총채벌레목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
총채벌레목(Thysanoptera)은 몸길이가 약 1mm 내외의 작은 곤충으로, 독특한 형태와 생활사를 지닌다. 전 세계적으로 분포하며, 식물 조직을 갉아먹거나 식물 질병을 매개하여 농작물에 해를 입히는 해충으로 알려져 있다.
총채벌레는 두 개의 아목(Terebrantia, Tubulifera)으로 나뉘며, 불완전변태를 거친다. 일부 종은 사회적 행동을 보이며, 다른 곤충이나 식물에 혹을 형성하기도 한다. 총채벌레는 살충제에 대한 저항성을 쉽게 획득하여 방제가 어렵지만, 생물학적 방제 연구가 진행 중이다.
더 읽어볼만한 페이지
- 총채벌레목 - 총채벌레아목
총채벌레아목은 곤충강 총채벌레목에 속하는 아목으로, 작고 가느다란 몸을 가진 곤충들로 구성된다.
| 총채벌레목 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Thysanoptera Haliday, 1836 |
| 이전 학명 | Physopoda |
| 분류 | |
| 계 | 동물계 |
| 문 | 절지동물문 |
| 강 | 곤충강 |
| 아강 | 유시아강 |
| 하강 | 신시하강 |
| 목 | 총채벌레목 (Thysanoptera) |
| 아목 및 과 | |
| 穿孔亜目 (Terebrantia) | Adiheterothripidae 시마아자미우마과 (Aeolothripidae) Fauriellidae † Hemithripidae Heterothripidae † Jezzinothripidae † Karataothripidae Melanthripidae 메로아자미우마과 (Merothripidae) † Scudderothripidae Stenurothripidae 아자미우마과 (Thripidae) † Triassothripidae Uzelothripidae |
| 有管亜目 (Tubulifera) | クダアザミウマ科 (Phlaeothripidae) † Rohrthripidae |
| 형태 및 생태 | |
| 크기 | 약 1mm |
| 화석 기록 | 페름기 – 현재 |
2. 형태
총채벌레라는 영어 이름은 고대 그리스어 θρίψ|트립스grc에서 직접 번역된 것으로, "좀"을 의미한다.[4] 'sheep', 'deer', 'moose'와 같이 영어에서 단수와 복수 형태가 같은 다른 동물 이름처럼, "thrips"라는 단어는 단수와 복수 모두를 나타내므로, 여러 마리의 총채벌레(thrips) 또는 단일 총채벌레(thrips)를 의미할 수 있다. 총채벌레의 다른 일반적인 이름으로는 thunderflies, thunderbugs, storm flies, thunderblights, storm bugs, corn fleas, corn flies, corn lice, freckle bugs, harvest bugs, physopods 등이 있다.[5][6][7] 더 오래된 그룹 이름인 "physopoda"는 다리 부절의 방광 모양 끝 부분을 가리킨다. 총채벌레목의 학명 ''Thysanoptera''는 고대 그리스어 θύσανος|티사노스grc ("술 또는 가장자리")와 πτερόν|프테론grc ("날개")에서 유래되었으며, 곤충 날개 가장자리의 술 장식을 지칭한다.[8][9][10]
총채벌레는 중생대에 곰팡이를 먹는 조상으로부터 유래한 것으로 보이며,[29] 많은 종류가 여전히 곰팡이 포자를 먹고 의도치 않게 재분배한다.[30] 이들은 낙엽이나 죽은 나무에서 살며 생태계의 중요한 구성원이며, 종종 꽃가루를 먹는다. 다른 종들은 원시적인 사회성 곤충이며 식물 충영을 형성하고, 또 다른 종들은 응애와 다른 총채벌레를 포식한다.[9][31] 브라질에서 ''Aulacothrips''의 두 종(''A. tenuis''와 ''A. levinotus'')이 에탈리온과 및 매미충과에 외부 기생충으로 발견되었다.[32] 오스트레일리아의 ''Akainothrips francisi''는 아카시아 나무에 실크 둥지나 굴을 만드는 다른 총채벌레 종인 ''Dunatothrips aneurae''의 군집 내 기생충이다.[33] Phlaeothripinae 아과에 속하는 많은 총채벌레는 아카시아를 숙주로 하며, 잎을 서로 접착하여 반사회적 군집이 서식하는 굴을 형성하는 실크를 생산한다.[34]
총채벌레는 독특한 시가 모양의 몸 구조를 가진 작고 불완전변태 곤충이다.[16] 몸은 길쭉하며 가로로 수축되어 있다. 대부분의 총채벌레는 길이가 약 1 mm이지만, 포식성 총채벌레의 경우 0.5mm에서 14mm 사이이다. 비행 가능한 총채벌레는 털이 달린 가장자리가 있는 두 쌍의 유사한 끈 모양의 날개를 가지고 있으며, 날개는 휴식 시 몸 위로 접혀진다. 다리는 일반적으로 족사가 2개인 마디로 끝나며, 발톱 앞부분에는 "arolium"이라고 하는 방광 모양의 구조가 있다. 이 구조는 혈림프 압력에 의해 뒤집힐 수 있어 곤충이 수직 표면을 걸을 수 있게 해준다.[11][12] 겹눈은 소수의 겹눈과 머리에 세 개의 홑눈 또는 단순한 눈으로 구성되어 있다.[62]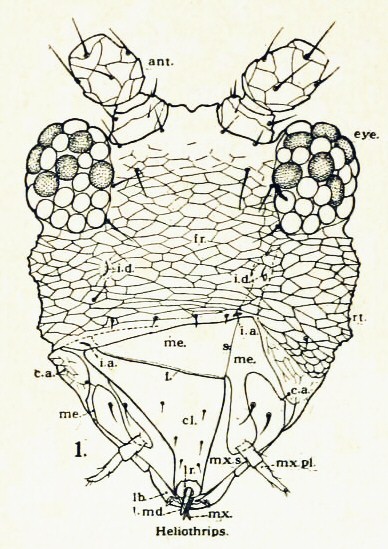
총채벌레는 이 그룹에 고유한 비대칭적인 구기를 가지고 있다. 노린재(진딧물)와 달리 총채벌레의 오른쪽 턱은 줄어들고 흔적 기관이며, 일부 종에서는 완전히 사라졌다.[13] 왼쪽 턱은 잠시 먹이 식물에 자르는데 사용되며, 침이 주입되고, 관을 형성하는 위턱 스타일렛이 삽입된 다음 반소화된 음식이 파열된 세포에서 펌핑된다. 이 과정은 세포를 파괴하거나 붕괴시키며, 총채벌레가 먹었던 줄기나 잎 표면에 독특한 은색 또는 청동색 흉터를 남긴다.[14] 총채벌레의 구기는 "긁어 먹는-빨아 먹는",[15] "구멍을 내서 빨아 먹는",[16] 또는 단순히 "뚫어 빨아 먹는" 구기의 특정 유형으로 묘사되어 왔다.[17]
총채벌레목(Thysanoptera)은 Terebrantia와 Tubulifera의 두 아목으로 나뉜다. 이들은 형태학적, 행동적, 발달적 특성으로 구분할 수 있다. 관총상목(Tubulifera)은 단일 과인 애총채벌레과(Phlaeothripidae)로 구성된다. 구성원은 특징적인 관 모양의 정단 복부 분절, 잎 표면 위에 알을 낳는 것, 세 개의 "번데기" 단계를 통해 식별할 수 있다. 애총채벌레과에서 수컷은 암컷보다 크고, 개체군 내에서 다양한 크기를 찾을 수 있다. 기록된 가장 큰 애총채벌레과의 종은 약 14mm이다. Terebrantia의 8개 과의 암컷은 모두 이름을 딴 톱 모양(참조 terebra)의 산란관을 정단 복부 분절에 가지고 있으며, 식물 조직 내에 알을 하나씩 낳고, 두 개의 "번데기" 단계를 거친다. 대부분의 Terebrantia에서 수컷은 암컷보다 작다. Uzelothripidae 과는 단일 종을 가지며 채찍 모양의 말단 안테나 분절을 갖는다는 점에서 독특하다.[62]
총채벌레는 가늘고 긴 몸을 가진 곤충이다. 머리 부분도 긴 것이 많아, 말의 이름은 여기서 연상된 것으로 보인다. 다리는 짧고, 운동은 그다지 활발하지 않다. 날개는 일반 곤충처럼 넓고 얇은 막이 아니라, 막대 모양의 주변에 온통 긴 털이 난 것이다. 총채벌레목의 옛 이름인 '''총시목'''은 이것에서 유래한다. 일반적으로 몸길이는 1mm 이하 정도이며, 비행 능력은 부족하지만 바람을 타고 멀리 이동한다. 입틀은 좌우 비대칭 부품이 원뿔 모양으로 조합되어 흡즙용으로 특수화되어 있으며, 모두 구멍을 뚫어 내용물을 빨아들이는 섭식법을 행하지만, 가까운 노린재목처럼 긴 입술을 가지고 있지 않기 때문에, 먹이 생물의 몸 깊숙한 곳에서 음식을 얻을 수는 없다. 식물 식자의 경우 표피에 구멍을 뚫어 그 아래 부드러운 조직의 세포 내용물을 빨아들인다. 꽃가루나 균류의 포자의 세포벽에 구멍을 뚫어, 내용물을 빨아들이는 종도 많다.
3. 어원
4. 생태
브라질에서 ''Mirothrips arbiter''는 종이말벌 둥지에서 발견되었으며, ''Mischocyttarus atramentarius'', ''Mischocyttarus cassununga'', ''Polistes versicolor''를 포함한 숙주의 알을 먹는다.[35] 특히 줄총채벌레과에 속하는 총채벌레는 포식자이기도 하며 사과굴나방과 같은 해충을 관리하는 데 유익한 것으로 간주된다.[36]
대부분의 연구는 경제적으로 중요한 작물을 먹는 총채벌레 종에 초점을 맞추고 있다. 일부 종은 포식성이지만, 대부분은 꽃가루와 식물 표피 및 엽육 세포의 외부 층에서 수확한 엽록체를 먹는다. 그들은 싹, 꽃 및 새 잎과 같은 식물의 연한 부분을 선호한다.[37][38] 꽃노랑총채벌레는 식물 조직을 먹는 것 외에도 꽃가루와 응애의 알을 먹는다. 애벌레가 이러한 방식으로 식단을 보충하면 발육 시간과 사망률이 감소하며, 응애 알을 섭취하는 성체 암컷은 번식력과 수명이 증가한다.[39]
꽃을 먹는 일부 총채벌레는 먹고 있는 꽃의 수분을 돕는다.[40] 일부 저자는 총채벌레가 숙주 식물과의 수분 관계를 진화시킨 최초의 곤충 중 하나였을 가능성이 있다고 추측한다.[41] 백악기 전기의 ''Gymnopollisthrips'' 화석은 ''Cycadopites''와 같은 꽃가루로 덮여 있는 것을 보여준다.[42] ''Scirtothrips dorsalis''는 상업적으로 중요한 고추의 꽃가루를 옮긴다.[43][44][45] 다윈은 더 큰 수분 매개체를 격리하는 실험을 수행했을 때 어떤 그물망으로도 총채벌레를 막을 수 없다는 것을 발견했다.[46] ''Thrips setipennis''는 호주 동부 열대 우림에서 자라는 작고 단성화의 일년생 나무 또는 관목인 ''Wilkiea huegeliana''의 유일한 수분 매개체이다. ''T. setipennis''는 ''Myrsine howittiana'' 및 ''M. variabilis''를 포함한 다른 호주 열대 우림 식물 종에 대한 필수적인 수분 매개체 역할을 한다.[47] 
총채벌레는 먹이를 먹는 동안 피해를 입힐 수 있다.[50] 이러한 피해는 숙주 선호도에 상당한 폭이 있고, 심지어 종 내에서도 숙주에 대한 충실도에 따라 다르기 때문에 다양한 먹이 항목에 걸쳐 나타날 수 있다.[37][51] 특히 총채벌레과(Thripidae)는 광범위한 숙주 범위를 가진 구성원들로 악명이 높으며, 해충 총채벌레의 대부분이 이 과에서 나온다.[52][53][54] 예를 들어, ''담배총채벌레(Thrips tabaci)''는 양파, 감자, 담배, 목화 작물에 피해를 준다.[38][55]
일부 총채벌레 종은 잎 조직에서 거의 항상 혹을 생성한다. 이것들은 잎이 말리거나, 굴러가거나 접히는 형태로 나타날 수 있으며, 조직의 확장을 변화시켜 잎의 날을 뒤틀리게 한다. 더 복잡한 예로는 로제트, 주머니, 뿔을 생성한다. 이러한 종의 대부분은 열대 및 아열대 지역에서 발생하며, 혹의 구조는 관련된 종을 진단하는 데 사용된다.[56] 호주에서는 ''아카시아(Acacia)'' 나무에서 총채벌레 종의 방사가 일어난 것으로 보인다. 이들 중 일부 종은 잎자루(petiole)에 혹을 생성하여 때로는 두 개의 잎자루를 함께 고정하는 반면, 다른 종은 나무 껍질의 모든 틈새에 산다. 같은 나라의 ''카수아리나(Casuarina)''에서는 일부 종이 줄기에 침입하여 오래 지속되는 목질 혹을 생성한다.[57] 
총채벌레는 가늘고 긴 몸을 가진 곤충이다. 머리 부분도 긴 것이 많아, 말의 이름은 여기서 연상된 것으로 보인다. 다리는 짧고, 운동은 그다지 활발하지 않다. 날개는 일반 곤충처럼 넓고 얇은 막이 아니라, 막대 모양의 주변에, 온통 긴 털이 난 것이다. 총채벌레목의 옛 이름인 '''총시목'''은 이것에서 유래한다.
일반적으로 몸길이는 1mm 이하 정도이며, 비행 능력은 부족하지만 바람을 타고 멀리 이동한다. 입틀은 좌우 비대칭 부품이 원뿔 모양으로 조합되어 흡즙용으로 특수화되어 있으며, 모두 구멍을 뚫어 내용물을 빨아들이는 섭식법을 행하지만, 가까운 노린재목처럼 긴 입술을 가지고 있지 않기 때문에, 먹이 생물의 몸 깊숙한 곳에서 음식을 얻을 수는 없다. 식물을 먹는 종의 경우 표피에 구멍을 뚫어 그 아래 부드러운 조직의 세포 내용물을 빨아들인다. 꽃가루나 균류의 포자의 세포벽에 구멍을 뚫어, 내용물을 빨아들이는 종도 많다.
일본어 이름은 "말이 나와라" 등이라고 말하면서, 엉겅퀴 꽃을 흔들어, 나온 꽃가루를 먹는 총채벌레를 세는 옛날 어린이들의 놀이에서 유래한다.
불완전변태를 하며, 유충은 성충과 비슷한 모습을 하고 있다. 단, 성충이 되기 전에 섭식하지 않고 행동도 둔해지는 용이라고 불리는 시기가 1기 또는 2기 있다. 이 시기의 형태는 유충과 큰 차이가 없으며, 성충의 형태도 유충에 단순히 날개를 붙인 듯한 모습이지만, 완전변태 곤충과 마찬가지로 대식세포에 의한 유충 기관의 많은 파괴와 소수의 세포에 의한 성충 기관의 신생이 일어나고 있는 것이 확인되었다.
총채벌레는 수정 없이 발생하면 수컷이 되고, 수정하면 암컷이 된다.
4. 1. 사회 행동
화학적 의사소통은 잘 알려져 있지는 않지만, 집단에게 중요하다고 여겨진다.[58] 항문 분비물은 뒷창자에서 생성되며,[59] 포식자 방어물로서 후방 강모를 따라 방출된다.[59][60] 오스트레일리아에서는 수컷 꽃노랑총채벌레의 집단이 ''중국 부용''과 ''목화''의 꽃잎에서 관찰되었으며, 암컷은 이 집단에 유인되었으므로 수컷이 페로몬을 생성했을 가능성이 높다.[61]
균류를 먹는 굴뚝총채벌레과에서 수컷은 암컷을 보호하고 교미하기 위해 경쟁하며, 알 덩어리를 방어한다. 수컷은 복부로 상대를 쳐내며 싸우고, 앞다리 발톱으로 죽일 수도 있다. 작은 수컷은 큰 수컷이 싸우는 동안 몰래 들어가 교미할 수 있다. 참총채벌레과와 애꽃노랑총채벌레과에서도 수컷은 크고 작은 형태로 다형성을 보이며, 아마도 짝을 얻기 위해 경쟁할 것이므로, 이 전략은 총채벌레목에서 조상 대대로 내려온 것일 수 있다.[62]
많은 총채벌레는 먹이를 먹거나 알을 낳을 때 식물에 혹을 형성한다. Kladothrips속[63]과 Oncothrips속[64]과 같은 혹을 형성하는 일부 굴뚝총채벌레과는 번식하는 여왕개미와 비번식 병정 계급을 가진 개미 군집과 유사한 진사회성 집단을 형성한다.[65][66][67]
총채벌레는 수정 없이 발생하면 수컷이 되고, 수정하면 암컷이 된다는 벌목과 유사한 성질을 가지고 있다. 1980년대에 벌류의 사회성 발달과 이러한 성 결정 방식에 깊은 관계가 있다고 여겨졌다. 그때 다른 곤충에서도 유사한 사례가 없는지 탐색이 이루어졌고, 그중 하나가 총채벌레류이다. 위의 성 결정 방식을 가지고 있으며, 자주 집중하여 생활하는 것이 알려져 있었기 때문에, 여기서 새로운 진사회성의 종이 발견될 것이라고 생각되었고, 그 후 오스트레일리아에서 약간의 사회성 종이 발견되었다.
5. 생애 주기
총채벌레는 길이가 약 0.2mm인 극도로 작은 알을 낳는다. Terebrantia 아목의 암컷은 산란관으로 식물 조직에 틈을 내어 알을 하나씩 삽입한다. Tubulifera 아목의 암컷은 알을 식물 바깥 표면에 하나씩 또는 소규모로 낳는다.[74] ''Elaphothrips tuberculatus''와 같은 일부 총채벌레는 선택적으로 난태생으로, 알을 내부에서 유지하고 수컷 새끼를 낳는 것으로 알려져 있다.[75] 많은 종의 암컷은 다른 암컷의 식충 및 포식자로부터 알을 보호한다.[76]
총채벌레는 불완전변태를 하며, 성충 형태로 점진적으로 변태한다. 유충 또는 약충이라고 불리는 처음 두 령은 생식기가 없는 작은 무익 성충과 같으며(종종 톡토기로 오인됨), 이들은 식물 조직을 먹는다. Terebrantia에서는 세 번째와 네 번째 령, Tubulifera에서는 다섯 번째 령도 있으며, 먹이를 먹지 않고 휴식을 취하는 단계는 번데기와 유사하다. 이 단계에서 신체 기관이 재형성되고, 날개 싹과 생식기가 형성된다.[74] 일부 종의 유충은 마지막 복부 마디에서 실을 생산하여 세포를 만들거나 번데기가 되는 고치를 형성하는 데 사용한다.[77] 성충 단계는 약 8~15일 안에 도달할 수 있으며, 성충은 약 45일 동안 살 수 있다.[87] 성충은 날개가 있는 형태와 날개가 없는 형태를 모두 가지고 있다. 예를 들어, 풀 총채벌레인 ''Anaphothrips obscurus''의 경우, 날개가 있는 형태는 봄(온대 지역)에 개체수의 90%를 차지하는 반면, 날개가 없는 형태는 늦여름에 개체수의 98%를 차지한다.[78] 총채벌레는 성충으로, 알 또는 번데기 휴면을 통해 겨울을 날 수 있다.[62]
총채벌레는 반수배수성으로, 무수정란에서 유래하는 수컷(벌목과 같이)과 단성생식이 가능한 이배체 암컷이 있으며, 많은 종이 수컷생식을 사용하고, 일부는 암컷생식을 사용한다.[79] ''Pezothrips kellyanus''에서는 암컷이 수컷보다 더 큰 알에서 부화하는데, 이는 수정될 가능성이 더 높기 때문일 수 있다.[80] 성을 결정하는 세균 공생체인 ''Wolbachia''는 생식 방식에 영향을 미치는 요인이다.[51][79][81] 여러 개의 정상적인 양성 종이 미국에서 암컷만 존재하는 상태로 정착되었다.[79][82]
불완전변태를 하며, 유충은 성충과 비슷한 모습을 하고 있다. 단, 성충이 되기 전에 섭식하지 않고 행동도 둔해지는 용이라고 불리는 시기가 1기 또는 2기 있다. 이 시기의 형태는 유충과 큰 차이가 없으며, 성충의 형태도 유충에 단순히 날개를 붙인 듯한 모습이지만, 완전변태 곤충과 마찬가지로 대식세포에 의한 유충 기관의 많은 파괴와 소수의 세포에 의한 성충 기관의 신생이 일어나고 있는 것이 확인되었다.
아자미무시(총채벌레)는 수정 없이 발생하면 수컷이 되고, 수정하면 암컷이 된다는 벌목과 유사한 성질을 가지고 있다. 진사회성의 사고방식이 주목받았던 1980년대에 벌류의 사회성 발달과 이러한 성 결정 방식에 깊은 관계가 있다고 여겨졌다. 그 때, 다른 곤충에서도 유사한 사례가 없는지 탐색이 이루어졌고, 주목받은 것 중 하나가 아자미무시류이다. 위의 성 결정 방식을 가지고 있으며, 자주 집중하여 생활하는 것이 알려져 있었기 때문에, 여기서 새로운 진사회성의 종이 발견될 것이라고 생각되었고, 그 후 오스트레일리아에서 약간의 사회성 종이 발견되었다.
6. 분류
2006년부터 총채벌레목은 크게 총채벌레아목(Terebrantia)과 관총채벌레아목(Tubulifera)의 두 아목으로 나뉜다.[25][26][62]
관총채벌레아목 (Tubulifera)은 관총채벌레과 (Phlaeothripidae) 한 과만을 포함하며, 2개의 아목, 447속으로 구성되어 있고, 곰팡이 균사와 포자를 먹는 것으로 알려져 있다.
- '''관총채벌레아목'''(Tubulifera)
- * 관총채벌레과 (Phlaeothripidae) - 2 아목, 447속
관총채벌레과에는 다음과 같은 속들이 있다.
- ''Bactrothrips'' - 밤나무관총채벌레(''B. brevitubus'')
- ''Ophthalmothrips'' - 억새관총채벌레(''O. miscanthicola'')
- ''Bagnalliella'' - 유카관총채벌레(''B. yuccae'')
- ''Haplothrips'' - 벼관총채벌레(''H. aculeatus''), 중국관총채벌레(''H. chinensis''), 무궁화관총채벌레(''H. kurdjumovi''), 토끼풀관총채벌레(''H. niger'')
- ''Liothrips'' - 콩관총채벌레(''L. glycinicola''), 돌나물관총채벌레(''L. piperinus''), 백합관총채벌레(''L. vaneeckei'')
- ''Pentagonothrips'' - 바둑총채벌레(''P. antennalis'')
- ''Podothrips'' - 대총채벌레(''P. sasacola'')
- ''Stephanothrips'' - 꼬리맵시총채벌레(''S. japonicus'')
- ''Xylaplothrips'' - 애백합총채벌레(''X. subterraneu'')
- 아카메가시와쿠다총채벌레
과거에는 "생명의 나무 프로젝트"에서 제안된 계통 분류도 있었으나,[100] 현재는 일반적으로 사용되지 않는다.
6. 1. 총채벌레아목 (Terebrantia)

2006년부터 인정되고 있는 총채벌레목의 하위 과는 다음과 같다.[25][26][62]
총채벌레 종 식별은 시간이 지남에 따라 품질이 다양한 슬라이드 표본으로 보존되기 때문에 어렵다. 또한, 많은 종이 오인될 정도로 상당한 변동성을 보인다. 분자 서열 기반 접근 방식이 점차 식별에 많이 적용되고 있다.[27][28]
총채벌레 (천공) 아목 Terebrantia에는 다음과 같은 과가 있다.
- Adiheterothripidae 과
- 시마총채벌레과 - 알리가타시마총채벌레
- Fauriellidae 과
- Heterothripidae 과
- Melanthripidae 과
- 메로총채벌레과 - 스베스베메로총채벌레
- 총채벌레과 - 미캉키이로총채벌레, 미나미키이로총채벌레, 네기총채벌레, 히라즈하나총채벌레
- Uzelothripidae 과
6. 2. 관총채벌레아목 (Tubulifera)
관총채벌레과는 2개의 아목, 447속으로 구성되어 있으며, 곰팡이 균사와 포자를 먹는다.[25][26][62]7. 계통 분류
다음은 2002년 "생명의 나무 프로젝트"(''The Tree of Life Web Project'')에 제안된 계통 분류이다.[100]
{| class="wikitable"
|-
! 신시류
|-
|
{| class="wikitable"
|-
|
{| class="wikitable"
|-
| 강도래목
|-
| 집게벌레목
|-
| 민벌레목
|-
! 망시상목
|-
|
|-
! 무시귀뚜라미붙이상목
|-
|
| 대벌레붙이목 |
| 귀뚜라미붙이목 |
|-
|
|-
! 메뚜기목
|-
|
|}
|-
|
{| class="wikitable"
|-
|
|-
| 내시상목
|}
|}
총채벌레는 일반적으로 노린재목(벌레)의 자매군으로 여겨진다.[23] 총채벌레과의 계통 발생은 거의 연구되지 않았다. 2013년에 3개의 유전자를 사용하여 37종을 예비 분석한 결과와 2012년의 리보솜 DNA와 3개의 단백질을 기반으로 한 계통 발생은 두 아목인 관총채아목과 숨총채아목의 단일 계통군을 지지한다. 숨총채아목에서 검정총채벌레과는 다른 모든 과의 자매군일 수 있지만, 다른 관계는 불분명하게 남아있다. 관총채아목에서 혹총채벌레과와 그 아과인 우상총채아과는 단일 계통군이다. 가장 큰 두 개의 총채벌레 아과인 혹총채벌레아과와 꽃노랑총채벌레아과는 측계통군이며, 그 구조를 결정하기 위한 추가 작업이 필요하다. 이러한 분석의 내부 관계는 아래와 같다.[24][25]
{| class="wikitable"
|-
! '''총채벌레목'''
|-
|
{| class="wikitable"
|-
! 숨총채아목
|-
|
| 검정총채벌레과 |
| 다른 숨총채아목 |
|-
! 관총채아목
|-
| 혹총채벌레과
|}
|}
8. 인간과의 관계

총채벌레는 식물을 가해하여 농작물에 피해를 주는 농업 해충으로 간주된다. 특히 온실 재배 작물에서 큰 피해를 입히고 있다.
하지만, 애리가타시마아자미우마는 미나미키이로아자미우마, 나미하드다니, 은잎가루이, 콩굴파리 등을 포식하기 때문에 생물 농약으로 활용되기도 한다.[94] 일본에서는 2003년 오이와 가지의 농약으로 등록되었으나,[95] 계절이나 온도 조건에 따라 효과가 제한적일 수 있다.[96]
8. 1. 농업 해충
많은 총채벌레는 발달 중인 꽃이나 채소를 갉아먹어 작물의 변색, 기형, 시장성 감소를 유발하여 상업 작물의 해충이 된다. 일부 총채벌레는 토스포바이러스와 같은 식물 질병의 매개체 역할을 한다.[83] 20가지가 넘는 식물 감염 바이러스가 총채벌레에 의해 전파되는 것으로 알려져 있지만, 반대로, 기술된 종 중 12종 미만이 토스포바이러스를 매개하는 것으로 알려져 있다.[84] 이 바이러스는 전 세계적으로 가장 치명적인 식물 병원체 중 하나로 간주되며, 해당 매개체 종은 인류의 농업에 막대한 영향을 미친다. 바이러스 구성원에는 토마토 반점 위조 바이러스와 봉선화 괴사 반점 바이러스가 포함된다. 꽃노랑총채벌레( ''Frankliniella occidentalis'')는 현재 전 세계적으로 분포할 정도로 확산되었으며, 토스포바이러스에 의해 유발되는 식물 질병의 주요 매개체이다.[85] 그들이 전파하는 다른 바이러스에는 ''Ilarvirus'', ''(Alpha'', ''Sobemovirus'' 및 ''Machlomovirus'' 속이 있다.[86]총채벌레는 작은 크기와 밀폐된 장소를 선호하는 성향 때문에 식물 검역으로 탐지하기 어렵고, 식물 조직 내에 알을 낳아 살충제 살포로부터 잘 보호된다.[87] 세계화되는 무역과 온실 농업의 성장이 결합됨에 따라, 총채벌레는 세계에서 가장 빠르게 증가하는 침입종 그룹에 속한다. 예로는 꽃노랑총채벌레(''F. occidentalis''), 글라디오러스총채벌레(''Thrips simplex''), ''Thrips palmi'' 등이 있다.[88]
꽃을 먹는 총채벌레는 밝은 꽃 색깔(흰색, 파란색, 특히 노란색 포함)에 정기적으로 유인되어 착륙하여 먹이를 먹으려고 한다. 일부 종(예: ''Frankliniella tritici'' 및 ''Limothrips cerealium'')이 그러한 상황에서 인간을 무는 것은 드문 일이 아니다. 어떤 종도 피를 먹지 않고 총채벌레에 의해 전파되는 동물 질병은 알려져 있지 않지만, 약간의 피부 자극은 보고된 바 있다.[89]
식물을 가해하는 종이 눈에 띄며, 농작물을 가해하는 종은 농업 해충으로 간주된다. 이들은 조건이 맞으면 종종 대 발생하며, 특히 최근에는 온실 재배 작물에서 중요한 해충이 되고 있다.
애리가타시마아자미우마는 미나미키이로아자미우마, 나미하드다니, 은잎가루이 및 콩굴파리를 포식하기 때문에 생물 농약으로의 응용이 연구되었고,[94] 일본에서는 2003년 오이와 가지의 농약으로 등록되었지만,[95] 계절이나 온도 조건의 제약도 있다.[96]
8. 2. 생물적 방제

총채벌레는 살충제에 대한 살충제 저항성을 쉽게 키우며, 이를 통제하는 방법에 대한 연구가 끊임없이 진행되고 있다. 이러한 특성 때문에 총채벌레는 새로운 살충제와 방법의 효과를 시험하는 데 이상적인 모델이 된다.[90]
총채벌레는 크기가 작고 번식률이 높아 고전적인 생물적 방제를 통해 제어하기 어렵다. 적합한 포식자는 총채벌레가 먹이를 찾기 위해 숨는 틈새를 파고들 수 있을 만큼 작고 가늘어야 하며, 효과를 보기 위해서는 알과 유충을 광범위하게 포식해야 한다. 기생성(parasitoid) 벌인 Eulophidae와 Trichogrammatidae 두 과만이 알과 유충에 기생한다. 성충과 유충의 다른 생물학적 방제제로는 ''Orius''속의 애꽃노린재와 phytoseiid 진드기가 있다. ''Beauveria bassiana''와 ''Verticillium lecanii''와 같은 생물 살충제는 총채벌레의 모든 생애 주기 단계에서 총채벌레를 죽일 수 있다.[91] 살충 비누 스프레이는 총채벌레에 효과적이다. 상업적으로 판매되거나 특정 종류의 가정용 비누로 만들 수 있다. 일본 과학자들은 식물에 적색광을 조사하면 멜론총채벌레의 유충과 성충이 현저하게 감소한다고 보고했다.[92]
애리가타시마아자미우마는 미나미키이로아자미우마, 나미하드다니, 은잎가루이 및 콩굴파리를 포식하기 때문에 생물 농약으로의 응용이 연구되었고[94], 일본에서는 2003년 오이와 가지의 농약으로 등록되었지만[95], 계절이나 온도 조건의 제약도 있다.[96]
참조
[1]
논문
Heinrich Uzel, the father of Thysanoptera studies
http://www.mapress.c[...]
[2]
서적
Monografie řádu Thysanoptera
https://archive.org/[...]
Hradec Králové
[3]
논문
Heinrich Uzel, the father of Thysanoptera studies
http://www.mapress.c[...]
[4]
LSJ
θρίψ
[5]
논문
Checklist of Nordic Thysanoptera
http://www.entomolog[...]
2014-10-25
[6]
서적
Thrips: Naturalists' Handbooks 25
The Richmond Publishing Company
[7]
서적
Bugs Britannica
https://books.google[...]
Chatto & Windus
[8]
웹사이트
Thysanoptera
https://www.merriam-[...]
Merriam-Webster
2017-02-15
[9]
서적
Encyclopedia of Entomology
https://books.google[...]
Springer Science & Business Media
[10]
LSJ
θύσανος, πτερόν
[11]
서적
Entomology
https://archive.org/[...]
Springer
[12]
논문
Functional morphology of the thysanopteran pretarsus
[13]
서적
Towards Understanding Thysanoptera. Proceedings of the International Conference on Thrips
USDA Technical Report NE-147
[14]
논문
A new look at thrips (Thysanoptera) mouthparts, their action and effects of feeding on plant tissue
[15]
웹사이트
Thrips: Biology and Rose Pests
https://entomology.c[...]
2024-10-14
[16]
웹사이트
Biology and Management of Thrips Affecting the Production Nursery and Landscape
https://extension.ug[...]
2024-10-14
[17]
웹사이트
Thrips (Thysanoptera: Thripidae)
https://agsci.colost[...]
[18]
서적
Biological diversity
CAB International
[19]
웹사이트
Thrips Wiki
http://thrips.info/w[...]
2017-02-19
[20]
논문
Thrips pollination of Mesozoic gymnosperms
Proceedings of the National Academy of Sciences
2012-05-21
[21]
논문
A new genus and species of Stenurothripidae (Insecta: Thysanoptera: Terebrantia) from mid-Cretaceous Myanmar amber
Elsevier BV
[22]
논문
New stenurothripid thrips from mid-Cretaceous Kachin amber (Thysanoptera, