웃는얼굴거미
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
웃는얼굴거미(Theridion grallator)는 몸길이가 5mm 미만인 하와이 제도 고유종 거미이다. 이 거미는 길고 가느다란 다리와 투명한 노란색 몸체를 가지며, 복부에는 다양한 색상과 무늬가 나타난다. 특히 복부에 웃는 얼굴과 유사한 패턴을 가진 개체는 '웃는얼굴'이라는 별칭으로 불린다. 웃는얼굴거미는 하와이 열대 우림에서 발견되며, 먹이에 따라 몸의 색깔이 변하며, 잎 아래에 작은 거미줄을 쳐서 먹이의 진동을 감지한다. 암컷은 알을 낳고 부화할 때까지 보호하며, 새끼를 돌보는 모성애를 보인다.
더 읽어볼만한 페이지
- 1900년 기재된 동물 - 예쁜꼬마선충
예쁜꼬마선충은 생물학 연구에 사용되는 작은 선충으로, 짧은 수명과 투명한 몸체, 유전자 조작의 용이성으로 인해 발생학, 유전학 등 다양한 분야에서 모델 생물로 활용된다.
| 웃는얼굴거미 | |
|---|---|
| 분류 정보 | |
| 학명 | Theridion grallator Simon, 1900 |
| 과 | 꼬마거미과 |
| 추가 정보 | |
| 특이 사항 | "웃는 얼굴" 모양의 무늬가 있음 |
2. 형태
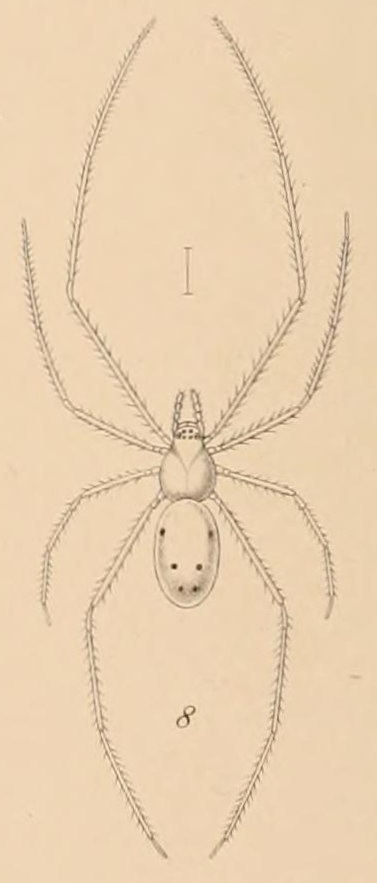
웃는얼굴거미(''T. grallator'')는 몸길이가 5mm 미만인 작은 거미이다.[4] 길고 가느다란 다리와 투명한 노란색 몸체를 가지고 있는 것이 특징이다.[2] 이 거미의 뚜렷하게 긴 다리는 분기도의 모든 구성원 중 가장 다른 신체 형태를 가지게 한다.[2] 복부는 옅은 투명한 노란색을 띠며, 다양한 빨간색, 흰색 및/또는 검은색 패턴을 포함할 수 있다.[3] 일부 변이형은 노란색 몸체에 웃는 얼굴 또는 웃는 광대 얼굴과 유사한 패턴을 가지고 있어 이러한 속칭이 생겼다. 섬마다 패턴이 다르며, 일부는 복부에 표시가 전혀 없다.[4] 복부 색상은 식단에 따라 투명한 노란색에서 녹색 또는 주황색으로 변한다.[5]
2. 1. 색상 변이
웃는얼굴거미의 복부에는 노란색, 빨간색, 검은색, 흰색 등 다양한 색상과 무늬가 나타나는데, 이는 섬마다, 개체마다 다르게 나타난다.[3][4][5] 이러한 다양성은 포식자의 눈을 피하기 위한 적응 전략으로 여겨진다.[5]웃는얼굴거미의 색상 다형성은 단순한 멘델 유전학을 따른다.[7] 가장 흔한 형태는 노란색으로, 개체군의 70%를 차지한다.[7] 노란색 형태는 모든 무늬 있는 형태에 대해 열성이다.[7] 무늬 있는 형태 내에서 복부에 존재하는 색소의 양은 관련된 대립 유전자의 우성과 상관관계가 있으며, 검은색, 빨간색 또는 흰색 색소와 관련된 대립 유전자는 계층적 구조로 배열되어 우성 효과를 나타낸다.[7] 흰색은 거의 모든 형태에 우성이다.[7]
흰색 형태는 외피에서 파생된 구조인 표피(hypodermis) 아래에 다량의 구아닌이 침착되어 생성된다.[8] 구아닌은 거미의 주요 질소질 배설물로, 이러한 침착물은 흰색 배경을 생성한다.[9] 구아닌 침착물과 신체 내 분포는 ''T. grallator''의 주요 유전자 좌위의 제어를 받는다.[9]
복부 색상 형질의 유전적 기반은 섬마다 다르다.[8] 마우이에서는 하나의 유전자좌에서 기원한 반면, 하와이에서는 최소 두 개의 연관되지 않은 유전자좌가 있다.[8] 또한, 마우이에서는 모든 다형성이 개별 대립유전자에 기인하는 반면, 하와이에서는 수컷과 암컷에서 차등적으로 발현되는 단일 유전자좌에 의존할 수 있는 두 쌍의 색상 형질이 있다.[8]
마우이와 하와이의 ''T. grallator'' 색상 형질의 서로 다른 유전적 배경은 두 섬의 연령과 그들의 정착의 차이 때문이다.[8] 하와이에서 일부 성 선택적 형질이 존재한다는 사실은 진화적 압력으로 인해 유전 패턴의 변화가 발생했음을 시사한다.[14]
웃는얼굴거미의 색상 다형성의 진화적 중요성은 아직 불분명하지만, 다양한 형태 비율에 작용하는 선택 압력이 존재한다.[7]
2. 2. 유전적 요인
웃는얼굴거미의 색깔과 무늬는 멘델 유전학을 따른다.[7] 가장 흔한 형태는 노란색인데, 전체의 70%를 차지한다.[7] 노란색 형태는 '무늬 없는' 형태로도 알려져 있으며, 다른 무늬가 있는 형태에 비해 열성이다.[7] 무늬가 있는 형태에서 색소의 양은 관련 대립 유전자의 우성과 관련이 있다.[7] 검은색, 빨간색, 흰색 색소와 관련된 대립 유전자는 우성 효과를 보이며 계층적으로 배열된다.[7] 흰색은 거의 모든 형태에 대해 우성이다.