of yeast mitochondria self-splices in vitro
1992-04-00
[140]
논문
Genome structure and gene content in protist mitochondrial DNAs
1998-02-00
[141]
논문
Mitochondria of protists
[142]
논문
Nuclear and mitochondrial tRNA-lookalikes in the human genome
[143]
논문
The single mitochondrial chromosome typical of animals has evolved into 18 minichromosomes in the human body louse, Pediculus humanus
2009-05-01
[144]
논문
Mitochondrial DNA: a tool for populational genetics studies
1998-12-01
[145]
논문
Mitochondrial DNA and human evolution
1987-01-01
[146]
논문
Harvesting the fruit of the human mtDNA tree
2006-06-01
[147]
논문
Reconstructing human origins in the genomic era
2006-09-01
[148]
논문
Neandertal DNA sequences and the origin of modern humans
1997-07-01
[149]
논문
Correcting for purifying selection: an improved human mitochondrial molecular clock
2009-06-01
[150]
논문
Estimate of the mutation rate per nucleotide in humans
2000-09-01
[151]
논문
Directed panspermia
http://profiles.nlm.[...]
2014-10-21
[152]
논문
A different genetic code in human mitochondria
1979-11-01
[153]
웹사이트
The Genetic Codes
https://www.ncbi.nlm[...]
2019-01-07
[154]
논문
The genetic code in mitochondria and chloroplasts
1990-12-01
[155]
논문
RNA editing in plant mitochondria
1989-12-01
[156]
논문
Parallel evolution of the genetic code in arthropod mitochondrial genomes
2006-05-01
[157]
서적
Parkinson's Disease
https://books.google[...]
CRC Press
[158]
논문
New insights into the role of mitochondria in aging: mitochondrial dynamics and more
2010-08-01
[159]
논문
Whole cell cryo-electron tomography suggests mitochondria divide by budding
2014-08-01
[160]
논문
Mitochondrial DNA can be inherited from fathers, not just mothers
2019-01-01
[161]
웹사이트
Sexual Reproduction in Humans: Copulation and Fertilization
http://home.comcast.[...]
2006-01-01
[162]
논문
Ubiquitin tag for sperm mitochondria
1999-11-01
[163]
논문
The Hows and Whys of Cytoplasmic Inheritance in Seed Plants
[164]
논문
The exceptional mitochondrial DNA system of the mussel family Mytilidae
2000-12-01
[165]
논문
The fate of paternal mitochondrial DNA in developing female mussels, Mytilus edulis: implications for the mechanism of doubly uniparental inheritance of mitochondrial DNA
1998-01-01
[166]
웹사이트
Male and Female Mitochondrial DNA Lineages in the Blue Mussel (Mytilus edulis) Species Group
http://mbe.library.a[...]
[167]
논문
Paternal transmission of mitochondrial DNA is (fortunately) rare
2003-10-01
[168]
논문
Mammalian mitochondria possess homologous DNA recombination activity
1996-11-01
[169]
논문
Animal mitochondrial DNA recombination
1997-05-01
[170]
논문
How clonal are human mitochondria?
1999-03-01
[171]
논문
Linkage disequilibrium and recombination in hominid mitochondrial DNA
1999-12-01
[172]
논문
A reduction of mitochondrial DNA molecules during embryogenesis explains the rapid segregation of genotypes
2008-02-01
[173]
논문
The mitochondrial bottleneck occurs without reduction of mtDNA content in female mouse germ cells
2007-03-01
[174]
논문
The mitochondrial DNA genetic bottleneck results from replication of a subpopulation of genomes
2008-12-01
[175]
논문
Stochastic modelling, Bayesian inference, and new in vivo measurements elucidate the debated mtDNA bottleneck mechanism
2015-06-01
[176]
논문
Nuclear and mitochondrial DNA repair in selected eukaryotic aging model systems
2012
[177]
논문
DNA damage related crosstalk between the nucleus and mitochondria
2017-06-00
[178]
논문
8-Oxoguanine accumulation in mitochondrial DNA causes mitochondrial dysfunction and impairs neuritogenesis in cultured adult mouse cortical neurons under oxidative conditions
2016-02-00
[179]
논문
Homologous recombination-mediated repair of DNA double-strand breaks operates in mammalian mitochondria
2018-05-00
[180]
논문
Involvement of mitochondrial-targeted RecA in the repair of mitochondrial DNA in the moss, Physcomitrella patens
2007-02-00
[181]
논문
Microhomology-mediated end joining is the principal mediator of double-strand break repair during mitochondrial DNA lesions
2016-01-00
[182]
논문
The unusual mitochondrial compartment of Cryptosporidium parvum
2005-02-00
[183]
논문
An aerobic eukaryotic parasite with functional mitochondria that likely lacks a mitochondrial genome
2019-04-00
[184]
웹사이트
Veritable powerhouse{{snd}}even without DNA: Parasitic algae from the dinoflagellate lineage have organized their genetic material in an unprecedented way
https://www.scienced[...]
2019-05-08
[185]
논문
Mitochondrial disorders
2004-10-00
[186]
논문
Mitochondrial DNA mutations in human disease
2005-05-00
[187]
논문
Targeting tumor phenotypic plasticity and metabolic remodeling in adaptive cross-drug tolerance
2019-08-00
[188]
논문
Intercellular nanotubes mediate mitochondrial trafficking between cancer and immune cells
2022-01-00
[189]
논문
Mitochondria
2003-09-00
[190]
논문
Environment, mitochondria, and Parkinson's disease
2002-06-00
[191]
논문
Pesticides and impairment of mitochondrial function in relation with the parkinsonian syndrome
2007-01-00
[192]
논문
Abeta and human amylin share a common toxicity pathway via mitochondrial dysfunction
2010-04-00
[193]
논문
Molecular basis of substrate recognition and degradation by human presequence protease
2014-07-00
[194]
논문
Mitochondrial disease
2006-07-00
[195]
논문
Mitochondrial dysfunction and molecular pathways of disease
2007-08-00
[196]
논문
Mitochondria in the diabetic heart
2010-11-00
[197]
논문
The Role of Mitochondria in Human Fertility and Early Embryo Development: What Can We Learn for Clinical Application of Assessing and Improving Mitochondrial DNA?
2022-02-00
[198]
논문
Embryo and Its Mitochondria
2021-01-00
[199]
간행물
Introduction
https://www.ncbi.nlm[...]
National Academies Press (US)
2023-12-05
[200]
논문
Normal oxidative damage to mitochondrial and nuclear DNA is extensive
1988-09-00
[201]
논문
Aging: a theory based on free radical and radiation chemistry
1956-07-00
[202]
논문
The biologic clock: the mitochondria?
1972-04-00
[203]
웹사이트
Mitochondria and Aging
http://www.circuitbl[...]
circuitblue.co
2006-10-23
[204]
논문
Decline with age of the respiratory chain activity in human skeletal muscle
1994-04-00
[205]
논문
Mitochondrial mutations in mammalian aging: an over-hasty about-turn?
[206]
논문
High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease
2006-05-00
[207]
논문
Targeting mitochondrial dysfunction in amyotrophic lateral sclerosis: a systematic review and meta-analysis
2019-08-00
[208]
논문
Mitochondrial bioenergetic deficits in C9orf72 amyotrophic lateral sclerosis motor neurons cause dysfunctional axonal homeostasis
2021-02-00
[209]
논문
Mitochondria in Ovarian Aging and Reproductive Longevity
2020-11-00
[210]
논문
Einige Bemerkungen über die Endigungen der Hautnerven und den Bau der Muskeln
https://www.biodiver[...]
2023-06-22
[211]
논문
Mitochondria: a historical review
1981-12-01
[212]
서적
Die Elementarorganismen und ihre Beziehungen zu den Zellen
https://www.deutsche[...]
Veit & Co.
2023-06-23
[213]
백과사전
mitochondria
http://www.etymonlin[...]
2013-05-23
[214]
논문
Ueber die Spermatogenese der Vertebraten und höherer Evertebraten. II. Theil: Die Histiogenese der Spermien.
https://www.biodiver[...]
2018-01-14
[215]
논문
Die vitale Farbung, eine Darstellungsmethode der Zellgranula
https://www.biodiver[...]
2023-06-23
[216]
논문
Über das Vorkommen von Mitochondrien bezw. Chondromiten in Pflanzenzellen
[217]
논문
Cytoplasmic fixation
https://www.biodiver[...]
2023-06-23
[218]
논문
Über sauerstoffatmende Körnchen aus Leberzellen und über Sauerstoffatmung in Berkefeld-Filtraten wässriger Leberextrake
https://www.biodiver[...]
2023-06-23
[219]
논문
Powerhouse of the cell
[220]
논문
Mitochondrial biology, targets, and drug delivery
2015-06-01
[221]
논문
Endosymbiotic theories for eukaryote origin
2015-09-01
[222]
서적
ミトコンドリアが進化を決めた
みすず書房
2007-01-01
[223]
웹사이트
mitochondria
http://www.etymonlin[...]
Online Etymology Dictionary
2018-11-27
[224]
문서
미토콘드리아의 매트릭스(mitochondrial matrix)
[225]
서적
標準組織学 総論(第3版)
医学書院
1988-02-01
[226]
논문
MAM: more than just a housekeeper
[227]
논문
Protein transport into mitochondria
[228]
논문
Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario
[229]
논문
Structure and dynamics of the mitochondrial inner membrane cristae
[230]
서적
ミトコンドリアはどこからきたか
日本放送出版
2000-06-30
[231]
서적
標準組織学 総論(第3版)
医学書院
1988-02-01
[232]
서적
標準組織学 総論(第3版)
医学書院
1988-02-01
[233]
서적
標準組織学 総論(第3版)
医学書院
1988-02-01
[234]
문서
미토콘드리아의 전자전달계 저해제들
[235]
문서
미토콘드리아와 에너지 생성
[236]
문서
포도당 한 분자로부터 ATP 생성량
[237]
논문
Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria
[238]
웹사이트
TCA회로
http://www.sc.fukuok[...]
