앤드류사쿠스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
앤드류사쿠스(Andrewsarchus mongoliensis)는 멸종된 포유류로, 1923년 내몽골의 이르딘 만하 지층에서 발견된 두개골 화석을 통해 처음 알려졌다. 처음에는 엔텔로돈의 두개골로 오인되었으나, 윌리엄 딜러 매튜에 의해 메소니키과에 속하는 크레오돈트로 분류되었고, 이후 오스본에 의해 앤드류사쿠스로 명명되었다. 분류학적으로는 메소니키드, 아르크토키온과, 그리고 고래 및 하마와 밀접한 관련이 있는 세탄코돈타모르파의 기저 구성원으로 여겨진다. 앤드류사쿠스는 커다란 두개골과 날카로운 이빨을 가지고 있었으며, 식성은 잡식성이거나 청소동물이었을 것으로 추정된다.
더 읽어볼만한 페이지
- 유제류 - 경하마형류
경하마형류는 분자생물학적 연구로 밝혀진 하마과와 고래하목을 포함하는 소목의 분류군으로, γ-피브리노겐 유전자 분석을 통해 공통 조상이 확인되었으며 반추동물, 라오엘리과와 자매 분류군으로 추정된다. - 염소 - 염소 (원소)
염소는 할로젠 원소로 황록색 기체이며 산화력과 살균 작용을 지니고 염화물 형태로 존재하며 전기분해를 통해 생산되어 살균, 표백, 화학 물질 생산 등에 사용되지만 독성이 강하고 오존층 파괴에도 영향을 미친다. - 염소 - 염소 소독
염소 소독은 물속 병원성 미생물을 제거하기 위해 염소를 사용하는 소독 방법으로, 수인성 질병 예방에 기여했으나 소독 부산물 생성 등의 문제점이 있어 안전하고 효과적인 사용을 위한 연구와 관리가 지속적으로 이루어지고 있다. - 에오세 아시아의 포유류 - 암불로케투스
암불로케투스는 유선형 몸과 특수한 귀 구조를 가진 초기 고래류 해양 포유류로, 악어와 유사한 방식으로 물 표면에서 매복하여 큰 포유류를 사냥했을 것으로 추정된다. - 에오세 아시아의 포유류 - 파키케투스
파키케투스는 5천만 년 전 에오세 초 파키스탄에 살았던 늑대 크기의 육상 포유류 고래류로, 육지 생활에 적응했으나 고래류 특유의 내이 구조와 치아 배열을 가져 고래 진화의 중요한 중간 단계이며, 고래가 육상 우제류에서 진화했음을 보여주는 중요한 증거이다.
| 앤드류사쿠스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Andrewsarchus |
| 명명자 | 헨리 페어필드 오즈본, 1924 |
| 모식종 | †Andrewsarchus mongoliensis |
| 모식종 명명자 | Osborn, 1924 |
| 속 하위 분류 | 다른 종 |
| 속 하위 분류 | †Andrewsarchus crassum Ding, Zheng, Zhang, & Tong, 1977 |
| 속 동의어 | Paratriisodon 저우밍전, 1959 |
| 종 동의어 | Paratriisodon henanensis Chow, 1959 Paratriisodon gigas Chow, Li, & Chang, 1973 |
| 분류 | |
| 상위 분류군 | 우제목목/경우제목목 |
| 시대 | |
| 화석 발견 시대 | 에오세 중기, |
| 발견 시대 | 약 4,500만 년 전 ~ 약 3,600만 년 전 (신생대고제3기에오세 중기-후기) |
| 특징 | |
| 예상 크기 | (두개골 길이) |
2. 발견 및 명명
앤드류사쿠스(''Andrewsarchus mongoliensis'')의 모식 표본은 거의 완전한 두개골(표본 번호 AMNH-VP 20135)이다.[6] 이 표본은 1923년 뉴욕 아메리카 자연사 박물관의 고생물학 탐사 중에 내몽골의 하부 이르딘 만하 지층에서 발견되었다.[2][7] 발견자는 현지 조수 간 추안바오[8]였으며, "벅샷"으로도 알려져 있다.[9] 이 표본은 처음 월터 W. 그레인저에 의해 엔텔로돈의 두개골로 확인되었다.[9] 두개골 그림이 박물관으로 보내졌고, 윌리엄 딜러 매튜에 의해 "메소니키과에 속하는 원시 크레오돈트"에 속하는 것으로 확인되었다.[2] 이후 1924년 오스본에 의해 기술되었다. 속명은 탐험대장 로이 채프먼 앤드루스의 이름을 기리고 있다.[2]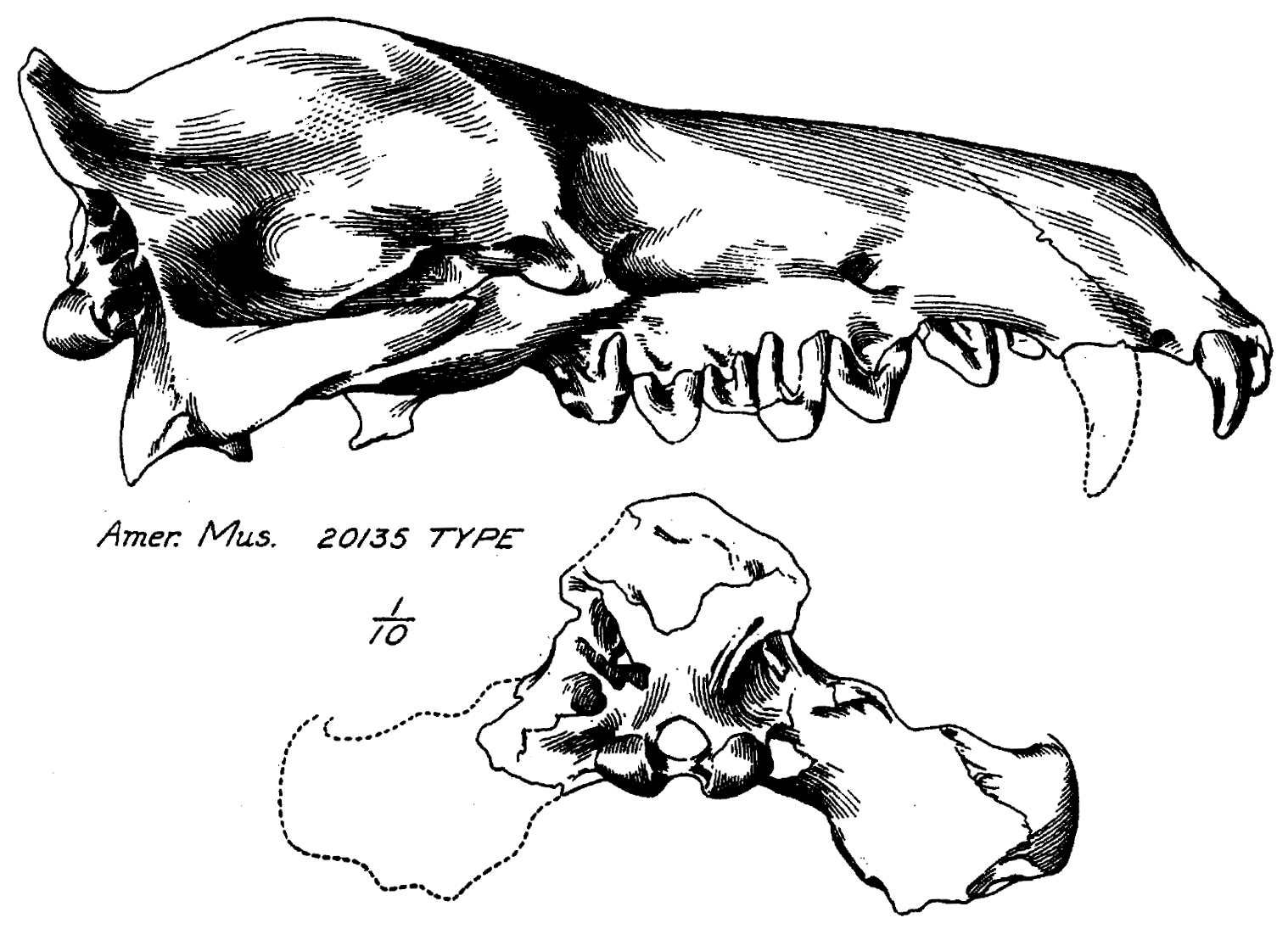
