데니소바인
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
데니소바인은 시베리아, 티베트, 라오스 등에서 발견된 멸종된 고대 인류이다. 2008년 러시아 데니소바 동굴에서 손가락뼈 화석이 발견되면서 존재가 처음 알려졌으며, 미토콘드리아 DNA 분석을 통해 현생 인류, 네안데르탈인과는 다른 새로운 종으로 분류되었다. 데니소바인은 네안데르탈인과 유전적으로 가깝고, 현생 인류와도 이종교배를 통해 유전자를 공유했다. 멜라네시아인, 오스트레일리아 원주민, 필리핀 아에타족 등은 데니소바인 DNA를 높은 비율로 가지고 있으며, 티베트인의 경우 데니소바인에게서 유래된 유전자가 고산 환경 적응에 기여했다.
더 읽어볼만한 페이지
- 아시아의 고고학 - 도유호
함경남도 함흥 출신 고고학자이자 정치인인 도유호는 오스트리아 빈 대학교에서 박사 학위를 받은 후 월북하여 김일성종합대학 교수로 재직하며 북한 고고학 연구를 주도했고, 최고인민회의 의장을 역임하며 《조선원시고고학》 등의 저서를 통해 북한 고고학 연구의 기초를 다지는 데 기여했다. - 알타이 변경주의 역사 - 카라수크 문화
카라수크 문화는 기원전 2300년에서 기원전 700년 사이 시베리아 남부에서 번성한 청동기 시대 문화로, 농업과 목축을 기반으로 움집에서 생활하며, 곡선형 칼과 마구를 특징으로 하는 청동기 제작 기술이 발달했다. - 알타이 변경주의 역사 - 아파나시에보 문화
아파나시에보 문화는 기원전 3300년경부터 기원전 2500년경까지 미누신스크 분지에서 준가리아에 이르는 지역에 존재했던 청동기 시대의 고고학 문화로서, 폰토스-카스피 스텝의 얌나야 문화와 유전적으로 가깝고 초기 목축 및 구리, 청동 야금술을 사용했으며 쿠르간 문화와의 연관성을 통해 초기 인도유럽어족과의 관련성이 제기된다. - 구석기 시대 아시아 - 호모 에렉투스
호모 에렉투스는 약 200만 년 전부터 10만 년 전까지 생존하며 직립 보행을 하고 아슐리안 석기를 제작했으며 불을 사용한 최초의 인류로 추정되는 종이다. - 구석기 시대 아시아 - 네안데르탈인
네안데르탈인은 유럽과 서아시아에 살았던 고대 인류로, 큰 뇌 용량과 독특한 신체적 특징을 지녔으며, 무스테리안 문화를 사용하고 동굴에서 거주하며 매장 의례를 치르는 등 복잡한 사회생활을 영위했지만, 현생 인류와의 경쟁, 기후 변화 등으로 약 4만 년 전 멸종한 종이며, 현생 인류와 이종교배하여 현생 인류 유전체에 일부 유전자가 남아있다.
| 데니소바인 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
| 학명 | Homo sp. |
| 영명 | Denisova hominin |
| 한글명 | 데니소바인 |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 목 | 영장목 |
| 아목 | 진원아목 |
| 상과 | 사람상과 |
| 과 | 사람과 |
| 아과 | 사람아과 |
| 족 | 사람족 |
| 아족 | 사람아족 |
| 속 | 사람속 |
| 특징 | |
| 시대 | 플라이스토세 |
| 지역 | 아시아 |
| 다른 인류와의 관계 | 네안데르탈인과 현생인류와 모두 관련이 있음 호모 에렉투스 와는 구별됨 |
| 유전적 특징 | 데니소바인 유전자는 현생 인류에게서 발견됨 특히 멜라네시아인, 오스트레일리아 원주민들에게서 높은 비율로 나타남 티베트인의 고지대 적응에 영향을 미치는 EPAS1 유전자 발견 |
| 발견 | |
| 최초 발견 장소 | 데니소바 동굴 (러시아 시베리아 남부) |
| 발견 연도 | 2008년 |
| 발견자 | 요하네스 크라우제와 스반테 파보 |
| 유물 | 손가락 뼈 조각 어금니 발가락 뼈 머리뼈 조각 |
| 추가 정보 | |
| 거주 추정 지역 | 시베리아 남부에서 동남아시아, 티베트 고원까지 확장되었을 가능성 |
| 문화 | 석기 제작 기술 사용 다른 지역의 석기 기술과 유사성 발견 |
| 연구 | 고인류학, 유전체학 연구 진행 중 네안데르탈인과의 혼혈 가능성 확인 |
| 참고 자료 | 칼 짐머 - 데니소바인 추적 기사 |
2. 발견 및 분류
2008년 7월, 시베리아 알타이산맥의 데니소바 동굴에서 발견된 손가락뼈, 치아 화석, 다리뼈 일부 화석에 대한 미토콘드리아 DNA 분석 결과, 2010년 3월 25일자 영국 과학 잡지 네이처에 현생 인류와 약 100만 년 전에 갈라진 새로운 계통의 인류라는 막스 플랑크 진화인류학 연구소 연구팀의 발표가 있었다.[77][78] DNA만으로 새로운 종의 인류가 발견된 것은 과학사상 처음 있는 일이다.[79]
2011년에는 데니소바 근처의 다른 동굴에서 4만 년 전의 네안데르탈인으로 추정되는 발가락 화석도 발견되었다.
2019년 1월 30일 네이처에 발표된 바에 따르면, 데니소바 동굴에서 발견된 데니소바 소녀의 DNA 분석, 탄소 측정, 100개 이상의 지층에서 28만 개의 사료 분석을 통한 OSL연대측정 결과, 데니소바 동굴에는 43만 년 전부터 인류가 살았던 것으로 추정된다. 43만 년 전부터 2~3만 년 전까지 간헐적으로 데니소바인이 살았고, 45만 년 전부터는 네안데르탈인이 살았으며, 어느 순간 네안데르탈인과 데니소바인이 간헐적으로 동굴을 교차 사용했으며, 13만 년 전에 최초로 네안데르탈인 어머니와 데니소바인 아버지를 둔 아이가 태어났다. 2~3만 년 전을 기점으로 데니소바인은 사라지고, 4만 9000년~4만 3000년 사이에 후기구석기 유물이 발견되었는데, 이는 현생인류만이 남겼다고 알려져 있다.
2019년 4월 11일 학술지 Cell에 발표된 논문에 따르면, 데니소바인은 3개의 독립적인 그룹으로 나뉘며, 이 중 한 그룹은 네안데르탈인과 데니소바인의 차이만큼 다른 두 그룹과 다르다.[80]
파푸아뉴기니와 솔로몬 제도 등 일부 멜라네시아인에게서 데니소바인의 유전자가 발견되면서 유전자 존재 여부에 대한 논란이 일기도 했다.