[239]
서적
Molecular Cell Biology, Fifth Edition
W. H. Freeman
[240]
서적
Illustrated 하퍼 생화학 (원서 27판)
마루젠
2007-01-30
[241]
논문
ATP는 어떻게 막을 통과하는가: 루프의 협조적 스윙에 의한 미토콘드리아 ADP/ATP 캐리어의 기능 발현
[242]
웹사이트
제5회 5. 에너지의 생산-사이트졸과 미토콘드리아
http://www.tmd.ac.jp[...]
[243]
서적
In: Biochemistry.
W.H. Freeman and Company
[244]
논문
The role of oxidative damage in mitochondria during aging: A review
[245]
논문
Vitamin C enters mitochondria via facilitative glucose transporter 1 (Glut1) and confers mitochondrial protection against oxidative injury
http://www.fasebj.or[...]
[246]
저널
Chemiosmotic hypothesis of oxidative phosphorylation
1967-01-14
[247]
저널
Proton current flow in mitochondrial systems
1967-06-24
[248]
웹사이트
Chemistry 1997
https://www.nobelpri[...]
Nobel Foundation
2007-12-16
[249]
서적
Illustrated ハーパー・生化学(原書27版)
丸善
2007-01-30
[250]
저널
新たな乳酸の見方
[251]
저널
Thermoregulation: What Role for UCPs in Mammals and Birds?
[252]
저널
Voltage-dependent anion channel (VDAC) as mitochondrial governator--thinking outside the box.
[253]
저널
アポトーシスの分子機構
https://doi.org/10.1[...]
[254]
저널
Mitochondria-endoplasmic reticulum choreography: structure and signaling dynamics
[255]
서적
Basic Neurochemistry
Lippincott Williams & Wilkins
[256]
저널
Mitochondria - the kraken wakes!
[257]
저널
T channels and steroid biosynthesis: in search of a link with mitochondria
[258]
저널
Mitochondrial calcium and its role in calcification.
[259]
저널
The role of mitochondria in growth plate calcification as demonstrated in a rachitic model.
[260]
저널
ミトコンドリアDNAに突然変異をもつ細胞は自然免疫により排除されることを発見
2011-10-12
[261]
저널
The genetic code in mitochondria and chloroplasts
1990-12-01
[262]
저널
RNA editing in plant mitochondria
[263]
서적
不思議な生き物-生命38億年の歴史と謎
角川学芸出版
2013-04-25
[264]
저널
Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues
[265]
저널
''Paracoccus denitrificans'' and the evolutionary origin of the mitochondrion.
[266]
저널
Mitochondrial origins
http://www.pnas.org/[...]
[267]
저널
The chaperone connection to the origins of the eukaryotic organelles.
[268]
저널
The genome sequence of Rickettsia prowazekii and the origin of mitochondria.
[269]
저널
Cultivation of the ubiquitous SAR11 marine bacterioplankton clade
[270]
저널
A robust species tree for the alphaproteobacteria.
http://jb.asm.org/cg[...]
[271]
저널
Warburgの「癌細胞の起原」に就いて
https://doi.org/10.4[...]
[272]
저널
ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis
[273]
저널
Mitochondrial defects trigger proliferation of neighbouring cells via a senescence-associated secretory phenotype in Drosophila (ショウジョウバエにおいて、ミトコンドリアの障害が、細胞老化関連分泌因子(SASP因子)を介して隣接細胞の増殖のトリガーになる)
https://www.nature.c[...]
2020-01-05
[274]
웹사이트
井垣達吏教授らの研究成果が、英国科学誌「Nature Communications」に掲載されました。
https://www.lif.kyot[...]
京都大学 大学院 生命科学研究科
2020-01-05
[275]
웹사이트
細胞間の相互作用で良性腫瘍ががん化する仕組みを解明
https://www.jst.go.j[...]
科学技術振興機構, 神戸大学
2020-01-05
[276]
웹인용
http://www.ncbi.nlm.[...]
[277]
저널
Evolutionary biology: Essence of mitochondria
[278]
웹인용
http://biology.plosj[...]
[279]
백과사전
미토콘드리아
글로벌 세계 대백과
[280]
문서
한국분자세포생물학회-미토콘드리아 품질관리기전과 노화관련 질병
http://www.ksmcb.or.[...]
[281]
저널
[282]
저널
[283]
저널
[284]
저널
The Hows and Whys of Cytoplasmic Inheritance in Seed Plants
[285]
저널
Paternal transmission of mitochondrial DNA is (fortunately) rare
2003
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com