[7]흰색 형태는 외피에서 유래된 표피(hypodermis) 아래에 구아닌이 많이 쌓여서 나타난다.[8] 구아닌은 거미의 주요 질소질 배설물이다.[9] 이러한 구아닌 침착은 밝은 색상 형태에 유리하게 작용한다.[9] 구아닌 침착물과 신체 내 분포는 ''T. grallator''의 주요 유전자 좌위의 제어를 받는다. 이 주요 유전자 좌위는 구아닌의 유무에 반응하고, 표피와 소화 게실 사이에서 화학적 신호를 보내 형태 색소 침착을 조절하는 두 가지 메커니즘의 제어를 받는다. 또한, 이 두 가지 메커니즘은 독립적으로 또는 함께 작동할 수 있다. 첫 번째 메커니즘은 색소 침착에 대한 구아닌의 효과를 억제하여 작동한다. 따라서 무색소 부위는 그 아래에 구아닌 층을 포함하게 된다. 두 번째 메커니즘은 빛을 이용하여 구아닌을 유도하여 무색소 부위 아래에 구아닌 침착물이 존재하게 하여 작동한다. 구아닌은 다양한 형태의 패턴을 형성하는 빨간색과 검은색 표피 색소 아래에서만 발견된다.[9] 흰색과 빨간색 선은 공동 우성을 보인다.[8] 성별 간 형태 분포에는 성 연관이 없는 것으로 보인다.[2]
''Theridion grallator''는 풍부한 갑각과 후체부 (복부) 패턴을 가진다. 후체부 형태는 상염색체 좌위에 있는 대립 유전자에 의해 결정되는 것으로 보인다. 우성은 보통 하나의 패턴을 다른 패턴 위에 겹쳐서 나타난다. 좌위 간의 연결은 갑각과 후체부 패턴 사이의 연관성을 만들 수 있다. 특정 좌위에 있는 대립 유전자의 다면발현 효과가 있을 수 있지만, 갑각과 복부에 관련된 패턴을 고려하면 가능성은 낮다. 구아닌 침착물과 함께 빨간색 및 검은색 후체부 색소 침착은 이러한 색상 다형성의 시각적 효과가 주는 이점을 보여준다.[10]
복부 색깔 형질의 유전적 기반은 섬마다 다르지만, 실제 복부 색깔 형질은 섬 전체에서 동일하게 보인다. 마우이섬에서는 ''T. grallator''의 색깔 형질이 하나의 유전자좌에서 나온 반면, 하와이섬에서는 색상 다형성에 관여하는 최소 두 개의 연관되지 않은 유전자좌가 있다. 마우이섬에서는 모든 다형성이 개별 대립유전자에 기인하지만, 하와이섬에서는 수컷과 암컷에서 다르게 발현되는 단일 유전자좌에 의존하는 두 쌍의 색깔 형질이 있다.[8] 이러한 차등 발현 형질의 한 쌍은 노란색과 붉은색 앞면인데, 암컷은 노란색, 수컷은 붉은색으로 나타난다. 하와이 개체군의 붉은색 덩어리와 붉은색 고리는 성별에 따라 암컷은 붉은색 덩어리, 수컷은 붉은색 고리로 다르게 나타난다.[7] 이러한 표현형의 차이는 성 연관이 아닌 차등적 발현 때문일 가능성이 높다.[13]
마우이섬과 하와이섬의 ''T. grallator'' 색깔 형질의 유전적 배경이 다른 이유는 두 섬의 나이와 정착의 차이 때문이다. 마우이섬이 먼저 나타났고, 그 다음이 하와이섬이다.[8] 하와이섬에서 일부 성 선택적 형질이 존재하는 것은 진화적 압력으로 인해 유전 패턴의 변화가 발생했음을 보여준다.[14] 현재 하와이 제도 섬들 간의 개체 교환은 거의 없으며, 각 섬에서 단일 계통 분지도를 형성한다.[14] 유전적 배경의 차이와 개체의 드문 교환에도 불구하고, 섬 간의 잡종 교배는 여전히 생존 가능한 자손을 낳을 수 있다. 이는 마우이섬과 하와이섬의 ''T. grallator''가 서로 크게 분화되지 않았음을 의미한다.[14]
3. 분포 및 서식지
웃는얼굴거미는 하와이 제도 고유종으로, 오아후, 몰로카이, 마우이, 하와이섬의 열대 우림에서 발견된다. 해발 300m에서 2000m 사이의 고도에서 서식한다.[2]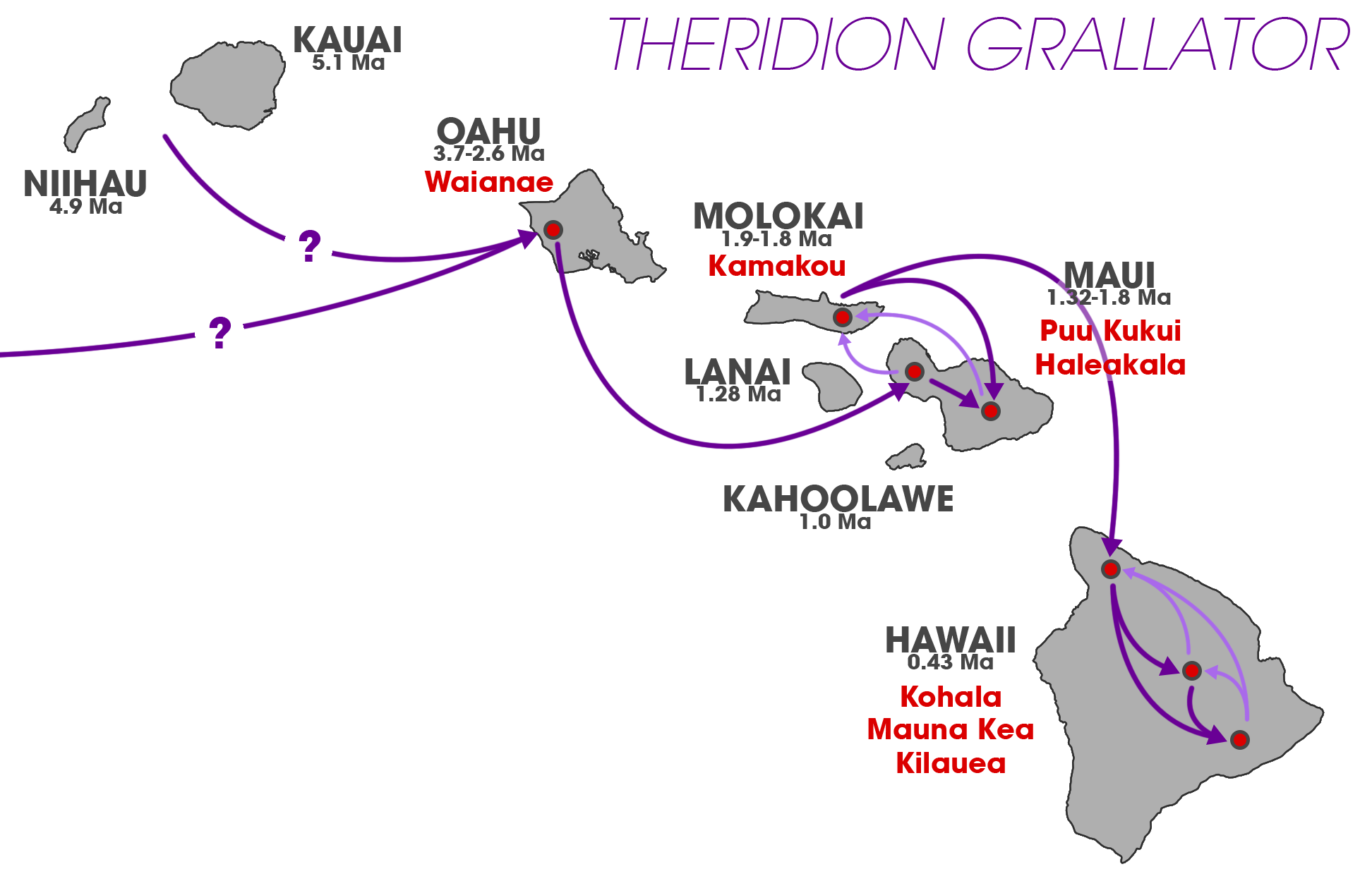
3. 1. 서식 환경
웃는얼굴거미(''T. grallator'')는 습하거나 중간 정도의 습한 환경에서 서식한다.[2] 습한 환경은 연간 강수량이 200cm에서 350cm 사이인 곳을 말하며, 중간 정도의 습한 환경은 연간 강수량이 100cm에서 200cm 사이인 곳을 말한다. 이 거미는 하와이 제도의 숲에서 발견된다.[2] 오아후, 몰로카이, 마우이, 하와이섬에서 발견되었다.[1] 웃는얼굴거미는 토착 식물인 ''Broussaisia arguta''와 ''Clermontia arborescens'', 그리고 외래종인 ''Hedychium coronarium''과 같은 식물의 잎 아랫면에서 주로 서식한다. 특히 ''H. coronarium''은 크고 미끄러운 잎을 가지고 있어 웃는얼굴거미가 포식자를 피하는 데 도움을 준다.[2]웃는얼굴거미는 용암류로 둘러싸인 키푸카 지역에서도 발견되었지만,[16] 그 지역을 둘러싼 용암류 자체에서는 발견되지 않는다.
