아그로박테리움
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
아그로박테리움은 식물에 혹병을 일으키는 세균 속으로, 유전자를 식물이나 균류에 전달하는 능력을 활용하여 생명공학, 특히 식물 개량에 사용된다. 1990년대까지는 쓰레기통 분류군으로 여겨졌으나, 16S rRNA 염기 서열 분석을 통해 분류가 재정립되었다. 아그로박테리움은 Ti 플라스미드에 포함된 T-DNA를 식물 세포에 전달하여 종양을 유발하며, 생명공학 기술을 통해 식물 유전자 조작에 활용된다. 최근에는 면역력이 약한 사람에게 기회 감염을 일으킬 수 있다는 연구 결과도 보고되었다.
더 읽어볼만한 페이지
| 아그로박테리움 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Agrobacterium |
| 최초 명명자 | Conn 1942 (Approved Lists 1980) |
| 모식종 | 아그로박테리움 라디오박테르 |
| 모식종 명명자 | (Smith and Townsend 1907) Conn 1942 (Approved Lists 1980) |
| 종 | |
| 이명 | |
2. 명명 역사
1990년대까지 쓰레기통 분류군으로 사용되던 ''Agrobacterium''(아그로박테리움) 속은 16S 염기 서열 분석으로 인해 여러 속으로 재분류되었다.[2][3] 2000년대 초, ''Agrobacterium''은 ''Rhizobium''(라이조비움) 속과 동의어가 되었으나,[4] 이는 논란을 일으켰다.[5][6] 이후 ''Agrobacterium'' 속이 ''Rhizobium''과 계통발생학적으로 뚜렷하게 구분된다는 것이 증명되면서 ''Agrobacterium'' 속이 복원되었다.[7][8][9]
2. 1. 생물변종의 변화
1990년대까지 ''아그로박테리움''(Agrobacterium) 속은 쓰레기통 분류군으로 사용되었다. 16S 염기 서열 분석이 나오면서, 많은 ''Agrobacterium'' 종(특히 해양 종)은 ''Ahrensia''(아렌시아), ''Pseudorhodobacter''(슈도호도박터), ''Ruegeria''(뢰게리아), ''Stappia''(스타피아) 속과 같은 속으로 다시 분류되었다.[2][3] 남아있는 ''Agrobacterium'' 종은 세 개의 생물변종으로 분류되었는데, 생물변종 1 (''Agrobacterium tumefaciens''(아그로박테리움 투메파시엔스)), 생물변종 2 (''Agrobacterium rhizogenes''(아그로박테리움 라이조제네스)), 그리고 생물변종 3 (''Agrobacterium vitis''(아그로박테리움 비티스))이었다. 2000년대 초, ''Agrobacterium''은 ''Rhizobium''(라이조비움) 속과 같은 속으로 취급되었다.[4] 이러한 움직임은 논란을 일으켰다.[5][6] 이 논쟁은 ''Agrobacterium'' 속이 ''Rhizobium''과 계통발생학적으로 뚜렷하게 구분된다는 것이 증명된 후 ''Agrobacterium'' 속이 복원되면서 최종적으로 해결되었다.