예쁜꼬마선충
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
예쁜꼬마선충은 생물학 연구에 널리 사용되는 작은 선충으로, 배아기, 유충기, 성체기를 거치는 생활사를 가진다. 자웅동체와 수컷의 두 가지 성을 가지며, 자가수정을 통해 번식하는 것이 특징이다. 예쁜꼬마선충은 단순한 구조와 짧은 수명, 투명한 몸체, 그리고 유전자 조작의 용이성 때문에 발생학, 유전학, 신경과학, 노화 연구 등 다양한 분야에서 모델 생물로 활용된다. 특히 세포 자살과 관련된 유전자 연구에 기여했으며, 인간 질병 연구에도 유용하게 사용되고 있다.
더 읽어볼만한 페이지
- 1900년 기재된 동물 - 웃는얼굴거미
웃는얼굴거미는 하와이 제도에서 발견되는 몸길이 약 5mm의 밝은 노란색 거미로, 사람의 웃는 얼굴을 닮은 무늬가 배에 있으며, 유전적, 환경적 요인에 의해 색상 변이를 보이고, 짝짓기, 번식, 양육 행동과 더불어 사회적 행동 및 기생 현상을 나타낸다. - 선형동물 - 편충
편충은 사람을 포함한 동물의 대장에 기생하며, 오염된 음식이나 물을 통해 감염되어 설사, 복통 등을 유발하고, 돼지 편충은 염증성 장 질환 치료에 연구되고 있는 기생충이다. - 선형동물 - 심장사상충
심장사상충은 모기를 통해 전파되어 개, 고양이 등의 심장과 폐동맥에 기생하며, 열대 및 온대 지역에 분포하고, 감염 시 호흡 곤란과 심부전 등을 유발하며, 예방약 투여가 권장된다. - 모델 생물 - 옥수수
옥수수는 멕시코 남부 원산의 벼과 식물로, 오래전부터 재배되어 전 세계로 퍼져나갔으며 식용, 사료용 등 다양한 용도로 활용되고, 옥수수 또는 maize라고 불린다. - 모델 생물 - 개
개는 회색늑대의 가축화된 아종으로, 오랜 가축화 과정을 거쳐 다양한 견종으로 분화되었으며, 반려동물과 다양한 직업 분야에서 인간과 함께 생활하며 여러 사회적 문제도 야기한다.
| 예쁜꼬마선충 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Caenorhabditis elegans |
| 학명 명명자 | Maupas, 1900 Dougherty, 1953 |
| 이명 | Rhabditis elegans Maupas, 1900 |
| 다른 이름 | C. elegans |
| 분류 | |
| 계 | 동물계 |
| 문 | 선형동물문 |
| 강 | 크로마도라강 |
| 아강 | 플레크티아아강 |
| 상목 | 랍디티스상목 |
| 목 | 간선충목 |
| 과 | 간선충과 |
| 속 | 카에노랍디티스속 |
| 특징 | |
| 크기 | 1mm (밀리미터) |
2. 생물학
예쁜꼬마선충은 선형동물로서, 분절되지 않은, 벌레 모양의, 좌우 대칭인 동물이다. 강한 외피인 큐티클, 네 개의 주요 표피 끈, 체액으로 채워진 가짜체강(체강)을 가지며, 더 큰 동물과 같은 일부 기관계를 가지고 있다.[17] 모든 선충류와 마찬가지로 순환계나 호흡계는 없다. 몸 전체 길이를 따라 달리는 네 개의 근육띠는 신경계에 연결되어 있으며, 머리를 제외하고는 몸을 등쪽이나 배쪽으로만 굽힐 수 있다.
지방 대사와 관련하여 예쁜꼬마선충은 포유류와 달리 특수한 지방 조직, 췌장, 간, 영양분을 전달하는 혈액이 없다. 중성 지질은 장, 표피, 배아에 저장되며, 표피는 주요 트리글리세리드 저장소로서 포유류의 지방세포에 해당한다.[18]
인두는 예쁜꼬마선충 머리에 있는 근육질의 먹이 펌프로서 단면이 삼각형이다. 음식을 갈아서 장으로 직접 운반하며, "밸브 세포"가 인두를 장에 연결한다. 소화 후 장 내용물은 직장을 통해 방출된다.[19] 인두와 배설관 사이에는 직접적인 연결이 없으며, 배설관은 액체 소변 방출 기능을 한다.
수컷은 단일엽 생식선, 정관, 교미침이 있는 특화된 꼬리를 가진다. 자웅동체는 두 개의 난소, 난관, 정자 저장소, 자궁을 가진다.
예쁜꼬마선충은 약 302개의 뉴런을 가지며, 이는 전체 체세포의 약 1/3에 해당한다.[20] 몸벽 근육에 연결되어 움직임을 조절하는 흥분성 콜린성 및 억제성 GABA성 운동 뉴런과 다양한 신경 전달 물질을 사용한다.[22]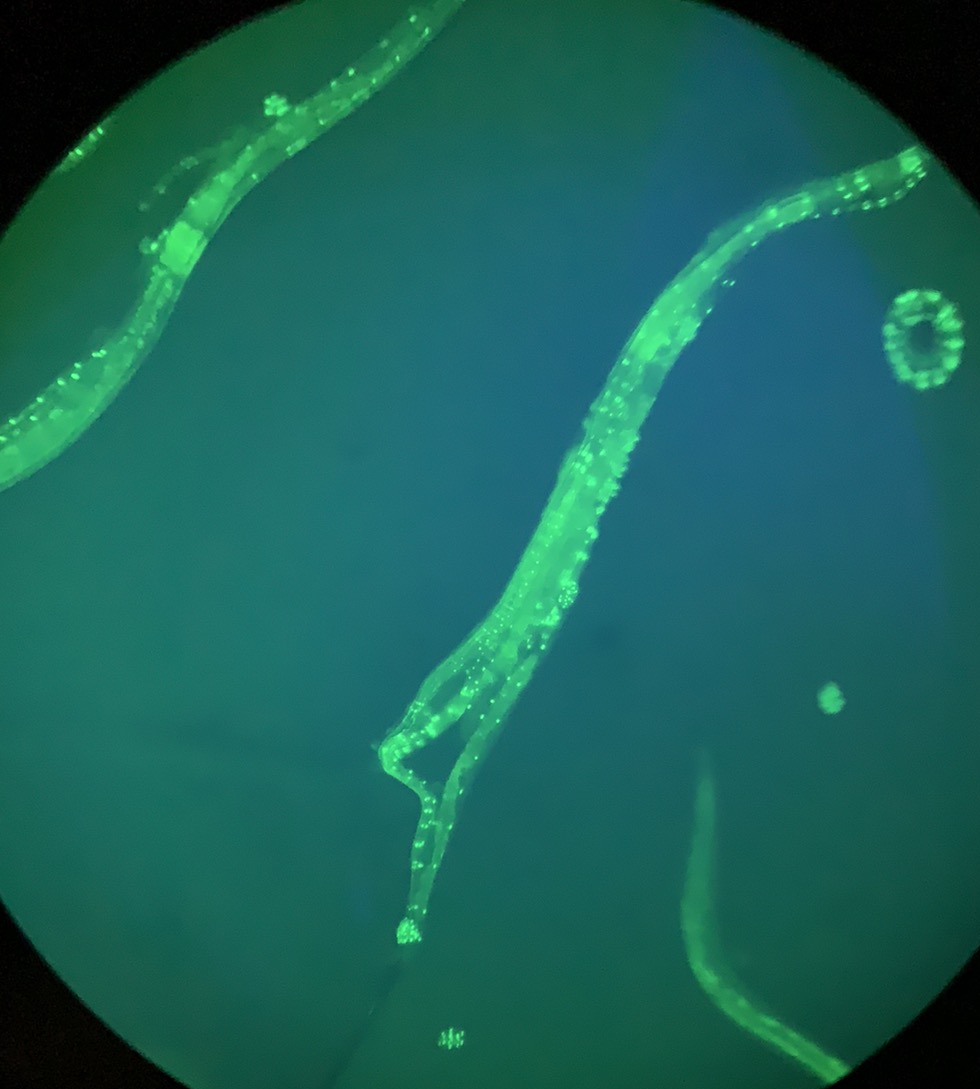
예쁜꼬마선충의 장에는 리소좀과 유사하지만 훨씬 큰 장 과립이 존재한다. 이 과립은 자외선 아래에서 강한 청색 형광을 방출하며, 선충이 죽으면 '죽음의 형광'이라는 강렬한 청색 형광을 방출한다. 장 과립은 글리코실화된 안트라닐산(AA) 형태의 청색 형광 물질을 포함하며, 항균 작용이나 광보호 기능을 할 수 있다.[23]

2. 1. 구조
예쁜꼬마선충은 다른 선형동물과 마찬가지로 체절이 없는 원통형 몸을 가지고 있으며 꼬리로 갈수록 가늘어진다. 입, 인두, 장, 생식선, 큐티클층을 가지고 있다. 예쁜꼬마선충의 단면은 외배엽에서 유래한 바깥 관과 내배엽에서 유래한 안쪽 관, 그리고 그 사이 공간인 의체강(pseudocoelomic space)으로 구성된다.[189] 바깥쪽 관은 큐티클층과 내피(hypodermis)층, 분비계, 신경계, 근육으로, 안쪽 관은 인두와 내장, 그리고 성체의 경우 생식기관으로 구성된다.[189]


예쁜꼬마선충 성체는 959개의 체세포와 1000~2000개의 생식세포를 가진다.[189] 몸은 콜라겐 성분 세포외 기질인 큐티클 층으로 덮여 있으며,[190][191] 이 큐티클은 내피에서 분비된다. 유충은 탈피 전 내피에서 오래된 큐티클 층 아래에 새 큐티클을 합성, 분비하고 오래된 큐티클 층은 탈피 과정에서 제거한다.[190][191]
예쁜꼬마선충은 분절되지 않은, 벌레 모양의, 좌우 대칭 동물이다. 큐티클(강한 외피, 외골격 같은), 네 개의 주요 표피 끈, 체액으로 채워진 가짜체강(체강)을 가지며, 더 큰 동물과 같은 일부 기관계를 가지고 있다.[17] 순환계나 호흡계는 없다. 몸 전체 길이를 따라 달리는 네 개의 근육띠는 신경계에 연결되어 몸을 등쪽 또는 배쪽으로만 굽힐 수 있다. 머리 부분에서는 네 개의 근육 사분면이 서로 독립적으로 연결되어 있다. 등쪽/배쪽 근육 수축 파동이 동물의 뒷부분에서 앞부분으로 진행되면 뒤로, 앞쪽에서 뒤쪽으로 진행되면 앞으로 추진된다. 이러한 굽힘 때문에 정상적인 살아있는 개체는 수평면을 가로지를 때 왼쪽이나 오른쪽으로 눕는 경향이 있다. 몸 큐티클 측면의 날개(alae)는 굽힘 동작 중 추가적인 견인력을 제공한다.[17]
지방 대사와 관련하여 예쁜꼬마선충은 포유류와 달리 특수한 지방 조직, 췌장, 간, 영양분 전달 혈액이 없다. 중성 지질은 장, 표피, 배아에 저장되며, 표피는 주요 트리글리세리드 저장소로 포유류의 지방세포에 해당한다.[18]
인두는 예쁜꼬마선충 머리에 있는 근육질 먹이 펌프로 단면이 삼각형이다. 음식을 갈아 장으로 직접 운반하며, "밸브 세포"가 인두를 장에 연결한다. 소화 후 장 내용물은 직장을 통해 방출된다.[19] 인두와 배설관 사이에는 직접적인 연결이 없으며, 배설관은 액체 소변 방출 기능을 한다.
수컷은 단일엽 생식선, 정관, 교미를 위한 교미침이 포함된 특화된 꼬리를 가진다. 자웅동체는 두 개의 난소, 난관, 정자 저장소, 자궁을 가진다.
예쁜꼬마선충은 약 302개의 뉴런을 가지며, 이는 전체 체세포의 약 1/3에 해당한다.[20] 많은 뉴런은 수상 돌기를 포함하며, 다른 뉴런과 시냅스 연결을 위해 신경 고리("뇌")로 뻗어 나온 돌기를 가진다.[21] 예쁜꼬마선충은 몸벽 근육에 연결되어 움직임을 조절하는 흥분성 콜린성 및 억제성 GABA성 운동 뉴런을 가진다. 또한, 이러한 뉴런과 다른 뉴런(예: 중간 뉴런)은 다양한 신경 전달 물질을 사용하여 행동을 조절한다.[22]
예쁜꼬마선충의 장에는 리소좀과 유사하지만 훨씬 큰 세포내이입 능력을 갖는 장 과립이 존재한다. 이 과립은 자외선 아래에서 강한 청색 형광을 방출하며, 선충이 죽으면 '죽음의 형광'이라는 강렬한 청색 형광을 방출한다. 장 과립은 글리코실화된 안트라닐산(AA) 형태의 청색 형광 물질을 포함하며, 항균 작용이나 광보호 기능을 할 수 있다.[23]
체세포 수는 자웅동체 성체의 경우 959개(단, 이는 핵의 수이며, 표피세포는 다핵이므로 세포 수는 더 적다)이고, 수컷의 경우 1033개이다.[175][176][177][178] 자웅동체의 장내에 공생 미생물이 풍부한 환경에서는 자손의 장세포가 1~3개 정도 증가할 가능성이 있다.[178] 예쁜꼬마선충의 세포 수를 셀 때, 세포융합에 의해 다핵세포가 되는 표피세포는 세포가 아닌 세포핵을 세지만, 세포분열을 수반하지 않는 핵분열에 의해 다핵세포가 되는 장세포는 세포핵이 아닌 세포를 센다.[175][176]
신경, 근육, 소화관, 표피, 생식소의 조직과 기관을 가진다. 체표는 각질하층이 분비하는 두꺼운 큐티클에 덮여 있다. 체내에는 가체강을 가지고 있다. 표피 안쪽을 종주근(체벽근)이 달린다.
소화관은 입에서 식도, 판을 지나 장, 직장을 거쳐 항문으로 이어진다. 식도는 식도 전부(corpus), 식도협(isthmus) 및 마개가 있는 후부 식도구(postcorporal bulb)로 나뉜다.[166] 구기에는 가동성 장식이 없다.[166]
신경계는 감각세포와 운동뉴런으로 구성된다. 복측신경삭이 발달하고, 중추인 신경환이 식도의 중앙부를 둘러싸고 있다.[166] 신경세포는 302개이며, 7600개의 시냅스가 모두 확인되었다. 이 정도 세포 수로도 물리적 자극 회피, 화학 물질/온도와 먹이의 연관 학습, 벤즈알데히드 등 유인성 휘발성 물질에 대한 순응 등의 행동을 보인다.[179][180][181] 개별 신경이 어떤 세포와 시냅스 또는 갭 정션을 형성하는지가 투과형 전자 현미경의 연속 절편 영상으로부터 완전히 재구성되었고, 레이저로 특정 신경세포를 파괴하는 실험 등을 통해 어떤 신경세포가 어떤 행동과 관련되는지도 알려져 있다.
성염색체에 의한 성 결정은 XO형이다. XX 개체는 자웅동체(두 종류의 생식 세포를 가진 암컷)가 되고, XO 개체는 수컷이 된다.[182]
수컷은 음경과 항문이 공유된 개구부인 총배출구를 가지며, 교접낭(bursa)가 발달한다.[166] 정자는 작고 둥근 편모가 없는 세포이며, 아메바와 같은 움직임으로 운동한다.
자웅동체는 항문과는 별도로 음문(vulva)을 가진다.[166] 유충기에 약 300개의 정자를 만들고, 성체가 되면 난자를 형성한다. 정자는 자궁 내 수정낭(저정낭)에 보관하며,[166] 보관해 둔 정자를 사용하여 자가수정을 한다.[182] 난자의 세포막과 정자가 융합하면, 수정된 난자에서는 키틴이 급속히 합성되어 다정(多精)이 억제된다. 수정란은 초기 분열 후 음문에서 밀려 나온다.
하나의 개체가 산란하는 자손은 약 300마리이다. 자웅동체이기 때문에 실험상 유전적 배경을 균일하게 하는 데 도움이 된다. 수컷은 약 0.1% 비율로 나타나며, 자웅동체와 교배 가능하다.
몸의 절반 이상을 차지하는 생식계열 세포는 1000개가 넘는 경우도 있다.
2. 2. 유전자
예쁜꼬마선충은 다세포 생물 중 최초로 전체 DNA의 염기서열이 분석된 생물이다. 약 1억 개의 염기 쌍을 가진 DNA를 가지고 있으며, 5쌍의 상염색체(I, II, III, IV, V)와 1쌍의 성염색체(X)를 가지고 있다. 진핵 생물이지만 원핵생물의 유전자에서 나타나는 오페론 형태의 유전자 배열을 가지고 있다.[192][193]예쁜꼬마선충의 유전자는 20,470개의 단백질을 암호화(encoding)하고 있다.[192] RNA 간섭(RNAi)은 특정 유전자의 기능을 방해하여 그 유전자의 기능을 추론할 수 있게 해주는 연구 방법이다. 예쁜꼬마선충은 RNAi 연구에 유용하게 사용되는 모델 생물이다.[88][89][90]
예쁜꼬마선충의 전유전체 서열은 1998년에 발표되었으며,[132] 2002년 10월에 최종 완성되었다. 예쁜꼬마선충 게놈은 약 20,470개의 단백질-코딩 유전자를 포함하고 있으며, 유전자 밀도는 5킬로염기쌍당 약 1개의 유전자이다. 인트론은 게놈의 26%를 차지한다.[136] 예쁜꼬마선충과 다른 선충류는 오페론을 가진 몇 안 되는 진핵생물에 속한다.[137]
예쁜꼬마선충 유전자의 약 35%는 사람의 상동 유전자를 가지고 있다. 사람 유전자가 예쁜꼬마선충에 도입되었을 때 예쁜꼬마선충 상동 유전자를 대체하는 것으로 나타났으며, 많은 예쁜꼬마선충 유전자는 포유류 유전자와 유사하게 기능할 수 있다.[47]
Tc1 전좌인자는 ''C. elegans''에서 활동하는 DNA 전좌인자이다.
2. 3. 성
예쁜꼬마선충은 크게 자웅동체와 수컷 두 가지 성별을 가진다. 자웅동체는 성 염색체 쌍(XX)을 가지며, 수컷(XO)은 감수 분열 시 염색체 비분리 현상으로 인해 성 염색체를 하나만 가지게 된다. 이러한 염색체 이상은 드물기 때문에 자연에서는 자웅동체가 대부분이고 수컷은 0.1% 정도로 매우 적다.[189]예쁜꼬마선충은 수컷이 있으면 유성생식을 하지만, 수컷이 없으면 자웅동체가 자신의 정자와 난자를 수정시켜 자신과 동일한 유전자를 가진 개체를 만든다.[189] 즉, 자가수정을 통해 번식한다.
자웅동체는 유충기에 약 300개의 정자를 만들고 성체가 되면 난자를 형성한다. 정자는 자궁 내 수정낭에 보관되며,[166] 보관된 정자를 이용해 자가수정을 한다.[182] 수컷은 자웅동체에 정액을 주입할 수 있으며, 자웅동체는 수컷 정자를 우선적으로 사용한다.[24]
예쁜꼬마선충의 정자는 편모가 없는 아메바 형태이며,[26] 자가 수정 시 약 300개의 알을 낳고, 수컷에 의해 수정되면 1,000개 이상의 자손을 낳을 수 있다. 자웅동체는 일반적으로 다른 자웅동체와 교배하지 않는다.
예쁜꼬마선충의 성 결정은 X0 성 결정 시스템에 기반한다. 자웅동체는 한 쌍의 성 염색체(XX)를, 수컷은 하나의 성 염색체(X0)를 가진다.[27]
수컷은 음경과 항문이 공유된 총배출구를 가지며, 교접낭이 발달한다.[166] 자웅동체는 항문과 별도로 음문을 가진다.[166]
2. 4. 생활 주기
예쁜꼬마선충의 삶은 크게 배아기, 유충기, 성체기로 나뉜다. 먼저 어미 선충의 자궁 안에서 알의 상태로 난할을 계속하여 상실기 단계(세포 수가 30개 정도)에 이른다. 알을 낳은 뒤에는 포배기 단계를 거치면서 세포들이 이동하여 배엽들을 형성한다. 외배엽은 피하 조직과 신경을, 중배엽은 인두와 근육을, 내배엽은 생식세포와 내장을 형성한다. 배아기가 거의 끝나갈 때 대략적인 몸의 형태를 갖추게 된다.[189]유충기는 크게 4단계로 구성되어 있으며 유충은 주변 환경에서 영양소를 섭취하면서 각 단계를 거쳐 성체로 성장한다. 유충기 동안에는 아직 완성되지 않은 신경 시스템과 생식기관이 발달한다. 유충은 주변 환경에 따라 몸의 상태를 변화시켜 그 환경에 적응할 수 있는데, 이러한 몸의 상태 변화는 발달과정에서 2번 일어날 수 있다. 먼저 제 1령 유충일 때 섭취할 수 있는 먹이가 없어 환경이 악화되면 유충은 몸의 발달을 멈추게 된다. 이러한 상태의 유충은 먹지 않고 6~10일을 견딜 수 있다. 또한 제 2령 유충의 마지막 단계에서 유충은 환경에 따라서 다우어(dauer) 유충 단계로 들어갈 수 있다. 다우어 유충기는 먹이가 부족하거나 특정 장소에서 개체의 밀도가 높을 때, 그리고 온도가 특정 수준 이상으로 높아졌을 때 발생한다.[194]
다우어 유충기로 들어가게 되면 4달 정도를 아무것도 섭취하지 않고 버틸 수 있다.[189] 다우어 유충의 몸은 다른 유충기에 비해서 가늘어지고 큐티클 층의 성분이 바뀌고 두께가 두꺼워지며 활동량이 극히 적어진다. 그리고 TCA 회로보다는 저장된 지방으로부터 에너지 대사를 하는 글리옥실레이트(glyoxylate) 회로가 사용된다. 또한 전체적으로 mRNA 전사율을 낮게 유지하여 쓸데없는 전사를 막아 에너지와 물질 소모를 막는다. 반면에 외부의 스트레스에 저항하는데 필요한 Hsp90(heat shock protein 90)과 산화 스트레스로부터 세포를 보호하는 효소(superoxide mutase와 카탈라아제)에 해당되는 mRNA의 전사 비율은 더 높아진다. 예쁜꼬마선충은 위와 같은 과정을 통해서 에너지 사용량을 낮추고 외부의 스트레스에 대한 저항력을 키운다.[194]
번식에 유리한 환경 조건 하에서 부화한 유충은 20 °C에서 단 3일 만에 L1, L2, L3 및 L4의 네 가지 유충 단계를 거친다. 먹이 부족, 과도한 개체 밀도 또는 고온과 같은 스트레스 조건에서는 ''예쁜꼬마선충''이 대체 3기 유충 단계인 L2d 단계, 즉 다우어 단계(독일어로 "영구적인"이라는 뜻)에 들어갈 수 있다. 다우어 유충은 스트레스에 강하며, 가늘고 특징적인 다우어 큐티클로 입이 막혀 먹이를 섭취할 수 없다. 이 단계는 몇 달 동안 지속될 수 있다.[47][48] 조건이 개선되어 유충의 성장을 촉진하면, 생식선 발달은 L2 단계에서 정지되어 있지만, 이제 L4 단계로 탈피한다.[49]
각 단계 전이는 벌레의 투명한 큐티클의 탈피에 의해 구분된다.

3. 생태학
예쁜꼬마선충은 비료나 정원의 흙과 같이 주로 사람이 사는 곳과 가까운 곳에서 발견되지만, 자연 내 서식지는 거의 알려지지 않았다. 최근에는 썩은 과일과 같은 유기 물질이 많은 곳에서 발견된다고 알려져 있다.[195] 흙 속의 유기 물질과 미생물을 먹고 살며, 구체적인 생태는 파악되지 않았다.[196]
간균류는 예쁜꼬마선충이 섭취할 수 없으며, 아메바의 특정 종의 포자는 예쁜꼬마선충이 소화시킬 수 없다는 것이 알려져 있다.[196][197] 예쁜꼬마선충은 환경이 악화되었을 때 다우어 유충기에 접어들며, 다우어 유충은 등각류, 절지류, 노래기류나 달팽이 같은 동물에 붙어서 장거리를 이동할 수 있다. 환경이 좋아지면 그 동물로부터 떨어져 나와 성장을 계속한다.[196]
다양한 ''예쁜꼬마선충속'' 종들은 다양한 영양분과 박테리아가 풍부한 환경에 서식한다. 이들은 부패하는 유기물에서 자라는 박테리아를 먹고 산다(미생물 섭식). 이들은 박테리아와 박테리아가 분비하는 대사산물(예: 철 시데로포어)을 감지할 수 있는 화학 감각 수용체를 가지고 있어 박테리아 먹이 쪽으로 이동할 수 있다.[59] 토양은 자체적으로 지속 가능한 개체군을 유지하기에 충분한 유기물이 부족하다. ''예쁜꼬마선충''은 다양한 박테리아를 먹고 살 수 있지만, 야생 생태는 대부분 알려져 있지 않다. 대부분의 실험실 균주는 정원과 퇴비 더미와 같은 인공 환경에서 채취되었다. 최근에 ''예쁜꼬마선충''이 다른 종류의 유기물, 특히 썩은 과일에서 번성하는 것으로 밝혀졌다.[60]
''예쁜꼬마선충''은 특히 미세한 나노플라스틱과 같은 오염 물질을 섭취할 수 있으며, 이는 항생제 내성 박테리아와의 연관성을 가능하게 하여 ''예쁜꼬마선충''에 의해 토양 전체로 나노플라스틱과 항생제 내성 박테리아가 퍼지는 결과를 초래할 수 있다.[61] ''예쁜꼬마선충''은 ''크립토코쿠스 로렌티''와 ''C. kuetzingii''를 포함한 여러 종의 효모를 유일한 먹이원으로 사용할 수도 있다.[62] 세균 섭식자이지만, ''예쁜꼬마선충''은 ''황색포도상구균''[63], ''녹농균''[64], ''살모넬라 엔테리카'' 또는 ''장구균''[65]을 포함한 여러 병원성 박테리아에 의해 죽을 수 있다. 병원성 박테리아는 생물막을 형성할 수도 있는데, 이 생물막의 점착성 엑소폴리머 매트릭스는 ''예쁜꼬마선충''의 운동성을 방해하고[66] 포식자의 탐지로부터 박테리아 쿼럼 센싱 유인 물질을 가릴 수 있다.[67]
지네류, 곤충류, 등각류류, 복족류류와 같은 무척추동물은 다우어 유충을 다양한 적합한 장소로 운반할 수 있다. 유충은 숙주가 죽으면 숙주를 먹는 것도 관찰되었다.[68]
선충류는 건조 상태에서도 생존할 수 있으며, ''예쁜꼬마선충''의 경우 이러한 능력의 메커니즘이 후기 배 발생 풍부 단백질임이 입증되었다.[69]
''예쁜꼬마선충''은 다른 선충류와 마찬가지로 포식성 선충류와 일부 곤충을 포함한 다른 잡식성 동물의 먹이가 될 수 있다.[70]
오르세이 바이러스는 ''예쁜꼬마선충''에 영향을 미치는 바이러스이며, 예쁜꼬마선충 Cer1 바이러스[71]와 예쁜꼬마선충 Cer13 바이러스도 마찬가지이다.
'''균류와의 상호작용'''
야생에서 분리된 ''예쁜꼬마선충''은 미포자충 균류 감염이 자주 발견된다. 그러한 종 중 하나인 ''네마토시다 파리시이''는 ''예쁜꼬마선충''의 장에서 복제된다.[72]
''아르트로보트리스 올리고스포라''는 균류와 선충류 사이의 상호 작용에 대한 모델 생물이다.[73] 가장 흔하고 널리 퍼져있는 선충 포획 균류이다. 선형동물문의 전통적인 분류에서 쌍기강은 대부분 자유생활성이지만, 이 종이 속한 쌍선강 선충류는 대부분의 종이 기생성이다.[174] 그러나 이 종은 자유생활성이다.[174] 토양에 서식하며 세균을 먹는 세균섭식성이다. 실험실에서는 한천배지에 배양한 대장균을 영양원으로 하여 사육된다.
4. 연구 분야에서의 활용
시드니 브레너는 1963년 동물의 신경 발달 연구를 위해 예쁜꼬마선충을 모델 생물로 사용할 것을 제안했다.[74] 브레너는 대량 배양이 쉽고 유전자 분석에 편리하다는 점 때문에 예쁜꼬마선충을 선택했다.[82] 예쁜꼬마선충은 다세포 진핵생물이지만, 현미경으로 자세히 관찰하기에 충분할 만큼 단순하며, 몸이 투명하여 세포 분화 및 기타 발달 과정을 온전한 생물체에서 연구하기 쉽다.[12] 또한, 번식 비용이 저렴하고 냉동 보관 후 해동해도 생존하여 장기간 보관이 가능하다는 장점이 있다.[12]
예쁜꼬마선충은 신경계, 대사 질환, 약물 및 독소 평가, 우주 연구 등 다양한 분야에서 활용되고 있다.
- 신경계 연구: 예쁜꼬마선충은 신경계를 가진 가장 단순한 생물 중 하나로, 자웅동체의 경우 302개의 뉴런으로 구성되어 있다.[75] 뉴런 연결망인 커넥톰은 자세하게 지도로 작성되었으며,[76][77] 소세계 네트워크를 이룬다는 것이 밝혀졌다.[78] 화학주성, 온도주성, 역학수용, 학습, 기억, 짝짓기 등 다양한 행동을 조절하는 신경 및 분자 메커니즘을 연구하는 데 활용된다.[79]
- 우주 연구: 우주왕복선 컬럼비아호 참사에서 생존한 표본이 발견되면서 우주 연구에도 활용되기 시작했다.[127] 국제우주정거장에서 근육 위축증의 유전적 기초 등 근육 발달과 생리에 대한 무중력의 영향을 탐구하는 연구가 진행되었다.[128]
예쁜꼬마선충 연구는 여러 차례 노벨 생리학·의학상과 노벨 화학상 수상으로 이어졌다. 2002년에는 기관 발생의 유전학 및 프로그램된 세포사 연구,[151] 2006년에는 RNA 간섭 발견,[151] 2024년에는 마이크로RNA와 유전자 조절에서의 역할 발견[152]으로 노벨 생리학·의학상이 수여되었다. 2008년에는 녹색 형광 단백질 연구로 마틴 챌피가 노벨 화학상을 받았다.[152]
4. 1. 예쁜꼬마선충의 특징
예쁜꼬마선충은 정상적인 조건에서 사멸 주기가 3주 정도로 짧고, 실험실에서 한천 접시나 대장균 배양액에서 쉽게 배양할 수 있으며 유지비가 적게 든다.[198] 또한 얼려서 보관할 수 있어 장기간 보관이 가능하다.[199] 짧은 생활 주기와 300개체가 넘는 많은 후손을 생성하며, 몸이 투명하고 체세포의 개수가 1000개 정도밖에 되지 않아 세포 관찰에 유용하다.[200] 유전자 연구가 집중적으로 이루어져 모든 유전자의 서열이 밝혀졌고, 유전자 결여 돌연변이 라이브러리나 돌연변이 유발, RNA 간섭, 유전자 이식 등에 관한 연구가 많이 진행되어 자료가 풍부하여 연구가 용이하다. 특히 RNA 간섭을 통해 특정 유전자의 발현을 억제할 수 있다는 장점이 있다.[201] 신경계가 단순하여 신경 연구 모델로도 많이 사용된다.''C. elegans''는 분절되지 않은, 벌레 모양의, 좌우 대칭인 생물이다. 큐티클(강한 외피, 외골격과 같은), 네 개의 주요 표피 끈, 그리고 체액으로 채워진 가짜체강(체강)을 가지고 있다. 또한, 더 큰 동물과 같은 일부 기관계를 가지고 있다. 약 천 마리 중 한 마리가 수컷이고 나머지는 자웅동체이다.[17] ''C. elegans''의 기본 해부학적 구조는 입, 인두, 장, 생식선, 그리고 콜라겐성 큐티클을 포함한다. 모든 선충류와 마찬가지로 순환계나 호흡계가 없다. 몸 전체 길이를 따라 달리는 네 개의 근육띠는 신경계에 연결되어 있으며, 이를 통해 근육은 동물의 몸을 등쪽 굽힘이나 배쪽 굽힘으로만 움직일 수 있지만, 머리를 제외하고는 좌우로 움직일 수 없다. 머리 부분에서는 네 개의 근육 사분면이 서로 독립적으로 연결되어 있다. 등쪽/배쪽 근육 수축의 파동이 동물의 뒷부분에서 앞부분으로 진행되면 동물은 뒤로 추진된다. 수축의 파동이 앞쪽에서 시작되어 몸을 따라 뒤쪽으로 진행되면 동물은 앞으로 추진된다. 이러한 등쪽/배쪽 편향의 몸 굽힘 때문에, 정상적인 살아있는 움직이는 개체는 수평면을 가로지를 때 왼쪽이나 오른쪽으로 옆으로 누워 있는 경향이 있다. 몸 큐티클의 측면에 있는 일련의 융기부인 날개(alae)는 이러한 굽힘 동작 중에 동물에게 추가적인 견인력을 제공하는 것으로 여겨진다.
지방 대사와 관련하여 ''C. elegans''는 포유류와 비교하여 특수한 지방 조직, 췌장, 간, 또는 영양분을 전달하는 혈액이 없다. 중성 지질은 대신 장, 표피 및 배아에 저장된다. 표피는 주요 트리글리세리드 저장소로서 포유류의 지방세포에 해당한다.[18]
인두는 ''C. elegans'' 머리에 있는 근육질의 먹이 펌프로서 단면이 삼각형이다. 이것은 음식을 갈아서 장으로 직접 운반한다. 일련의 "밸브 세포"가 인두를 장에 연결하지만, 이 밸브가 어떻게 작동하는지는 알려져 있지 않다. 소화 후, 장 내용물은 직장을 통해 방출되며, 이는 다른 모든 선충류의 경우와 같다.[19] 인두와 배설관 사이에는 직접적인 연결이 없으며, 배설관은 액체 소변의 방출 기능을 한다.
수컷은 단일엽 생식선, 정관, 그리고 교미를 위해 특화된 꼬리를 가지고 있으며, 여기에는 교미침이 포함된다. 자웅동체는 두 개의 난소, 난관, 정자 저장소, 그리고 단일 자궁을 가지고 있다.
''C. elegans''에는 약 302개의 뉴런이 있으며, 이는 전체 신체의 모든 체세포의 약 1/3에 해당한다.[20] 많은 뉴런은 신경 전달 물질이나 다른 신호를 받기 위해 세포에서 뻗어 나온 수상 돌기를 포함하며, 다른 뉴런과 시냅스 연결을 위해 신경 고리("뇌")로 뻗어 나온 돌기를 가지고 있다.[21] ''C. elegans''는 몸벽 근육에 연결되어 움직임을 조절하는 흥분성 콜린성 및 억제성 GABA성 운동 뉴런을 가지고 있다. 또한, 이러한 뉴런과 다른 뉴런(예: 중간 뉴런)은 다양한 신경 전달 물질을 사용하여 행동을 조절한다.[22]
4. 2. 세포 자살 연구 분야
예쁜꼬마선충은 세포 수가 적어 세포 발달 과정을 관찰하기 용이하여 세포자살 연구에 많이 사용되는 모델 생물이다. H. 로버트 호비츠는 예쁜꼬마선충의 발달 과정에서 1090개의 세포 중 131개가 프로그램된 세포 자살(apoptosis)을 한다는 것을 발견했다.[202] 또한, 예쁜꼬마선충에서 발견된 세포 자살 관련 유전자(ced-3, ced-4, ced-9)와 유사한 유전자들이 인간에게도 존재한다는 것이 밝혀졌다.[203]프로그램 세포사멸(아포토시스)은 예쁜꼬마선충(자웅동체)에서 131개의 세포(대부분 뉴런이 될 세포)를 제거한다.[87] 이러한 예측 가능한 아포토시스 덕분에, 세포 사멸 촉진 유전자와 억제 유전자가 밝혀졌다.[87]
이러한 연구를 통해 세포 자살에 대한 이해가 높아졌고, 암과 같은 질병과의 연관성이 밝혀지면서 생물학 및 약학계에 큰 영향을 주었다.[203] 로버트 호로비츠는 시드니 브레너, 존 설스턴과 함께 예쁜꼬마선충을 이용한 기관 발생과 아포토시스 유전자 조절 연구로 2002년 노벨 생리학·의학상을 수상했다.
4. 3. 질병
예쁜꼬마선충은 질병 연구에도 많이 사용된다. 예쁜꼬마선충과 인간의 질병이 직접적인 연관은 없지만, 여러 변인들을 통제하기 쉽고 빠른 결과를 얻을 수 있기 때문이다. 또한 인간과 예쁜꼬마선충의 유전자를 비교한 결과 60~80% 정도의 유전자가 비슷하며, 인간 질병 관련 유전자 533개가 예쁜꼬마선충에게서 발견되어, 예쁜꼬마선충 연구가 인간 질병 연구로 이어질 수 있음을 보여준다.[204] 예쁜꼬마선충 연구를 통해 질병의 분자적 기작을 발견할 수 있다는 점도 질병 연구 대상으로 많이 사용되는 이유이다.[204]예쁜꼬마선충은 당뇨병이나 비만 연구와 같은 에너지 대사 증후군 연구,[205] 신호전달 체계와 관련된 암 연구, 알츠하이머 병, 헌팅턴 병, 파킨슨 병과 같은 신경 분해성 질환(neurodegration disease) 연구, 근 위축증같은 유전병연구, 우울증과 같은 신경관련 질환 연구에 사용된다.[204]
RNA 간섭(RNAi)은 특정 유전자의 기능을 방해하는 비교적 간단한 방법이다. 유전자의 기능을 침묵시키는 것은 때때로 연구자가 그 유전자의 가능한 기능을 추론할 수 있게 해준다. 예쁜꼬마선충은 관심 있는 이중 가닥 RNA를 발현하는 유전적으로 형질전환된 박테리아에 담그거나, 주입하거나,[88] 또는 먹일 수 있으며, 그 서열은 연구자가 비활성화하려는 유전자의 서열을 보완한다.[89] RNAi는 기능 유전체학 연구에서 강력한 도구로 부상했다. 예쁜꼬마선충은 유전자 기능을 분석하고 체계적인 유전적 상호 작용에 대한 미래의 발견 가능성을 주장하는 데 사용되었다.[90]
니코틴에 대한 의존성 또한 예쁜꼬마선충을 이용하여 연구할 수 있는데, 이는 예쁜꼬마선충이 포유류와 유사한 니코틴에 대한 행동 반응을 보이기 때문이다. 이러한 반응에는 급성 반응, 내성, 금단 증상 및 민감화가 포함된다.[99] 크기가 작고 수명 주기가 짧은 예쁜꼬마선충은 다세포 생물에서 약물 및 독소의 화학 라이브러리를 평가하기 위한 생체 내 고처리량 스크리닝(HTS) 플랫폼을 가능하게 하는 소수의 유기체 중 하나이다.[125] 예쁜꼬마선충에서 관찰 가능한 인간 질병의 상동 표현형은 기존 승인 약물의 인간 치료 적응증에 대한 재목적화 가능성을 알려줄 수 있는 약물 라이브러리 프로파일링을 가능하게 할 수 있다.[126]
2015년 규슈 대학의 연구 그룹은, 예쁜꼬마선충을 사용하여 피험자의 소변 냄새를 이용하여 조기에 고정확도로 암 검진에 성공했다는 것을 발표했다.[186]
4. 4. 노화 연구 분야
예쁜꼬마선충은 노화 연구에 중요한 모델 생물로 사용되어 왔다. 인슐린 유사 성장 인자 신호 전달 경로를 억제하면 성체의 수명이 3배 증가하는 반면,[101][102] 포도당 섭취는 산화 스트레스를 유발하여 수명을 절반으로 줄인다.[81] 인슐린/IGF-1 수용체의 유도 분해는 늦은 시기에 벌레의 기대 수명을 크게 연장시킨다.[103]''예쁜꼬마선충''의 유전자 중 *age-1* 유전자에 돌연변이가 발생하면 평균 수명이 약 2배 증가하는데, 이는 체내 슈퍼옥사이드 디스뮤타제(SOD)와 카탈라아제 양이 증가하여 산화 스트레스를 견디는 능력이 향상되기 때문이다.[206] 또한, 나이가 들면서 손상되는 미토콘드리아 유전자가 *age-1* 돌연변이에서는 덜 손상된다는 사실도 밝혀졌다.[207]
장수하는 ''예쁜꼬마선충'' 돌연변이체는 산화 스트레스와 자외선에 대한 저항성이 높고,[104] 야생형보다 DNA 복구 능력이 뛰어나다.[1] 뉴클레오티드 절단 복구 유전자 *xpa-1*의 기능이 저하되면 자외선에 대한 민감도가 증가하고 장수 돌연변이체의 수명이 감소한다.[1] 이러한 결과는 DNA 복구 능력이 장수의 기초가 됨을 시사한다.
엑소좀을 통한 슈퍼옥사이드 디스뮤타제(SOD) 전달은 활성 산소종(ROS) 수준을 감소시켜 수명을 연장하고 노화를 지연시킨다.[105] 뉴클레오티드 절단 복구 과정에 의한 DNA 손상 복구 능력은 나이가 들면서 감소한다.[106]
5mM 염화 리튬(LiCl)에 노출된 ''예쁜꼬마선충''은 수명이 연장되었고,[107] 10μM LiCl 노출 시 사망률 감소가 관찰되었다.[108]
''예쁜꼬마선충''은 알츠하이머병 관련 유전자(예: 프레세닐린) 기능을 확인하는 데 중요한 역할을 했다.[109] 또한, RNA 결합 단백질이 생식세포와 초기 배아 발달에 필수적임을 밝혀냈다.[110]
텔로미어는 다양한 생물에서 수명 연장 및 노쇠 지연과 관련이 있는데,[111][112][113] ''예쁜꼬마선충''은 다른 진핵생물과 유사하게 텔로미어를 유지한다. 초파리와는 다르게 ''예쁜꼬마선충''은 레트로트랜스포존을 사용하지 않는다.[114] 텔로머라제의 촉매적 소단위 (*trt-1*) 녹아웃 시, ''예쁜꼬마선충''은 대체 텔로미어 연장(ALT) 능력을 얻는다.[115] ''예쁜꼬마선충''은 전형적인 텔로머라제 경로 녹아웃 후 ALT 기능을 얻은 최초의 진핵생물이다.[115] ALT는 임상 암의 약 10-15%에서 관찰되므로,[116] ''예쁜꼬마선충''은 ALT 연구의 주요 대상이다.[117][118][119] 텔로미어가 짧아지는 동안에도 벌레의 수명이 약간 증가하는 현상이 관찰되었는데, 이는 *trt-1* 과발현 시 텔로미어가 역설적으로 짧아지기 때문이며, 이는 거의 불임에 이른다.[120]
참조
[1]
논문
Modes et formes de reproduction des nématodes
[2]
논문
Les modalités de la reproduction et le déterminisme du sexe chez quelques nematodes libres.
1949
[3]
논문
Spontaneous unstable unc-22 IV mutations in C. elegans var. Bergerac
1984-12-01
[4]
논문
Tc1 transposition and mutator activity in a Bristol strain of ''Caenorhabditis elegans''
1990-06-01
[5]
논문
Structural analysis of Tc1 elements in Caenorhabditis elegans var. Bristol (strain N2)
1989-07-01
[6]
서적
The Nematode ''Caenorhabditis elegans''
Cold Spring Harbor Laboratory Press
[7]
논문
Phylogeny of ''Rhabditis'' subgenus ''Caenorhabditis'' (Rhabditidae, Nematoda)
[8]
문서
[9]
웹사이트
''Caenorhabditis elegans''
http://plpnemweb.ucd[...]
University of California, Davis
2013-11-19
[10]
논문
A cladistic analysis of pseudocoelomate (aschelminth) morphology.
1996
[11]
웹사이트
Introduction to sex determination
http://www.wormbook.[...]
2017-03-15
[12]
논문
The genetics of ''Caenorhabditis elegans''
1974-05-01
[13]
논문
The structure of the nervous system of the nematode ''Caenorhabditis elegans''
1986-11-01
[14]
논문
Getting into the mind of a worm--a personal view
2013-06-01
[15]
잡지
The Connectome Debate: Is Mapping the Mind of a Worm Worth It?
https://www.scientif[...]
2014-01-18
[16]
뉴스
These Tiny Worms Account for at Least 4 Nobel Prizes
https://www.nytimes.[...]
New York Times
2024-10-17
[17]
서적
Molecular Biology of the Cell
Garland Science
[18]
논문
Investigating Connections between Metabolism, Longevity, and Behavior in Caenorhabditis elegans
2016-08-01
[19]
웹사이트
The ''C. elegans'' pharynx: a model for organogenesis
http://www.wormbook.[...]
2017-03-15
[20]
서적
Neurotoxicity of Metals
[21]
웹사이트
About the nematode Caenorhabdtis elegans
https://web.archive.[...]
[22]
논문
Specification of the nervous system
[23]
논문
The mysterious case of the ''C. elegans'' gut granule: death fluorescence, anthranilic acid and the kynurenine pathway
[24]
논문
fog-2 and the evolution of self-fertile hermaphroditism in ''Caenorhabditis''
2005-01-01
[25]
논문
Serotonin-deficient mutants and male mating behavior in the nematode ''Caenorhabditis elegans''
1993-12-01
[26]
논문
Transformation: how do nematode sperm become activated and crawl?
2012-10-01
[27]
논문
A CUL-2 ubiquitin ligase containing three FEM proteins degrades TRA-1 to regulate C. elegans sex determination
2007-07-01
[28]
웹사이트
Handbook - Male Introduction
https://www.wormatla[...]
2021-03-30
[29]
논문
C. elegans outside the Petri dish
2015-03-01
[30]
논문
The TRA-1A Sex Determination Protein of C. elegans Regulates Sexually Dimorphic Cell Deaths by Repressing the egl-1 Cell Death Activator Gene
1999-08-06
[31]
논문
The evolution of nematode sex determination: C. elegans as a reference point for comparative biology
2005-12-01
[32]
논문
xol-1: a gene that controls the male modes of both sex determination and X chromosome dosage compensation in C. elegans
1988-10-01
[33]
웹사이트
mix-1 (gene) - WormBase : Nematode Information Resource
https://wormbase.org[...]
2021-04-23
[34]
논문
Sexual and temporal dynamics of molecular evolution in C. elegans development
2005-01-01
[35]
논문
Specification of the anteroposterior axis in Caenorhabditis elegans
https://pubmed.ncbi.[...]
1996-05-01
[36]
서적
Developmental biology
Sinauer
[37]
논문
par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed
1995-05-00
[38]
논문
Asymmetric cell division and axis formation in the embryo
2005-10-00
[39]
간행물
Germline proliferation and its control
https://www.ncbi.nlm[...]
2005-08-15
[40]
백과사전
WBbt:0006773 (anatomy term)
2014-05-14
[41]
서적
Developmental biology
Sinauer
[42]
논문
Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm
1997-08-00
[43]
논문
Chiral forces organize left-right patterning in C. elegans by uncoupling midline and anteroposterior axis
2010-09-00
[43]
논문
Quantification of sebaceous excretion in volunteers: influence of chronological age, sex and race
[43]
서적
Developmental biology
Sinauer
[44]
논문
Cell lineages, developmental timing, and spatial pattern formation in embryos of free-living soil nematodes
https://pubmed.ncbi.[...]
1992-06-00
[45]
서적
Developmental biology
Sinauer
[46]
논문
Ascaroside signaling in C. elegans
2013-01-18
[47]
웹사이트
Introduction to C. Elegans
http://avery.rutgers[...]
Rutgers University
2014-08-15
[48]
웹사이트
Handbook - Introduction
http://www.wormatlas[...]
[49]
웹사이트
Dauer
http://www.wormbook.[...]
2018-09-27
[50]
논문
miRNAs give worms the time of their lives: small RNAs and temporal control in Caenorhabditis elegans
2010-05-00
[51]
서적
Developmental transitions in C. elegans larval stages
https://pubmed.ncbi.[...]
Academic Press
[52]
논문
The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14
1993-12-00
[53]
논문
Developmental timing in C. elegans is regulated by kin-20 and tim-1, homologs of core circadian clock genes
2005-02-00
[54]
논문
Post-embryonic cell lineages of the nematode, Caenorhabditis elegans
https://linkinghub.e[...]
1977-03-00
[55]
논문
The embryonic cell lineage of the nematode Caenorhabditis elegans
https://linkinghub.e[...]
1983-11-00
[56]
논문
Glia-derived neurons are required for sex-specific learning in C. elegans
2015-10-00
[57]
논문
Small RNAs couple embryonic developmental programs to gut microbes
http://biorxiv.org/l[...]
2020-11-14
[58]
서적
Invertebrate Zoology
Cengage Learning
[59]
논문
Bacterivorous nematodes decipher microbial iron siderophores as prey cue in predator–prey interactions
2024-01-16
[60]
논문
The natural history of Caenorhabditis elegans
2010-11-00
[61]
논문
Simultaneous Dissemination of Nanoplastics and Antibiotic Resistance by Nematode Couriers
https://pubs.acs.org[...]
2023-06-02
[62]
논문
Killing of Caenorhabditis elegans by Cryptococcus neoformans as a model of yeast pathogenesis
2002-11-00
[63]
논문
Caenorhabditis elegans as a model host for Staphylococcus aureus pathogenesis
2003-04-00
[64]
논문
Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis
1999-01-00
[65]
논문
Long-lived C. elegans daf-2 mutants are resistant to bacterial pathogens
2003-06-00
[66]
논문
Biofilm matrix disrupts nematode motility and predatory behavior
2021-01-00
[67]
논문
Biofilm matrix cloaks bacterial quorum sensing chemoattractants from predator detection
2022-05-00
[68]
논문
Ecology of Caenorhabditis species
2006-01-00
[69]
논문
An LEA group 3 family member is involved in survival of C. elegans during exposure to stress
2004-11-00
[70]
웹사이트
Soil biology primer
https://www.nrcs.usd[...]
USDA
[71]
논문
Genomic Analysis of ''Caenorhabditis elegans'' Reveals Ancient Families of Retroviral-like Elements
[72]
논문
Microsporidian genome analysis reveals evolutionary strategies for obligate intracellular growth
2012-12-01
[73]
논문
''Arthrobotrys oligospora'': A model organism for understanding the interaction between fungi and nematodes
[74]
논문
Voltage-gated sodium channels as therapeutic targets
2000-11-01
[75]
논문
Dynamics of the Model of the Caenorhabditis Elegans Neural Network
[76]
논문
Whole-animal connectomes of both ''Caenorhabditis elegans'' sexes
US National Library of Medicine, National Institutes of Health
2019-12-03
[77]
논문
Mapping the brain to understand the mind
https://knowablemaga[...]
2022-04-21
[78]
논문
Collective dynamics of 'small-world' networks
1998-06-01
[79]
논문
Deciphering the neural and molecular mechanisms of C. elegans behavior
2005-09-01
[80]
논문
Whole-animal connectomes of both Caenorhabditis elegans sexes
2019-07-01
[81]
논문
Caenorhabditis elegans respond to high-glucose diets through a network of stress-responsive transcription factors
2018-01-01
[82]
웹사이트
Sydney Brenner
http://elegans.swmed[...]
Southwestern Medical Center
[83]
논문
The genetics of Caenorhabditis elegans.
[84]
웹사이트
Behavior
http://www.wormbook.[...]
2018-09-26
[85]
논문
Post-embryonic cell lineages of the nematode, Caenorhabditis elegans
1977-03-01
[86]
논문
The postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans
1979-06-01
[87]
논문
Cell death specification in C. elegans
2008-08-01
[88]
Youtube
Injection of C. elegans Gonads
https://www.youtube.[...]
2015-03-05
[89]
논문
Systematic functional analysis of the Caenorhabditis elegans genome using RNAi
2003-01-01
[90]
논문
Uncover genetic interactions in Caenorhabditis elegans by RNA interference
2005-01-01
[91]
논문
RNA interference in nematodes and the chance that favored Sydney Brenner
2008-11-01
[92]
논문
Caenorhabditis elegans SID-2 is required for environmental RNA interference
2007-06-01
[93]
논문
Hyper-resistance of meiotic cells to radiation due to a strong expression of a single recA-like gene in Caenorhabditis elegans
2000-11-01
[94]
논문
Efficient repair of DNA damage induced by heavy ion particles in meiotic prophase I nuclei of Caenorhabditis elegans
2003-09-01
[95]
논문
C. elegans mre-11 is required for meiotic recombination and DNA repair but is dispensable for the meiotic G(2) DNA damage checkpoint
2001-03-01
[96]
논문
BRCA1/BRC-1 and SMC-5/6 regulate DNA repair pathway engagement during Caenorhabditis elegans meiosis
2024-08-01
[97]
논문
High local genetic diversity and low outcrossing rate in Caenorhabditis elegans natural populations
2005-07-01
[98]
논문
Evolutionary Origin of Recombination during Meiosis
2010-07-01
[99]
논문
A C. elegans model of nicotine-dependent behavior: regulation by TRP-family channels
2006-11-01
[100]
웹사이트
Alliance of Genome Resources Community Forum
https://community.al[...]
2024-08-01
[101]
논문
Regulation of C. elegans life-span by insulinlike signaling in the nervous system
2000-10-01
[102]
논문
Dauer-independent insulin/IGF-1-signalling implicates collagen remodelling in longevity
2015-03-01
[103]
논문
End-of-life targeted degradation of DAF-2 insulin/IGF-1 receptor promotes longevity free from growth-related pathologies
2021-09-10
[104]
논문
Longevity and resistance to stress correlate with DNA repair capacity in Caenorhabditis elegans
[105]
논문
Exosome-mediated delivery of superoxide dismutase for anti-aging studies in Caenorhabditis elegans
2023-06-01
[106]
논문
Decline of nucleotide excision repair capacity in aging Caenorhabditis elegans
[107]
논문
Pharmacogenetic analysis of lithium-induced delayed aging in Caenorhabditis elegans
2008-01-00
[108]
논문
Low-dose lithium uptake promotes longevity in humans and metazoans
2011-08-00
[109]
논문
Understanding the molecular basis of Alzheimer's disease using a Caenorhabditis elegans model system
2010-03-00
[110]
논문
PGL proteins self associate and bind RNPs to mediate germ granule assembly in C. elegans
2011-03-00
[111]
논문
The polygenic nature of telomere length and the anti-ageing properties of lithium
2019-03-00
[112]
논문
Uncoupling of longevity and telomere length in C. elegans
2005-09-00
[113]
논문
Telomere length: how the length makes a difference
2020-09-00
[114]
논문
Retrotransposons that maintain chromosome ends
2011-12-20
[115]
논문
trt-1 is the Caenorhabditis elegans catalytic subunit of telomerase
2006-02-00
[116]
논문
Alternative lengthening of telomeres: models, mechanisms and implications
https://pubmed.ncbi.[...]
2010-05-00
[117]
논문
Null allele mutants of trt-1, the catalytic subunit of telomerase in Caenorhabditis elegans, are less sensitive to Mn-induced toxicity and DAergic degeneration
https://pubmed.ncbi.[...]
2016-12-00
[118]
논문
Caenorhabditis elegans POT-1 and POT-2 repress telomere maintenance pathways
2013-02-00
[119]
논문
Paradoxical delay of senescence upon depletion of BRCA2 in telomerase-deficient worms
2016-10-00
[120]
논문
Effects of telomerase overexpression in the model organism Caenorhabditis elegans
https://pubmed.ncbi.[...]
2020-03-30
[121]
논문
The microarchitecture of C. elegans behavior during lethargus: homeostatic bout dynamics, a typical body posture, and regulation by a central neuron
2013-03-00
[122]
논문
Cellular stress induces a protective sleep-like state in C. elegans
2014-10-00
[123]
웹사이트
Teensy, Eyeless Worms Have Completely New Light-Detecting Cells
https://www.livescie[...]
[124]
간행물
Scientific American
2018-08-00
[125]
논문
In vivo quantitative high-throughput screening for drug discovery and comparative toxicology.
2023-00-00
[126]
논문
Caenorhabditis elegans for rare disease modeling and drug discovery: strategies and strengths.
2021-00-00
[127]
뉴스
Worms survived Columbia disaster
http://news.bbc.co.u[...]
2008-07-11
[128]
뉴스
University sends worms into space
http://news.bbc.co.u[...]
2009-07-09
[129]
뉴스
Legacy Space Worms Flying on Shuttle
https://web.archive.[...]
2011-05-17
[130]
논문
Microfluidics-integrated spaceflight hardware for measuring muscle strength of Caenorhabditis elegans on the International Space Station
2022-11-07
[131]
논문
Regulation of the X chromosomes in ''Caenorhabditis elegans''
2014-03-00
[132]
논문
Genome sequence of the nematode ''C. elegans'': a platform for investigating biology
1998-12-00
[133]
논문
Biochemistry and genetics of von Willebrand factor
Annual Reviews
[134]
논문
Aryl hydrocarbon receptors: diversity and evolution
Elsevier
[135]
논문
Regulatory and Signaling Properties of the Vav Family
American Society for Microbiology
[136]
논문
A global analysis of ''Caenorhabditis elegans'' operons
2002-06-00
[137]
논문
Operons in eukaryotes
2004-11-00
[138]
웹사이트
WS227 Release Letter
https://archive.toda[...]
WormBase
2013-11-19
[139]
논문
Large-scale sequencing reveals 21U-RNAs and additional microRNAs and endogenous siRNAs in ''C. elegans''
2006-12-00
[140]
논문
''C. elegans'' noncoding RNA genes
2005-06-00
[141]
웹사이트
WS202 Release Letter
https://archive.toda[...]
WormBase
2009-05-29
[142]
웹사이트
WS197 Release Letter
https://web.archive.[...]
WormBase
2008-11-27
[143]
웹사이트
Genome sequence changes
https://web.archive.[...]
WormBase
2011-06-15
[144]
논문
The genome sequence of ''Caenorhabditis briggsae'': a platform for comparative genomics
2003-11
[145]
웹사이트
''Caenorhabditis remanei'': Background
http://genome.wustl.[...]
Washington University School of Medicine
[146]
웹사이트
''Caenorhabditis japonica'': Background
http://genome.wustl.[...]
Washington University School of Medicine
[147]
논문
A strategy of DNA sequencing employing computer programs
1979-06
[148]
웹사이트
UCSC genome browser
http://genome.ucsc.e[...]
[149]
논문
The UCSC Genome Browser Database: update 2009
2009-01
[150]
논문
A streamlined system for species diagnosis in Caenorhabditis (Nematoda: Rhabditidae) with name designations for 15 distinct biological species
[151]
논문
Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans
http://www.dspace.ca[...]
1998-02
[152]
논문
Medicine Nobel awarded for gene-regulating 'microRNAs'
https://www.nature.c[...]
Nature Publishing Group
2024-10-07
[153]
논문
WormBase: a comprehensive resource for nematode research
2010-01
[154]
웹사이트
A 線虫とは何か
https://iss.jaxa.jp/[...]
[155]
서적
動物学の百科事典
丸善出版
2018-09-30
[156]
서적
動物学の百科事典
丸善出版
2018-09-30
[157]
간행물
寄生虫和名表
https://jsparasitol.[...]
日本寄生虫学会
1995
[158]
서적
生物学名命名法辞典
平凡社
1994-11-01
[159]
서적
国際動物命名規約 第4版 日本語版 [追補]
http://ujssb.org/icz[...]
日本分類学会連合
2005-10
[160]
서적
国際動物命名規約 第4版 日本語版 [追補]
http://ujssb.org/icz[...]
日本分類学会連合
2005-10
[161]
웹사이트
線虫に驚異的な加速耐性
https://www.nikkei-s[...]
日経サイエンス
[162]
보도자료
環境の酸素濃度が体の低温適応を調節する―複数の感覚情報を統合する神経の仕組みの理解に繋がる―
https://www.amed.go.[...]
国立研究開発法人日本医療研究開発機構(AMED)
2019-02-07
[163]
보도자료
低温環境に弱い線虫が氷点下で生き延びた!
https://www.aist.go.[...]
国立研究開発法人産業技術総合研究所
2019-05-15
[164]
보도자료
気温上昇により線虫に多世代遺伝子変異が生じる
https://www.eurekale[...]
アメリカ科学振興協会
2017-04-20
[165]
보도자료
ClCチャネルによる味覚応答の調節
https://www.s.u-toky[...]
東京大学大学院理学系研究科・理学部 広報室
2021-01-27
[166]
서적
無脊椎動物の多様性と系統
裳華房
2000-11-30
[167]
서적
生物の種多様性
裳華房
1997-09-20
[168]
서적
はじめに線虫ありき そして、ゲノム研究が始まった
青土社
2006-02-20
[169]
웹사이트
ライフサイエンス実験 Space Aging
https://iss.jaxa.jp/[...]
2015-04-13
[170]
서적
線虫学実験
京都大学学術出版会
2014-10-01
[171]
논문
自活性土壌線虫の分離と措養および遺伝学の試み
2005
[172]
서적
多様性からみた生物学
裳華房
2008
[173]
논문
昆虫の低分子量GTP結合蛋白質(Rab)
https://www.jstage.j[...]
2014
[174]
서적
動物の系統分類と進化
裳華房
2010-04-28
[175]
논문
Post-embryonic cell lineages of the nematode, Caenorhabditis elegans
https://linkinghub.e[...]
1977-03
[176]
저널
The embryonic cell lineage of the nematode Caenorhabditis elegans
https://linkinghub.e[...]
1983-11
[177]
저널
Glia-derived neurons are required for sex-specific learning in C. elegans
http://www.nature.co[...]
2015-10
[178]
저널
Small RNAs couple embryonic developmental programs to gut microbes
http://biorxiv.org/l[...]
2020-11-14
[179]
저널
Neural regulation of thermotaxis in Caenorhabditis elegans
http://www.nature.co[...]
1995-07
[180]
저널
Plasticity of chemotaxis revealed by paired presentation of a chemoattractant and starvation in the nematode Caenorhabditis elegans
https://jeb.biologis[...]
2001-05-15
[181]
저널
Odorant-specific adaptation pathways generate olfactory plasticity in C. elegans
https://linkinghub.e[...]
1995-04
[182]
저널
Somatic sexual differentiation in ''Caenorhabditis elegans''
2008
[183]
서적
動物学の百科事典
丸善出版
2018-09-30
[184]
웹사이트
Caenorhabditis Genetics Center
http://biosci.umn.ed[...]
[185]
저널
Green fluorescent protein as a marker for gene expression
https://www.sciencem[...]
1994-02-11
[186]
뉴스
九大プレスリリース20150312
九州大学プレスリリース
2015-03-12
[187]
저널
Modes et formes de reproduction des nématodes
[188]
서적
벌레의 마음
[189]
웹사이트
Introduction
2009
[190]
웹사이트
The cuticle
http://www.wormbook.[...]
2007-03-19
[191]
저널
Cuticle of Caenorhabditis elegans: its isolation and partial characterization
1981
[192]
서적
THE CELL: A Molecular Approach Fifth Edition
ASM PRESS
[193]
저널
Operons in eukaryotes
2004
[194]
웹사이트
Dauer
http://www.wormbook.[...]
2007-08-08
[195]
저널
The natural history of Caenorhabditis elegans
http://dx.doi.org/10[...]
2010
[196]
웹사이트
Ecology of Caenorhabditis species
http://www.wormbook.[...]
2006-01-09
[197]
저널
How cellular slime molds evade nematodes
http://www.pnas.org/[...]
1996
[198]
웹사이트
C.elegans, 환상적인 생명공학 모델 동물
http://www.kormb.or.[...]
2012
[199]
저널
The Genetics of CAENORHABDITIS ELEGANS
1974-05
[200]
서적
Background on Caenorhabditis elegans
Oxford Universit Press, NY
1999
[201]
저널
Caenorhabditis elegans: an emerging model in biomedical and environmental toxicology
2008
[202]
저널
Genetic control of programmed cell death in the nematode C.elegans
1986
[203]
서적
THE CELL: A Molecular Approach Fifth Edition
ASM PRESS
[204]
저널
Finding fuction in novel targets: C.elegans as a model organism
2006
[205]
저널
Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C.elegans insulin gene family
[206]
저널
Aging and resistance to oxidative damage in Caenorhabditis elegans
1993
[207]
저널
Increased frequency of deletions in the mitochondrial genome with age of Caenorhabdtis elegans
1995
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com