1977년 딩 수이인과 동료들은 광시 성 둥쥔 지층에서 발견된 한 쌍의 치아(두 번째와 세 번째 아래 앞니)인 IVPP V5101을 기반으로 두 번째 종인 ''A. crassum''을 명명했다.[10][11][12]
1957년 저우 밍전과 동료들은 중국 허난 성 루시 지층에서 이르딘 만하 지층과 관련된 하악골, 파편적인 상악골, 몇 개의 고립된 치아를 발견했다. 상악골은 으스러진 두개골에 속해 있었고, 하악골과 같은 개체에서 온 것일 가능성이 높다.[4] 저우는 1959년 이를 ''Paratriisodon henanensis''로 기술하고 아르크토키온과에 할당했다. 또한 어금니와 앞니가 ''트리이소돈''의 것과 유사하다는 점을 근거로 트리이소돈과의 아과인 트리이소돈아과 (현재 트리이소돈과)에 분류했다.[4] 1973년 저우와 동료들이 루시 지층에서 발견된 어금니를 기준으로 두 번째 종인 ''P. gigas''를 명명했다. 이후 이르딘 만하 지층에서 발견된 세 개의 어금니와 절치가 ''P. gigas''로 분류되었다.[13] 1969년 프레데릭 살라이는 두 속이 동일한 아르크토키온 조상으로부터 진화했거나 수렴 진화의 예시라고 제안했다.[14] 1978년 리 반 발렌에 의해 ''Paratriisodon''은 ''Andrewsarchus''와 동의어가 되었으며,[15] 1998년 모린 오리어리는 두 속의 어금니와 앞니의 유사점과 비슷한 몸 크기를 근거로 그들의 동의어를 유지했다.[12]
2. 1. 추가 발견
앤드류사쿠스(''Andrewsarchus mongoliensis'')의 모식 표본은 거의 완전한 두개골(표본 번호 AMNH-VP 20135)이다.[6] 이 표본은 1923년 뉴욕 아메리카 자연사 박물관의 고생물학 탐사 중에 내몽골의 하부 이르딘 만하 지층에서 발견되었다.[2][7] 발견자는 현지 조수 간 추안바오[8]였으며, "벅샷"으로도 알려져 있다.[9] 이 표본은 처음 월터 W. 그레인저에 의해 엔텔로돈의 두개골로 확인되었다.[9] 두개골 그림이 박물관으로 보내졌고, 윌리엄 딜러 매튜에 의해 "메소니키과에 속하는 원시 크레오돈트"에 속하는 것으로 확인되었다.[2] 이후 1924년 오스본에 의해 기술되었다. 속명은 탐험대장 로이 채프먼 앤드루스의 이름을 기리고 있다.[2]1977년 딩 수이인과 동료들은 광시 성 둥쥔 지층에서 발견된 한 쌍의 치아(두 번째와 세 번째 아래 앞니)인 IVPP V5101을 기반으로 두 번째 종인 ''A. crassum''을 명명했다.[10][11][12]
1957년 저우 밍전과 동료들은 중국 허난 성 루시 지층에서 이르딘 만하 지층과 관련된 하악골, 파편적인 상악골, 몇 개의 고립된 치아를 발견했다. 상악골은 으스러진 두개골에 속해 있었고, 하악골과 같은 개체에서 온 것일 가능성이 높다.[4] 저우는 1959년 이를 ''Paratriisodon henanensis''로 기술하고 아르크토키온과에 할당했다. 또한 어금니와 앞니가 ''트리이소돈''의 것과 유사하다는 점을 근거로 트리이소돈과의 아과인 트리이소돈아과 (현재 트리이소돈과)에 분류했다.[4] 1973년 저우와 동료들이 루시 지층에서 발견된 어금니를 기준으로 두 번째 종인 ''P. gigas''를 명명했다. 이후 이르딘 만하 지층에서 발견된 세 개의 어금니와 절치가 ''P. gigas''로 분류되었다.[13] 1969년 프레데릭 살라이는 두 속이 동일한 아르크토키온 조상으로부터 진화했거나 수렴 진화의 예시라고 제안했다.[14] 1978년 리 반 발렌에 의해 ''Paratriisodon''은 ''Andrewsarchus''와 동의어가 되었으며,[15] 1998년 모린 오리어리는 두 속의 어금니와 앞니의 유사점과 비슷한 몸 크기를 근거로 그들의 동의어를 유지했다.[12]
3. 분류학적 변천

''앤드류사쿠스''(''Andrewsarchus'')는 처음에 메소니키드(mesonychid)로 여겨졌고,[2] ''파라트리이소돈''(Paratriisodon)은 아르크토키온과(arctocyonid)로 여겨졌다.[13] 1995년에 전자는 아과(subfamily) 앤드류사르키나에(Andrewsarchinae)의 유일한 구성원이 되었으며, 이는 메소니키아(Mesonychia) 내에 속한다.[16] 이 아과는 1998년 필립 D. 진저리치(Philip D. Gingerich)에 의해 과(family) 수준으로 격상되었으며, 그는 잠정적으로 ''파라트리이소돈''을 이에 배정했다.[17] 1988년, 도널드 프로테로(Donald Prothero)와 동료들은 ''앤드류사쿠스''를 고래의 자매군(sister taxon)으로 회복시켰다.[18]
화석은 1923년 6월, 중국 고비 사막 내몽골의 이르딘 만하(Irdin Mahna)에서 앞서 언급한 조사대의 일원이었던 간 추엔 파오(Kan Chuen Pao)에 의해 발견되었다.[36] 거기서 발견된 것은 하악골이 없는 두개골과 불과 2, 3개의 골편뿐이었으며, 이어진 발견 사례는 거의 없다. 그러나 치아와 두개골의 형태학적 분석을 통해 중치류와의 유연성이 지적되어 당시에는 하위 분류에 추가되었다.[37]
본 종은 이듬해인 1924년, 고생물학자 헨리 페어필드 오스본에 의해 기재(학술상의 정식 명명)되었다. 중치류는 한때 치아의 유사성 등으로 고래목의 조상 계통으로 생각하는 것이 주류였다. 그러나 분자계통학에 의한 지견, 그리고 다른 화석의 발견 등에 기초한 형태학적 지견에 의해 (비교적 가깝다고는 할 수 있지만) 직접적인 조상 계통이나 가장 근연이라고 할 수 있는 계통은 아니라는 것이 판명되었다.[38]
그러나 1966년 시점에서 두개골 형태와 치아의 엔테로돈과와의 유사성으로, 중치류를 바탕으로 복원한 오스본의 복원상은 잘못되었다는 지적이 있었다.[39]
이 지적을 뒷받침하듯이 2009년의 계통분석에서는 이 속은 중치류와는 거리가 멀고, 오히려 엔테로돈과나 고래하마형류에 근연하다는 결과가 나왔다.[40]
그 이후, 이는 세탄코돈타모르파(Cetancodontamorpha)의 더 기저(basal)적인 구성원으로 회복되었으며, 엔텔로돈트(entelodont), 하마(hippos), 그리고 고래(whales)와 가장 밀접한 관련이 있다.[19][20]
2023년, 유(Yu)와 동료들은 엔텔로돈트과(entelodontid) 우제목(ungulates)에 특히 초점을 맞춰 우제목의 계통발생학적 분석을 수행했다. ''앤드류사쿠스''는 자기 자신, ''아케노돈(Achaenodon)'', ''얼리안히우스(Erlianhyus)'', ''프로엔텔로돈(Proentelodon)'', ''우투히우스(Wutuhyus)'', 그리고 엔텔로돈트과(Entelodontidae)로 구성된 분지군의 일부로 회복되었다. 이는 ''아케노돈''과 ''얼리안히우스''와 가장 밀접한 관련이 있으며, 이들과 함께 폴리토미(polytomy)를 형성하는 것으로 밝혀졌다.[19]
아래는 그들의 계통발생학에 기초한 분기도(cladogram)이다.[19]
{| class="wikitable"
|-
! 우제목(Artiodactyla)
|-
|
{| class="wikitable"
|-
| 낙타상목(Camelidamorpha)
|-
|
{| class="wikitable"
|-
| 돼지상목(Suinamorpha)
|-
|
{| class="wikitable"
|-
| 반추상목(Ruminantiamorpha)
|-
! 세탄코돈타모르파(Cetancodontamorpha)
|-
|
{| class="wikitable"
|-
| 샴모테리움 크라비엔시스(Siamotherium krabiensis)
|-
|
{| class="wikitable"
|-
|
{| class="wikitable"
|-
|
| 얼리안히우스 프리미티부스(Erlianhyus primitivus) |
| 앤드류사쿠스 몽골리엔시스(Andrewsarchus mongoliensis) |
| 아케노돈 유인텐시스(Achaenodon uintensis) |
|-
|
{| class="wikitable"
|-
| 우투히우스 프리미티부스(Wutuhyus primitivus)
|-
|
| 프로엔텔로돈(Proentelodon) |
| 엔텔로돈트과(Entelodontidae) |
|}
|}
|-
|
| 하마상목(Hippopotamidamorpha) |
| 고래상목(Cetaceomorpha) |
|}
|}
|}
|}
|}
2022년의 견해에서도 앤드류사쿠스가 엔테로돈류와 중치목 중 어느 쪽에 속하는지는 의견이 갈리고 있으며, 분류의 혼란은 당분간 계속될 가능성이 있다.[36]
덧붙여 2011년에 트리이소돈과 (triisodontids)와 중치과를 대상으로 한 연구에서는 안드류사르쿠스가 비교 재료 중 하나로 언급되었다.[42]
3. 1. 2009년 계통 분석
''앤드류사쿠스''(''Andrewsarchus'')는 처음에는 메소니키드(mesonychid)로 여겨졌으며,[2] 1995년에 아과(subfamily) 앤드류사르키나에(Andrewsarchinae)의 유일한 구성원이 되었고, 이는 메소니키아(Mesonychia) 내에 속한다.[16] 2009년의 계통분석에서는 이 속이 중치류와는 거리가 멀고, 오히려 엔테로돈과나 고래하마형류에 근연하다는 결과가 나왔다.[40]
이후, 세탄코돈타모르파(Cetancodontamorpha)의 더 기저(basal)적인 구성원으로 회복되었으며, 엔텔로돈트(entelodont), 하마(hippos), 그리고 고래(whales)와 가장 밀접한 관련이 있다.[19][20] 2023년, 유(Yu)와 동료들은 엔텔로돈트과(entelodontid) 우제목(ungulates)에 특히 초점을 맞춰 우제목의 계통발생학적 분석을 수행했다. ''앤드류사쿠스''는 자기 자신, ''아케노돈(Achaenodon)'', ''얼리안히우스(Erlianhyus)'', ''프로엔텔로돈(Proentelodon)'', ''우투히우스(Wutuhyus)'', 그리고 엔텔로돈트과(Entelodontidae)로 구성된 분지군의 일부로 회복되었다. 이는 ''아케노돈''과 ''얼리안히우스''와 가장 밀접한 관련이 있으며, 이들과 함께 폴리토미(polytomy)를 형성하는 것으로 밝혀졌다.[19]
2022년의 견해에서도 안드류사르쿠스가 엔테로돈류와 중치목 중 어느 쪽에 속하는지는 의견이 갈리고 있으며, 분류의 혼란은 당분간 계속될 가능성이 있다.[36]
3. 2. 2023년 계통 분석
2023년, 유(Yu)와 동료들은 엔텔로돈트과(entelodontid) 우제목(ungulates)에 초점을 맞춰 우제목의 계통발생학적 분석을 수행했다.[19] ''앤드류사쿠스''는 아케노돈(Achaenodon), 얼리안히우스(Erlianhyus), 프로엔텔로돈(Protentelodon), 우투히우스(Wutuhyus), 그리고 엔텔로돈트과(Entelodontidae)로 구성된 분지군의 일부로 회복되었으며, ''아케노돈''과 ''얼리안히우스''와 가장 밀접한 관련이 있으며, 이들과 함께 폴리토미(polytomy)를 형성하는 것으로 밝혀졌다.[19]아래는 그들의 계통발생학에 기초한 분기도(cladogram)이다.[19]
{| class="wikitable"
|-
! 우제목(Artiodactyla)
|-
|
{| class="wikitable"
|-
| 낙타상목(Camelidamorpha)
|-
|
{| class="wikitable"
|-
| 돼지상목(Suinamorpha)
|-
|
{| class="wikitable"
|-
| 반추상목(Ruminantiamorpha)
|-
! 세탄코돈타모르파(Cetancodontamorpha)
|-
|
{| class="wikitable"
|-
| 샴모테리움 크라비엔시스(Siamotherium krabiensis)
|-
|
{| class="wikitable"
|-
|
{| class="wikitable"
|-
|
| 얼리안히우스 프리미티부스(Erlianhyus primitivus) |
| 앤드류사쿠스 몽골리엔시스(Andrewsarchus mongoliensis) |
| 아케노돈 유인텐시스(Achaenodon uintensis) |
|-
|
{| class="wikitable"
|-
| 우투히우스 프리미티부스(Wutuhyus primitivus)
|-
|
| 프로엔텔로돈(Proentelodon) |
| 엔텔로돈트과(Entelodontidae) |
|}
|}
|-
|
| 하마상목(Hippopotamidamorpha) |
| 고래상목(Cetaceomorpha) |
|}
|}
|}
|}
|}
2022년의 견해에서도 앤드류사쿠스가 엔테로돈류와 중치목 중 어느 쪽에 속하는지는 의견이 갈리고 있으며, 분류의 혼란은 당분간 계속될 가능성이 있다.[36]
4. 형태
앤드류사쿠스는 커다란 길쭉한 코와, 날카로운 이빨 그리고 납작한 볼을 가진 네발 짐승으로, 길쭉한 코와 날카로운 이빨은 다른 짐승의 뼈를 부수기 위해서라고 생각된다. 발톱은 없고 발굽이 있으며, 속도가 느린 편이라 시체를 주로 먹었을 것으로 추정된다. 앤드류사쿠스의 화석은 주로 몽골과 중국 북부 만리장성 이북 지역에서 발견되었으며, 첫 발견자인 채프먼 앤드류의 이름을 따서 앤드류사쿠스라 이름 붙였다.
앤드류사쿠스는 아직 몇 개의 해골밖에 알려지지 않아서, 이 동물이 포식자였는지, 혹은 이 동물이 하이에나 같은 청소부 역할을 했을지에 대해서 논쟁이 붙고 있다. 두개골의 길이는 1m이고, 몸길이는 3m 가량으로 추정된다. 앤드류사쿠스는 육식성이었으나, 그와 가까운 친척이던 아메리카 대륙의 오레오돈트는 초식동물이었다. 현재의 염소와 양에 가까운 근연종으로, 염소와 양의 직계 조상인 오레오돈트와 공통 조상에서 갈려나간 종이었다.
오스본은 처음 ''앤드루사쿠스''를 묘사할 때, 가장 큰 육상 육식성 포유류라고 믿었다. ''A. mongoliensis'' 모식표본 두개골의 길이를 기반으로, ''메소닉스''의 비율을 사용하여, 전체 몸길이는 3.82m, 몸 높이는 1.89m로 추정했다.[2] 그러나, 두개골과 치아의 유사성을 엔텔로돈트와 비교하여, 프레데릭 설레이와 스티븐 제이 굴드는 이 동물이 메소닉스과보다는 엔텔로돈트와 더 유사한 비율을 가졌을 것이라고 제안했으며, 따라서 오스본의 추정은 부정확할 가능성이 높다고 보았다.[1]
''안드류사쿠스''의 모식 표본 두개골의 전체 길이는 83.4cm이며, 관골궁에서 너비는 56cm이다. 주둥이는 매우 길쭉하며, 두개 기저 길이의 1.5배에 달하고,[2] 송곳니 앞쪽의 주둥이 부분은 엔텔로돈트과와 유사하다.[1] 그러나 엔텔로돈트과와 달리, 안와후연은 불완전하다.[19] 시상 능선은 축소되었고, 하악와는 비교적 평평하다. 이러한 특징들은 약한 측두근과 상당히 약한 물어뜯는 힘을 시사한다. 경구개는 길고 좁다.[1] 하악와는 또한 두개 기저에서 측면과 복면으로 오프셋되어 있는데, 이는 중치류에서 보이는 상태와 유사하다.[21] 하악골 자체는 길고 얕으며, 곧고 비교적 얕은 수평지가 특징이다.[4] 두 하악골에 교근이 부착되는 하악골의 함몰 부위인 교근와는 얕다. 두 하악골 사이의 결합 접촉은 제한적이다.[1]
''안드류사쿠스''의 모식 표본 두개골은 측면당 3개의 앞니, 1개의 송곳니, 4개의 소구치 및 3개의 어금니로 구성된 전형적인 치아 배열을 보여준다.[1] 하지만 하악골에도 동일하게 적용되는지는 불분명하다. 위쪽 앞니는 송곳니 앞에서 반원 형태로 배열되어 있으며, 이는 엔텔로돈트와 공유하는 특징이다. 두 번째 앞니는 커져서 송곳니 크기와 거의 비슷하다.[1] 이는 원래 송곳니가 "엄청난 크기"로 묘사되었지만,[2] 치아 전체에 비례하여 상대적으로 작기 때문이다.[1] 위쪽 소구치는 길쭉하며 단일 융기로 구성되어 있으며, 엔텔로돈트의 소구치와 유사하다. 네 번째 소구치는 원뿔을 유지하지만, 흔적 기관 형태이다.[2] 뿌리는 합류되지 않고 상아질 플랫폼이 없는데, 이는 모두 치아의 마모 후 치아의 기능적 수명을 연장하기 위한 적응으로 보인다. 첫 번째 어금니가 가장 작다. 두 번째는 가장 넓지만 화석화 이후 심하게 마모되었다. 세 번째는 그 마모를 크게 피했다. 소구치와 어금니는 치관이 주름져 있는데, 이는 멧돼지과 및 기타 잡식성 우제목에서 보이는 상태와 유사하다.[1] 하악골(IVPP V5101)의 치아 구조는 거의 모두 치아 마모되었거나 부러져서 파악하기 어렵다. 첫 번째 소구치를 제외한 오른쪽 하악골의 모든 치아가 보존되어 있으며, 첫 번째 소구치는 대신 왼쪽 하악골에 보존되어 있다. 아래쪽 송곳니와 첫 번째 소구치는 모두 앞쪽을 향하고 있다.[1] 세 번째 어금니는 크며, 두 개의 융기를 가진 탈론을 가지고 있다.[4]
안드류사르쿠스는 현재 알려진 모든 육식성 육상 포유류 중에서 가장 큰 턱을 가진 동물이다. 긴 주둥이와 잘 발달된 턱을 가지고 있으며, 그 턱에 달린 이빨은 모두 컸다. 앞니, 굽고 날카로운 송곳니, 그리고 튼튼한 어금니를 갖추고 있다.
4. 1. 크기
앤드류사쿠스는 커다란 길쭉한 코와, 날카로운 이빨 그리고 납작한 볼을 가진 네발 짐승으로, 길쭉한 코와 날카로운 이빨은 다른 짐승의 뼈를 부수기 위해서라고 생각된다.[43] 발톱은 없고 발굽이 있으며, 속도가 느린 편이라 시체를 주로 먹었을 것으로 추정된다. 앤드류사쿠스의 화석은 주로 몽골과 중국 북부 만리장성 이북 지역에서 발견되었으며, 첫 발견자인 채프먼 앤드류의 이름을 따서 앤드류사쿠스라 이름 붙였다.두개골의 길이는 1m이고, 몸길이는 3m 가량으로 추정된다. 두개골은 길이가 83.4cm[43], 최대 폭 56cm로 거대하다. 두개골 형태가 유사한 메소니쿠스 오브투시덴스를 참고하여 두개골 길이로부터 단순 계산된 앤드류사쿠스의 크기는, 몸길이 약 382cm, 어깨 높이 약 189cm 정도로 추정되었다. 그러나 몸통 화석은 미발견 상태이며, 실제로는 메소니쿠스와의 유연성이 지지받지 못하게 됨에 따라 실제 전장은 아직 불명이다. 체형의 비율은 엔테로돈류에 가까울 것이라는 추측은 1966년에 이루어졌지만, 구체적인 값은 제시되지 않았다[39]。
5. 식성
앤드류사쿠스는 커다란 길쭉한 코와 날카로운 이빨, 그리고 납작한 볼을 가진 네발 짐승이었다. 길쭉한 코와 날카로운 이빨은 다른 짐승의 뼈를 부수기 위한 것으로 추정된다. 발톱은 없고 발굽이 있었으며, 속도가 느려 시체를 주로 먹었을 것으로 보인다. 앤드류사쿠스의 화석은 주로 몽골과 중국 북부 만리장성 이북 지역에서 발견되었다.
앤드류사쿠스는 육식성이었으나, 그와 가까운 친척이던 아메리카 대륙의 오레오돈트는 초식동물이었다. 현재의 염소와 양에 가까운 근연종으로, 염소와 양의 직계 조상인 오레오돈트와 공통 조상에서 갈려나간 종이었다. 앤드류사쿠스는 날카로운 송곳니와 넓고 튼튼한 어금니를 갖고 있었다. 주로 육식성이었으나 작은 동물을 직접 사냥하기도 했고, 죽은 동물의 사체, 즙이 많은 나뭇잎과 나무줄기, 산딸기 종류, 곤충의 유충과 땅을 기어다니는 벌레류, 작은 설치류와 도마뱀 등을 씹어먹었다.
얼굴 앞부분에 난 윗송곳니는 턱 밖으로 나와 있으며, 안쪽으로 구부러져 있고, 어금니는 고기를 잘 씹도록 칼날과 같은 형태를 띠고 있었다. 위턱의 어금니는 상당히 넓었는데, 뼈를 부술 수 있을 수준이었다. 기타 이빨은 곰의 이빨과 비슷하다.
오스본은 ''안드류사쿠스''를 잡식성일 가능성이 있다고 제안했다.[2] 몰라와 앞니의 심하게 주름진 치관을 증거로 사용한 설레이와 굴드는 이러한 결론을 뒷받침했다.[1] 1990년 R.M. 조켈은 이를 "잡식성-청소동물"일 가능성이 높으며, 앤텔로돈트와 생태학적으로 유사하다고 제안했다.[22] 라르스 베르델린은 이를 청소동물이라고 제안했으며, 브론토테르류를 잡아먹었을 가능성도 제시했다.[23]
6. 고생태학
앤드류사쿠스는 커다란 길쭉한 코, 날카로운 이빨, 납작한 볼을 가진 네 발 짐승이었다. 길쭉한 코와 날카로운 이빨은 다른 짐승의 뼈를 부수는 데 사용되었을 것으로 추정된다. 발굽이 있어 속도가 느렸을 것으로 보이며, 시체를 주로 먹었을 것으로 추정된다. 앤드류사쿠스의 화석은 주로 몽골과 중국 북부 만리장성 이북 지역에서 발견되었다.
앤드류사쿠스는 몇 개의 해골만 알려져 있어 포식자였는지, 하이에나 같은 청소부 역할을 했는지에 대한 논쟁이 있다. 두개골 길이는 1m, 몸길이는 3m 가량으로 추정된다. 앤드류사쿠스는 육식성이었으나, 가까운 친척인 아메리카 대륙의 오레오돈트는 초식동물이었다. 염소와 양의 직계 조상인 오레오돈트와 공통 조상에서 갈라져 나간 종으로, 현재의 염소와 양에 가까운 근연종이었다.
에오세 대부분 동안에는 습하고 열대적인 환경과 지속적으로 높은 강수량을 보이는 온실 기후가 우세했다. 기제목, 우제목, 그리고 영장류 (또는 부속과 유영장류)를 포함한 현대 포유류의 목은 이미 초기 에오세에 나타나 급속히 다양화되었고, 잎을 먹는 데 특화된 치아를 발달시켰다. 잡식성 형태들은 대부분 잎을 먹는 식단으로 바꾸거나, 구식 "유제류"와 함께 중기 에오세 (루테티아 시대–바토니아 시대, 4800만 년 전~3800만 년 전)에 멸종되었다.[24][25]
''안드루사쿠스''의 모식표본이 발견된 이르딘 만하 지층은 중기 에오세에 해당하는 이르딘만한 지층으로 구성되어 있다.[26] ''안드루사쿠스 몽골리엔시스''는 하부 이르딘만한에 해당하는 IM-1 지역에서 발견되었으며,[27] 여기에서는 하이에노돈족 ''프로프테로돈'', 중치류 ''하르파골레스테스'', 적어도 세 종의 이름 없는 중치류,[1] 우제목 ''얼리안히우스'',[28] 기제목 ''데페레텔라''와 ''로피알레테스'', 오모미과 ''타르콥스'', 글라이레스 ''곰포스'', 설치류 ''탐콰미스''와 다양한 불확정적 글라이레스가 알려져 있다.[26][27] ''파라트리이소돈 헤난엔시스'' 표본이 발견된 루시 지층은 이르딘 만하 지층과 거의 같은 시기에 퇴적되었다. 중치류 ''메소닉스'', 판토돈트 ''유디노케라스'', 디코부니드 ''디코부네'', 헬로히과 ''고비오히우스'', 브론토테르 ''리노티탄'' 및 ''마이크로티탄'', 기제목 ''아미노돈'' 및 ''로피알레테스'', 테노닥틸리드 ''친링고미스''와 토끼목 ''루실라구스''가 루시 지층에서 확인되었다.[29] ''A. 크라섬''이 기원하는 동준 지층 역시 중기 에오세이다.[30] 이곳에서는 님라브과 ''에우스밀루스'', 안트라코테리과 ''프로브라키오두스'', 판토돈트 유디노케라스, 브론토테르 ''메타텔마테리움'' 및 cf. ''프로티탄'', 데페레텔리드 ''데페레텔라''와 ''텔레올로푸스'', 히라코돈과 ''포르스터쿠페리아'', 코뿔소과 ''일리안오돈'' 및 ''프로히라코돈'', 아미노돈과 ''아미노돈'', ''기가타미노돈''과 ''파라미노돈''이 보존되어 있다.[31]
참조
[1]
논문
Asiatic Mesonychidae (Mammalia, Condylartha)
http://hdl.handle.ne[...]
1966
[2]
논문
''Andrewsarchus'', giant mesonychid of Mongolia
http://hdl.handle.ne[...]
1924
[3]
논문
The age and characteristic of the Liuniu and the Dongjun faunas, Bose Basin of Guangxi
http://www.ivpp.ac.c[...]
1977
[4]
논문
A new arctocyonid from the Upper Eocene of Lushih, Honan
http://www.ivpp.cas.[...]
2022-03-20
[5]
논문
Late Eocene mammalian faunas of Honan and Shansi with notes on some vertebrate fossils collected therefrom
http://www.ivpp.ac.c[...]
2023-03-27
[6]
논문
Impact of increased character sampling on the phylogeny of Cetartiodactyla (Mammalia): combined analysis including fossils
https://onlinelibrar[...]
2008-08-07
[7]
웹사이트
PBDB Collection
https://paleobiodb.o[...]
2024-08-18
[8]
웹사이트
American Museum of Natural History Research Library: Chuen-pao, Kan (Buckshot) (amnhp_1002386)
https://data.library[...]
2024-10-11
[9]
서적
The New Conquest of Central Asia: A Narrative of the Explorations of the Central Asiatic Expeditions in Mongolia and China, 1921–1930
New York, American Museum of Natural History.
[10]
웹사이트
PBDB Taxon
https://paleobiodb.o[...]
2024-10-10
[11]
논문
The age and characteristics of the vertebrate fauna from Liuniu and Oongjun Formations of the Bose Basin, Zhuang autonomous region
1977
[12]
서적
The Emergence of Whales
Springer
1998
[13]
논문
Irdin Manha Upper Eocene and its mammalian fauna at Huhebolhe Cliff in central Inner Mongolia
http://www.ivpp.cas.[...]
1980
[14]
서적
The Hapalodectinae and a phylogeny of the Mesonychidae (Mammalia, Condylarthra)
https://www.biodiver[...]
American Museum of Natural History
2024-10-11
[15]
논문
The Beginning of the Age of Mammals
https://www.mn.uio.n[...]
2024-08-16
[16]
간행물
Evolution of Paleocene-Eocene Mesonychidae (Mammalia, Mesonychia)
https://hdl.handle.n[...]
University of Michigan
1995
[17]
간행물
Paleobiological Perspectives on Mesonychia, Archaeoceti, and the Origin of Whales
https://link.springe[...]
Springer US
2024-08-16
[18]
서적
The Phylogeny and Classification of the Tetrapods
1988
[19]
논문
A new entelodont (Artiodactyla, Mammalia) from the late Eocene of China and its phylogenetic implications
2023
[20]
논문
Relationships of Cetacea (Artiodactyla) among mammals: Increased taxon sampling alters interpretations of key fossils and character evolution
2009
[21]
서적
The affinities of Apterodon (Mammalia, Deltatheridia, Hyaenodontidae)
https://www.biodiver[...]
American Museum of Natural History
2024-10-11
[22]
논문
A functional interpretation of the masticatory system and paleoecology of entelodonts
https://www.cambridg[...]
2024-10-11
[23]
간행물
Carnivore Behavior, Ecology, and Evolution
https://www.degruyte[...]
Cornell University Press
2024-10-11
[24]
논문
Mountain uplift explains differences in Palaeogene patterns of mammalian evolution and extinction between North America and Europe
[25]
논문
Western European middle Eocene to early Oligocene Chiroptera: systematics, phylogeny and palaeoecology based on new material from the Quercy (France)
[26]
웹사이트
PBDB Collection
https://paleobiodb.o[...]
2024-08-18
[27]
논문
Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China
https://link.springe[...]
2010-12-01
[28]
논문
A new middle Eocene bunodont artiodactyl from the Erlian Basin (Nei Mongol, China)
https://www.tandfonl[...]
2024-10-11
[29]
웹사이트
Mammalian faunal lists of East Asia formatted for AEO analysis (Alroy, 1992, 1994, 1996, 1998, 2000).
https://doc.rero.ch/[...]
null
[30]
웹사이트
PBDB Collection
https://paleobiodb.o[...]
2024-10-13
[31]
논문
A preliminary report on the Eocene mammals of the Pondaung fauna, Myanmar
https://repository.k[...]
2000
[32]
논문
The age and characteristic of the Liuniu and the Dongjun faunas, Bose Basin of Guangxi
http://www.ivpp.ac.c[...]
[33]
서적
絶滅哺乳類図鑑
https://books.google[...]
丸善
2002-03-20
[34]
웹사이트
小学館の図鑑NEO 大むかしの生物
https://www.shogakuk[...]
2023-07-19
[35]
문서
[36]
서적
特別展「化石ハンター展 ~ゴビ砂漠の恐竜とヒマラヤの超大型獣~」公式図録
国立科学博物館
2022-07-16
[37]
학술지
Andrewsarchus, giant mesonychid of Mongolia
https://cir.nii.ac.j[...]
[38]
웹사이트
"「デメテール」群馬県立自然史博物館だより No.72 2頁"
http://www.gmnh.pref[...]
群馬県立自然史博物館群馬県立
2023-07-19
[39]
학술지
Asiatic Mesonychidae (Mammalia, Condylarthra). Bulletin of the AMNH ; v. 132, article 2
https://hdl.handle.n[...]
[40]
학술지
Relationships of Cetacea (Artiodactyla) Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution
https://www.ncbi.nlm[...]
2009-09-23
[41]
학술지
A new entelodont (Artiodactyla, Mammalia) from the late Eocene of China and its phylogenetic implications
https://www.tandfonl[...]
2023-01-01
[42]
학술지
A structural intermediate between triisodontids and mesonychians (Mammalia, Acreodi) from the earliest Eocene of Portugal
https://doi.org/10.1[...]
2011-02-01
[43]
문서
現生種では[[ライオン]]が大きくても約43cm程度。つまり、本種の頭蓋骨はその特徴的な細長い形で尺を稼ぐとは言え、ライオンのほぼ倍の大きさである。
[44]
서적
よみがえる恐竜・古生物: 超ビジュアルCG版
https://ci.nii.ac.jp[...]
ソフトバンク クリエイティブ株式会社
2006-07-15
[45]
웹사이트
BBC - Science & Nature - Making of Walking with Beasts
https://www.bbc.co.u[...]
2023-07-19
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com