[7][8][9] 또한 ''Agrobacterium'' 종은 독특한 공유파생형질, 즉 속의 모든 구성원이 선형 크로미드를 갖도록 하는 프로텔로메라제 유전자 ''telA''의 존재로 통합되었다.[10] 그러나 이 시점에서 세 개의 ''Agrobacterium'' 생물변종은 효력을 잃었다. 생물변종 1은 ''Agrobacterium''에 남았고, 생물변종 2는 ''Rhizobium rhizogenes''(라이조비움 라이조제네스)로, 생물변종 3은 ''Allorhizobium vitis''(알로라이조비움 비티스)로 이름이 변경되었다.3. 식물 병원성
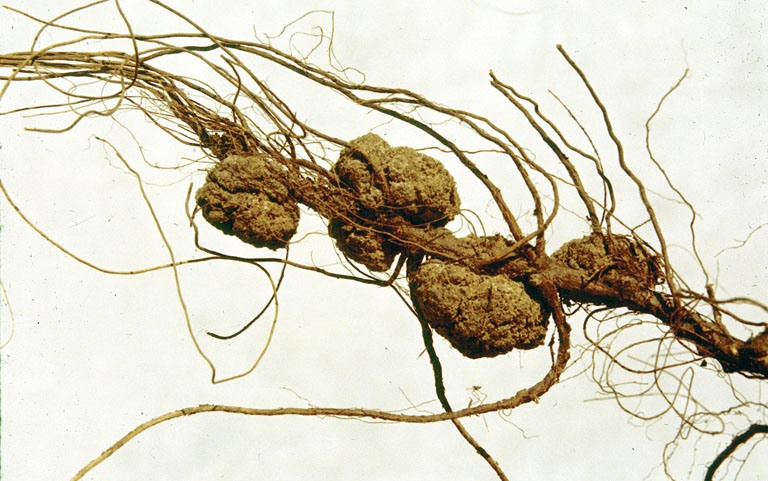
''아그로박테리움 투메파시엔스''(Agrobacterium tumefaciens)는 식물에 관모양 혹병(crown gall disease)을 일으킨다. 이 질병은 감염된 식물, 종종 뿌리와 줄기 사이의 접합부에서 종양과 같은 성장 또는 혹으로 특징지어진다. 종양은 세균성 종양 유도(Ti) 플라스미드에서 DNA 세그먼트(T-DNA)의 세균 접합 전달에 의해 유발된다.[11]
T-DNA는 식물 호르몬인 옥신과 시토키닌, 그리고 오파인 생합성을 위한 유전자를 가지고 있어, 선택적 이점을 제공한다.[13] 식물 세포 내 호르몬 균형 변화로 인해 세포 분열이 제어되지 않아 종양이 형성되며, 옥신 대 시토키닌 비율에 따라 종양 형태가 결정된다.[13] T-DNA는 숙주 세포의 게놈에 반무작위적으로 통합되며,[12] T-DNA의 종양 형태 유전자들이 발현되어 혹이 형성된다.
3. 1. 아그로박테리움의 종류와 플라스미드
''아그로박테리움 투메파시엔스''(Agrobacterium tumefaciens)는 식물에 관모양의 혹병을 일으킨다. 이 질병은 감염된 식물, 종종 뿌리와 줄기 사이의 접합부에서 종양과 같은 성장 또는 혹으로 특징지어진다. 종양은 세균성 종양 유도(Ti) 플라스미드에서 DNA 세그먼트(T-DNA)의 세균 접합 전달에 의해 유발된다.[11] 밀접하게 관련된 종인 ''아그로박테리움 라이조제네스''(Agrobacterium rhizogenes)는 뿌리 종양을 유발하며 별개의 Ri(뿌리 유도) 플라스미드를 가지고 있다.[11]
일반적으로 ''아그로박테리움 투메파시엔스'', ''아그로박테리움 라이조제네스'' 및 ''아그로박테리움 비티스''의 3가지 생물형이 존재한다. ''아그로박테리움 투메파시엔스''와 ''아그로박테리움 라이조제네스'' 내의 균주는 Ti 또는 Ri-플라스미드를 모두 가질 수 있으며, 일반적으로 포도나무에 국한된 ''아그로박테리움 비티스''의 균주는 Ti-플라스미드를 가질 수 있다.[11] Ri-플라스미드를 가지는 비 ''아그로박테리움'' 균주는 환경 샘플에서 분리되었으며, 실험 연구를 통해 비 ''아그로박테리움'' 균주 또한 Ti-플라스미드를 가질 수 있음이 밝혀졌다.[11] 일부 환경 ''아그로박테리움'' 균주는 Ti 또는 Ri-플라스미드를 모두 가지고 있지 않다. 이러한 균주는 비병원성이다.[11]
플라스미드 T-DNA는 숙주 세포의 게놈에 반무작위적으로 통합되며,[12] T-DNA의 종양 형태 유전자들이 발현되어 혹이 형성된다. T-DNA는 특이한 아미노산인 옥토핀 또는 노파린의 생성을 위한 생합성 효소에 대한 유전자를 가지고 있다. 또한 식물 호르몬인 옥신과 시토키닌의 생합성 및 오파인의 생합성을 위한 유전자를 가지고 있어, 대부분의 다른 미생물이 사용할 수 없는 탄소 및 질소원을 제공하여 ''아그로박테리움''에 선택적 이점을 제공한다.[13] 식물 세포 내 호르몬 균형을 변화시킴으로써, 해당 세포의 분열은 식물에 의해 제어될 수 없으며 종양이 형성된다. 종양 유전자에 의해 생성되는 옥신 대 시토키닌의 비율은 종양의 형태(뿌리 모양, 무질서 또는 줄기 모양)를 결정한다.[13]
4. 인간에 대한 영향
일반적으로 식물 감염으로 여겨지지만, ''아그로박테리움''은 면역 체계가 약화된 사람에게 기회 감염을 일으킬 수 있다.[14][15] 하지만 건강한 개인에게는 주요 병원체로 나타나지 않았다. ''아그로박테리움 라디오박터''에 의해 발생한 인간 질병의 가장 초창기 연관성은 1988년 스코틀랜드의 J. R. 케인 박사에 의해 보고되었다.[16] 이후 연구에서는 ''아그로박테리움''이 T-DNA를 인간 세포 유전자 속에 통합함으로써 여러 유형의 인간 세포에 부착되어 유전적으로 형질전환을 일으킨다는 것을 시사했다. 이 연구는 배양된 인간 조직을 사용하여 수행되었으며 자연 상태에서의 관련 생물학적 활동에 대한 어떠한 결론도 도출하지 않았다.[17]
5. 생명공학에서의 이용
아그로박테리움은 유전자를 식물과 균류에 전달하는 능력을 가지고 있어 생명공학, 특히 식물 개량을 위한 유전자 조작에 활용된다. 식물과 균류의 유전체는 T-DNA 이진 벡터를 이용하여 조작할 수 있다. 식물 형질전환에는 T-DNA의 두 개의 작은(25 염기쌍) 경계 반복 서열 중 적어도 하나가 필요하다.[27][28]
''아그로박테리움''을 사용한 균류의 형질전환은 주로 연구 목적으로 사용되며,[23][24] 식물 형질전환과 유사한 방식을 따른다. T-DNA 이진 벡터 시스템은 ''아그로박테리움'' 균주를 균류와 함께 배양한 후 형질전환된 균류 균주를 선택하는 방식으로 이루어진다.
5. 1. 형질전환 작물
''아그로박테리움''은 유전자를 식물과 균류에 전달하는 능력을 가지고 있어 생명공학, 특히 식물 개량을 위한 유전자 조작에 활용된다.[27][28] 이 과정에서는 종양 유발 유전자가 제거되어 '무장 해제'된 Ti 또는 Ri 플라스미드가 사용된다. 형질전환에 필요한 유전자는 식물 이진 벡터에 클론되며, 여기에는 T-DNA 영역과 형질전환 성공 여부를 판별할 수 있는 항생제 내성과 같은 선택 표지가 포함된다. 형질전환된 식물은 항생제가 포함된 배지에서 배양되며, T-DNA가 통합되지 않은 식물은 생존하지 못한다. 아그로침투는 또 다른 형질전환 방법이다.[18][19]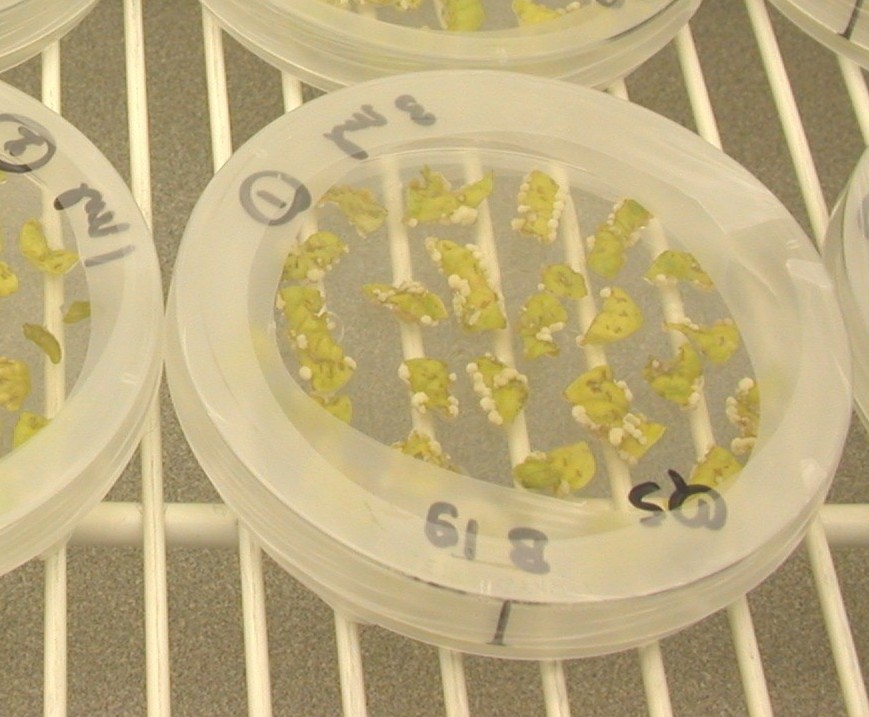
''아그로박테리움''을 이용한 형질전환은 원형질체나 잎 디스크를 ''아그로박테리움''과 함께 배양한 후 식물 조직 배양을 통해 완전한 식물을 재생하는 방식으로 이루어진다. 또는 아그로침투와 같이 ''아그로박테리움''을 식물 잎 조직에 직접 주입하는 방법을 사용할 수도 있다. 아그로침투는 박테리아와 직접 접촉하는 세포만 형질전환시키며, 플라스미드 DNA의 일시적인 발현을 유도한다.[20]
