마카이로두스아과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
마카이로두스아과는 고대 그리스어에서 '칼'과 '이빨'을 의미하는 단어에서 유래되었으며, 검치호랑이아과로도 불린다. 시어도어 길에 의해 명명되었으며, 마카이로두스족, 스밀로돈족, 호모테리움족, 메타일루루스족의 4개 족으로 분류된다. 이들은 중신세에 기원하여 검 모양의 송곳니를 특징으로 하며, 멸종된 종을 포함한다. 검치호라는 명칭은 오해를 불러일으킬 수 있으며, 이들은 현생 고양이과와는 다른 계통을 가지고 있다. 마카이로두스아과는 두개골과 체골격의 특징, 그리고 물기 힘, 식성, 사회적 행동, 송곳니의 기능 등 다양한 특징을 가지고 연구되어 왔다.
더 읽어볼만한 페이지
- 마카이로두스아과 - 제노스밀루스
제노스밀루스는 초기 플라이스토세 시대 북미에 살았던 멸종된 고양이과 동물로, 튼튼한 체격과 독특한 송곳니 형태를 지니며 사회적 동물이었을 가능성이 제기된다. - 마카이로두스아과 - 시미타고양이
시미타고양이는 플라이오세 초기부터 플라이스토세 후기까지 유라시아, 아프리카, 북아메리카, 남아메리카 등지에 서식했던 멸종된 검치호랑이의 한 속으로, 긴 다리와 짧은 꼬리, 톱니 모양의 송곳니를 특징으로 하며, 큰 먹이를 사냥하고 집단 사냥을 했을 것으로 추정된다. - 1872년 기재된 화석 분류군 - 하스트수리
하스트수리는 뉴질랜드에 서식했던 멸종된 대형 맹금류로, 흰점어깨수리, 쇠독수리와 가까운 관계이며, 모아를 주로 사냥했으나 마오리족의 등장과 모아의 멸종으로 멸종되었다. - 1872년 기재된 화석 분류군 - 아가타우마스
아가타우마스는 1872년 와이오밍 주에서 발견된 각룡류 공룡의 한 속으로, 한때 육상 동물 중 가장 큰 것으로 여겨졌으나, 현재는 불확실한 분류로 트리케라톱스의 화석일 가능성이 제기된다. - 시어도어 길이 명명한 분류군 - 가시두더지
가시두더지는 오스트레일리아와 뉴기니섬에 서식하며 가시로 덮인 몸, 긴 주둥이, 발톱을 가진 단공류 포유류로, 알을 낳고 개미 등을 먹으며, 위협 시 몸을 둥글게 말거나 땅에 숨는 특징을 지닌다. - 시어도어 길이 명명한 분류군 - 쥐가오리과
쥐가오리과는 날개 모양 가슴지느러미와 머리지느러미를 가진 연골어류로, 여과 섭식을 하며, 분자생물학적 연구를 통해 매가오리과에서 독립된 과로 인정받았고, 쥐가오리속을 유일한 현존 속으로 가지며 암초대왕쥐가오리, 대왕쥐가오리 등을 포함하고, 멸종된 화석 속인 아르키아오만타도 알려져 있다.
| 마카이로두스아과 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Machairodontinae |
| 명명자 | Gill, 1872 |
| 지질 시대 | 중신세 중기 - 홀로세 초기 |
| 분류 | |
| 하위 그룹 | 호모테리움족 마카이로두스족 메타일루루스족 스밀로돈족 |
2. 어원
마카이로두스(Μαχαιροῦς)는 고대 그리스어에서 유래되었으며, 마카이라(μάχαιρα)는 '칼'을 의미하고, 오돈토스(oδόντος)는 "이빨"을 의미한다.
시어도어 길은 1872년에 마카이로두스아과(Machairodontinae)라는 아과명을 명명하였다. 기준 속이 되는 마카이로두스(Machairodus)는 μάχαιρα|machaira|단검grc οδούς|odous|이grc에서 유래되었으며, "단검의 이"라는 의미를 갖는다. 명명 규약에 따라 ''Machairodus''를 속격화하고 아과명 어미 "-inae"를 붙여 이 아과명이 되었다.[64]
3. 하위 분류
족 속 종 [6][7] | †호모테리니족(Homotherini) 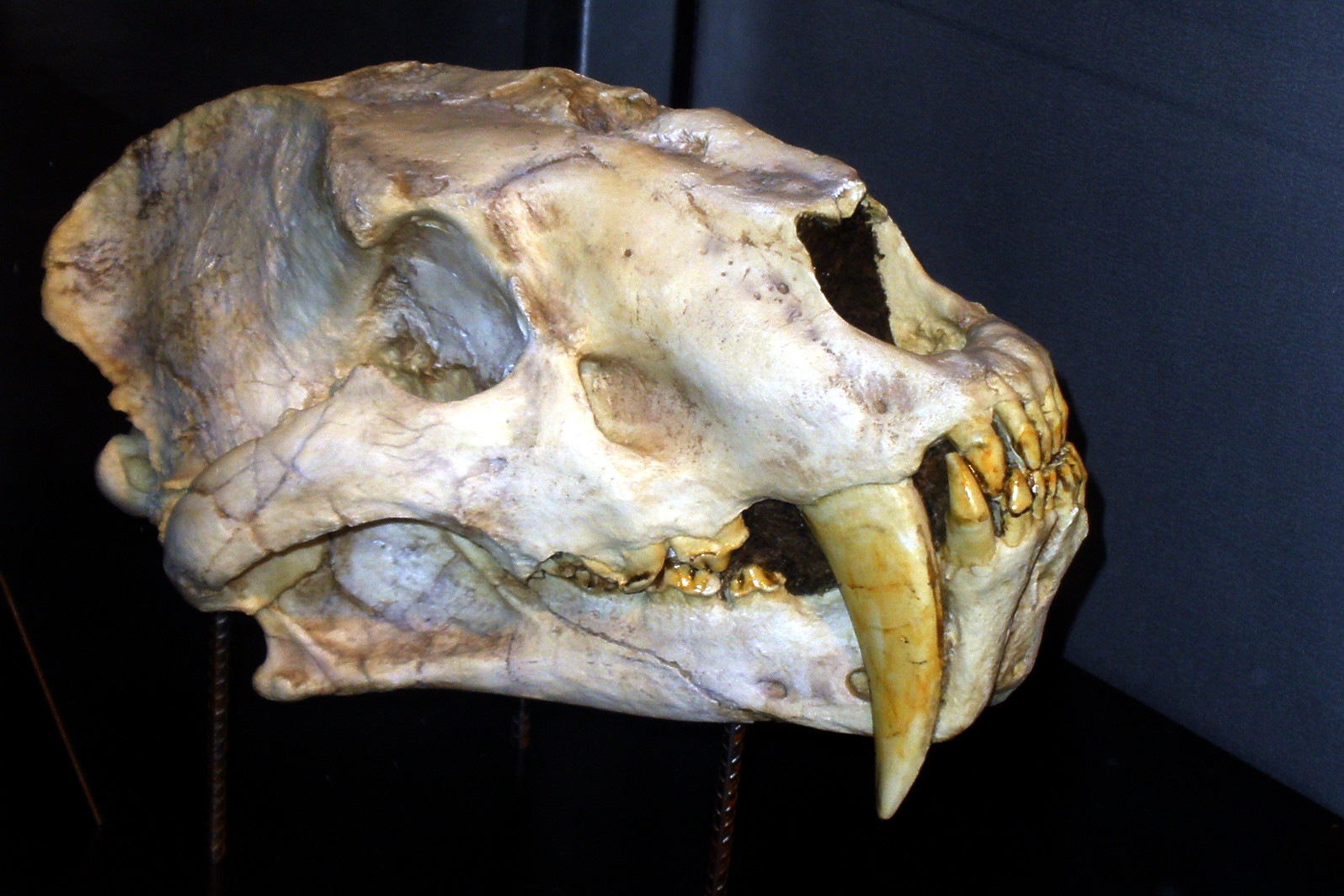
†'암피마카이로두스(Amphimachairodus)'[16] | 
†'호모테리움(Homotherium)' | 
†'로코툰자일루루스(Lokotunjailurus)' | 
†'님라비데스(Nimravides)'[16] | 
†'제노스밀루스(Xenosmilus)' | †마카이로돈티니족(Machairodontini) †'헤미마카이로두스(Hemimachairodus)' | 
†'마카이로두스(Machairodus)' | †'롱추안스밀루스(Longchuansmilus)' | †'아데일로스밀루스(Adeilosmilus)' | †'타오우(Taowu)' | †'미오마카이로두스(Miomachairodus)' | †메타일루리니족(Metailurini) †'아델파일루루스(Adelphailurus)' | 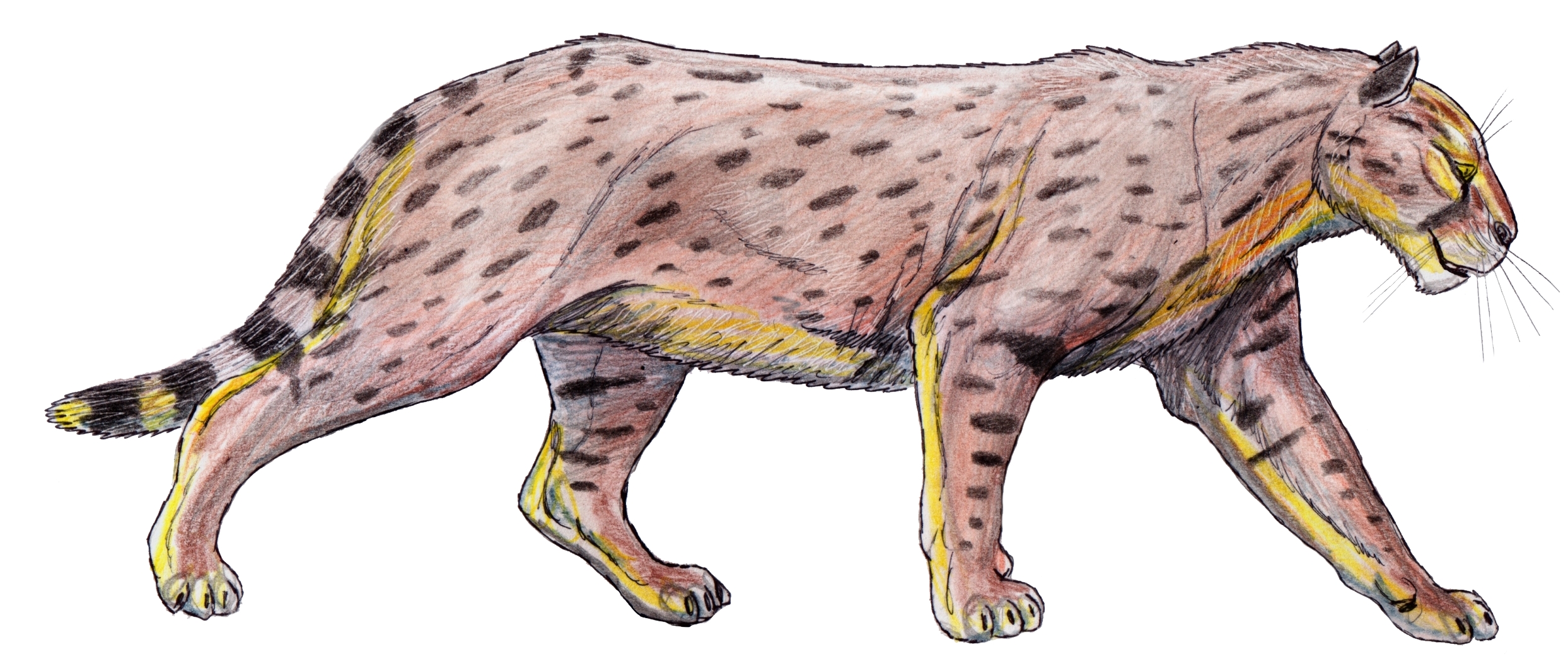
†'디노펠리스(Dinofelis)' | 
†'메타일루루스(Metailurus)' | †'스테나일루루스(Stenailurus)' | 
†'포르투니크티스(Fortunictis)' | †'요시(Yoshi)'[8] | †스밀로돈티니족(Smilodontini) 
†'메간테레온(Megantereon)' | †'파라마카이로두스(Paramachairodus)' | 
†'프로메간테레온(Promegantereon)'[16] | 
†'리조스밀로돈(Rhizosmilodon)' | 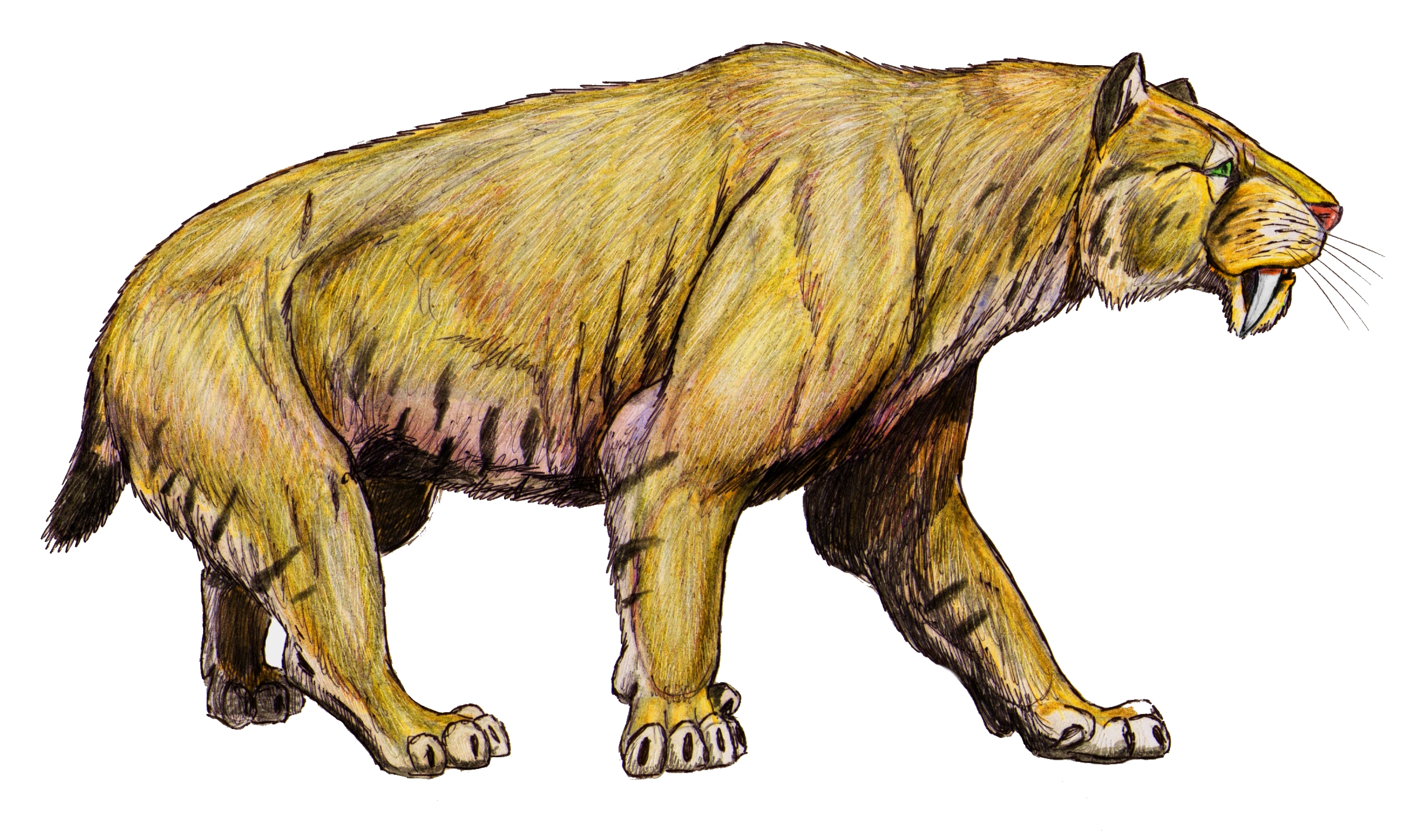
†'스밀로돈(Smilodon)' |
4. 계통 분류
족 속 종 [6][7] | †호모테리니족(Homotherini) †'암피마카이로두스(Amphimachairodus)'[16] | †'호모테리움(Homotherium)' | †'로코툰자일루루스(Lokotunjailurus)' | †'님라비데스(Nimravides)'[16] | †'제노스밀루스(Xenosmilus)' | †마카이로돈티니족(Machairodontini) †'헤미마카이로두스(Hemimachairodus)' | †'마카이로두스(Machairodus)' | †'롱추안스밀루스(Longchuansmilus)' | †'아데일로스밀루스(Adeilosmilus)' | †'타오우(Taowu)' | †'미오마카이로두스(Miomachairodus)' | †메타일루리니족(Metailurini) †'아델파일루루스(Adelphailurus)' | †'디노펠리스(Dinofelis)' | †'메타일루루스(Metailurus)' | †'스테나일루루스(Stenailurus)' | †'포르투니크티스(Fortunictis)' | †'요시(Yoshi)'[8] | †스밀로돈티니족(Smilodontini) †'메간테레온(Megantereon)' | †'파라마카이로두스(Paramachairodus)' | †'프로메간테레온(Promegantereon)'[16] | †'리조스밀로돈(Rhizosmilodon)' | †'스밀로돈(Smilodon)' |
마카이로두스아과의 계통 발생 관계는 다음 분기도를 통해 나타낸다.[9][10][11][12][13]
프로아일루루스와 Pseudaelurus영어 및 현생 고양이과의 3개 그룹을 외군으로 하는 마카이로두스아과의 계통 발생을 각 속에 대한 간략한 설명과 함께 나타낸다.[68][69][70][71]。이하, Ma는 백만 년 전을 나타낸다.