현생인류, 네안데르탈인, 호모 솔로엔시스 외에 시베리아에 존재했던 미지의 인류라는 점에서 한때 화제가 되었다. 네안데르탈인과 그들의 공통 조상으로 추정되는 호모 하이델베르켄시스가 가까운 근연종이다. 2013년 12월 4일에는 에스파니아 시마 동굴에서 발견된 일부 호모 하이델베르켄시스 유골에서 데니소바인의 특징이 나타나기도 했다.[95]
데니소바인은 새로운 종의 ''호모'' 또는 ''호모 사피엔스''(현생 인류)의 고대 아종일 수 있지만, 적절한 분류군을 정할 만큼 화석이 충분하지 않다. ''H. denisova''[2] 또는 ''H. altaiensis''[3]가 데니소바인에 대해 제안된 종명이다. 중국 연구자들은 데니소바인이 ''호모 롱이''의 구성원이라고 제안했고, 고생물학자 크리스 스트링거가 이를 지지했다.[4] 2024년, 고인류학자 크리스토퍼 베이와 쉬지에 우는 쉬자야오 인류 화석을 ''호모 줄루엔시스'' 종의 표준 표본으로 지정하고, 데니소바인을 이 종으로 통합할 것을 제안했다.[5]
2. 1. 발견
2008년 7월, 시베리아의 알타이산맥에 위치한 데니소바 동굴에서 30~50세 가량으로 추정되는 손가락뼈, 치아 화석 및 다리뼈 일부 화석이 처음 발견되었다.[94] 미토콘드리아 DNA 검사 결과 현생인류와는 다른 점이 발견되었으며, 네안데르탈인과 가까운 점이 발견되었다. 처음에 동굴곰의 뼈로 인식했던 새끼손가락뼈는 6~7살 정도의 여자 어린이의 것으로 확인되었다. 다리뼈 일부와 어금니 화석은 성인의 것으로 밝혀졌다. 유전자 검사 결과 미토콘드리아 DNA에 따르면 네안데르탈인과 현생 인류와 관련이 있으며 공통 조상은 약 100만년 전에 있었던 것으로 확인되었다. 호모 하이델베르겐시스의 후손이자 네안데르탈인과 근연종으로 추정되며, 네안데르탈인, 현생인류와 동시대에 산 것으로 보이며 서로 교배도 한 것으로 보인다.2019년에는 티베트고원 바이시야 카르스트 동굴(Baishiya Karst Cave)에서 시베리아 데니소바 동굴 밖의 데니소바인이 처음으로 발견되었다. 이 아래턱 화석은 1980년에 발견되었으나 2019년에 이르러서야 DNA 분석 결과 데니소바인 화석인 것으로 확인되었다. 이 화석은 데니소바인이 아시아에 폭넓게 거주했다는 설과 티베트의 고산 적응 유전자가 데니소바인에서 유래했다는 설을 뒷받침하는 증거이다.

데니소바 동굴은 러시아 알타이 크라이에 위치하며, 시베리아 남중부, 알타이 산맥 서쪽 가장자리에 있다. 이 동굴은 18세기에 그곳에 살았던 러시아 구교도 은둔자 데니스(디오니시)의 이름을 따서 명명되었다.
2019년, 그리스 고고학자 카테리나 두카와 동료들은 데니소바 동굴의 표본을 방사성탄소 연대 측정하여 데니소바 2(가장 오래된 표본)가 19만 5000년~12만 2700년 전에 살았던 것으로 추정했다.[9] 동굴에서 발견된 유물을 바탕으로, 호미닌 거주(아마도 데니소바인에 의한)는 28만 7000±41년 또는 20만 3000±14년 전에 시작되었을 것으로 추정된다. 네안데르탈인 또한 19만 3000±12년 전과 9만 7000±11년 전에 존재했는데, 아마도 데니소바인과 동시에 존재했을 가능성이 있다.[48]
2018년, 안남 산맥(Annamite Mountains) 라오스 정글의 동굴들을 발굴해 온 라오스, 프랑스, 미국 인류학자팀이 현지 아이들의 안내를 받아 탐 응우 하오 2 동굴("코브라 동굴")에서 인간의 치아를 발견했다. 이 치아는 아마도 16만 4천 년 전에서 13만 1천 년 전 사이의 것으로 추정된다.[15]
| 이름 | 화석 부위 | 연대 | 발견 연도 | 발견 장소 | 성별 및 연령 | 발표 연도 | 이미지 | GenBank 접근 번호 |
|---|---|---|---|---|---|---|---|---|
| 데니소바 3 (또는 X 여성)[24][18][8] | 약손가락의 원위 지골 | 76,200–51,600년 전[9] | 2008 | 데니소바 동굴 (러시아) | 13.5세 소녀 | 2010 |  | [https://www.ncbi.nlm.nih.gov/nuccore/NC013993 NC013993] |
| 데니소바 4[24][20] | 영구 상악 제2 또는 제3 대구치 | 84,100–55,200년 전[9] | 2000 | 데니소바 동굴 (러시아) | 성인 남성 | 2010 | 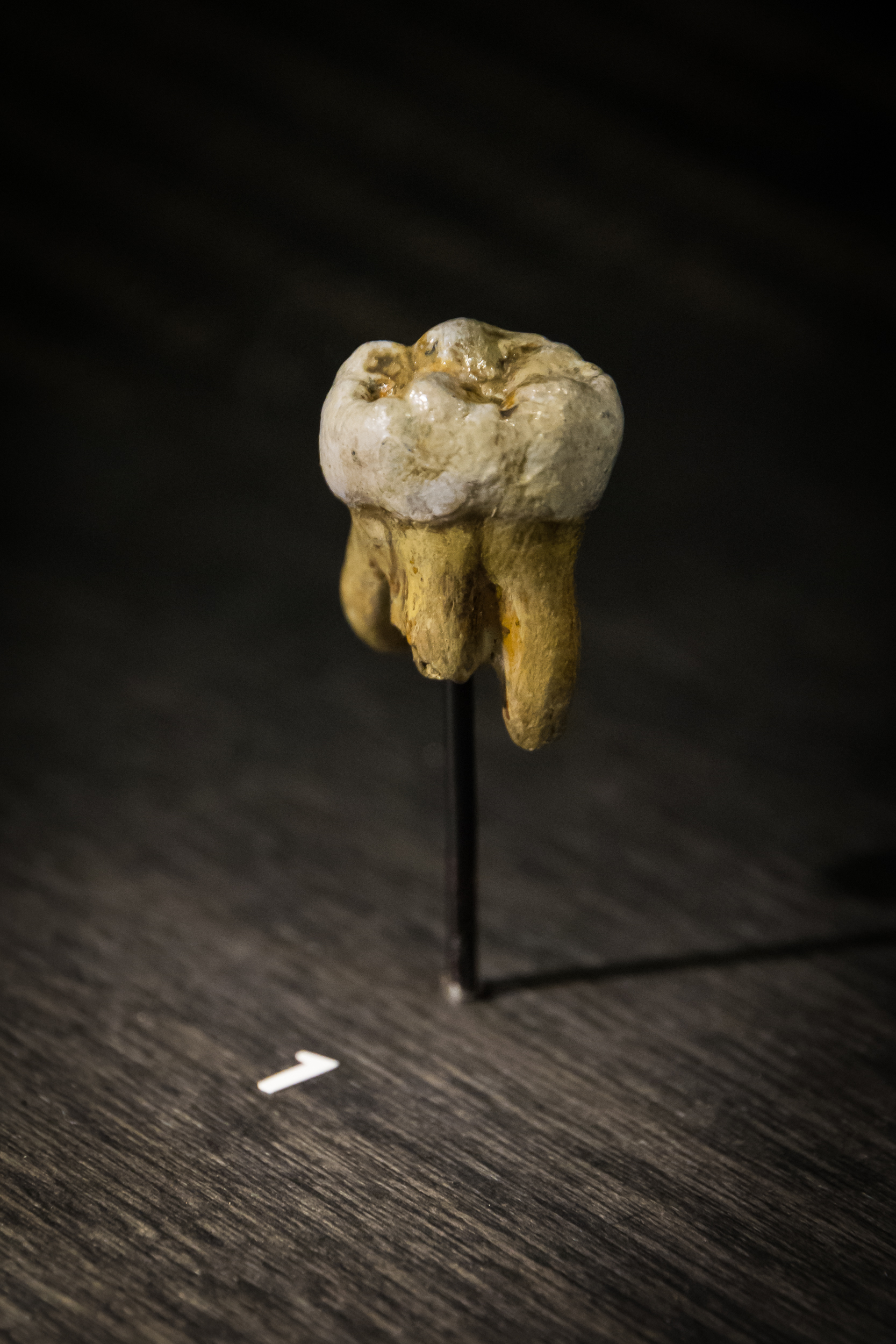 | [https://www.ncbi.nlm.nih.gov/nuccore/FR695060 FR695060] |