4. 생태
웃는얼굴거미(''T. grallator'')는 먹이에 따라 복부 색깔이 변한다. 복부의 반투명성 때문에 먹이에 포함된 색소가 거미의 표피에 침착되어 색깔 변화를 일으킨다.[5] 반투명한 노란색에서 주황색으로 변하는 경우가 흔하며, 이는 주로 파리 (쌍시목)를 많이 먹기 때문으로 보인다.[9] 먹은 후 2~6일 동안 색소가 복부에 남아있다가 소화와 배설을 거치면 다시 원래의 옅은 노란색으로 돌아온다.[5]
웃는얼굴거미는 거미줄을 쳐서 먹이를 잡는 대신, 자유롭게 먹이를 찾아다니며 실을 이용해 먹이를 잡는다.[17] 파리매과와 초파리과가 주된 먹이이다.[17] 서식하는 잎의 종류에 따라 먹이에 대한 공격성이 달라지기도 한다.[17]
4. 1. 포식자
자나방속의 육식성 애벌레는 웃는얼굴거미를 공격하는 것이 관찰되었다.[18] 하와이 제도에는 웃는얼굴거미를 잡아먹는 여러 종의 ''Eupithecia''가 있다. 이 애벌레는 잎 위에 숨어 있다가 몸의 끝부분과 접촉하는 거미를 공격할 수 있다. 공격을 받으면 웃는얼굴거미는 애벌레를 물고 도망가려고 한다.엘레우테로닥틸루스 코키는 푸에르토리코에서 온 침입종으로, 웃는얼굴거미를 잡아먹는다. 1980년대에 하와이에서 발견되었다.[19]
4. 2. 거미줄
웃는얼굴거미(''T. grallator'')는 식물의 잎 아래에 비교적 작은 2차원 거미줄을 친다.[20] 거미줄은 보통 잎 아랫면이나 나무 틈새에서 발견된다. 웃는얼굴거미의 거미줄은 매우 얇고 엉성한데, 이는 전형적인 가슴거미과의 특징이다. 웃는얼굴거미는 대부분의 가슴거미류가 짓는 거미줄보다 훨씬 더 얇은 작은 거미줄을 짓는다. 거미줄이 많이 사용되지 않는 이유는 하와이 기후의 진화적 압력으로 인해 거미줄이 불리하게 작용했기 때문일 수 있다. 높은 강수량은 거미줄 실의 접착제를 손상시켜 먹이 포획 효율을 떨어뜨린다. 웃는얼굴거미는 거미줄을 먹이 감지 수단으로 사용하는 대신, 먹이가 잎을 통해 전달하는 진동을 통해 먹이를 감지한다. 그런 다음 거미는 먹이의 위치와 방향을 구별할 수 있다.[17]작은 거미줄을 짓는 것은 종종 먹이 유형의 전문화와 관련이 있지만, 웃는얼굴거미의 경우에는 관찰되지 않는다. 낮 동안 웃는얼굴거미는 조류의 포식을 피하기 위해 잎 아랫면에 꽉 달라붙는다.[20] 밤에는 주행성 포식 조류가 잠들면, 이 거미들은 잎 아래에서 실을 늘어뜨려 매달린다. 웃는얼굴거미는 거미줄을 최소한으로만 사용하지만, 실을 사용하여 먹이를 잡을 수 있다. 웃는얼굴거미는 진동을 감지하여 먹이를 파악하고 관심 있는 먹이 근처로 몸을 돌린다. 그런 다음 거미는 빠르게 몸을 돌려 실을 먹이에게 던져 풀어놓는다. 이 실은 효율적인 먹이 포획을 가능하게 하는 끈적한 물질로 구성되어 있다.[17] 또한, 어미 웃는얼굴거미는 거미줄을 사용하여 알집을 보호하거나 새끼를 위해 잡은 먹이를 보관할 수 있다.[20]
5. 번식
암컷 웃는얼굴거미(''T. grallator'')는 교미 후 몇 주 뒤에 알집을 낳고, 알이 부화할 때까지 알집을 지킨다.[20]
웃는얼굴거미의 개체군은 계절에 따라 크기와 성별 구성이 변한다. 겨울철(10월~3월)에는 작고 미성숙한 거미가 많고, 봄철(5월~8월)에는 성체가 많아지며 대부분 어미 암컷이다. 봄철에는 개체군의 최대 85%가 알집을 가진 어미 암컷일 수 있다.[20]
성숙한 거미는 새끼 거미에게서 보이는 검은색 또는 적갈색 무늬가 없고, 배 전체를 덮는 붉은색 색소로 특징지어지는 붉은색 덩어리 형태가 더 많이 나타난다.[17]