{{clade| style=font-size:90%;line-height:100%
|label1='''고양이과'''
|1={{clade
|1=프로아일루루스 25Ma: 유럽・아시아: 1종: 체중 9 kg
|2={{clade
|1=Pseudaelurus영어 18Ma: 유럽・아시아・북미: 12종: 평균 체중 40 kg
|2={{clade
|1={{clade
|label1='''고양이아과'''(광의)
|1=(현생 고양이류 10Ma- 현세: 북미・남미・유럽・아시아・아프리카: 44종)
|label2='''마카이로두스아과'''
|2={{clade
|1={{clade
|label1=메타일루루스족
|1={{clade
|1=Adelphailurus영어 10-5Ma: 북미: 1종: 체중 50 kg
|2=Pontosmilus영어 4종
|3=Stenailurus영어'' 7Ma: 유럽
|4={{clade
|1=메타일루루스 9Ma - 11,000년 전: 아프리카・북미・유럽・아시아: 6종
|2=디노펠리스 (또는 ''Therailurus'') 5-1Ma: 유럽・아시아・아프리카・북미: 8종: 평균 체중 90 kg}} }}
|2={{clade
|label1=스밀로돈족
|1={{clade
|1=Paramachairodus영어(또는 파라메간테레온 ''Paramegantereon'', 프로메간테레온 ''Promegantereon'') 15-9Ma: 유럽・아시아: 2종: 평균 체중 55 kg
|2={{clade
|1=메간테레온 6-2Ma: 북미・아프리카・아시아: 12종: 평균 체중 120 kg
|2=스밀로돈 
2.5Ma - 10,000년 전: 북미・남미: 3종: 평균 체중 400 kg}}}}
|2={{clade
|1={{clade
|label1=호모테리움족
|1={{clade
|1=Lokotunjailurus영어 10Ma: 아프리카
|2={{clade
|1=호모테리움
5Ma- 10,000년 전: 북미・남미・유럽・아시아・아프리카: 14종: 평균 체중 190 kg
|2= 1Ma: 북미: 1종: 300 kg}} }}
|label2=마카이로두스족
|2={{clade
|1=Miomachairodus영어 12Ma: 유럽・아시아・아프리카・북미: 1종
|2=마카이로두스
(또는 님라비데스 ''Nimravides'', 암피마카이로두스 ''Amphimachairodus'') 11Ma- 126,000년 전: 유럽・아시아・아프리카・북미: 20종: 평균 체중 300 kg
}} }} }} }} }} }} }} }} }} }}
'''분류'''
Amphimachairodus영어
Lokotunjailurus영어
Nimravides영어
호모테리움
Adelphailurus영어
디노펠리스
Pontosmilus영어
Stenailurus영어
메타일루루스
메간테레온
Paramachairodus영어
Rhizosmilodon영어
스밀로돈
Hemimachairodus영어
Miomachairodus영어
마카이로두스
5. 진화
마카이로두스아과는 중기 미오세 유럽에서 기원했다.[2] 초기 고양이과 동물 ''Pseudaelurus quadridentatus''는 위턱 송곳니가 길어지는 경향을 보였으며, 마카이로두스아과의 진화의 기초로 여겨진다.[3] 가장 초기에 알려진 마카이로두스 속은 아프리카와 터키에서 발견된 중기 미오세 ''Miomachairodus''이다.[1] 후기 미오세까지, 마카이로두스아과는 긴 검치아를 가진 고대 대형 육식동물인 바르부로펠리스과와 함께 여러 지역에서 공존했다.[1]
전통적으로 마카이로두스아과는 스밀로돈티니, 마카이로돈티니(호모테리니), 메타일루리니의 세 부족으로 분류되었다. 스밀로돈티니는 ''Megantereon''과 ''Smilodon''과 같은 일반적인 검치 형태를 가졌고, 마카이로돈티니(호모테리니)는 ''Machairodus'' 또는 ''Homotherium''과 같은 낫 모양의 이빨을 가진 고양이를 포함하며, 메타일루리니는 ''Dinofelis'' 및 ''Metailurus''와 같은 속을 포함한다. 그러나 과거에는 일부 학자들이 메타일루리니를 현생 고양이와 함께 고양이과 펠리나에아과로 재분류하기도 했다.[1] 2022년 계통학적 연구에 따르면 ''Paramachairodus'' 속이 두 그룹의 조상이며, 메타일루리니와 스밀로돈티니 사이의 다계통 관계를 시사한다.[4]
미토콘드리아 DNA 염기서열을 기반으로 한 화석에서 추출한 마카이로두스는 약 2천만 년 전에 현존하는 고양이와 분기되었으며, 마지막으로 생존한 마카이로두스 속인 ''호모테리움''과 ''스밀로돈''은 약 1천 8백만 년 전에 서로 분기된 것으로 추정된다.[5]
'검치호'라는 이름은 오해의 소지가 있다. 마카이로두스는 호랑이와 같은 아과에 속하지 않았고, 호랑이와 같은 털 무늬를 가졌다는 증거도 없으며, 이 광범위한 동물군이 현대의 호랑이와 같은 방식으로 모두 살거나 사냥하지 않았기 때문이다. 2005년에 발표된 DNA 분석은 분기도 분석을 통해 마카이로두스아과가 현대 고양이의 조상과 일찍 분기되었으며 현존하는 고양이 종과 밀접한 관련이 없음을 보여주었다.[1]
| 족 | 속 | 종 |
|---|---|---|
| Incertae sedis (분류 불확실) | †'차다일루루스(Tchadailurus)'[6][7] | |
| †호모테리니족(Homotherini) | †'암피마카이로두스(Amphimachairodus)'[16] | |
| †'호모테리움(Homotherium)' | ||
| †'로코툰자일루루스(Lokotunjailurus)' | ||
| †'님라비데스(Nimravides)'[16] | ||
| †'제노스밀루스(Xenosmilus)' | ||
| †마카이로돈티니족(Machairodontini) | †'헤미마카이로두스(Hemimachairodus)' | |
| †'마카이로두스(Machairodus)' | ||
| †'롱추안스밀루스(Longchuansmilus)' | ||
| †'아데일로스밀루스(Adeilosmilus)' | ||
| †'타오우(Taowu)' | ||
| †'미오마카이로두스(Miomachairodus)' | ||
| †메타일루리니족(Metailurini) | †'아델파일루루스(Adelphailurus)' | |
| †'디노펠리스(Dinofelis)' | ||
| †'메타일루루스(Metailurus)' | ||
| †'스테나일루루스(Stenailurus)' | ||
| †'포르투니크티스(Fortunictis)' | ||
| †'요시(Yoshi)'[8] | ||
| †스밀로돈티니족(Smilodontini) | †'메간테레온(Megantereon)' | |
| †'파라마카이로두스(Paramachairodus)' | ||
| †'프로메간테레온(Promegantereon)'[16] | ||
| †'리조스밀로돈(Rhizosmilodon)' | ||
| †'스밀로돈(Smilodon)' |
마카이로두스아과의 계통 발생 관계는 다음 분기도와 같다.[9][10][11][12][13]
{{clade |style=font-size:85%;line-height:85%;background-color:#eeeeFF;border:1px solid;
|label1= †'''마카이로두스아과'''
|1={{clade
|label1= †'''메타일루리니''' |style1=background-color:#eeccFF;
|1={{clade
|1={{clade
|label1= †''디노펠리스''
|1={{clade
|1={{clade
|1=†''디노펠리스 팔레온카''
|2=†''디노펠리스 페테리''
}}
|2=†''디노펠리스 아로노키''
|3=†''디노펠리스 바를로위''
|4=†''디노펠리스 크리스타타''
|5=†''디노펠리스 다르티''
|6=†''디노펠리스 디아스테마타''
|7=†''디노펠리스 피베토이''
}}
|label2= †''메타일루루스''
|2={{clade
|1=†''메타일루루스 부우돈''
|2=†''메타일루루스 메이저''
|3=†''메타일루루스 몽골리엔시스''
|4=†''메타일루루스 울티무스''
}} }}
|label2= †''아델파일루루스''
|2=†''아델파일루루스 칸센시스''
|label3= †''스테나일루루스''
|3=†''스테나일루루스 테일하르디''
|label4= †''요시''[14]
|4={{clade
|1=†''요시 가레브스키''
|2=†''요시 마이너''
}} }}
|label2= †''차다일루루스'' |style2=background-color:#ddddFF; |state2=dashed
|2=†''차다일루루스 아데이''
|3={{clade
|label1= †'''스밀로돈티니''' |style1=background-color:#ccccFF;
|1={{clade
|1={{clade
|1={{clade
|label1= †''메간테레온''
|1={{clade
|1={{clade
|1={{clade
|1=†''메간테레온 컬트리덴스''
|2=†''메간테레온 에키도이트''
}}
|2=†''메간테레온 화이티''
}}
|2=†''메간테레온 헤스페루스''
|3=†''메간테레온 이넥스펙타투스''
|4=†''메간테레온 마이크로타''
|5=†''메간테레온 니호와넨시스''
|6=†''메간테레온 박센시스''
}}
|label2= †''스밀로돈''
|2={{clade
|1=†''스밀로돈 파탈리스''
|2={{clade
|1=†''스밀로돈 그라킬리스''
|2=†''스밀로돈 포풀라토르''
}} }} }}
|label2= †''리조스밀로돈''
|2=†''리조스밀로돈 피테아이''[15]
}}
|label2= †''파라마카이로두스''
|2={{clade
|1=†''파라마카이로두스 막시밀리아니''
|2=†''파라마카이로두스 오리엔탈리스''
|3=†''파라마카이로두스 트란스아시아티쿠스''
}} }}
|label2= †''프로메간테레온''
|2=†''프로메간테레온 오기기아''[16]
}}
|2={{clade
|label1= †'''마카이로돈티니''' |style1=background-color:#ccFFcc;
|1={{clade
|label1= †''마카이로두스''
|1={{clade
|1=†''마카이로두스 알베르디에''
|2=†''마카이로두스 아파니스트스''
|3=†''마카이로두스 호리빌리스''
|4=†''마카이로두스 라스케레비''
|6=†''마카이로두스 로빈소니''
}}
|label2= †''미오마카이로두스''
|2=†''미오마카이로두스 슈도아엘루로이데스''
|label3= †''헤미마카이로두스''
|3=†''헤미마카이로두스 즈비에르지키''
}}
|label2= †'''호모테리니''' |style2=background-color:#FFdddd;
|2={{clade
|label1= †''암피마카이로두스''[16]
|1={{clade
|1=†''암피마카이로두스 쿠르테니''
