양산 세포
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
양산 세포는 눈의 망막에 위치하며, 시각 시스템에서 물체의 위치와 움직임을 감지하는 데 중요한 역할을 하는 망막 신경절 세포의 한 유형이다. 파라솔 신경절 세포라고도 불리며, 크고 빠르게 신호를 전달하는 특징을 가진다. 양산 세포는 거대 세포 경로를 통해 정보를 전달하며, 무채색 정보, 높은 시간 주파수, 낮은 공간 주파수에 민감하다. 이러한 특성으로 양산 세포는 시각적 탐색, 움직임 감지, 깊이 지각에 기여하며, 난독증과 같은 질환과 관련되기도 한다.
더 읽어볼만한 페이지
- 인간 세포 - 무축삭 세포
무축삭 세포는 망막 내부에서 양극 세포와 망막 신경절 세포 간의 시냅스 연결을 조절하며, 연결 영역, 내부 얼기층 위치, 신경 전달 물질 종류에 따라 분류되고, 다양한 기능을 수행한다. - 인간 세포 - B세포
B세포는 골수에서 성숙하여 항체를 생산하고 체액성 면역에 관여하는 림프구의 일종으로, 항원에 반응하여 형질 세포로 분화하거나 면역 기억을 형성하는 기억 B 세포로 남으며 기능 이상 시 자가면역질환이나 림프종을 유발할 수 있다. - 시각계 - 눈 (해부학)
눈은 빛을 감지하여 시각 정보를 뇌로 전달하는 감각 기관이며, 다양한 종류가 존재하고, 안구, 시신경, 부속기로 구성되며, 진화 과정을 통해 다양한 환경에 적응해왔다. - 시각계 - 시각 영역
시각 영역은 눈으로 볼 수 있는 시야의 범위로, 정상 범위는 수평 180~200도, 수직 130~140도이며, 시야 결손은 다양한 질환이나 뇌 손상으로 발생하고 시야 검사로 진단한다.
| 양산 세포 | |
|---|---|
| 일반 정보 | |
 | |
| 일부 | 눈의 망막 |
| 계통 | 시각 시스템 |
| 추가 정보 | |
| 전구체 | 알 수 없음 |
| 동맥 | 알 수 없음 |
| 정맥 | 알 수 없음 |
| 신경 | 알 수 없음 |
| 림프 | 알 수 없음 |
2. 구조
파라솔 신경절 세포는 눈의 망막에 위치하며, 전체 망막 신경절 세포의 약 10%를 차지한다.[3] 이 세포는 큰 세포체와 광범위하게 퍼져 서로 겹치는 가지 모양의 수상돌기를 가진다.[3][8] 또한 두껍고 수초화가 잘 된 축삭을 가지고 있어, 빠른 신호 전달에 유리한 구조적 특징을 보인다.[4][7] 중심와를 포함한 망막 전반에 걸쳐 비교적 고른 밀도로 분포하는 특징도 지닌다.[8]
2. 1. 파라솔 세포 vs. 미idget 세포
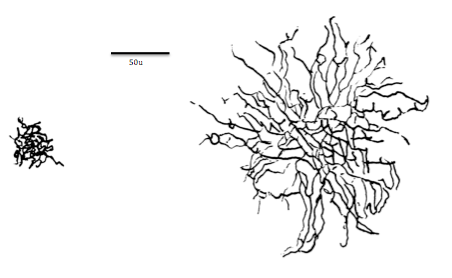
파라솔 신경절 세포는 눈의 망막에 위치하며, 전체 망막 신경절 세포의 약 10%를 차지한다.[3] 이 세포는 넓게 퍼져 서로 겹치는 가지 모양의 수상돌기를 가진 큰 세포체와[3][8] 두껍고 수초화가 잘 된 축삭을 가지고 있다. 이러한 구조적 특징 덕분에 파라솔 세포는 P 경로에 정보를 전달하는 왜소 세포보다 훨씬 빠르게 신호를 보낼 수 있다.[4][7]