| 데니소바 8[11] | 영구 상악 제3 대구치 | 136,400–105,600년 전[9] | 2010 | 데니소바 동굴 (러시아) | 성인 남성 | 2015 | [https://www.ncbi.nlm.nih.gov/nuccore/KT780370 KT780370] | |
| 데니소바 2[21] | 유치 하악 제2 대구치 | 194,400–122,700년 전[9] | 1984 | 데니소바 동굴 (러시아) | 소녀 | 2017 | [https://www.ncbi.nlm.nih.gov/nuccore/KX663333 KX663333] | |
| 하허 하악골(Xiahe mandible)[13] | 하악골 일부 | 16만 년 이상 | 1980 | 바이샤 동굴(Baishiya Cave) (중국) | 2019 |  | ||
| 데니소바 11 (또는 데니, 데니소바인 × 네안데르탈인 잡종)[22] | 팔 또는 다리뼈 조각 | 118,100–79,300년 전[9] | 2012 | 데니소바 동굴 (러시아) | 13세 소녀 | 2016 |  | |
| 데니소바 13[23] | 두정골 조각 | 22층에서 발견[23] (약 285,000 ± 39,000년 전)[48] | 2019 | 데니소바 동굴 (러시아) | 미정 | |||
| TNH2-1[15] | 영구 하악 좌측 제1 또는 제2 대구치 | 164,000–131,000년 전 | 2018 | 탐 응우 하오 2 동굴 (라오스) | 3.5~8.5세 소녀 | 2022 | 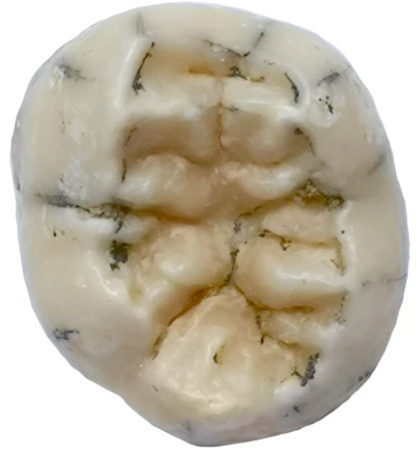 | |
| BSY-19-B896-1 (하허 2) | 원위 갈비뼈 조각 | 48,000–32,000년 전 | 1980 | 바이샤 동굴 (중국) | 불명 | 2024 |  | |
| 데니소바 25[10] | 대구치 | 20만 년 전 | 2024 | 데니소바 동굴 (러시아) | 남성 | 미정 |
2. 2. 분류
데니소바인은 새로운 종의 ''호모'' 또는 ''호모 사피엔스''(현생 인류)의 고대 아종을 나타낼 수 있지만, 적절한 분류군을 설정할 만큼 화석이 충분하지 않다. 데니소바인에 대해 적극적으로 제안된 종명으로는 ''H. denisova''[2] 또는 ''H. altaiensis''[3]가 있다. 중국 연구자들은 데니소바인이 ''호모 롱이''의 구성원이라고 제안했고, 고생물학자 크리스 스트링거가 이러한 견해를 지지했다.[4] 2024년, 고인류학자 크리스토퍼 베이와 쉬지에 우는 쉬자야오 인류 화석을 ''호모 줄루엔시스'' 종의 표준 표본으로 지정하고, 데니소바인을 이 종으로 통합할 것을 제안했다.[5]3. 유전적 특징 및 진화
2010년 12월 23일, 막스 플랑크 진화인류학 연구소 등의 국제 연구팀은 네이처에 발표한 논문에서 데니소바인이 네안데르탈인과 근연 관계에 있으며, 80만 4천 년 전 현생 인류인 호모 사피엔스와의 공통 조상으로부터 네안데르탈인·데니소바인의 조상이 분기하고, 64만 년 전에 네안데르탈인으로부터 분기한 인류라고 추정했다.[83] 데니소바 동굴은 네안데르탈인 화석 발견지 중 가장 가까운 이라크 북부 샤니다르 유적과 약 4000km 떨어져 있다.
멜라네시아인 게놈의 4~6%가 데니소바인 고유의 것과 일치하며,[84] 스웨덴 웁살라 대학교 연구팀은 중국 남부 거주민 유전자 구조의 약 1%가 데니소바인 유래라는 연구 결과를 발표했다.[85] 네안데르탈인과 분기한 시기는 약 35만 년 전이라는 설도 제기된다.[76]
네안데르탈인과 데니소바인은 멸종했지만, 네그로이드를 제외한 현생 인류 유전자의 수 %는 네안데르탈인에게서 유래했다. 2010년 5월에는 중동에서 현생 인류 조상과 네안데르탈인의 혼혈을 보여주는 연구 결과가 발표되었고, 12월에는 아시아 내륙부에서 데니소바인과 현생 인류 조상이 혼혈했다는 연구 결과가 나왔다. 이는 과거 이종의 인류 조상 간 혼혈·공존이 일반적인 일이었을 가능성을 보여준다.[86]
아시아 내륙부에서 데니소바인과 혼혈한 현생 인류 조상은 오랜 시간에 걸쳐 멜라네시아 등으로 남하했고, 중국으로 이주한 집단은 한족이, 고지로 이주한 집단은 티베트인이 되었다고 추정된다.[87]
2018년 8월 22일, 과학 저널 「네이처」는 2012년 러시아에서 발견된 10대 소녀의 화석이 네안데르탈인 어머니와 데니소바인 아버지 사이에서 태어났다는 사실을 발표했다.[89][90][91]