아그로침투는 주로 담배(''Nicotiana'') 형질전환에 사용되며, 애기장대의 경우 꽃 침지법이 일반적인 형질전환 프로토콜이다.[21] 이 방법은 꽃차례를 ''아그로박테리움'' 현탁액에 담가 생식세포를 만드는 생식 계열 세포를 형질전환시킨다. 이후 씨앗에서 항생제 내성 등 원하는 표지를 가진 개체를 선별한다. 플라스미드 DNA가 통합되지 않은 식물은 항생제에 의해 제거된다.[18]
''아그로박테리움''은 미국의 아래 GMO 작물에 유전 물질을 전달하는 벡터로 사용되었다:[22]
6. 유전체학
''아그로박테리움'' 게놈은 원형 염색체, 선형 염색체/크로미드, (일부 종에서) Ti 플라스미드의 세 부분으로 구성된다.[25]
여러 종의 ''아그로박테리움'' 게놈의 염기서열 분석을 통해 이들 유기체의 진화 역사를 연구할 수 있게 되었으며, 병원성, 생물학적 방제 및 공생에 관여하는 유전자와 시스템에 대한 정보를 제공했다. 중요한 발견 중 하나는 이러한 많은 박테리아에서 플라스미드로부터 염색체가 진화하고 있을 가능성이다. 또 다른 발견은 이 그룹의 다양한 염색체 구조가 공생과 병원성 생활 방식을 모두 지원할 수 있는 것으로 보인다는 것이다. ''아그로박테리움'' 종의 게놈 염기서열 정보의 활용은 계속 증가하여, 이 식물 관련 미생물 그룹의 기능과 진화 역사에 대한 상당한 통찰력을 얻을 수 있을 것이다.[26]
7. 역사
마르크 반 몬테규와 요제프 셸은 벨기에 겐트 대학교에서 ''아그로박테리움''과 식물 사이의 유전자 전달 메커니즘을 발견했다. 이는 ''아그로박테리움''을 식물 유전자 공학의 효율적인 전달 시스템으로 변경하는 방법 개발로 이어졌다.[27][28] 1983년, 메리-델 칠턴이 이끄는 연구팀은 병원성 유전자를 제거해도 ''아그로박테리움''이 자체 DNA를 식물 게놈에 삽입하는 능력에 부정적인 영향을 미치지 않는다는 것을 처음으로 입증했다.[29]
참조
[1]
논문
Proposal for rejection of the generic name ''Polymonas'' Lieske 1928
[2]
논문
Phylogenetic position of the marine subdivision of ''Agrobacterium'' species based on 16S rRNA sequence analysis
1997-08
[3]
논문
Reclassification of marine ''Agrobacterium'' species: Proposals of ''Stappia stellulata'' gen. nov., comb. nov., ''Stappia aggregata'' sp. nov., nom. rev., ''Ruegeria atlantica'' gen. nov., comb. nov., ''Ruegeria gelatinovora'' comb. nov., ''Ruegeria algicola'' comb. nov., and ''Ahrensia kieliense'' gen. nov., sp. nov., nom. rev.