|2=†''암피마카이로두스 알바레지''
|3=†''암피마카이로두스 콜로라덴시스''
|4=†''암피마카이로두스 기가스''
|5=†''암피마카이로두스 카비르''
}}
|label2= †''님라비데스''[16]
|2={{clade
|1=†''님라비데스 카타콥시스''
|2=†''님라비데스 갈리아니''
|3=†''님라비데스 히바르디''
|4=†''님라비데스 페디오노무스''
|5=†''님라비데스 티노바테스''
}}
|3={{clade
|label1= †''로코툰자일루루스''[16]
|1={{clade
|1=†''로코툰자일루루스 에마게리투스''
|2=†''로코툰자일루루스 파노네이''
}}
|2={{clade
|label1= †''제노스밀루스''
|1=†''제노스밀루스 호드소네이''
|label2= †''호모테리움''
|2={{clade
|1=†''호모테리움 이스치루스''
|2=†''호모테리움 라티덴스''
|3={{clade
|1=†''호모테리움 세룸''
|2=†''호모테리움 베네주엘렌시스''
}} }} }} }} }} }} }}
5. 1. 진화 역사와 표현형의 기원
1990년대 바탈로네스-1에서 마이오세 후기 화석 매장지가 발견되기 전까지, 스밀로돈티니와 호모테리니 조상의 표본은 희귀하고 단편적이어서 검치 표현형의 진화 역사는 대체로 알려지지 않았다.[17][18] 바탈로네스-1 발굴 이전에는 고도로 파생된 검치 표현형이 다면발현 진화를 통해 빠르게 나타났다는 가설이 지배적이었다.[19]바탈로네스-1에서는 스밀로돈티니 조상인 ''Promegantereon ogygia''와 호모테리니 조상인 ''Machairodus aphanistus''의 새로운 표본이 발굴되어 진화 역사에 대한 새로운 통찰력을 제공했다.[17][18] 표범 크기의 ''P. ogygia''는 900만 년 전에 스페인(및 추가 영토일 수 있음)에 서식했다.[21]
바탈로네스-1을 통해 가능해진 검치 표현형의 진화에 대한 현재 가설은 이 표현형이 모자이크 진화를 통해 시간이 지남에 따라 점진적으로 나타났다는 것이다.[17][18] 정확한 원인은 불확실하지만, 현재 연구 결과는 먹이를 빠르게 죽여야 할 필요성이 진화적 시간 동안 이 표현형의 발달을 이끄는 주요 압력이었다는 가설을 뒷받침한다. 부러진 이빨이 많이 나타나는 것으로 볼 때, 검치 고양이의 생물 환경은 치열한 경쟁이 특징이었다.[22][23]
부러진 이빨은 이빨이 뼈와 접촉하는 빈도를 나타낸다. 이빨-뼈 접촉의 증가는 시체의 소비 증가, 먹이의 빠른 소비 또는 사냥에 대한 공격성 증가를 시사하며, 이 세 가지 모두 먹이 가용성의 감소를 시사하여 포식자 간의 경쟁을 심화시킨다. 이러한 경쟁 환경은 먹이를 더 빨리 죽이는 것을 선호하는데, 이는 소비 전에 먹이를 빼앗기면 해당 먹이를 잡는 데 드는 에너지 비용이 보상되지 않고, 포식자의 생애 동안 이러한 일이 자주 발생하면 과로 또는 기아로 죽을 수 있기 때문이다. 먹이를 죽이는 속도를 개선하는 초기 적응은 ''P. ogygia''와 ''M. aphanistus''의 두개골과 아래턱에 존재하며,[17][18] ''P. ogygia''의 경추[18]와 앞다리[24]에도 존재한다. 이는 검치 표현형 진화에서 속도의 중요성을 뒷받침하는 추가적인 형태학적 증거를 제공한다.[24]
마카이로두스아과는 전기 또는 중기 중신세의 아프리카에서 기원했다. 초기의 고양이과 동물인 ''Pseudaelurus quadridentatus''는 송곳니가 길어지는 경향을 보였으며, 마카이로두스아과 진화의 기본이 된 것으로 생각된다.[66] 이 아과에 속하는 최초의 동물은 중기 중신세의 미오마카이로두스 (''Miomachairodus'')로, 아프리카와 터키에서 화석이 발견되었다.[67]
전통적으로 마카이로두스아과는 스밀로돈족, 마카이로두스족(호모테리움족), 메타일루루스족의 3개 족으로 분류되었다. 그러나 최근 메타일루루스족을 현생 고양이류와 같은 고양이아과로 재분류하는 사례가 나타나고 있다.[62] 마지막 마카이로두스아과 속인 스밀로돈과 호모테리움은 1만 년 전, 홍적세의 북아메리카와 남아메리카 대륙에도 생존해 있었다.
2005년에 발표된 DNA 분석에서는, 마카이로두스아과가 현재 고양이류의 조상과 이른 시기에 분리되었고, 현생 고양이아과와의 근연성은 낮다는 분지학적 결론이 나왔다.[62]
마카이로두스류 그룹에서 가장 잘 연구된 것은 두개골, 특히 그 이빨이다. 마카이로두스아과는 포식자와 피식자의 관계, 특수화, 강한 육식성의 분석 연구 방법으로 최상의 것 중 하나가 되었다.[99]
thumb로 나기 시작하고 있다]]
마카이로두스아과는 다크형 송곳니 그룹과 시미터형 송곳니 그룹으로 나뉜다. 다크형 송곳니 그룹은 가늘고 길게 뻗은 위턱 송곳니를 가지며, 일반적으로 튼튼한 체형을 하고 있다. 시미터형 송곳니 그룹은 폭이 넓고 짧은 위턱 송곳니와 대체로 소형의 몸과 긴 사지를 가지고 있다. 긴 송곳니를 가진 종은 종종 뻗은 골질의 칼날을 아래턱에 가지고 있다.
식육목은 식물이나 곤충을 씹어 먹는 것에서 고기를 먹는 것으로 특화됨에 따라 그 치아 수를 줄여나간다. 고양이과는 기본 치식으로 3/3・1/1・3/2・1/1의 치식을 가지지만[79], 스밀로돈과 같은 몇몇 속에서는 소구치가 1개 소실되거나 흔적적인 예가 알려져 있다.[80]
긴 송곳니를 가지면 크게 입을 벌려야 한다. 마카이로두스류는 수렴 진화에 의해 유사한 송곳니를 얻은 다른 그룹과 마찬가지로, 그러한 송곳니를 수용하기 위해 두개골에 몇 가지 개량을 가할 필요가 있었다.
thumb
thumb 두개골의 최대 개구 (80°)]]
포유류에서 크게 입을 벌리는 데 주요 장애물은 턱 뒤쪽에 있는 교근과 측두근이다. 더 크게 입을 벌리기 위해 이 동물들은 그 근육을 소형화하고 형상 변화를 해야 했다. 그 첫 번째 단계는 하악골 근육돌기의 축소이다. 근육이 축소될수록 더 큰 유연성을 얻고 더 큰 개구가 가능해진다.
마카이로두스류 두개골에서는 측두근의 형상에 다른 변화도 나타난다. 입을 벌릴 때의 주된 제약은 개구 시 측두근이 턱 관절 주변에 과도하게 신장되면 파열이 일어난다는 것이다. 현생 고양이과 동물에서는 후두골이 뒤로 뻗어 그 표면에 부착되어 있는 측두근이 크게 입을 벌렸을 때 관절 주위에 감겨 늘어나 버린다. 측두근의 신장을 경감시키기 위해 마카이로두스류는 더 수직적인 후두골을 가진 두개골을 진화시켰다. 스밀로돈에서는 개구 각도가 128°가 되었고, 하악저와 후두골이 이루는 각도는 100°이다.
마카이로두스류를 포함한 많은 검치호류의 두개골은 위아래로 길고 앞뒤로 짧아졌다. 협골궁은 압축되었고, 얼굴의 각 부분은 눈이 높은 위치에 있고 콧등이 짧아지는 등의 특징을 가진다.[81]
6. 골격
마카이로두스아과는 크게 검치형과 단검형 두 가지 유형으로 나뉜다. 단검형 고양이는 길고 좁은 위쪽 송곳니를 가지고 있으며 일반적으로 몸집이 튼튼했다. 반면 단도형 고양이는 더 넓고 짧은 위쪽 송곳니와 일반적으로 다리가 더 긴 유연한 몸매를 가졌다. 더 긴 치아를 가진 고양이는 종종 아래턱에서 뻗어나온 뼈로 된 플랜지를 가지고 있었다. 그러나 크세노스밀루스는 이러한 일반적인 특징에서 벗어나 단검형 고양이의 튼튼한 사지와 단도형 고양이의 튼튼한 송곳니를 모두 소유한 독특한 종이었다.[51]
육식동물은 식물이나 곤충을 갈아 먹는 대신 고기를 먹는 데 특화되면서 치아의 수가 줄어들었다. 고양이는 모든 육식 동물 그룹 중 가장 적은 수의 치아를 가지고 있으며, 마카이로두스아과는 그 수를 더욱 줄였다. 대부분의 마카이로두스아과는 각 턱에 6개의 앞니, 2개의 송곳니, 6개의 앞어금니를 유지하고, 위턱에만 2개의 어금니를 가지고 있다. 스밀로돈과 같은 일부 속은 아래턱에 앞어금니가 하나 적어, 아래턱에 4개의 큰 앞어금니와 2개의 뭉툭한 송곳니, 6개의 튼튼한 앞니만 남겨두었다. 송곳니는 부드럽게 뒤로 휘어져 있으며, 톱니가 있지만 미미하고 나이가 들면서 닳아 없어졌다.
더 긴 송곳니는 더 큰 입을 필요로 했다. 마카이로두스아과는 수렴 진화를 통해 유사한 치아를 얻은 다른 동물 그룹과 함께, 송곳니를 수용하기 위해 두개골을 여러 가지 방식으로 변경해야 했다. 포유류가 큰 입을 벌리는 것을 막는 주요 요인은 턱 뒤쪽의 측두근과 교근이다. 이 근육들은 강력하지만 탄성이 좋지 않아 넓게 벌어지는 입에 저항했다. 이를 극복하기 위해 마카이로두스아과는 아래턱의 근육돌기를 줄이고 측두근의 모양을 변경하여 근육의 탄성을 높였다. 이러한 변화는 물기 힘을 약화시키기도 했다.
마카이로두스아과의 두개골은 측두근의 모양에 또 다른 변화가 있음을 보여준다. 턱을 벌리는 주요 제약은 입을 벌릴 때 측두근이 관절와 주위에서 늘어나 찢어지는 것이었다. 이를 줄이기 위해 마카이로두스아과는 더 수직적인 후두골을 가진 두개골로 진화했다. 집고양이는 80°, 사자는 91° 벌어지는 반면, ''스밀로돈''은 128°까지 입을 벌릴 수 있었다. 마카이로두스아과를 포함한 많은 검치 포식자의 두개골은 위아래로 길고 앞뒤로 짧다. 관골궁이 압축되고 눈과 같은 안면 특징을 가진 두개골 부분은 더 높고 주둥이는 더 짧아[27] 이러한 변화는 입을 더 크게 벌리는 것을 보완하는 데 도움이 된다.[28] 마카이로두스아과는 또한 아래쪽 송곳니를 줄여 위턱과 아래턱 사이의 거리를 유지했다.
마카이로두스류의 턱은, 특히 스밀로돈이나 메간테레온과 같은 긴 송곳니를 가진 진화된 종에서는 매우 약했다. 컴퓨터 복원 결과, 스밀로돈은 날뛰는 먹이를 붙잡아 두기 위한 턱 힘이 부족했다.[83] 스밀로돈은 턱 근육만 사용했다면 사자의 1/3밖에 안 되는 교합력을 가졌을 것이다. 그러나 두개골 후단에 연결된 목 근육은 강력하여 머리를 눌러 송곳니로 먹이를 관통할 수 있었다.[83]
6. 1. 두개골
마카이로두스아과는 크게 두 그룹, 즉 스밀로돈, 메간테레온, 파라마카이로두스 등이 속한 어두운 형태의 송곳니 그룹과 마카이로두스, 호모테리움, 제노스밀루스 등이 속한 시미터형 송곳니 그룹으로 나뉜다. 이 두 그룹은 두개골 이외의 체골격 특징에서 차이를 보인다.어두운 형태의 송곳니 그룹은 튼튼하고 힘이 센 체격을 가졌다. 가장 원시적인 파라마카이로두스는 파생된 스밀로돈보다 작고 유연하며, 메간테레온은 그 중간 정도의 체격을 가진다. 이들은 짧은 중족골과 튼튼한 몸통을 가져 지구력이 좋은 주자는 아니었으며, 현생 사자와 비교했을 때 흉곽은 앞쪽이 좁고 뒤쪽이 넓은 통 모양이다. 견갑골은 특히 스밀로돈에서 잘 발달하여 강력한 어깨 근육과 삼두근이 부착될 수 있는 넓은 면적을 제공한다. 경추는 매우 튼튼하며 근육 부착 부위도 튼튼하다. 척추의 요추 부분은 짧고, 꼬리는 진화할수록 짧아져 스밀로돈에서는 보브캣과 같은 꼬리가 되었다. 체골격만 보면 현생 고양이과보다는 현생 곰과 유사하다.
시미터형 송곳니 그룹은 더 다양하며, 대부분의 마카이로두스아과가 여기에 속한다. 이들의 송곳니는 짧고 폭이 넓다. 호모테리움은 한때 족행성으로 여겨졌지만 지행성으로 밝혀졌으며, 이 그룹의 마카이로두스는 모든 마카이로두스아과 중에서 가장 크다. 어두운 형태와 달리 큰 성적 이형성을 보이기도 한다. 호모테리움은 현생 점박이하이에나처럼 기울어진 등을 가져 장거리 주행에 뛰어났을 수 있다. 이들은 일반적으로 긴 사지와 유연한 몸을 가지며, 어두운 형태보다 많은 치아를 가졌다. 마카이로두스는 도약의 명수였을지도 모른다. 체골격만 보면 현생 표범아과 동물(표범속과 구름표범속)과 유사하다.
6. 2. 체골격
마카이로두스아과는 검치형과 단검형 두 가지 유형으로 나뉜다. 단검형 고양이는 길고 좁은 위쪽 송곳니를 가지고 있으며 일반적으로 몸집이 튼튼했다. 단도형 고양이는 더 넓고 짧은 위쪽 송곳니와 일반적으로 다리가 더 긴 유연한 몸매를 가졌다. 더 긴 치아를 가진 고양이는 종종 아래턱에서 뻗어나온 뼈로 된 플랜지를 가지고 있었다. 그러나 두 개의 비교적 완전한 화석만 알려진 ''크세노스밀루스''라는 한 속은 이러한 틀을 깼다. 크세노스밀루스는 단검형 고양이와 관련된 튼튼하고 무거운 사지와 단검형 고양이의 튼튼한 송곳니를 모두 소유했다.[51]육식동물은 식물이나 곤충 물질을 갈아먹는 대신 고기를 먹는 데 특화되면서 치아의 수를 줄였다. 고양이는 모든 육식 동물 그룹 중 가장 적은 수의 치아를 가지고 있으며, 마카이로두스아과는 그 수를 더욱 줄였다. 대부분의 마카이로두스아과는 각 턱에 6개의 앞니, 2개의 송곳니, 6개의 앞어금니를 유지하고, 위턱에만 2개의 어금니를 가지고 있다. ''스밀로돈''과 같은 일부 속은 아래턱에 하나가 적은 8개의 앞어금니만 가지고 있어, 아래턱에 4개의 큰 앞어금니와 2개의 뭉툭한 송곳니, 6개의 튼튼한 앞니만 남겨두었다. 송곳니는 부드럽게 뒤로 휘어져 있으며, 톱니가 있지만 미미하고 나이가 들면서 닳아 없어지기 때문에, 대부분의 중년 마카이로두스아과(약 4~5세)는 톱니가 없다. 이러한 뼈의 힌트는 고생물학자가 멸종된 동물의 개체군 연구를 위해 개체의 나이를 추정하는 데 도움이 된다.

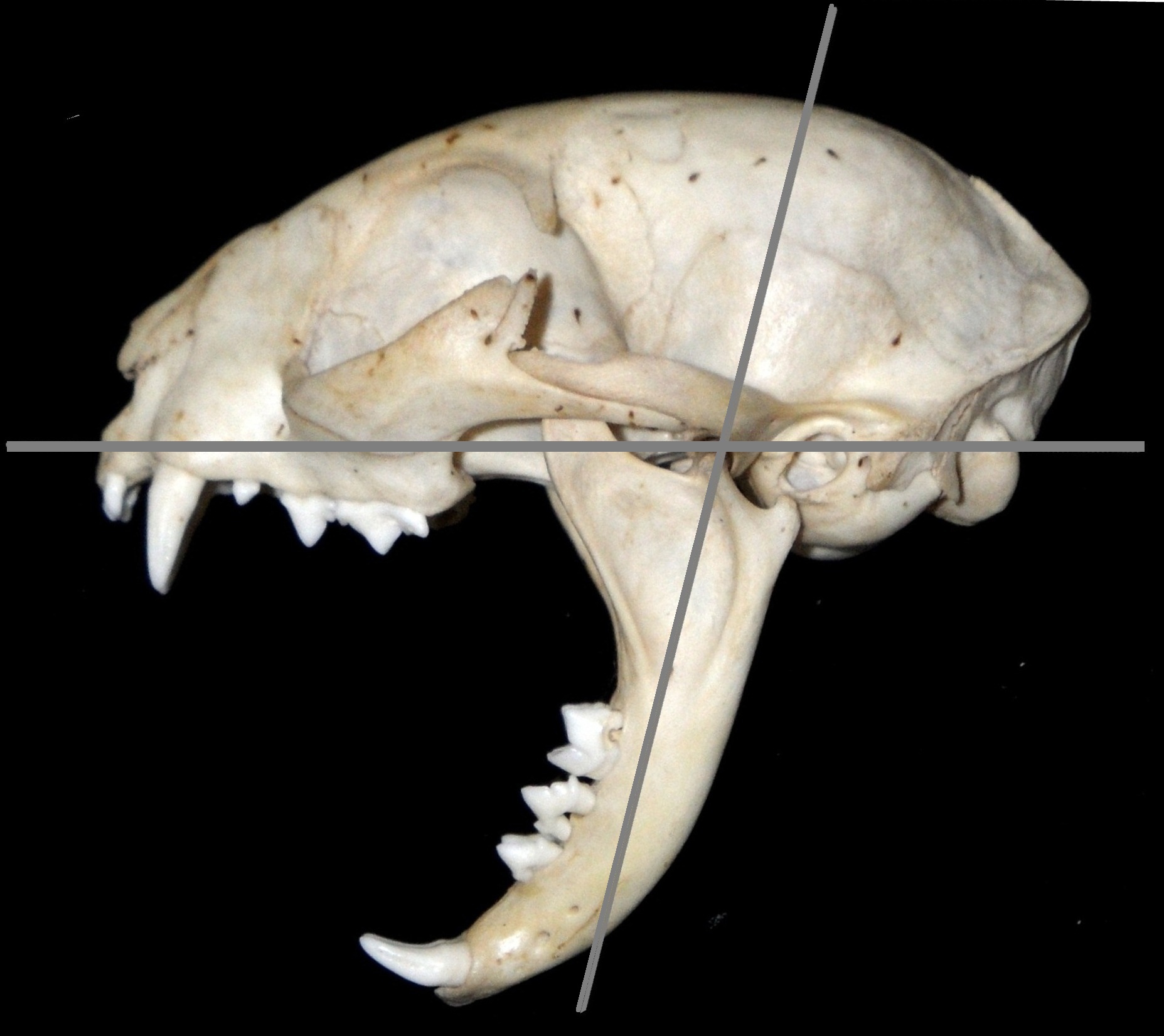
포유류가 큰 입을 벌리는 것을 막는 주요 요인은 턱 뒤쪽의 측두근과 교근이다. 이 근육들은 강력할 수 있고 다양한 물기 힘에 대해 큰 변형을 겪을 수 있지만, 두께, 위치, 강도로 인해 탄성이 좋지 않다. 입을 더 넓게 벌리기 위해, 이 종들은 근육을 더 작게 만들고 모양을 변경해야 했다. 이 과정의 첫 번째 단계는 아래턱의 근육돌기를 줄이는 것이었다. 교근, 특히 측두근은 이 뼈의 돌출부에 삽입되므로, 이 과정을 줄이면 근육도 줄어들게 된다. 각 근육의 질량이 감소하면 탄성이 커지고 넓은 입에 대한 저항이 줄어든다. 이와 관련하여 측두근의 모양을 변경하면 기시점과 삽입점 사이의 거리가 더 멀어져 근육이 더 길고 콤팩트해지는데, 이는 일반적으로 이러한 유형의 스트레칭에 더 적합한 형식이다. 이러한 감소는 물기 힘을 약화시켰다.
마카이로두스아과의 두개골은 측두근의 모양에 또 다른 변화가 있음을 시사한다. 턱을 벌리는 주요 제약은 입을 벌릴 때 측두근이 관절와 주위에서 임계점을 넘어서 늘어나면 찢어진다는 것이다. 현대 고양이과에서는 후두골이 뒤쪽으로 뻗어 있지만, 이 표면에 부착된 측두근은 입을 넓게 벌릴 때 근육이 관절와 주위를 감싸기 때문에 부담을 받는다. 움직일 수 없는 돌기 주위에서 측두근의 스트레칭을 줄이기 위해, 마카이로두스아과는 더 수직적인 후두골을 가진 두개골로 진화했다. 집고양이는 80°의 입을 벌리며, 사자는 91°의 입을 벌린다. ''스밀로돈''에서는 입이 128°이고, 아래턱의 가지와 후두골 사이의 각도는 100°이다. 이 각도는 입을 벌리는 주요 제한 요소이며, ''스밀로돈''에서 볼 수 있듯이 입의 구개에 상대적인 후두골의 각도를 줄이면 입을 더 넓게 벌릴 수 있다.

스밀로돈, 메간테레온, 파라마카이로두스 등 어두운 형태의 송곳니 그룹에 속하는 마카이로두스류는 튼튼함과 힘으로 특징지어진다. 가장 원시적인 파라마카이로두스는 파생된 스밀로돈보다 작고 유연한 몸을 가지고 있으며, 진화 과정에서 이 둘의 중간에 있는 메간테레온은 체격 또한 중간 형태를 보인다. 이들은 짧은 중족골과 튼튼한 몸통을 가지고 있어 지구력이 좋은 주자는 아니었다. 현생 사자와 비교했을 때, 이들의 흉곽은 앞쪽이 좁고 뒤쪽이 넓어진 통 모양을 하고 있다. 견갑골은 특히 스밀로돈에서 잘 발달되어 강력한 어깨 근육과 삼두근이 부착될 수 있는 넓은 면적을 제공한다. 경추는 매우 튼튼하고 근육 부착 부위도 튼튼하다. 척추의 요추 부분은 단축되어 있다. 꼬리는 가장 원시적인 것에서 가장 진화된 것으로 갈수록 점점 짧아져, 그 결과 스밀로돈에서는 보브캣과 같은 꼬리가 되었다. 체골격만 보았을 때, 이들은 현생 고양이류보다는 현생 곰류와 유사한 구조를 가지고 있다.
마카이로두스, 미오마카이로두스, 호모테리움, 아델파일루스, 디노펠리스, 메타일루루스, 폰토스밀루스, 테라일루스, 로코툰자일루스, 제노스밀루스 등을 포함하는 시미터형 송곳니 그룹은 더 다양한 그룹이며, 대부분의 마카이로두스아과는 이 덜 특수화된 유형에 속한다. 이 다수의 구성원을 포함하는 그룹의 송곳니는 명확하게 짧고, 대체로 폭이 넓다. 그룹 내의 다양성 때문에, 전형적인 예를 제시하기는 어렵다. 호모테리움은 한때 족행성으로 여겨졌지만, 지행성인 것으로 밝혀졌다. 이 그룹은 일반적으로 평균보다 날씬하고 작지만, 이 그룹의 마카이로두스는 모든 마카이로두스아과 중에서 가장 큰 것(최소한 가장 큰 것 중 하나)이다. 어두운 형태의 송곳니 그룹과는 달리, 큰 성적 이형성을 보이는 것도 있다. 호모테리움은 현생 점박이하이에나와 매우 유사한 기울어진 등을 가지고 있으며, 마찬가지로 장거리 주행에 뛰어났을 수도 있다. 그룹 전체적으로 긴 사지와 유연한 몸을 가지는 것이 일반적이었다. 시미터형 송곳니 그룹은 평균적인 어두운 형태의 송곳니 그룹보다 많은 치아를 가지고 있었다. 마카이로두스는 도약의 명수였을지도 모른다. 시미터형 송곳니 그룹의 두개골 이후의 체골격만 보면, 그 형태는 현생 표범아과 동물(표범속과 구름표범속)과 비교적 잘 유사하다.
7. 해부학적 특징과 식성
마카이로두스아과는 크게 단검 모양 이빨(단검니) 그룹과 시미터(곡선) 모양 이빨(검치) 그룹으로 나뉜다. 단검니 그룹은 스밀로돈, 메간테레온, 파라마카이로두스 등을 포함하며, 튼튼하고 강인한 체격이 특징이다. 이들은 짧은 발목뼈와 발허리뼈를 가졌고, 늑골은 통 모양으로 앞쪽이 좁고 뒤쪽이 넓다. 특히 스밀로돈은 잘 발달된 어깨뼈를 가져 강력한 어깨 근육과 삼두근이 부착될 수 있었다. 목뼈는 매우 튼튼했고, 허리뼈는 짧아졌다. 꼬리는 진화할수록 짧아져 스밀로돈은 삵과 비슷한 꼬리를 가지게 되었다. 이들의 골격 구조는 현대 곰과 유사하다.[29]
검치 그룹은 마카이로두스, 호모테리움, 제노스밀루스 등을 포함하며, 더 다양한 형태를 보인다. 이들의 송곳니는 현대 고양이보다 길지만 단검니보다는 짧고 굵다. 마카이로두스는 뛰어난 점프 능력을 가졌을 것으로 보이며, 호모테리움은 점박이 하이에나처럼 경사진 등을 가지고 있어 장거리 달리기에 유리했을 수 있다. 제노스밀루스는 검 모양 이빨을 가졌지만, 체형은 단검니와 유사했다. 메타일루리니는 현대 고양이와 유사한 형태를 유지하며 긴 꼬리, 긴 뒷다리, 긴 척추를 가졌다.[46] 검치 고양이는 단검니보다 이빨이 더 많은 경향이 있었다.[29]
조지 밀러는 마카이로두스아과, 특히 스밀로돈의 귓바퀴가 낮게 위치하고, 퍼그처럼 눌린 코를 가졌으며, 입술이 50% 더 길다는 가설을 제시했다. 그러나 귓바퀴 위치 가설은 현대 고양이과 동물과의 비교에서 근거가 부족하며, 코 모양 가설 역시 사자와 호랑이의 코뼈 비교에서 반박된다. 입술 길이 가설은 현대 고양이의 입술 탄력성과 교근 위치를 고려할 때 논란의 여지가 있지만, 일부 복원도에서는 긴 입술을 가진 모습으로 묘사되기도 한다.[36]
스밀로돈과 사자의 설골 비교 연구에 따르면, 스밀로돈을 포함한 마카이로두스류는 현생 대형 고양이과 동물처럼 포효했을 가능성이 제기되었다.[40][41] 안정 동위원소 분석 결과, 스밀로돈은 주로 들소와 말을 사냥했으며, 때로는 땅늘보와 매머드를 사냥한 반면, 호모테리움은 매머드가 없을 때는 어린 매머드와 영양 및 큰뿔양과 같은 다른 초식 동물을 사냥했다는 것이 밝혀졌다.[34]
7. 1. 물기 힘
마카이로두스아과의 송곳니는 형태에 따라 크게 두 그룹으로 나뉜다.1. 단검니 마카이로두스류 (스밀로돈족)
- 대표 속: 스밀로돈, 메간테레온, 파라마카이로두스
- 특징:
- 튼튼하고 강인한 체격
- 짧은 발목뼈와 발허리뼈
- 통 모양의 늑골: 앞쪽은 좁고 뒤쪽은 넓음
- 잘 발달된 어깨뼈: 특히 스밀로돈은 거대한 어깨와 삼두근 근육 부착
- 튼튼한 목뼈와 근육 부착 부위
- 짧아진 허리뼈
- 꼬리: 원시 종에서 진화된 종으로 갈수록 짧아짐 (스밀로돈은 삵과 유사)
- 골격 구조: 현대 곰과 유사[29]
- 지구력이 좋은 주자는 아니었음
2. 검치 마카이로두스류 (마카이로돈티니, 호모테리니, 메타일루리니)
- 대표 속: 마카이로두스, 호모테리움, 제노스밀루스, 로코툰자일루루스, 메타일루루스
- 특징:
- 다양한 형태
- 송곳니: 현대 고양이보다 길지만, 단검니보다는 짧고 굵음
- 마카이로두스: 뛰어난 점프 능력
- 호모테리움:
- 초기 분류군은 현대 사자 크기에 다리가 길고 날씬함
- 플리오세-플라이스토세 속은 지구보행[12]
- 점박이 하이에나처럼 경사진 등: 장거리 달리기에 유리했을 가능성
- 잘 발달된 시각 피질, 큰 비강, 작고 부분적으로만 후퇴 가능한 발톱: 활동적인 주행성 생활 방식 시사[30]
- 제노스밀루스: 검 모양 이빨, 단검니와 유사한 체형[34]
- 메타일루리니:
- 현대 고양이와 유사
- 긴 꼬리, 긴 뒷다리, 긴 척추 유지[46]
- 평균적으로 검치 고양이는 단검니보다 이빨이 많음 (아래턱에 6개의 앞니)[29]
- 골격 형태: 현대 표범아과 (표범, 네오펠리스)와 유사[29]
7. 2. 식성

스밀로돈, 메간테레온, 파라마카이로두스 등 어두운 형태의 송곳니 그룹에 속하는 마카이로두스류는 튼튼함과 힘으로 특징지어진다. 이들은 짧은 중족골과 튼튼한 몸통을 가지고 있어 지구력이 좋은 주자는 아니었다. 현생 사자와 비교했을 때, 이들의 흉곽은 앞쪽이 좁고 뒤쪽이 넓어진 통 모양을 하고 있다. 견갑골은 특히 스밀로돈에서 잘 발달되어 강력한 어깨 근육과 삼두근이 부착될 수 있는 넓은 면적을 제공한다. 경추는 매우 튼튼하고 근육 부착 부위도 튼튼하다. 척추의 요추 부분은 단축되어 있다. 꼬리는 가장 원시적인 것에서 가장 진화된 것으로 갈수록 점점 짧아져, 그 결과 스밀로돈에서는 보브캣과 같은 꼬리가 되었다.[29]
마카이로두스, 미오마카이로두스, 호모테리움, 아델파일루스, 디노펠리스, 메틸루스, 폰토스밀루스, 테라일루스, 로코툰자일루루스, 제노스밀루스 등을 포함하는 시미터형 송곳니 그룹은 더 다양한 그룹이며, 대부분의 마카이로두스아과는 이 덜 특수화된 유형에 속한다. 이 그룹은 일반적으로 평균보다 날씬하고 작지만, 이 그룹의 마카이로두스는 모든 마카이로두스아과 중에서 가장 큰 것(최소한 가장 큰 것 중 하나)이다. 호모테리움은 현생 점박이하이에나와 매우 유사한 기울어진 등을 가지고 있으며, 마찬가지로 장거리 주행에 뛰어났을 수도 있다.[30] 그룹 전체적으로 긴 사지와 유연한 몸을 가지는 것이 일반적이었다. 마카이로두스는 도약의 명수였을지도 모른다.[12]
양호한 보존 상태의 화석화된 포식동물의 뼈에는 생존 시에 포식했던 종에 기인하는 식별 가능한 단백질이 보존되어 있는 경우가 있다.[84][85] 이러한 단백질의 안정 동위체 분석을 통해 스밀로돈은 주로 들소·말을 포식했으며, 경우에 따라 땅늘보나 매머드를 먹었고, 반면 호모테리움은 거의 매머드에만 의존했다는 것을 알 수 있었다.
7. 3. 얼굴
조지 밀러는 마카이로두스아과, 특히 스밀로돈의 연조직에 대해 이전에는 고려되지 않았던 특징들을 제시했다.[99]그가 제안한 첫 번째 외형 변화는 귀가 더 낮아졌다는 것이다. 이는 정확히 말하면 더 높은 시상 능선으로 인해 귀가 낮아 보이는 착시였다. 그러나 귀의 위치는 현대 고양이과 동물에서 항상 유사하며, 검치호와 비슷한 크기의 능선을 가진 개체에서도 마찬가지이다.[86] 귓바퀴의 위치는 털 색깔과 함께 복원하는 사람에 따라 달라진다. 크고 작고, 뾰족하거나 둥글고, 높거나 낮거나, 화석은 이러한 특징을 기록하지 않으므로 해석의 여지가 있다.
밀러는 또한 퍼그와 같은 코를 제안했다. 퍼그와 유사한 개를 제외하고는, 현대 육식동물 중 선택적 번식에서 비롯된 부자연스럽게 만들어진 특성인 퍼그 코를 가진 동물은 없다. 퍼그 코의 비교적 낮은 분포로 인해 일반적으로 무시되었다.[37] 밀러의 이론적 근거는 ''스밀로돈''의 코뼈가 뒤로 물러났다는 데 기반을 두고 있다. 밀러의 이론에 대한 비판은 사자와 호랑이의 코뼈를 비교하는 것이다. 호랑이와 비교했을 때 사자 또한 코뼈가 강하게 뒤로 물러나 있지만, 사자의 콧등은 호랑이보다 더 뒤로 물러나지 않았다. 따라서 밀러가 제안한 ''스밀로돈''의 퍼그 코는 유사한 동물의 신체 구조에서 증거가 거의 없다. Antón, García-Perea 및 Turner (1998)에 따르면, 살아있는 고양이과 동물의 콧구멍은 코뼈의 길이에 관계없이 항상 유사한 위치까지 확장되며, 이는 ''스밀로돈''에서 현대 종에서 관찰되는 범위 내에 있다.[87]
제안된 세 번째 아이디어는 입술을 50% 연장하는 것이다. 그의 다른 가설은 대부분 폐기되었지만, 마지막 가설은 현대 묘사에 크게 사용된다. 밀러는 입술이 길면 입을 더 넓게 벌려 먹이를 물 때 필요한 더 큰 탄성을 가질 수 있다고 주장한다. 이 주장은 과학계에서 논쟁의 대상이 되었지만, 그럼에도 불구하고 예술가들의 지지를 받고 있다. 과학적 비판은 현대 고양이, 특히 더 큰 종의 입술이 놀라운 탄성을 보이며, 입이 더 크게 벌어지더라도 일반적인 입술 길이가 적절하게 늘어날 것이고,[88] 살아있는 육식 동물에서 입술 선은 항상 교근보다 앞쪽에 있으며, ''스밀로돈''에서는 열육치 바로 뒤에 위치해 있다는 점을 지적한다.[89] 그럼에도 불구하고 ''스밀로돈'', ''마카이로두스'' 및 기타 종의 재구성은 종종 큰 개의 볼을 닮은 긴 입술을 가지고 있는 것으로 나타난다.
7. 4. 발성
스밀로돈과 사자의 설골을 비교한 결과, 스밀로돈과 다른 마카이로두스류도 현생 대형 고양이과 동물과 마찬가지로 포효할 수 있었을 것으로 생각된다.[90][91] 2023년 연구에 따르면, 마카이로두스류는 "포효하는" 고양이와 같은 수의 설골을 가지고 있었지만, 그 형태는 "가르랑거리는" 고양이와 더 유사했다.[42][40][41]8. 사회적 행동
마카이로두스아과의 사회적 행동에 대해서는 여전히 논란이 많다.
스밀로돈은 라 브레아 타르 핏에서 발견된 화석을 통해 사회적이었을 가능성이 제기되었다. 그러나 스밀로돈의 뇌가 비교적 작다는 점은 집단 사냥과 같은 복잡한 협동 행동에 어려움이 있었을 것이라는 반론을 제시한다.[98]
호모테리움은 텍사스 주 프리센한 동굴에서 어린 맘모스 유해와 함께 발견되어, 집단 사냥을 했을 가능성이 제기되었다. 이들은 뛰어난 야간 시력을 바탕으로 밤에 사냥을 했을 것으로 추정된다.[44]
마카이로두스는 화석 증거가 부족하지만, 송곳니 손상 비율이 높고 치유 흔적이 발견되어 사회적이었을 가능성이 제기되기도 한다.
8. 1. 스밀로돈
2009년, 남아프리카 공화국과 탄자니아의 보호 구역에 있는 사회성 식육목과 단독성 식육목의 비율을 홍적세 화석층으로 유명한 라 브레아 타르 핏 화석의 비율과 비교하여[92], 죽어가는 먹이의 녹음된 소리에 어떻게 반응하는지를 조사한 연구를 통해 스밀로돈이 사회성이었는지 단독성이었는지를 판단하였다. 과거 라 브레아 타르 핏은 깊은 타르로 이루어져 있었고, 그곳에 동물들이 갇혔다. 갇힌 동물들이 죽어갈 때, 그들의 울음소리가 육식동물을 끌어들였고, 그 결과 그들도 갇히게 되었다. 이곳은 수많은 동물들을 타르에 의해 포획하고 보존해 온, 북미에서 가장 상태가 좋은 홍적세 화석층으로 여겨지며, 이 연구의 상황과 유사했을 수 있다. 단독성 식육목은 다른 포식자와 마주칠 위험이 있기 때문에 그러한 소리의 발생원에 접근하지 않을 것이라는 가정이 있었다. 사회성 식육목(예: 사자)은 두려워할 다른 포식자가 거의 없기 때문에 즉시 그러한 부름에 접근할 것이다. 이 연구에서는 라 브레아 타르 핏에서 발견되는 동물들의 비율이 후자의 상황과 잘 일치하며, 따라서 스밀로돈은 아마 사회성이었을 것이라고 결론 내렸다[92]。라 브레아 타르 핏에서 발견된 사냥 시 부상을 입은 스밀로돈 화석은 사회성을 보여주는 고병리학적 증거이다. 사냥 시의 삠에 기인하는 부상 외에도, 더 심한 부상은 사회성을 강력하게 시사한다. 부상자는 부상이 나을 때까지 오랫동안 장애를 가진 채로, 부어오른 발목을 질질 끌며 걷고, 몇 년 동안 행동에 제한이 있었을 것으로 생각된다[90]。어떤 예에서는 골반을 복잡 골절한 젊은 개체가 치유된 건이 알려져 있다. 그 개체는 부상당한 다리를 거의 사용할 수 없었고, 다른 세 다리로 느릿느릿 기어갈 뿐, 분명히 단독으로는 사냥을 할 수 없었다[97]。만약 단독성 포식자가 그러한 중상을 입고 살아남았다면, 그것은 매우 드문 일이었음에 틀림없다. 몇 달 동안 같은 장소에서 움직이지 못하고, 먹을 것을 가져다주거나, 동료가 쓰러뜨린 먹이까지 기어가서 간신히 살아남았다고 생각하는 것이 훨씬 더 타당할 것이다.
8. 2. 호모테리움

텍사스 주 프리센한 동굴에서 거의 400마리에 달하는 어린 맘모스의 유해와 함께 ''호모테리움''의 골격이 발견되었다. ''호모테리움'' 집단은 어린 맘모스를 사냥하는 데 특화되었으며, 사냥감을 탁 트인 곳에서 벗어나 은밀한 동굴로 끌고 들어가 먹었을 것으로 추정된다.[44] 이들은 뛰어난 야간 시력을 유지했으며, 북극 지역에서 밤에 사냥하는 것이 주요 사냥 방법이었을 것이다.[44]
현대 사자는 많은 수로 약해진 성체 코끼리와 건강한 아성체 코끼리를 죽일 수 있으므로, 비슷한 크기의 ''호모테리움''도 어린 맘모스를 상대로 같은 행동을 했을 것으로 추정된다.[45] 이는 동위원소 분석으로 뒷받침된다. 하지만 매우 크고 사회적이었을 수도 있는 고양이가 약 181.44kg (약 181kg)의 맘모스 새끼를 이빨을 손상시키지 않고 동굴 안으로 끌고 들어갈 수 있었을지에 대해서는 큰 비판이 제기되었다. 뒤쪽이 경사진 척추와 강력한 요추는 곰과 유사한 체형을 시사하므로, 무게를 끌 수 있었을 수도 있지만, ''마카이로두스''와 ''스밀로돈''에게서 종종 나타나는 송곳니 파손은 ''호모테리움''에서는 관찰되지 않는다.
어린 맘모스의 뼈는 ''호모테리움'' 앞니의 뚜렷한 자국을 보여주며, 이는 이들이 사체의 대부분의 고기를 효율적으로 처리할 수 있었음을 나타낸다. 또한 사체를 동굴로 끌고 들어간 것은 청소 동물이 아니라 이들이었음을 나타낸다.[46] 뼈 검사 결과, 이 고양이들은 사냥감을 안전한 지역으로 옮기고 청소 동물로부터 힘들게 얻은 식사를 빼앗기지 않기 위해 사체를 끌고 가기 전에 해체했다는 것을 시사한다.[47] 또한 고양이들이 뼈에서 살점을 효과적으로 발라내어 눈에 띄는 자국을 남겼다는 증거도 있다.[47]
8. 3. 반론
마카이로두스는 화석에서 사회성을 나타내는 증거가 거의 없지만, 송곳니가 다른 종에 비해 손상된 비율이 높고, 광범위한 치유 흔적도 보인다. Babiarz Institute of Paleontological Studies에 소장된 중국산 수컷 ''암피마카이로두스 giganteus'' 화석은 손상 후에도 사용되어 마모된 송곳니를 가진 노령 개체이다. 그러나 이 개체는 사회성 포식자였다면 치료 기회가 있었을 심각한 부비동염으로 사망했으며, 이 사실은 이 화석의 해석에 몇 가지 다른 설을 가져왔다.[96] 마카이로두스 새끼의 영구치 송곳니가 돋아나는 데는 오랜 시간이 걸리고, 그 전까지는 어미의 돌봄에 의존한다.라 브레아 타르 핏에서 발견된 사냥 시 부상을 입은 스밀로돈 화석이 많다는 점은 사회성을 시사하는 고병리학적 증거이다. 사냥 시의 삠으로 인한 부상 외에도, 더 심한 부상은 사회성을 강력하게 시사한다. 부상자는 부상이 나을 때까지 오랫동안 장애를 가진 채로, 부어오른 발목을 질질 끌며 걷고, 몇 년 동안 행동에 제한이 있었을 것으로 생각된다.[90] 어떤 예에서는 골반을 복잡 골절한 젊은 개체가 치유된 건이 알려져 있다. 그 개체는 부상당한 다리를 거의 사용할 수 없었고, 다른 세 다리로 느릿느릿 기어갈 뿐, 단독으로는 사냥을 할 수 없었다.[97] 만약 단독성 포식자가 그러한 중상을 입고 살아남았다면, 그것은 매우 드문 일이었을 것이다. 몇 달 동안 같은 장소에서 움직이지 못하고, 먹을 것을 가져다주거나, 동료가 쓰러뜨린 먹이까지 기어가서 간신히 살아남았다고 생각하는 것이 훨씬 더 타당하다.
마카이로두스아과가 사회성을 가졌는지에 대해서는 여전히 논란이 많다. 스밀로돈이 전통적인 단독 생활을 했다는 설을 지지하는 증거는 뇌에서 발견된다. 사람, 회색늑대, 사자 등 대부분의 사회적 포식자는 동종의 단독 생활 포식자보다 약간 더 큰 뇌를 가지고 있다. 스밀로돈은 오히려 비교적 작은 뇌를 가지고 있어, 집단 사냥과 같은 복잡한 협동 행동을 하지 못했을 것이라는 점이 시사되었다.[98]
골절 흔적 또한 사회성을 시사하는 것처럼 보이지만, 단독 생활을 하는 동물이 중상을 입고 회복하는 과정에 대한 가장 좋은 해석은, 충분한 영양을 비축해두었고 필요에 따라 사용할 수 있었다는 것이다. 치타가 그 반례처럼 보이지만, 다른 고양이과 동물보다 가냘픈 체격을 갖도록 특화된 종이기 때문이다. 사자나 표범과 같이 더 크고 건장한 체격의 고양이과 동물은 턱 골절이나 근육 파열과 같은 중상에서도 회복된 사례가 있다.
9. 송곳니의 기능
마카이로두스아과의 송곳니 기능에 대해서는 여러 가설이 제기되어 왔다.
- 찌르기: 마카이로두스아과는 큰 송곳니를 마치 칼처럼 사용하여 먹이의 피부와 살갗에 구멍을 냈을 것이라는 주장이 있었으나,[51] 치아 에나멜이 약하고 뼈에 쉽게 부러질 수 있으며, 아래턱 구조상 효과적인 찌르기가 어려웠을 것이라는 반론에 의해 과학계에서 부정되었다.[99]
- 성적 특징: 송곳니가 사자의 갈기처럼 성 선택의 결과로, 구애, 성적 과시, 사회적 지위를 위해 사용되었을 수 있다는 주장이 있다.[52] 그러나 마카이로두스 속 종은 암수 모두 송곳니를 가졌고 크기 차이도 거의 없다는 점에서 반박된다.[51]
- 사체 섭취: 대부분의 마카이로두스류가 청소동물(사체를 먹는 동물)이었을 것이라는 추측도 있다. 뛰어난 후각과 청력으로 사체를 찾거나 다른 포식자의 사냥감을 훔쳤을 수 있으며, 튼튼한 체형으로 보아 빨리 달릴 필요가 없었을 것이다.[51] 찰스 R. 나이트가 묘사한 라 브레아 타르 핏 동물군 그림에는 기회주의적 청소동물 역할을 하는 두 마리의 ''스밀로돈''이 등장한다.
- 목 물기: 가장 널리 받아들여지는 가설로, 현대 고양잇과 동물처럼 목을 물어 기도를 압박해 질식시키는 방식이다.[51] 이 가설에는 다음과 같은 변형이 있다.
- 일반적인 물기: 먹이를 제압한 후 목을 물어 혈관을 손상시키고 과다 출혈로 죽게 한다.[54]
- 물고 후퇴하기: 먹이를 물어 상처를 입힌 후, 출혈로 죽을 때까지 기다린다.
- 물고 압박하기: 송곳니로 기관을 압박하여 질식시키고, 혈관 손상으로 출혈을 유발한다.
- 정확한 절단 물기: 한 번의 물기로 목의 주요 혈관 4개를 모두 절단하여 빠르게 죽인다.[12]
- 배 가르기: 1985년 윌리엄 에이커스턴이 제안한 가설로, 하이에나처럼 먹이의 복강을 물어뜯어 찢어 열어 과다 출혈로 죽이는 방식이다.[56][57] 이 방법은 사회적 행동을 전제로 하며, 송곳니가 부러질 위험이 적다는 장점이 있다.[58][59]
9. 1. 찌르기 (Stabbing)
마카이로두스아과는 사냥하는 동안 큰 어금니를 사용해 동물을 잡고 입을 벌린 다음 머리를 충분한 힘으로 내려쳐 동물의 피부와 살갗에 구멍을 냈을 것이라고 제안되어 왔다. 한때는 큰 어금니가 마치 칼처럼 사용되었을 것이라고 제안되었다.[51] 송곳니는 처음에는 척추를 부수거나 글립토돈과 같이 갑옷으로 덮인 동물을 찢는 데 사용되는 강력하고 파괴적인 도구처럼 보였다.그러나 치아는 지지되지 않는 에나멜로 만들어졌으며, 뼈와 같은 단단한 물질에 쉽게 부러졌을 것이다. 또한 하악골과 입을 크게 벌릴 수 없는 점이 효과적인 찌르기를 방해했을 것이라는 주장도 제기되었다.[51] 이러한 이유로, 이 개념은 과학계에서 거부되었다.
마카이로두스아과 및 기타 검치호(剣歯虎) 고양이류의 매우 긴 송곳니 사용법에 대해서는 다양한 설이 제기되고 있다. 이 설은 송곳니를 찔러 넣는 데 사용했다는 것이다. 마카이로두스아과는 먹이에 달라붙어 입을 벌리고, 목을 휘둘러 먹이의 피부와 살을 관통할 정도의 힘을 실어 머리를 내리쳤을 것이다. 한때 이 검치호의 이는 사람이 손으로 칼을 다루는 것과 같은 방식으로 사용되었다고 생각되기도 했다.[99] 이 송곳니는 언뜻 보기에는 척추를 파괴하거나 글립토돈과 같은 동물의 갑옷을 찢는 데에도 사용할 수 있는 매우 유용하고 강력한 도구처럼 보인다. 마카이로두스류를 다룬 초기 논문의 대부분은 먹이를 그렇게 죽였다고 한다. 그것은 일반에도 널리 받아들여져 주요 설이 되었고, 그림이나 영화에서도 그렇게 그려졌다.
그러나, 이는 금속이 아닌 지지대가 없는 에나멜질로 만들어져 뼈와 같은 단단한 소재에 닿으면 쉽게 부러진다. 또한, 아래턱이 효과적인 자상(刺傷)의 장애가 된다는 점도 논의되었다.[99] 이러한 이유로, 자상설은 과학계에서 부정되었다.
9. 2. 성적 특징 (Sexual characteristic)
마카이로두스아과의 길고 큰 송곳니는 성 선택의 결과일 수 있으며, 사자의 갈기와 마찬가지로 구애, 성적 과시 및 사회적 지위를 위해 사용되었을 것으로 보인다. 송곳니는 상대적으로 약하고 턱 근육이 강하지 않아 포식 기능은 불확실하다.[52]하지만 성적 매력을 높이기 위해 특성이 채택될 때, 일반적으로 수컷을 포함한 한쪽 성별만 이 특징을 나타낸다. 모든 마카이로두스 속 종에서 수컷과 암컷 모두 이러한 송곳니를 가지고 있으며, ''마카이로두스''와 같은 사소한 예외를 제외하고는 모양이 비슷하다. 일반적으로 성별 간의 크기 차이도 있지만, 수컷과 암컷 마카이로두스는 같은 크기였던 것으로 보인다. 또한, 개인의 식사와 일반적인 기능에 심각한 손상을 남길 수 있다는 점을 감안할 때, 이러한 수준의 성 선택은 극단적인 것으로 보인다.[51]
마카이로두스아과 및 기타 검치호(검치호랑이) 고양이과 동물의 매우 긴 송곳니 사용법에 대해서는 다양한 설이 제기되고 있다. 그 중 하나는 이 이빨이 실제로 사냥에 사용되었는지에 대한 의문을 제기하고, 성 선택 때문이 아니냐는 논의이다. 사자의 갈기와 같은 몇몇 특징은 수컷 간의 경쟁과 암컷이 독자적인 판단 기준에 따라 잠재적이고 건강한 후보 중에서 배우자를 선택함으로써 진화해 왔다.
마카이로두스류에서 이 긴 송곳니에 대한 선택은 사슴의 뿔이나 사자의 갈기와 유사한 것이라고 생각할 수 있으며, 마카이로두스류는 구애, 성적 과시, 사회적 지위를 위해 사용된 특징을 성 선택을 통해 진화시키도록 유도되었다. 이들은 아마도 구애라는 게임을 위해 기묘한 특징을 진화시킨 그룹 중 하나일 뿐이며, 그들의 송곳니에는 특별히 기능적인 목적이 없을 수도 있다. 송곳니는 충분히 발달했지만, 다소 부서지기 쉽고, 턱 근육은 강하지 않았기 때문에, 실제 기능적인 용도에 대해서는 명확하지 않은 것으로 보인다.[100]
이 설에 대한 중요한 논쟁이 몇 가지 진행되었다. 성적 매력을 강화하기 위해서만 그러한 기능성과 무관한 특징을 가진 종의 대부분은 한쪽 성별(보통 수컷)에만 그 특징이 나타난다. 마카이로두스류의 종은 모두 수컷과 암컷 모두 그러한 송곳니를 가지고 있으며, 마카이로두스를 예외로 한다면 체격도 동일하다. 성적인 특징을 고도로 발전시켜온 대부분의 종에서는 적어도 또 다른 신체적 차이가 보인다. 그러한 종에서는 보통 체격이 다르다. 수컷 사슴은 뿔을 가질 뿐만 아니라, 일반적으로 암컷보다 몸집이 크다. 수컷 사자는 갈기를 가질 뿐만 아니라, 암컷보다 크다. 미국황조롱이의 수컷은 짝인 암컷보다 밝고 빛나는 체색을 가지고 있을 뿐만 아니라, 암컷보다 작다. 마카이로두스류의 수컷과 암컷은 같은 크기이다. 이러한 특징을 성적 과시의 목적으로만 발달시켜 왔다면, 마카이로두스류는 식사나 일반적인 기능에 매우 큰 장애가 있었을 것이다.[99]
9. 3. 사체 섭취 (Scavenging)
대부분의 마카이로두스류가 청소동물(사체를 먹는 동물)이었을 것이라는 추측이 있다. 이는 송곳니가 대부분 기능을 하지 못한다는 점을 설명하며, 종종 성 선택 가설과 연결된다. 많은 현대 육식동물들은 정도의 차이는 있지만 사체 섭취를 한다. 뛰어난 후각과 청력은 사체를 찾거나 다이어울프나 북극곰과 같은 다른 포식자들의 사냥감을 훔치는 데 도움이 되었을 수 있으며, 대부분의 마카이로두스류에서 보이는 튼튼한 체형을 볼 때, 질주할 필요는 없었을 것이다.[51]- -]
많은 현대 고양이들이 이러한 특성들을 혼합하여 보여준다. 사자는 튼튼한 사냥꾼이지만 기회가 있으면 훔치기도 한다. 호랑이와 퓨마는 사냥감을 묻어두고 며칠이 지난 후에도 다시 돌아와 먹이를 먹는다. 모든 고양이는 아프거나 부상당한 동물을 사냥하는 것을 선호하며, 움직일 수 없는 아픈 동물과 죽은 동물 사이에는 미묘한 차이가 있다. 캘리포니아 라 브레아 타르 핏에서 발견된 ''스밀로돈''의 풍부한 골격 역시 이 가설을 뒷받침한다. 웅덩이에 갇힌 동물들은 죽어가거나 죽었을 것이며, 이는 현대의 치타와 같은 진정한 초육식 동물이 외면할 종류의 먹이일 것이다. 이 가설은 가장 오래되었지만 여전히 타당한 것으로 간주된다.
이 개념에 대한 반론은 고양이의 여러 부분에서 찾아볼 수 있다. 이빨은 순전히 육식성이며,[51] 개와 곰의 잡식성 이빨처럼 식물 재료를 갈 수 없다. 절치는 뼈를 부수는 것이 아니라 살을 효율적으로 베도록 모양이 잡혀 있는데, 이는 현대의 점박이하이에나와 다르다. 두 성별 모두 이러한 송곳니를 가지고 있고 두개골에 추가적인 변형이 존재하기 때문에, 마카이로두스류는 어느 정도 기회주의적이었을 것이다.
9. 4. 목 물기 가설 (Neck-biting hypotheses)
마카이로두스아과의 사냥 방식에 대해 가장 널리 받아들여지는 가설은 목을 물어뜯는 것이다. 현대 고양잇과 동물들은 목 윗부분을 물어 기도를 압박하여 먹이를 질식시키는 목 조르기를 사용한다.[51] 이들의 송곳니는 주로 피부를 뚫고 먹이를 붙잡는 데 사용되며, 먹이에게 큰 상처를 입히지는 않는다. 반면 마카이로두스아과는 이와 같은 기술을 사용했다면 큰 피해를 입혔을 것이다.[53]이 방식의 주요 단점은 많은 양의 피가 흘러나와 주변의 다른 육식동물, 예를 들어 다른 마카이로두스아과 동물이나 다이어 울프에게 냄새를 맡게 할 수 있다는 것이다. 포식자들은 종종 경쟁 관계를 형성하며, 아프리카의 현대 사자와 점박이하이에나에서 볼 수 있듯이 지배력이 한 종에서 다른 종으로 바뀔 수 있다. 이러한 상황에서 싸움은 드물지 않다. 이러한 최상위 포식자들 사이의 권력 균형과 지배력은 사회적 요인 때문에 여전히 미스터리로 남아있다. 수적으로 우세한 것은 이러한 싸움에서 중요할 수 있다. 예를 들어, 다이어 울프는 작은 무리를 지어 이동한 것으로 여겨지며, 개별적으로는 약했지만 그들의 수는 마카이로두스아과가 사냥한 먹이를 빼앗기에 충분했을 수 있다.
그러나 고양잇과 동물은 다이어 울프가 사냥한 먹이를 먹고 살았을 수도 있다. 두 마리의 고독한 마카이로두스아과는 빠르게 서열을 형성하여 첫 번째 개체가 지배적이 될 것이다. 이러한 불확실성 때문에 마카이로두스아과의 생태적 지위의 많은 부분이 아직 알려지지 않았다. 이 가설에 대한 여러 변형은 모두 억제되고 움직이지 않는 동물을 필요로 한다.
목 물기 가설의 변형
- 일반적인 물기: 마카이로두스아과는 동물을 제압한 다음, 위치에 크게 구애받지 않고 목을 물어 주요 혈관에 손상을 입힌 후, 동물이 피를 흘려 죽도록 내버려 둔다. 척추와의 접촉으로 이빨이 부러질 수 있는 목 뒤쪽을 물지 않고, 목의 어느 곳이든 깊게 물면 치명적이라는 것이다.[54] 이러한 일반적인 물기는 물 수 있는 모든 곳에 사용되었을 것이며, 더 적은 수의 포식자를 필요로 한다. 한 마리의 ''메간테레온''은 큰 사슴, 심지어 말도 송곳니가 부러질 위험 없이 죽일 수 있었다. 육식 동물이 먹이 뒤에 몸을 유지하면서 물 수 있기 때문이며, 다리를 휘두르는 것을 피하면서도 몸무게로 압박하여 움직이지 못하게 할 수 있기 때문이다. 이는 매복 사냥에 적합한 빠른 물기였을 것이며, 대부분의 마카이로돈트의 크고 강한 몸집으로 유추할 수 있다. 혼자 있는 마카이로돈트가 이러한 방식으로 큰 먹이 동물을 부상시킨 다음, 풀어주고 충격으로 쓰러질 때까지 따라가는 것도 가능했을 것이다.
- 물고 후퇴하기: 피가 많이 나는 점과, 발버둥치는 먹이가 그 지역의 모든 포식자와 청소 동물을 끌어들였을 것이라는 점 때문에 비판받아왔다. 단독 동물이 먹이 동물을 물어 부상시킨 후 풀어주고 따라갔을 것이라는 생각은 더욱 강력하게 반박되어 왔다. 고양잇과 동물은 배를 채울 때까지 먹이로부터 거의 떠나지 않으며, 다른 포식자에게 빼앗길 위험이 있었을 것이다.
- 물고 압박하기: 마카이로두스가 물어 뜯고 압착하는 방식은 다음과 같다. 마카이로두스의 이빨에 물린 동물이 상처를 입었을 때, 송곳니는 기관 뒤에 삽입되고 앞 어금니는 기관을 감쌌을 것이다. 이 방식은 마카이로두스가 물어뜯은 후 기관을 압착하여 먹잇감을 질식시키고 상처를 입히는 역할을 했다는 것이다. 목의 큰 혈관을 뚫어 대량 출혈을 일으키면 동물의 죽음을 재촉했을 것이다. 현대 고양이, ''Pseudaelurus''와 ''Proailurus''와 같은 모든 고양이의 기저 속들은 먹잇감을 처리하는 일반적인 방법으로 목 조르기를 사용한다. 질식은 당황한 먹잇감의 소리를 억제하는데, 이는 현대 치타와 표범이 사용하는 방법이다. 송곳니에서 생긴 상처와 공기 부족은 먹잇감을 죽게 했을 것이다. 이 방법은 송곳니에 의해 만들어진 상처의 완전한 효과를 방해할 수 있다. 송곳니를 상처에 계속 유지하면 신체의 혈류를 막아 먹잇감이 소리를 낼 수 없더라도 더 오래 살아있게 할 수 있다. 짧고 원뿔 모양의 송곳니를 가진 조상 고양이와 비교했을 때, 더 긴 송곳니가 갖는 뚜렷한 이점이 없다. 잘 제압된 상태에서도 당황한 동물의 목에 이빨이 부러질 위험이 잠재적인 이점보다 크기 때문에, 개연성이 없는 것으로 간주된다.
- 정확한 절단 물기: 진화된 검치호랑이들은 매우 특화되어 있어서, 한 번의 물기로 먹잇감 동물의 목에 있는 4개의 주요 혈관을 관통할 수 있는 특정한 기하학적 구조를 갖추고 있었다.[12] 물고 압박하는 방식의 가설과 유사하지만, 더 정확하게 혈관을 관통하는 조심스러운 물기를 포함하며, 검치호랑이는 물고 나서 물러나 먹잇감이 매우 빠르게 출혈로 죽도록 내버려 두었을 것이다. 피가 많이 났겠지만, 이 방법은 동물을 죽이는 데 가장 짧은 시간이 걸렸을 것이다. 검치호랑이가 사냥했을 가능성이 있는 종들 간의 해부학적 차이 때문에, 예를 들어 말을 죽이는 데 필요한 기하학적 구조는 들소를 죽이는 데 효과가 없을 수 있다. 이는 속, 심지어 특정 종이 하나의 먹잇감 동물 유형에 매우 특화되어야 함을 의미한다. 이는 그들의 멸종에 대한 설명을 제공할 수 있는데, 먹잇감 종의 이동이나 멸종은 그 전문 포식자의 죽음으로 이어질 수 있기 때문이다.
9. 5. 배 가르기 (Belly shearing)
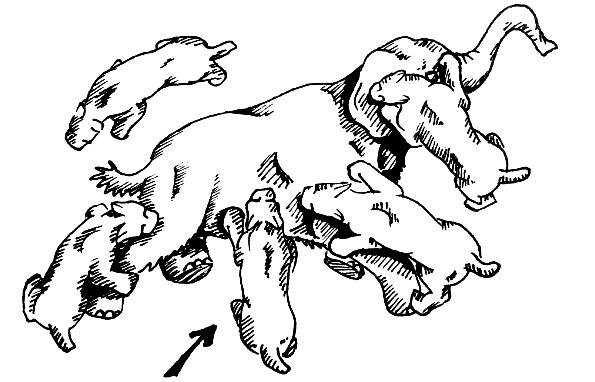
1985년, 미국의 고생물학자 윌리엄 에이커스턴은 절단 물기(shearing bite)를 제안했다.[56][57] 이 살해 방식은 오늘날 하이에나와 개과 동물에게서 볼 수 있는 것과 유사하다. 마카이로두스 무리가 먹잇감을 포획하여 완전히 제압한 후, 무리 중 한 마리가 복강을 물어뜯어 몸을 찢어 열었다.이 기술은 특정 동작 순서를 따른다. 먼저 동물을 완전히 제압해야 하며, 여러 개체가 먹잇감을 붙잡기 위해 마카이로두스는 사회적이어야 한다. 살해 물기를 가할 개체는 입을 최대한 벌리고, 아래턱으로 배의 피부를 위로 눌러 올린다. 아래 송곳니와 앞니가 피부를 누르는 곳에 움푹한 곳을 만들고, 아래턱을 위로 밀면서 아래 치아 위 피부에 약간의 주름을 만든다. 다음으로, 위 송곳니를 피부에 누르고 목 근육을 사용하여 머리를 누른다. 턱을 '위로' 당기는 대신, 두개골을 '아래로' 누르는 것이다. 송곳니가 피부를 뚫으면, 입을 45° 정도 벌릴 때까지 낮추고, 두개골을 계속 누르면서 아래턱을 위로 당긴다. 대부분의 마카이로두스 아래턱 앞부분의 작은 플랜지는 두개골을 누르는 데 사용된다. 동물의 입이 닫히면 송곳니 뒤 턱 사이에 두꺼운 피부 조각이 잡히고, 동물은 허리와 앞다리 근육을 사용하여 뒤로 당겨 피부 조각을 몸에서 뜯어낸다. 이 틈새가 열리면 내장이 노출되고 동맥과 정맥이 찢어진다. 피를 흘리는 동물은 몇 분 안에 죽고, 반복적인 물기로 몸에서 내장이 찢어지는 충격이 과정을 가속화할 수 있다.[58]
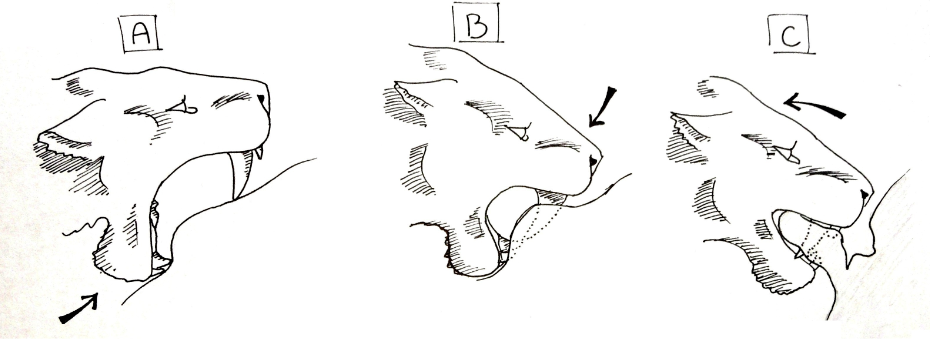
이 방법은 사회적 마카이로두스가 먹잇감에게 큰 상처를 입힐 수 있게 한다. 대량 출혈이 발생하며, 사회적 집단은 그 지역에 끌리는 거의 모든 동물을 막을 수 있다. 물기는 특정할 필요가 없으며, 동물의 죽음을 재촉하기 위해 반복될 수 있다. 이는 점박이하이에나와 같은 몇몇 현존하는 종의 살해 방법에서 이미 관찰되고 있다. 송곳니는 목에 비해 배의 부드러운 특성 때문에 부러질 가능성이 적으며, 목에서처럼 배에서 갑작스러운 움직임이 증폭되지 않는다. 복부 절단 가설은 일반적으로 매우 그럴듯한 것으로 간주된다. 라 브레아 타르 구덩이에서 ''스밀로돈''의 부러진 송곳니는 드물게 발생하며, 이 덜 위험한 방법이 이에 기여했을 수 있다.[59]
그러나 절단 물기는 마카이로두스에게 여러 문제가 되었을 수 있다. 대부분의 유제류는 배와 뒷다리 부위에 매우 민감하며, 대부분의 포식자는 머리와 앞다리를 조작하여 국내 소와 유사한 동물을 포획하고 제압하는 것이 더 쉽다고 생각한다. 동물을 땅에 눕히고 다리 사이에 위치함으로써, 마카이로두스는 발에 차일 위험이 컸다. 발차기의 힘은 이빨, 아래턱, 다리를 부러뜨리고 고양이를 불구로 만들거나 죽일 수 있다.
사회성은 다른 개체들이 동물을 붙잡고 있는 동안 한 개체가 살해 물기를 가함으로써 이 문제를 해결했을 수 있다. 게다가, 바이슨과 같은 큰 유제류의 복부 직경이 너무 클 수 있고, 피부가 너무 팽팽해서 마카이로두스가 피부 조각을 잡을 수 없을 뿐만 아니라 몸에서 뜯어낼 수 없을 수도 있다. 절단 물기의 세 번째 문제는 송곳니가 동물의 배에 큰 구멍을 뚫어야 하지만, 대신 피부를 벗겨내고 두 개의 긴 틈새를 만들 수 있다는 것이다. 이 상처는 고통스럽고 피를 흘리게 하지만, 동물은 피를 흘려 죽을 가능성은 낮고 탈출하여 생존할 수 있다.
2004년 실험에서는 라 브레아 타르 구덩이에서 나온 ''스밀로돈 파탈리스''의 CT 스캔을 통해 주조된 한 쌍의 기계 알루미늄 턱을 사용하여 신선한 국내 소 사체에 절단 물기를 포함하여 ''스밀로돈''이 사용할 수 있는 여러 물기 기술을 시뮬레이션했다.[60] 소의 배는 송곳니가 피부를 뚫을 수 없을 정도로 직경이 컸고, 대신 턱이 접근을 막으면서 몸에서 빗나갔다. 그러나 모델은 현대 고양이가 물듯이 턱을 위로 당겼지만, 마카이로두스는 목 근육으로 두개골을 아래로 눌렀을 것이므로 그렇게 하지 않았을 가능성이 크다. 이 절차의 결함은 결과를 무효화하고 복부 절단 가설을 손상되지 않은 채로 남겨둘 수 있다.
참조
[1]
논문
Fossils explained 52 Majestic killers: the sabre-toothed cats
2006-07
[2]
서적
Biology and Conservation of Wild Felids
https://www.research[...]
Oxford University Press
2010-01
[3]
서적
Mammoths, Sabertooths and Hominids 65 Million Years of Mammalian Evolution in Europe
Columbia University Press
2002
[4]
논문
A dwarf sabertooth cat (Felidae: Machairodontinae) from Shanxi, China, and the phylogeny of the sabertooth tribe Machairodontini
https://www.scienced[...]
2022-05-15
[5]
논문
Evolutionary History of Saber-Toothed Cats Based on Ancient Mitogenomics
2017-10-19
[6]
논문
New sabre toothed Felidae (Carnivora, Mammalia) in the hominid-bearing sites of Toros Menalla (late Miocene, Chad)
https://sciencepress[...]
2022-11-30
[7]
웹사이트
Paleo Profile: The Chad Cat
https://blogs.scient[...]
2018-04-05
[8]
논문
A New Felid from the Late Miocene of the Balkans and the Contents of the Genus Metailurus Zdansky, 1924 (Carnivora, Felidae)
[9]
웹사이트
Paleobiology Database
https://paleobiodb.o[...]
2011-06-16
[10]
논문
The evolution of the guild of larger terrestrial carnivores during the Plio-Pleistocene in Africa
[11]
논문
Three Ways To Be a Saber-Toothed Cat
[12]
서적
The Big Cats and their fossil relatives
https://archive.org/[...]
Columbia University Press
[13]
논문
A New Machairodont from the Palmetto Fauna (Early Pliocene) of Florida, with Comments on the Origin of the Smilodontini (Mammalia, Carnivora, Felidae)
[14]
논문
A New Felid from the Late Miocene of the Balkans and the Contents of the Genus Metailurus Zdansky, 1924 (Carnivora, Felidae)
[15]
웹사이트
Rhizosmilodon fiteae :: Florida Museum of Natural History
https://www.flmnh.uf[...]
2016-10-04
[16]
서적
Sabertooth
University of Indiana Press
2013
[17]
논문
First known complete skulls of the scimitar-toothed cat Machairodus aphanistus (Felidae, Carnivora) from the Spanish late Miocene site of Batallones-1
[18]
논문
Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat Paramachairodus ogygia (Kaup, 1832) (Felidae, Machairodontinae) from the Late Miocene of Spain: implications for the origins of the machairodont killing bite
[19]
논문
Machaeroides simpsoni, new species, oldest known sabertooth credont (Mammalia), of Lost Cabin Eocene
[20]
논문
Systematic revision of the late Miocene sabre-toothed felid Paramachaedrodus in Spain
[21]
서적
The big cats and their fossil relatives: an illustrated guide to their evolution and natural history
Columbia University Press
[22]
논문
Tough times at La-Brea – tooth breakage in large carnivores of the Late Pleistocene
[23]
논문
Costs of carnivory: tooth fracture in Pleistocene and Recent carnivorans
https://zenodo.org/r[...]
[24]
논문
Functional anatomy of the forelimb in Promegantereon ogygia (Felidae, Machairodontinae, Smilodontini) from the Late Miocene of Spain and the origins of the sabre-toothed felid model
[25]
논문
Evolution of the sabertooth mandible: A deadly ecomorphological specialization
https://linkinghub.e[...]
2018-05
[26]
논문
Mummy of a juvenile sabre-toothed cat ''Homotherium latidens'' from the Upper Pleistocene of Siberia
2024-11-14
[27]
논문
Evolution of Skull and Mandible Shape in Cats (Carnivora: Felidae)
[28]
웹사이트
Greg Laden's Blog: About
http://scienceblogs.[...]
[29]
논문
How to Build a Mammanian Super-Predator
2006-05-04
[30]
논문
Co-existence of scimitar-toothed cats, lions and hominins in the European Pleistocene. Implications of the post-cranial anatomy of Homotherium latidens (Owen) for comparative palaeoecology
https://www.scienced[...]
2005-05-01
[31]
웹사이트
Saber-Toothed Cat Had Wimpy Bite
http://www.livescien[...]
2011-07-05
[32]
웹사이트
Stable Isotopes in Archaeology
http://archaeology.a[...]
2011-09-10
[33]
웹사이트
Stable Isotopes and Diet: You Are What You Eat
http://luna.cas.usf.[...]
2011-09-10
[34]
서적
Sabertooth
2013
[35]
간행물
Sabertooth carcass consumption behavior and the dynamics of Pleistocene large carnivoran guilds
https://doi.org/10.1[...]
2022
[36]
논문
Reconstructed facial appearance of the sabretoothed felid Smilodon
[37]
서적
Sabertooth
University of Indiana Press
2013
[38]
논문
The smilodon's smile
http://www.nature.co[...]
1999-01-28
[39]
논문
Concealed weapons: A revised reconstruction of the facial anatomy and life appearance of the sabre-toothed cat ''Homotherium latidens'' (Felidae, Machairodontinae)
2022
[40]
웹사이트
Saber-Toothed Tales
http://discovermagaz[...]
1993-04-01
[41]
웹사이트
Saber-toothed Cat Sculpture
http://www.utexas.ed[...]
2011-09-10
[42]
논문
The roar of Rancho La Brea? Comparative anatomy of modern and fossil felid hyoid bones
[43]
논문
Parallels between playbacks and Pleistocene tar seeps suggest sociality in an extinct sabretooth cat, Smilodon
2009-02-23
[44]
웹사이트
Late Pleistocene Climate and Proboscidean Paleoecology
http://ir.lib.uwo.ca[...]
2011-09-10
[45]
서적
Sabertooth
University of Indiana Press
2013
[46]
서적
Sabertooth
University of Indiana Press
2013
[47]
웹사이트
The Diet of Saber-Toothed Cats
https://nimravid.wor[...]
2008-03-08
[48]
웹사이트
Sabertooth Cat, Chinese ''Machairodus giganteus'' Skull
http://www.boneclone[...]
2011-07-03
[49]
논문
Healed Massive Pelvic Fracture in a Smilodon from Ranco La Brea, California
http://anthropologie[...]
[50]
논문
Assessing behavior in extinct animals: was Smilodon social?
https://zenodo.org/r[...]
[51]
서적
Big Cats and their Fossil Relatives
Columbia University Press
[52]
웹사이트
Revised Repost: What big teeth you have
http://scienceblogs.[...]
ScientificBlogs
2012-10-05
[53]
논문
Sabretoothed Carnivores and the Killing of Large Prey
[54]
논문
Supermodeled sabercat, predatory behavior in ''Smilodon fatalis'' revealed by high-resolution 3D computer simulation
[55]
웹사이트
Xenosmilus hodsonae
https://www.floridam[...]
Florida Museum
2017-04-03
[56]
서적
Do Cats Hear with Their Feet?: Where Cats Come From, what We Know about Them
[57]
웹사이트
Saber-toothed cats
http://exhibits.muse[...]
2011-09-10
[58]
Youtube
Prehistoric Predators: Sabertooth Part 4
https://www.youtube.[...]
2016-11-26
[59]
서적
Monster of God: the man-eating predator in the jungles of history and the mind
https://archive.org/[...]
W. W. Norton & Company
[60]
논문
Simulated bite marks
[61]
웹사이트
Paleobiology Database: ''Machairodontinae'' Basic info
http://paleodb.org/c[...]
[62]
간행물
Fossils explained 52 Majestic killers: the sabre-toothed cats.
http://www.blackwell[...]
Blackwell Publishing Ltd
2006-07
[63]
서적
絶滅哺乳類図鑑
丸善
2002
[64]
웹사이트
A pdf biography of T.H. Gill
http://www.nasonline[...]
2016-05-01
[65]
웹사이트
Arrangement of the Families of Mammals
https://archive.org/[...]
2016-05-01
[66]
서적
Mammoths, Sabertooths and Hominids 65 Million Years of Mammalian Evolution in Europe
Columbia University Press
2002
[67]
논문
Fossils explained 52 Majestic killers: the sabre-toothed cats
2006-07
[68]
웹사이트
Paleobiology Database
http://www.paleodb.o[...]
[69]
논문
The evolution of the guild of larger terrestrial carnivores during the Plio-Pleistocene in Africa
[70]
논문
Three Ways To Be a Saber-Toothed Cat
[71]
문서
Turner, (1997) p60
[72]
논문
First known complete skulls of the scimitar-toothed cat Machairodus aphanistus (Felidae, Carnivora) from the Spanish late Miocene site of Batallones-1
[73]
논문
Aspects of the functional morphology in the cranial and cervical skeleton of the sabre-toothed cat Paramachairodus ogygia (Kaup, 1832) (Felidae, Machairodontinae) from the Late Miocene of Spain: implications for the origins of the machairodont killing bite
[74]
논문
Machaeroides simpsoni, new species, oldest known sabertooth credont (Mammalia), of Lost Cabin Eocene
[75]
논문
Systematic revision of the late Miocene sabre-toothed felid Paramachaedrodus in Spain
[76]
논문
Tough times at La-Brea – tooth breakage in large carnivores of the Late Pleistocene
[77]
논문
Costs of carnivory: tooth fracture in Pleistocene and Recent carnivorans
[78]
논문
Functional anatomy of the forelimb in Promegantereon ogygia (Felidae, Machairodontinae, Smilodontini) from the Late Miocene of Spain and the origins of the sabre-toothed felid model
[79]
서적
動物大百科1 食肉類
平凡社
[80]
웹사이트
Smilodon fatalis
https://www.flmnh.uf[...]
2016-05-01
[81]
논문
Evolution of Skull and Mandible Shape in Cats (Carnivora: Felidae)
http://www.plosone.o[...]
[82]
논문
How to Build a Mammanian Super-Predator
2006-05-04
[83]
웹사이트
Saber-Toothed Cat Had Wimpy Bite
http://www.livescien[...]
2016-05-01
[84]
웹사이트
Stable Isotopes in Archaeology
http://archaeology.a[...]
2016-05-01
[85]
웹사이트
Stable Isotopes and Diet: You Are What You Eat
http://luna.cas.usf.[...]
2016-05-01
[86]
문서
Antón, García-Perea, & Turner (1998), 369–386, esp. 377-378.
[87]
문서
Antón, García-Perea, & Turner (1998), 375-377.
[88]
웹사이트
The smilodon's smile
http://www.nature.co[...]
2016-05-01
[89]
문서
Antón, García-Perea, & Turner (1998), 375.
[90]
웹사이트
Saber-Toothed Tales
http://discovermagaz[...]
2016-05-01
[91]
웹사이트
Saber-toothed Cat Sculpture
http://tmm.utexas.ed[...]
2016-05-01
[92]
논문
Parallels between playbacks and Pleistocene tar seeps suggest sociality in an extinct sabretooth cat, Smilodon
2009-02-23
[93]
웹사이트
Late Pleistocene Climate and Proboscidean Paleoecology
http://ir.lib.uwo.ca[...]
2016-05-01
[94]
웹사이트
Elephant Kill at the Savute Safari Lodge
https://lionprides.w[...]
2016-05-01
[95]
웹사이트
Elephant Eaters of the Savuti
http://www.go2africa[...]
2016-05-01
[96]
웹사이트
Sabertooth Cat, Chinese ''Machairodus giganteus'' Skull
http://www.boneclone[...]
2016-05-01
[97]
논문
Healed Massive Pelvic Fracture in a Smilodon from Ranco La Brea, California
http://anthropologie[...]
[98]
논문
Assessing behavior in extinct animals: was Smilodon social?
[99]
문서
Turner, (1997)
[100]
웹사이트
Revised Repost: What big teeth you have
http://scienceblogs.[...]
ScientificBlogs
2016-05-01
[101]
논문
Sabretoothed Carnivores and the Killing of Large Prey
[102]
논문
Supermodeled sabercat, predatory behavior in ''Smilodon fatalis'' revealed by high-resolution 3D computer simulation
http://www.pnas.org/[...]
[103]
서적
Do Cats Hear with Their Feet?: Where Cats Come From, what We Know about Them
[104]
웹사이트
Saber-toothed cats
http://exhibits.muse[...]
2016-05-01
[105]
서적
Monster of God: the man-eating predator in the jungles of history and the mind
[106]
웹사이트
Simulated bite marks [digital simulation]
http://ieeexplore.ie[...]
2016-05-01
[107]
저널
The Evolution of CATS
http://sehrg.at.ua/_[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com