2019년 9월, 히브리 대학교와 스탠퍼드 대학교의 데이비드 고크만은 DNA 메틸화를 조사하여 골격에 관한 32가지 특징을 추출, 데니소바인의 골격을 제안했다.[92][93] 이 연구에 따르면, 데니소바인은 좁은 이마와 튼튼한 턱 등 네안데르탈인과 매우 유사한 특징을 가졌을 가능성이 있지만, 두정골 폭이 넓은 등 다른 특징도 보인다.[79]
3. 1. 유전적 특징
Denisovan영어의 미토콘드리아 DNA(mtDNA)는 현생인류 및 네안데르탈인의 mtDNA와 상당한 차이를 보인다.[77][78] 2010년 발표된 내용에 따르면, 세포핵 DNA 분석 결과, 데니소바인은 네안데르탈인과 가까운 관계이며, 현생인류와는 약 80만 4천 년 전에 분기된 것으로 추정되었다.[83] 이들은 네안데르탈인과 64만 년 전에 분기하였다.[83]2019년 4월 11일자 학술지 『Cell』에 발표된 논문에 따르면, 데니소바인에게는 독립적인 3개의 그룹이 존재하며, 이 그룹 중 하나는 네안데르탈인과 데니소바인의 차이만큼 다른 두 그룹의 데니소바인과 다르다는 것이 시사되고 있다.[80]
데니소바인 게놈에는 검은 피부, 갈색 머리카락, 갈색 눈과 관련된 유전자 변이가 존재한다. 또한 티베트인의 고산 적응에 관여하는 EPAS1 유전자 주변의 변이 영역이 데니소바인 게놈에도 존재한다.[94]
3. 2. 진화
데니소바인은 아프리카에서 이주한 초기 호모 에렉투스의 후손으로 추정되며, 호모 하이델베르겐시스의 후예이자 네안데르탈인의 근연종으로 여겨진다.미토콘드리아 DNA(mtDNA) 분석에 따르면, 데니소바인은 현생 인류 및 네안데르탈인과 약 100만 년 전에 공통 조상에서 갈라졌다. 그러나 핵 DNA(nDNA) 분석 결과, 데니소바인과 네안데르탈인은 현생 인류보다 서로 더 가까운 관계이며, 약 80만 4천 년 전에 현생 인류와 갈라졌고, 64만 년 전에 서로 분리되었다.[24][83] 염기쌍(bp)당 연간 돌연변이율과 세대당 돌연변이율을 고려하면, 네안데르탈인/데니소바인의 분리는 약 23만~74만 4천 년 전에 일어났을 것으로 추정된다.[28][25]
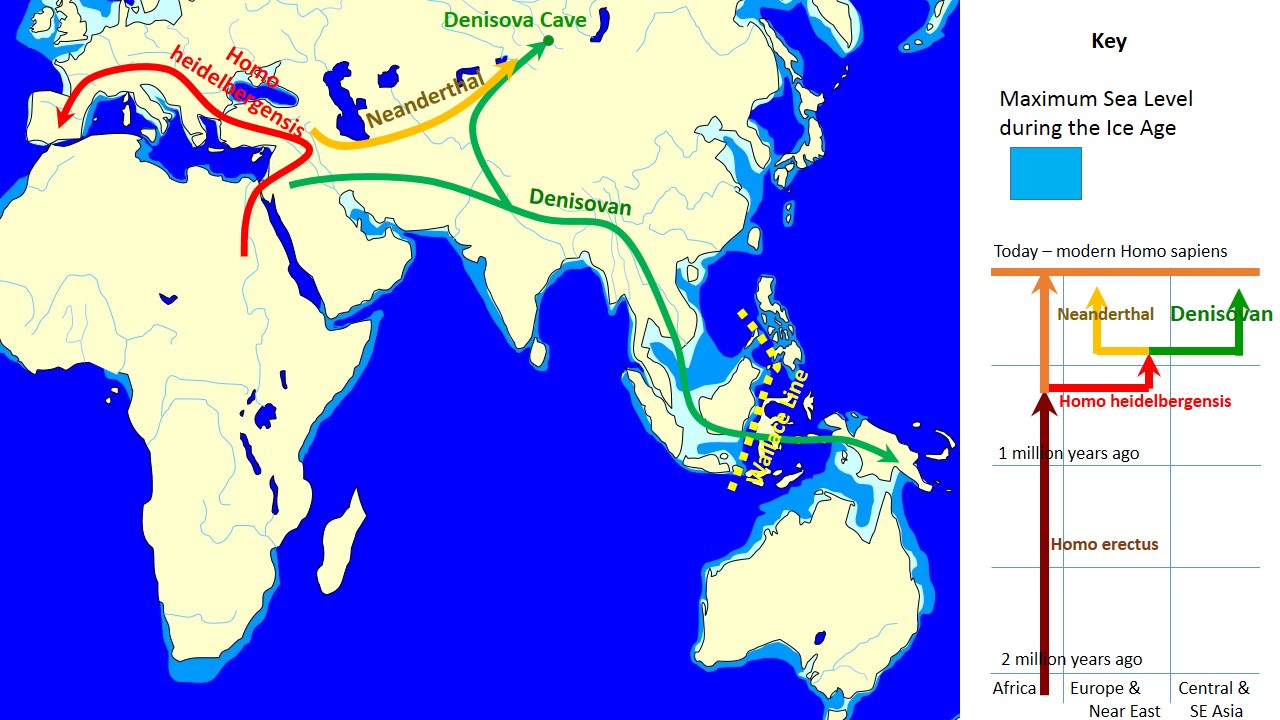
막스 플랑크 진화인류학 연구소의 연구에 따르면, 데니소바인은 독립적인 3개의 그룹으로 구성되어 있었으며, 이들 간에는 상당한 유전적 차이가 존재했다.[80] 이는 데니소바인이 여러 지역에 걸쳐 고립된 채 진화했음을 시사한다.
데니소바인은 네안데르탈인과 마찬가지로 멸종했지만, 현생 인류의 일부(특히 멜라네시아인, 애보리지니, 중국 남부 거주민)에게 유전자를 남겼다.[84][85] 이는 과거 이종 간의 혼혈이 빈번했음을 보여주는 증거이다.[86]
2018년에는 데니소바 동굴에서 네안데르탈인 어머니와 데니소바인 아버지 사이에서 태어난 10대 소녀의 화석이 발견되어, 두 인류 간의 혼혈이 확인되었다.[89][90][91]
4. 분포 및 생활상
2008년 7월, 시베리아의 알타이산맥에 위치한 데니소바 동굴에서 30~50세 가량으로 추정되는 손가락뼈, 치아 화석 및 다리뼈 일부 화석이 처음 발견되었다.[94] 미토콘드리아 DNA 검사 결과 현생인류와는 다른 점이 발견되었으며, 네안데르탈인과 가까운 점이 발견되었다. 유전자 검사 결과에 따르면 네안데르탈인과 현생 인류와 관련이 있으며 공통조상은 약 100만년 전에 있었던 것으로 확인되었다. 호모 하이델베르겐시스의 후예이자 네안데르탈인과 근연종으로 추정되며, 이들은 네안데르탈인, 현생인류와 동시대에 살았고 서로 교배도 한 것으로 보인다.
기원전 6만년 경 이후 유라시아로 이동한 현생인류 중에 데니소바인이 흡수되었고, 동남아시아로 이주한 것으로 추정된다. 라오스 북부의 한 동굴에서 164,000년에서 131,000년 전에 사망한 것으로 보이는 데니소바인 소녀의 것으로 추정되는 어금니들이 발견되었다.[94] 시베리아에 남아있던 데니소바인은 2~3만년 전에 사멸된 것으로 추정된다. 현재 멜라네시아인들과 오스트레일리아 원주민인 애보리지니들의 DNA 중 6%가 이들로부터 유래한 것으로 보인다.
2019년 티베트고원 바이샤 카르스트 동굴(Baishiya Karst Cave)에서 시베리아 데니소바 동굴 밖의 데니소바인이 처음으로 발견되었다. 이 아래턱 화석은 1980년에 발견되었으나 2019년에 이르러서야 DNA 분석 결과 데니소바인 화석인 것으로 확인되었다.