1998-06
[4]
논문
A revision of ''Rhizobium'' Frank 1889, with an emended description of the genus, and the inclusion of all species of ''Agrobacterium'' Conn 1942 and ''Allorhizobium undicola'' de Lajudie ''et al''. 1998 as new combinations: ''Rhizobium radiobacter'', ''R. rhizogenes'', ''R. rubi'', ''R. undicola'' and ''R. vitis''
2001-01
[5]
논문
''Agrobacterium'' is a definable genus of the family Rhizobiaceae
2003-09
[6]
논문
Classification and nomenclature of ''Agrobacterium'' and ''Rhizobium''
2003-09
[7]
논문
History and current taxonomic status of genus ''Agrobacterium''
[8]
논문
Phylogeny of the ''Rhizobium''-''Allorhizobium''-''Agrobacterium'' clade supports the delineation of ''Neorhizobium'' gen. nov.
[9]
논문
Revised phylogeny of ''Rhizobiaceae'': Proposal of the delineation of ''Pararhizobium'' gen. nov., and 13 new species combinations
[10]
논문
Single acquisition of protelomerase gave rise to speciation of a large and diverse clade within the ''Agrobacterium''/''Rhizobium'' supercluster characterized by the presence of a linear chromid
[11]
논문
Proposal for rejection of ''Agrobacterium tumefaciens'' and revised descriptions for the genus ''Agrobacterium'' and for ''Agrobacterium radiobacter'' and ''Agrobacterium rhizogenes''
1993-10
[12]
논문
Identification of ''Arabidopsis thaliana'' transformants without selection reveals a high occurrence of silenced T-DNA integrations
2005-02
[13]
논문
New insights into an old story: ''Agrobacterium''-induced tumour formation in plants by plant transformation
2010-03
[14]
논문
''Agrobacterium'' infections in humans: experience at one hospital and review
1993-01
[15]
논문
Recovery of a strain of ''Agrobacterium radiobacter'' with a mucoid phenotype from an immunocompromised child with bacteremia
1993-09
[16]
논문
A case of septicaemia caused by ''Agrobacterium radiobacter''
1988-03
[17]
논문
Genetic transformation of HeLa cells by ''Agrobacterium''
2001-02
[18]
논문
Genetic Engineering of Plants
http://www.eolss.net[...]
2016-07-17
[19]
논문
Efficient agroinfiltration of plants for high-level transient expression of recombinant proteins
2013-07
[20]
논문
Optimization and utilization of ''Agrobacterium''-mediated transient protein production in ''Nicotiana''
2014-04
[21]
논문
Floral dip: a simplified method for ''Agrobacterium''-mediated transformation of ''Arabidopsis thaliana''
1998-12
[22]
웹사이트
The FDA List of Completed Consultations on Bioengineered Foods
http://www.cfsan.fda[...]
[23]
논문
Agrobacterium-mediated transformation as a tool for functional genomics in fungi
2005-07
[24]
논문
''Agrobacterium''-mediated transformation of fungi
2017
[25]
논문
Genomic analysis of Agrobacterium radiobacter DSM 30147T and emended description of A. radiobacter (Beijerinck and van Delden 1902) Conn 1942 (Approved Lists 1980) emend. Sawada et al. 1993
2014-01-02
[26]
서적
Plant Pathogenic Bacteria: Genomics and Molecular Biology
Caister Academic Press
[27]
서적
Genetic Engineering for Nitrogen Fixation
[28]
논문
Genetic analysis of transfer and stabilization of ''Agrobacterium'' DNA in plant cells
[29]
논문
Agrobacterium. A Memoir
https://www.jstor.or[...]
2001
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com