5. 1. 암컷/수컷 상호작용
성숙한 수컷은 암컷을 찾아 숲의 식생을 적극적으로 이동하며, 암컷은 더 정착하는 경향이 있다. 구애는 주로 진동과 후각에 의존한다.[7] 예를 들어, 수컷은 신체적 움직임과 거미줄 뜯기를 포함하는 구애 춤을 수행할 수 있다. 구애 행동 동안의 이러한 진동은 잠재적인 암컷 배우자에 의해 평가된다. 교미는 밤에 이루어지며, 두 거미는 잎의 밑면에서 매달린다. 수컷은 교미 후 곧 죽지만 암컷은 더 오래 살며, 알이 부화할 때까지 알을 지키고 새끼를 위해 먹이를 잡는다.[20]''T. grallator''는 시력이 매우 낮은 거미과에 속하므로, 수컷은 진동 및 후각 신호를 사용하여 암컷에게 구애한다.[7]
5. 2. 부모의 돌봄
암컷 웃는얼굴거미(''T. grallator'')는 알집을 보호하고 새끼를 돌보며, 위탁된 알집이나 새끼도 받아들여 돌본다.[20] 어미는 포식, 기생 말벌, 서식하는 잎이 떨어질 가능성으로부터 새끼를 보호한다.[20] 거미 새끼가 부화하면 어미는 계속해서 새끼를 방어하고 돌보는데, 모든 새끼에게 함께 먹이를 주고 포식자로부터 보호하는 탁월한 모성애를 보여준다.[20] 거미 새끼는 약 40~100일 동안 어미와 같은 잎에 머물며, 이 시기 동안 스스로 먹이를 잡을 수 없어 어미가 없으면 죽는다.[20]어미 거미는 기꺼이 위탁된 알집을 받아들이며, 새끼 거미가 다른 무리로 옮겨지면 새로운 어미는 이 새끼 거미들을 가족으로 '입양'하여 마치 자신의 새끼처럼 돌본다.[20] 새끼 거미의 입양은 관련 어미가 죽었을 경우 발생할 수 있으며, 어미를 잃은 새끼 거미는 종종 다른 잎으로 이동하여 다른 무리에 합류한다.[20]
부모-자식 간 갈등은 어미 거미가 새끼를 보호하는 데 드는 비용에서 발생할 수 있다. 어미 암컷 ''T. grallator''가 두 번째 새끼를 낳는 경우, 첫 번째 새끼가 스스로 먹이를 먹을 수 없기 때문에 부화 후 일정 기간 동안 함께 머물러야 하므로, 첫 번째 새끼에 대한 부모의 투자가 필요하여 두 번째 새끼가 위협받을 수 있다.[20]
6. 사회 행동
암컷 웃는얼굴거미는 주로 움직이지 않고 잎 아랫면에 머무르는 반면, 수컷은 짝을 찾아 이동한다. 수컷은 이동성 때문에 포식자에게 더 눈에 띄기 쉽다.[7] 임신한 암컷과 알집을 지키는 암컷은 다른 성체 ''T. grallator''와 잎을 공유하지 않는다.[20]
같은 집단 내에서는 먹이 경쟁, 형제 살해, 동족 포식이 관찰되지 않았다.[20]
7. 기생
웃는얼굴거미(''T. grallator'')는 ''Baeus'' 속 말벌에 의해 기생당하는 비율이 높다.[21] 이 말벌은 ''Clubiona robusta''를 포함한 다른 거미에게도 기생한다고 알려져 있다.[20] 기생은 알의 사망률을 높이는 원인 중 하나이다. 말벌의 알이 작기 때문에 거미의 높은 기생률을 설명할 수 있다. 어미 거미는 자신의 알 주머니가 기생당했는지 알아차리기 어려울 수 있으며, ''Baeus''의 기생 행동은 어미가 알을 지키고 있을 때도 일어난다.[20]
참조
[1]
논문
Molecular insights into the phylogenetic structure of the spider genus ''Theridion'' (Araneae, Theridiidae) and the origin of the Hawaiian Theridion-like fauna
http://doi.wiley.com[...]
2007-07
[2]
논문
Maintaining a happy face: stable colour polymorphism in the spider ''Theiridion grallator'' (Araneae, Theridiidae)
1990-08
[3]
PhD
Comparative Behavioral Ecology of the Spider Theridion Grallator (Araneae: Theridiidae) in the Hawaiian Archipelago
University of California, Davis
[4]
논문
What makes a happy face? Determinants of colour pattern in the Hawaiian happy face spider ''Theridion grallator'' (Araneae, Theridiidae)
[5]
논문
Diet-induced color change in the Hawaiian happy-face spider ''Theridion grallator'', (Araneae, Theridiidae)
https://www.research[...]
1989
[6]
논문
Selection on the color polymorphism in Hawaiian happy-face spiders: evidence from genetic structure and temporal fluctuations
1998
[7]
논문
Portraits of evolution: studies of coloration in Hawaiian spiders
2001
[8]
논문
Quantum shifts in the genetic control of a colour polymorphism in ''Theridion grallator'' (Araneae: Theridiidae), the Hawaiian happy-face spider
1996-03
[9]
논문
Guanine as a colorant in spiders: development, genetics, phylogenetics and ecology
https://www.research[...]
1997
[10]
논문
Genetics of a colour polymorphism in Theridion grallator (Araneae: Theridiidae), the Hawaiian happy-face spider, from Greater Maui
1996-03
[11]
논문
The Evolution of Exuberant Visible Polymorphisms
https://www.jstor.or[...]
2009
[12]
논문
Stabilizing selection maintains exuberant colour polymorphism in the spider Theridion californicum (Araneae, Theridiidae): SELECTION MAINTAINS SPIDER COLOUR POLYMORPHISM
http://doi.wiley.com[...]
2011-01
[13]
논문
Mating in ''Theridion varians'' Hahn and related species (Araneae: Theridiidae)
http://www.tandfonli[...]
1998-04
[14]
논문
De novo characterization of the gene-rich transcriptomes of two color-polymorphic spiders, Theridion grallator and T. californicum (Araneae: Theridiidae), with special reference to pigment genes
2013
[15]
논문
Colonization History and Population Genetics of the Color-Polymorphic Hawaiian Happy-Face Spider Theridion Grallator (Araneae, Theridiidae)
2012-04
[16]
논문
Non-native spiders change assemblages of Hawaiian forest fragment kipuka over space and time
https://neobiota.pen[...]
2020-03-23
[17]
논문
Foraging behavior of the Hawaiian happy face spider (Araneae: Theridiidae)
https://academic.oup[...]
1994-11-01
[18]
논문
Can Hawaiian carnivorous caterpillars attack invasive ants or vice versa?
2010-12-08
[19]
논문
Temporal Foraging Patterns of Nonnative Coqui Frogs (Eleutherodactylus coqui) in Hawaii
https://meridian.all[...]
2016-12-01
[20]
논문
Costs and benefits of brood care in the Hawaiian happy face spider ''Theridion grallator'' (Araneae, Theridiidae)
https://www.jstor.or[...]
1990-04
[21]
논문
Two new species of ''Baeus'' (Hymenoptera: Scelionidae) from Southeastern Brazil parasitoids of ''Anelosimus'' (Araneae: Theridiidae)
http://dx.doi.org/10[...]
2006-03-30
[22]
문서
Taxon details ''Theridion grallator'' Simon, 1900
http://www.wsc.nmbe.[...]
Natural History Museum Bern
2016-01-30
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com