데니소바 동굴은 데니소바인이 살았던 시기 동안 따뜻하고 습한 소나무, 자작나무 숲에서 툰드라, 숲-툰드라 지형으로 환경이 변화했다.[48] 바이샤 카르스트 동굴은 티베트 고원의 고지대에 위치하여 춥고 산소가 부족한 환경이었는데, 데니소바인은 이러한 혹독한 환경에 적응했을 것으로 보인다.[45][13] 데니소바인 게놈에는 고지대의 저산소 환경 적응에 유리한 EPAS1 유전자 변이가 있는데,[45][13] 이는 티베트인에게서도 발견된다. WARS2와 TBX15 유전자좌 영역에는 이누이트의 체지방 분포에 영향을 주는 변이가 있다.[46] 라오스의 탐 응우 하오 2 동굴(코브라 동굴)에서 발견된 데니소바인의 치아는 이들이 동남아시아의 정글에도 서식했음을 보여준다.[15]
티베트 퀘상 온천 근처에서는 데니소바인 어린이가 남긴 것으로 추정되는 손자국과 발자국이 발견되었는데,[51] 이는 전에서 전의 것으로 가장 오래된 암벽화의 예로 간주될 수 있다.[51] 발자국은 평균 192.3mm로 약 7~8세 어린이에 해당하며, 손자국은 평균 161.1mm로 12세 어린이에 해당한다.[51]
4. 1. 분포
데니소바인은 시베리아, 티베트, 라오스 등지에 서식했던 것으로 알려져 있다.[15] 현생인류의 데니소바인 DNA 흔적은 동아시아 전역,[36][60] 오세아니아, 그리고 잠재적으로 서유라시아[69]까지 분포했음을 시사한다.2019년, 유전학자 가이 제이콥스(Guy Jacobs)와 동료들은 현대 인류에게 유전자 혼합을 일으킨 세 개의 독립적인 데니소바인 집단을 확인했는데, 각각 시베리아와 동아시아, 뉴기니와 인근 섬들, 그리고 오세아니아와 아시아 전역(다소 적은 정도)에 원주민으로 살고 있는 집단이다. 연합 모델링을 사용하여, 데니소바 동굴의 데니소바인은 두 번째 집단과 약 28만 3천 년 전, 세 번째 집단과 약 36만 3천 년 전에 분리되었다. 이는 데니소바인 집단 간에 상당한 생식적 격리가 있었음을 나타낸다.[37]
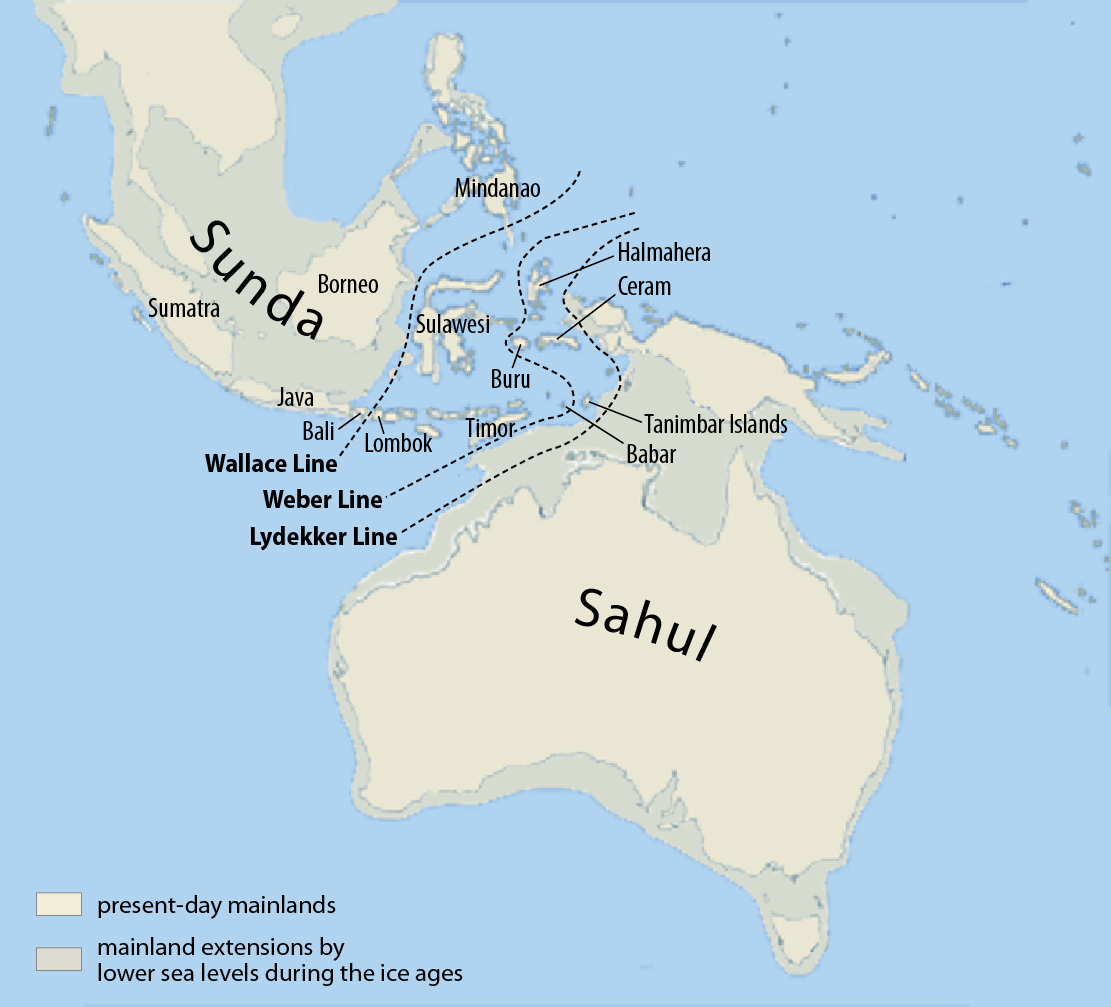
현대 파푸아인과 호주 원주민에게서 높은 비율의 데니소바인 DNA가 발견된 것을 바탕으로, 데니소바인은 왈라세선을 넘어 이동했을 가능성이 있으며,[39] 이는 그 이전의 ''호모 플로레시엔시스''에 이어 두 번째로 그러한 이동을 한 인종이다. 이러한 논리에 따르면, 그들은 필리핀에도 들어가 ''H. 루손엔시스''와 함께 살았을 수 있는데, 만약 그렇다면 이는 동일하거나 밀접하게 관련된 종일 수 있다.[64]
지수 분포 분석을 해플로타입 길이에 적용하여, 제이콥스는 현대 인류에 대한 유전자 유입이 뉴기니인의 조상이 되는 데니소바인 집단과는 약 2만 9900년 전에, 뉴기니인과 오세아니아인의 조상이 되는 집단과는 약 4만 5700년 전에 발생했음을 계산했다. 뉴기니 집단의 이렇게 늦은 시기는 데니소바인이 최대 1만 4500년 전까지 생존했을 수 있음을 시사하는데, 이는 그들을 가장 최근까지 생존한 구인류로 만든다.[37]
4. 2. 생활상
데니소바 동굴은 데니소바인이 살았던 시기 동안 따뜻하고 습한 소나무, 자작나무 숲에서 툰드라, 숲-툰드라 지형으로 환경이 변화했다.[48] 바이샤 카르스트 동굴은 티베트 고원의 고지대에 위치하여 춥고 산소가 부족한 환경이었는데, 데니소바인은 이러한 혹독한 환경에 적응했을 것으로 보인다.[45][13] 데니소바인 게놈에는 고지대의 저산소 환경 적응에 유리한 EPAS1 유전자 변이가 있는데,[45][13] 이는 티베트인에게서도 발견된다. 또한 WARS2와 TBX15 유전자좌 영역에는 이누이트의 체지방 분포에 영향을 주는 변이가 있다.[46] 라오스의 탐 응우 하오 2 동굴(코브라 동굴)에서 발견된 데니소바인의 치아는 이들이 동남아시아의 정글에도 서식했음을 보여준다.[15]데니소바 동굴에서는 중기 구석기 시대와 상기 구석기 시대의 다양한 석기들이 발견되었다. 초기 중기 구석기 시대 석기는 핵석, 긁개, 톱니형 석기, 노치된 석기 등이며,[48] 중기 구석기 시대 유물은 르발루아 핵석이 주를 이루었고, 측면 긁개, 노치된 톱니형 석기, 끌과 같은 도구도 있었다.[48] 초기 상기 구석기 시대 유물은 석판 생산과 르발루아 생산이 있었지만, 긁개가 다시 주를 이루었다.[48] 상기 구석기 시대 지층에서는 뼈 도구와 대리석 반지, 상아 펜던트, 붉은사슴 이빨 펜던트, 녹렴석 팔찌, 뼈 바늘 등 장신구도 발견되었다.[49][48] 그러나 이러한 유물들이 현생인류와 관련이 있는지는 불분명하다.[49][48]

바이샤 석회암 동굴에서는 염소과 동물, 소, 사슴, 말, 털코뿔소 등 다양한 동물 뼈가 발견되었으며, 큰 육식동물, 마멋, 토끼, 독수리의 뼈도 발견되었다.[50] 이들은 긴 뼈를 사용하여 뼈 도구를 만들었을 수 있으며, 석기 유물도 발견되었다.[50]
티베트 퀘상 온천 근처에서는 데니소바인 어린이가 남긴 것으로 추정되는 손자국과 발자국이 발견되었는데,[51] 이는 전에서 전의 것으로 가장 오래된 암벽화의 예로 간주될 수 있다.[51] 발자국은 평균 192.3mm로 약 7~8세 어린이에 해당하며, 손자국은 평균 161.1mm로 12세 어린이에 해당한다.[51]
5. 다른 인류와의 관계
2008년 시베리아의 알타이산맥에 위치한 데니소바 동굴에서 발견된 화석을 통해 처음 알려진 데니소바인은, 미토콘드리아 DNA 검사 결과 네안데르탈인 및 현생 인류와는 다른 계통의 인류로 밝혀졌다. 이들은 약 100만 년 전에 현생 인류와 공통 조상에서 갈라졌으며, 네안데르탈인과는 약 64만 년 전에 분기한 것으로 추정된다.[83]
데니소바인은 현생 인류와도 교배한 것으로 보이는데, 특히 멜라네시아인과 애보리지니의 DNA에서 데니소바인의 유전자가 발견된다. 이는 데니소바인이 기원전 6만 년경 유라시아로 이동한 현생 인류에게 흡수되었고, 일부는 동남아시아로 이주했음을 시사한다. 실제로 라오스 북부 동굴에서 발견된 어금니는 시베리아 밖에서 발견된 두 번째 데니소바인 화석이다.[94] 파푸아뉴기니와 솔로몬 제도 등지에 거주하는 일부 멜라네시아인에게서 데니소바인의 유전자가 발견된 것은, 현생 인류가 아프리카에서 이주해 나오면서 데니소바인과 만나 유전자를 교환했음을 보여준다.
5. 1. 네안데르탈인과의 관계
2010년 미토콘드리아 DNA 검사 결과, 데니소바인은 네안데르탈인과 가까운 관계이며, 호모 하이델베르겐시스의 후손이자 네안데르탈인과 근연종으로 추정된다. 이들은 네안데르탈인 및 현생 인류와 동시대에 살았으며, 서로 교배하기도 했다.[94] 2013년 12월 4일에는 에스파니아의 시마 동굴에서 발견된 일부 호모 하이델베르켄시스의 유골에서 데니소바인의 특징을 가진 유골이 존재한다는 것이 확인되었다.[95]2018년 국제 학술지 네이처에는 네안데르탈인과 데니소바인 두 인류종 간의 1세대 혼혈 개체가 데니소바 동굴에서 처음 발견되었다는 보고가 발표되었다. 2019년 네이처 1월 30일자 발표에 따르면, 데니소바 동굴에서 발견된 데니소바 소녀의 DNA 분석과 탄소 측정, 그리고 100개 이상의 지층에서 28만 개의 사료 분석을 통해 OSL 연대 측정을 한 결과, 13만 년 전에 네안데르탈인 어머니와 데니소바인 아버지를 둔 아이가 태어났다는 사실이 밝혀졌다. 데니소바 동굴에서 데니소바인과 네안데르탈인 1대1 혼혈 화석이 발견되어 혼혈은 확실한 것으로 확정되었다.
2019년 4월 11일자 학술지 『Cell』에 발표된 논문에 따르면, 데니소바인에게는 독립적인 3개의 그룹이 존재하며, 이 그룹 중 하나는 네안데르탈인과 데니소바인의 차이만큼 다른 두 그룹의 데니소바인과 다르다는 것이 시사되고 있다.[80]
2010년 12월 23일, 막스 플랑크 진화인류학 연구소 등의 국제 연구팀은 『네이처』에 게재한 논문에서, 세포핵 DNA 분석 결과 데니소바인이 80만 4천 년 전 현생 인류인 호모 사피엔스와의 공통 조상으로부터 네안데르탈인·데니소바인의 조상이 분기하고, 64만 년 전에 네안데르탈인으로부터 분기한 인류라고 추정했다.[83] 데니소바 동굴은 네안데르탈인 화석 발견지 중 가장 가까운 이라크 북부 샤니다르 유적과 약 4000km 떨어져 있다.
현생 인류 게놈 분석 결과, 적어도 두 종류의 구인류, 네안데르탈인[52]과 데니소바인[24][53]과의 과거 이종교배가 있었고, 이러한 이종교배 사건이 여러 차례 발생했음을 알 수 있다. 데니소바인, 네안데르탈인, 현생 인류의 게놈 비교를 통해 이들 계통 간의 복잡한 이종교배 네트워크의 증거가 밝혀졌다.[52]
데니소바 동굴에서 발견된 데니소바인 게놈의 최대 17%가 현지 네안데르탈인 집단의 DNA를 나타낸다.[52] 데니소바 11은 데니소바인/네안데르탈인 잡종 1세대(F1) 개체였다. 이러한 개체가 발견되었다는 사실은 이곳에서 이종교배가 흔한 일이었음을 시사한다. 데니소바인 게놈은 크로아티아의 빈디야 동굴(Vindija Cave) 네안데르탈인 게놈이나 코카서스의 메즈마이스카야 동굴(Mezmaiskaya cave) 네안데르탈인 게놈보다 시베리아의 알타이 네안데르탈인 게놈과 더 많은 유도 대립 유전자를 공유하는데, 이는 유전자 흐름이 현지 알타이 네안데르탈인과 더 밀접한 관련이 있는 집단에서 비롯되었음을 시사한다.[56] 그러나 데니의 데니소바인 아버지는 전형적인 알타이 네안데르탈인의 유전자 혼입(introgression)을 가지고 있었던 반면, 그녀의 네안데르탈인 어머니는 빈디야 네안데르탈인과 더 밀접한 관련이 있는 집단을 나타냈다.[55] 20만 년 전 것으로 추정되는 데니소바 25는 이전에 알려지지 않은 네안데르탈인 집단으로부터 게놈의 5%를 물려받았으며, 더 어린 표본들과는 다른 데니소바인 집단에서 유래한 것으로 추정된다.[10]
2018년 8월 22일, 고유전학자 비비안 슬론은 2012년 러시아에서 발견된 10대 소녀의 화석이 어머니가 네안데르탈인, 아버지가 데니소바인이었다고 과학 저널 「네이처」에서 발표했다.[89][90][91]
5. 2. 현생인류와의 관계
현생 인류 게놈 분석 결과, 네안데르탈인[52]과 데니소바인[24][53]과 같은 구인류와의 이종교배가 여러 차례 발생했음이 밝혀졌다. 데니소바인, 네안데르탈인, 현생 인류의 게놈을 비교한 결과, 이들 계통 사이에 복잡한 이종교배가 있었음을 알 수 있다.[52]멜라네시아인 유전체의 약 4~6%[24] 또는 1.9~3.4%[62]가 데니소바인 유전자에서 유래한다. 2021년 연구에서는 필리핀의 아에타족에게서 파푸아인보다 30~40% 더 많은 데니소바인 조상 유전자를 발견했는데, 이는 유전체의 약 5%에 해당한다. 루손 섬의 아에타 마그부콘족은 세계 어느 인구 집단보다 데니소바인 조상 유전자 비율이 가장 높다.[64]
반면, 아시아 본토인과 아메리카 원주민에게서는 데니소바인 조상 유전자가 0.2% 유래한다.[65] 남아시아인은 동아시아인과 유사한 수준의 데니소바인 혼합 유전자를 가지고 있는 것으로 밝혀졌다.[67] 2018년 한족, 일본인, 다이족 유전체 연구에 따르면 현대 동아시아인은 두 개의 다른 데니소바인 집단에서 유래한 DNA를 가지고 있다. 하나는 파푸아인 유전체에서 발견된 데니소바인 DNA와 유사하고, 다른 하나는 데니소바 동굴의 데니소바인 유전체와 더 가깝다. 이는 두 개의 다른 데니소바인 집단과 관련된 두 개의 별개의 유전자 혼입 사건을 나타낼 수 있다. 남아시아 유전체에서는 파푸아인에서 관찰되는 것과 같은 단일 데니소바인 유전자 혼입에서만 DNA가 유래했다.[67] 2019년 연구에서는 동아시아인에게 혼입된 세 번째 데니소바인 집단을 발견했다.[37]
데니소바인 유전자는 아프리카에서 이주한 초기 현생 인류가 기후에 적응하는 데 도움이 되었을 수 있다. 고산 환경에서 저산소 환경에서 생활할 수 있게 해주는 현대 티베트인의 EPAS1 하프로타입은 데니소바인에서 유래했을 가능성이 높다.[45][13]
5. 3. 알려지지 않은 고대 인류와의 관계
데니소바인 게놈의 약 4%는 확인되지 않은 초기 인류(archaic hominin)에서 유래하는데, 아마도 변칙적인 고대 mtDNA의 원천일 것이며, 이 종은 100만 년 전에 네안데르탈인과 현생 인류로부터 갈라졌음을 나타낸다.[52] 후기 플라이스토세(Late Pleistocene) 아시아에서 확인된 유일한 호모 속(Homo) 종은 *H. erectus*와 *H. heidelbergensis*이다.[56][57] 그러나 2021년에 후자의 종으로 분류된 표본은 *H. longi*와 *H. daliensis*로 재분류되었다.[58]네안데르탈인의 조상("네안데르소반")이 유럽으로 이주하면서 이미 그곳에 존재했던 확인되지 않은 "초고대(superarchaic)" 인류 종과 이종교배를 했던 것으로 보인다. 이 초고대 인류는 약 190만 년 전 아프리카에서 매우 초기의 이주로부터 내려온 후손이었다.[59]
6. 한국과의 관련성 (추가)
한국인의 유전체 분석 결과에서는 데니소바인의 유전적 영향이 미미한 것으로 나타난다. 이는 데니소바인이 주로 멜라네시아 등 동남아시아와 오세아니아 지역으로 확산되었고, 한반도를 포함한 동북아시아 지역에는 큰 영향을 미치지 않았음을 시사한다. 다만, 스웨덴 웁살라 대학교 연구팀의 연구 발표에 따르면 중국 남부 거주민의 유전자 구조의 약 1%가 데니소바인에게서 유래했다고 한다.[85] 또한, 중국 쪽으로 이주한 집단은 한족이 되고, 고지로 이주한 집단은 티베트인이 되었다고도 한다.[87]
참조
[1]
뉴스
On the Trail of the Denisovans - DNA has shown that the extinct humans thrived around the world, from chilly Siberia to high-altitude Tibet — perhaps even in the Pacific islands.
https://www.nytimes.[...]
2024-03-02
[2]
서적
Exploring Human Biology in the Laboratory
https://books.google[...]
Morton Publishing Company
2020-10-25
[3]
간행물
The Morphology of Permanent Molars from the Paleolithic Layers of Denisova Cave
https://www.research[...]
[4]
뉴스
Scientists link elusive human group to 150,000-year-old Chinese 'dragon man'
https://www.theguard[...]
2024-03-30
[5]
서적
The Paleoanthropology of Eastern Asia
University of Hawai‘i Press
[6]
간행물
A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum
[7]
서적
Who We Are and How We Got Here
Oxford University Press
[8]
간행물
The complete mitochondrial DNA genome of an unknown hominin from southern Siberia
[9]
간행물
Age estimates for hominin fossils and the onset of the Upper Palaeolithic at Denisova Cave
https://ro.uow.edu.a[...]
2019
[10]
간행물
The most ancient human genome yet has been sequenced—and it's a Denisovan's
https://www.science.[...]
2024-07-11
[11]
간행물
Nuclear and mitochondrial DNA sequences from two Denisovan individuals
[12]
간행물
First fossil jaw of Denisovans finally puts a face on elusive human relatives
[13]
간행물
A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau
https://kar.kent.ac.[...]
[14]
간행물
Denisovan DNA in Late Pleistocene sediments from Baishiya Karst Cave on the Tibetan Plateau
[15]
간행물
A Middle Pleistocene Denisovan molar from the Annamite Chain of northern Laos
[16]
간행물
An updated age for the Xujiayao hominin from the Nihewan Basin, North China: Implications for Middle Pleistocene human evolution in East Asia
[17]
간행물
Biggest Denisovan fossil yet spills ancient human's secrets
[18]
간행물
Morphology of the Denisovan phalanx closer to modern humans than to Neanderthals
[19]
간행물
Late Middle Pleistocene Harbin cranium represents a new Homo species
2021-06-25
[20]
간행물
Fossil genome reveals ancestral link
[21]
간행물
A fourth Denisovan individual
[22]
간행물
Identification of a new hominin bone from Denisova Cave, Siberia using collagen fingerprinting and mitochondrial DNA analysis
[23]
간행물
A parietal fragment from Denisova cave
https://meeting.phys[...]
[24]
간행물
Genetic history of an archaic hominin group from Denisova Cave in Siberia
http://repositori.up[...]
[25]
간행물
Early history of Neanderthals and Denisovans
[26]
간행물
Hominin interbreeding and the evolution of human variation
[27]
간행물
Adaptive evolution of the ''Homo'' mitochondrial genome
2011
[28]
간행물
Approximate Bayesian computation with deep learning supports a third archaic introgression in Asia and Oceania
[29]
간행물
The complete genome sequence of a Neanderthal from the Altai Mountains
[30]
간행물
Ancient gene flow from early modern humans into Eastern Neanderthals
[31]
간행물
Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals
[32]
간행물
Genomic analysis of Andamanese provides insights into ancient human migration into Asia and adaptation
[33]
간행물
Hominin DNA baffles experts
[34]
서적
The Strange Case of the Rickety Cossack and other Cautionary Tales from Human Evolution
Palgrave Macmillan
[35]
간행물
Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins
[36]
논문
First Aboriginal genome sequenced
[37]
논문
Multiple Deeply Divergent Denisovan Ancestries in Papuans
[38]
웹사이트
Denisovan DNA may help modern humans adapt to different environments
https://www.newscien[...]
2024-08-11
[39]
논문
Did the Denisovans Cross Wallace's Line?
[40]
논문
Siberia's ancient ghost clan starts to surrender its secrets
[41]
논문
Reconstructing the DNA methylation maps of the Neandertal and the Denisovan
[42]
논문
Reconstructing Denisovan Anatomy Using DNA Methylation Maps
[43]
논문
Late Pleistocene archaic human crania from Xuchang, China
[44]
논문
A High-Coverage Genome Sequence from an Archaic Denisovan Individual
[45]
논문
Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA
[46]
논문
Archaic Adaptive Introgression in TBX15/WARS2
[47]
논문
Neanderthal and Denisovan ancestry in Papuans: A functional study
[48]
논문
Timing of archaic hominin occupation of Denisova Cave in southern Siberia
https://ro.uow.edu.a[...]
2019-01-01
[49]
논문
Dating of hominin discoveries at Denisova
[50]
논문
Middle and Late Pleistocene Denisovan subsistence at Baishiya Karst Cave
2024-07-03
[51]
논문
Earliest parietal art: Hominin hand and foot traces from the middle Pleistocene of Tibet
[52]
논문
More Genomes from Denisova Cave Show Mixing of Early Human Groups
[53]
논문
A draft sequence of the Neandertal genome
http://www.eva.mpg.d[...]
[54]
논문
Mum's a Neanderthal, Dad's a Denisovan: First discovery of an ancient-human hybrid – Genetic analysis uncovers a direct descendant of two different groups of early humans.
[55]
논문
Mum's a Neanderthal, Dad's a Denisovan: First discovery of an ancient-human hybrid
[56]
논문
The complete genome sequence of a Neanderthal from the Altai Mountains
[57]
논문
Outstanding questions in the study of archaic hominin admixture
[58]
논문
Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage
[59]
논문
Neanderthal-Denisovan ancestors interbred with a distantly related hominin
[60]
논문
Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania
[61]
논문
A genetic history of migration, diversification, and admixture in Asia
https://www.pivotsci[...]
2022-01-06
[62]
논문
Excavating Neandertal and Denisovan DNA from the genomes of Melanesian individuals
[63]
논문
An Aboriginal Australian genome reveals separate human dispersals into Asia
[64]
논문
Philippine Ayta possess the highest level of Denisovan ancestry in the world
2021-08-01
[65]
논문
The complete genome sequence of a Neanderthal from the Altai Mountains
[66]
논문
DNA analysis of an early modern human from Tianyuan Cave, China
[67]
논문
Analysis of Human Sequence Data Reveals Two Pulses of Archaic Denisovan Admixture
[68]
논문
Genome of a middle Holocene hunter-gatherer from Wallace
[69]
논문
The nature of Neanderthal introgression revealed by 27,566 Icelandic genomes
[70]
논문
The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans
[71]
논문
The combined landscape of Denisovan and Neanderthal ancestry in present-day humans
[72]
논문
Could the Denisovan Genes have conferred enhanced Immunity Against the G614 Mutation of SARS-CoV-2?
https://www.research[...]
2021-07-19
[73]
논문
Denisovan introgression has shaped the immune system of present-day Papuans
2022-12-08
[74]
뉴스
Morning Person? You Might Have Neanderthal Genes to Thank. - Hundreds of genetic variants carried by Neanderthals and Denisovans are shared by people who like to get up early.
https://www.nytimes.[...]
2023-12-14
[75]
기타
[76]
웹사이트
데니소와인, 현생인류와 교잡의 가능성
https://natgeo.nikke[...]
2010-12-24
[77]
논문
The complete mitochondrial DNA genome of an unknown hominin from southern Siberia
[78]
뉴스
인류에 미발견의 신계통인가, 4만년 전의 시베리아에
https://natgeo.nikke[...]
2010-03-25
[79]
뉴스
이것이 데니소와인이다 DNA에서 골격을 재현, 초
https://natgeo.nikke[...]
2019-09-24
[80]
웹사이트
데니소와인에 별개의 그룹, 아시아에서 또 놀라운 발견 현생인류와 교배한 “구인”은 다양했다, 현대인의 DNA에서 판명, 인도네시아
https://natgeo.nikke[...]
2019-04-15
[81]
서적
Exploring Human Biology in the Laboratory
https://books.google[...]
Morton Publishing Company
[82]
논문
The Morphology of Permanent Molars from the Paleolithic Layers of Denisova Cave
https://www.research[...]
[83]
뉴스
「데니소와인」, 아시아에도 분포인가=5만〜3만년전―세포핵 게놈 해독・국제팀
http://www.asahi.com[...]
2010-12-23
[84]
뉴스
현대인의 조상, 별개의 인류「데니소와인」과 교잡
http://www.yomiuri.c[...]
2010-12-23
[85]
웹사이트
아시아에서도 데니소와인과 교잡의 가능성
https://natgeo.nikke[...]
2011-10-31
[86]
웹사이트
인류 3종이 수만년도 공존, 데니소와인 연구에서 판명
https://natgeo.nikke[...]
[87]
뉴스
티베트인의 고지 적응능력, 멸종 인류 계통으로부터 획득인가 국제 연구
https://www.afpbb.co[...]
2014-07-03
[88]
서적
네안데르탈인은 우리와 교배했다
분게이슌주
2015-06
[89]
웹사이트
소녀의 부모는, 네안데르탈인과 데니소와인
https://natgeo.nikke[...]
[90]
웹사이트
「어머니는 네안데르탈인, 아버지는 데니소와인」 러시아에서 발견의 화석 DNA감정에서
https://www.bbc.com/[...]
[91]
웹사이트
어머니는 네안데르탈인, 아버지는 데니소와인 DNA분석에서 첫 확인
https://www.jiji.com[...]
[92]
웹사이트
수수께끼의 고대 인류 데니소와인의 골격을 DNA에서 복원
https://style.nikkei[...]
2019-10-07
[93]
웹사이트
Reconstructing Denisovan Anatomy Using DNA Methylation Maps
https://www.scienced[...]
2019-09-19
[94]
뉴스
https://www.nature.c[...]
2022-05-17
[95]
웹사이트
40만년 전 인류 DNA, 데니소와인과 유사
http://www.yonhapnew[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com