파라솔 신경절 세포는 막대 세포와 원추 세포 모두로부터 정보를 받는 넓은 수용 영역을 가진다.[3][7] 하지만 원추 세포로부터 입력을 받음에도 불구하고 색상에 대한 정보는 처리하지 않는다.[3][7] 왜소 세포와 달리, 파라솔 세포의 수용 영역 중심과 주변부 모두 동일한 색상 유형의 원추 세포를 포함한다. 이러한 색상 특이성의 부족으로 인해 파라솔 세포는 물체에서 반사되는 서로 다른 빛의 파장을 구별할 수 없으며, 명암과 같은 무채색 정보만을 전달한다.[10]
망막의 중심부인 중심와에서도 파라솔 신경절 세포는 주변부와 거의 동일한 밀도로 존재하는데, 이는 중심와에 밀집하는 왜소 세포와 구별되는 또 다른 특징이다.[8]
파라솔 세포와 왜소 세포는 각각 병렬적으로 거대 세포 경로와 소세포 경로를 시작하며 시각 정보 처리에 기여한다. 두 세포 모두 시각 시스템에서 중요한 역할을 하지만, 아래 표와 같이 해부학적 구조와 기능적 측면에서 뚜렷한 차이를 보인다.[3][11][15][12]
| RGC 유형 | 파라솔 세포 | 왜소 세포 |
|---|---|---|
| 관여하는 경로 | 거대 세포 경로 | 소세포 경로 |
| 세포체 크기 | 큼 | 작음 |
| 수상돌기 나무 | 복잡함 | 덜 복잡함 |
| 전도 속도 | ~1.6 ms | ~2 ms |
| 시각 시스템에서의 기능 | 물체가 "어디"에 있는지; 물체를 "어떻게" 잡는지 | 미세한 세부 사항에 따라 물체가 "무엇"인지 |
| 공간 주파수에 대한 민감도 | 낮음 | 중간에서 높음 |
| 시간 주파수 | 높음 | 낮음 |
| 색상 반대성 | 무채색 | 적-녹 반대성 |
양산 망막 신경절 세포는 세밀한 정보나 색상 정보는 제공하지 못하지만,[4] 유용한 정적, 깊이, 움직임 정보를 제공한다. 이 세포는 높은 명암 대비 감지 능력을 가지며,[13] 높은 공간 주파수보다 낮은 공간 주파수에서 더 민감하다. 이러한 대비 정보로 인해 이 세포는 휘도 변화를 감지하는 데 뛰어나며, 따라서 시각적 탐색 작업 수행과 가장자리 감지에 유용한 정보를 제공한다.[14]
일반적으로 뉴런은 세포 외부에서 금속 전극을 이용해 연구하지만, 망막 신경절 세포는 특히 생체 외( in vitro|인 비트로la ) 환경에서 연구된다. 이러한 방법을 통해 양산 세포의 복잡하게 얽힌 구조를 세포 내부에서 분석할 수 있다. 1941년 폴리악은 골지 염색법을 이용하여 망막 신경절 세포를 처음으로 확인했으며, 수상돌기의 형태를 면밀히 분석하여 큰 수상돌기 나무 구조를 밝혀냈다. 이후 1986년 카플란과 셰플리는 양산 세포를 시각 시스템과 처음으로 연결시킨 연구 결과를 발표했다. 이들은 외측 슬상핵(LGN)에 있는 망막 신경절 세포(RGC)의 축삭 말단에서 S-전위(S-potential)를 기록하여, 영장류의 magnocellular layer|거대 세포층영어으로 이어지는 세포들이 높은 명암 감도를 보이고, parvocellular layer|소세포층영어으로 이어지는 세포들이 낮은 명암 감도를 보인다는 사실을 밝혔다.[3]
거대세포 경로의 비정상적인 신호 전달은 난독증과 조현병과 관련이 있는 것으로 알려져 있다.[21][22]
[1]
서적
The central nervous system : structure and function
https://archive.org/[...]
Oxford University Press
2010
3. 기능
양산 망막 신경절 세포는 또한 물체의 위치에 대한 정보를 제공하는 데 중요하다. 이 세포는 공간에서 물체의 방향과 위치를 감지할 수 있으며,[5][15] 이 정보는 결국 배측 경로를 통해 전송된다.[16] 이 정보는 양안 깊이 지각에 중요한 도구인 각 눈의 망막에서 물체의 위치 차이를 감지하는 데에도 유용하다.[5][17]
양산 세포는 높은 시간 주파수를 감지하는 능력이 있으며,[18] 따라서 물체의 위치에서 빠른 변화를 감지할 수 있다.[7] 이것이 움직임 감지의 기초이다.[5][13][19] 두정엽 내구 (IPS)로 전송된 정보는 거대 세포 경로가 주의를 기울이고 시야에서 중요한 움직이는 물체를 따라 빠른 안구 운동을 안내할 수 있게 한다.[4][14][19] 눈으로 물체를 따라가는 것 외에도, IPS는 손과 팔이 크기, 위치 및 위치에 따라 물체를 올바르게 잡을 수 있도록 움직임을 조정할 수 있게 하는 전두엽의 일부에 정보를 보낸다.[16] 이러한 능력으로 인해 일부 신경 과학자들은 거대 세포 경로의 목적이 공간적 위치를 감지하는 것이 아니라 물체의 위치 및 움직임과 관련된 행동을 안내하는 것이라고 가설을 세웠다.[20]
4. 마그노세포 경로

양산 세포는 시각 시스템에서 마그노세포 경로의 첫 단계를 담당한다. 이 세포들은 망막에서 시작하여 시신경을 통해 시상의 특정 핵인 LGN의 가장 아래쪽 두 개 층으로 정보를 전달한다. 이 LGN 층들은 주로 마그노세포로 채워져 있으며, 이 마그노세포들은 다시 선조 피질 (V1)의 4Cα 층으로 정보를 보낸다.[7]
궁극적으로 양산 세포가 망막에서 수집한 정보는 시각 피질의 여러 부분으로 전달된다. 주요 경로는 두 가지가 있다. 하나는 후두정 피질과 V5 영역을 거치는 배쪽 경로이고, 다른 하나는 하측 측두회와 V4 영역을 거치는 복쪽 경로이다.[6]
5. 연구 및 실험
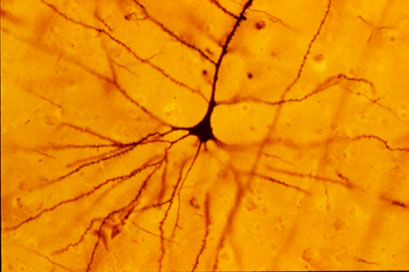
영장류와 기타 모델 시스템인간 시각 연구의 모델 시스템으로 사용되는 구세계원숭이와 신세계원숭이는 양산 세포 연구에도 중요한 기여를 했다.[8] 예를 들어, 역행성 표지( retrograde labeling|역행성 표지영어 ) 기법을 이용한 여러 원숭이 연구는 양산 세포가 거대 세포 경로(magnocellular pathway)와, 먼지 세포(midget cell)가 소세포 경로(parvocellular pathway)와 각각 연결된다는 점을 밝혀냈다. 또한 이러한 연구들은 color opponency theory|색상 반대 이론영어의 기초를 마련하는 데 기여했다.[3][8] 1996년 데이시는 생체 외 영장류 망막 세포에 염료를 주입하는 실험을 통해 이러한 결과를 뒷받침했다. 그의 연구에서 거대 세포 경로와 연결된 양산 세포는 색상 정보를 처리하지 않는 무채색(achromatic) 특징을 보이는 것으로 나타났다.[3] 다른 연구에서는 마모셋과 같은 신세계원숭이를 이용하여 외측 슬상핵(LGN) 거대 세포층의 공간 및 시간 주파수 특성에 대한 이해를 높이는 데 기여했다. 니슬 염색법( Nissl staining|니슬 염색법영어 )을 이용한 연구 결과, 거대 세포층의 세포체는 소세포층이나 코니오세포층( koniocellular layer|코니오세포층영어 )의 세포체보다 더 어둡고 밀도가 높은 것으로 확인되었다.[11]
고양이의 망막 신경절 세포 역시 영장류 및 인간의 시각 시스템과 비교 연구되었다. 고양이 연구를 통해, 양산 세포는 그 구조적 특징으로 인해 먼지 세포보다 더 큰 receptive field|수용 영역영어을 가진다는 사실이 밝혀졌다. 이는 인간의 망막 세포에서도 유사할 것으로 추정되며, 더 넓은 수용 영역은 spatial localization|공간적 위치 파악영어에 유리할 수 있다.[3]
6. 관련 질환
특히, 양산 신경절 세포의 발달이 부족한 것이 난독증을 유발하는 데 영향을 미칠 수 있다는 이론이 있다. 양산 신경절 세포는 시각 시스템에 운동 정보를 제공하여 뇌가 눈의 도약 운동을 조절하도록 돕는다. 만약 도약 운동에 문제가 생기면 시야가 흐려지고 글을 읽는 데 어려움을 겪을 수 있다. 이러한 발달 부전은 영양 결핍이나 6번 염색체에 있는 KIAA0319 유전자의 변이 등 다양한 요인에 의해 발생할 수 있다. 또한, 항신경항체에 의한 자가 면역 공격이 양산 신경절 세포의 정상적인 발달을 방해할 수도 있는데, 이는 면역 체계가 약한 사람들에게서 난독증이 자주 나타나는 이유를 설명할 수 있다.[4]
참조
[2]
서적
Neuroscience
https://archive.org/[...]
Sinauer
2004
[3]
논문
Structure and function of parallel pathways in the primate early visual system
2005-07
[4]
논문
Dyslexia: the Role of Vision and Visual Attention
2014-01-01
[5]
논문
Early visual development: differential functioning of parvocellular and magnocellular pathways
1992-01-01
[6]
논문
Two functional channels from primary visual cortex to dorsal visual cortical areas
2001-04-13
[7]
논문
Parallel processing strategies of the primate visual system
2009-05
[8]
논문
Receptive field structure in the primate retina
1996-03
[9]
논문
Projection of rods and cones within human visual cortex
2000-01-01
[10]
논문
The circuitry of V1 and V2: integration of color, form, and motion
2005-01-01
[11]
논문
Tracking blue cone signals in the primate brain
2013-05
[12]
논문
On the prevalence of magnocellular deficits in the visual system of non-dyslexic individuals
2004-01
[13]
논문
Review: steady and pulsed pedestals, the how and why of post-receptoral pathway separation
2011-07-07
[14]
논문
The role of the magnocellular pathway in serial deployment of visual attention
2004-10-01
[15]
논문
On identifying magnocellular and parvocellular responses on the basis of contrast-response functions
2011-01-01
[16]
논문
What visual information is processed in the human dorsal stream?
2012-06-13
[17]
논문
The analysis of stereopsis
1984
[18]
논문
Efficiently Measuring Magnocellular and Parvocellular Function in Human Clinical Studies
2015-09-01
[19]
논문
Neural underpinnings of dyslexia as a disorder of visuo-spatial attention
2004-01-01
[20]
논문
An evolving view of duplex vision: separate but interacting cortical pathways for perception and action
2004
[21]
논문
Dyslexia: the Role of Vision and Visual Attention
2014-01-01
[22]
논문
Face recognition in schizophrenia disorder: A comprehensive review of behavioral, neuroimaging and neurophysiological studies
2015-06-01
[23]
서적
The central nervous system : structure and function
https://archive.org/[...]
Oxford University Press
2010
[24]
서적
Neuroscience
https://archive.org/[...]
Sinauer
2004
[25]
논문
Structure and function of parallel pathways in the primate early visual system
https://archive.org/[...]
2005-07
[26]
논문
Dyslexia: the Role of Vision and Visual Attention
2014-01-01
[27]
논문
Early visual development: differential functioning of parvocellular and magnocellular pathways
1992-01-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com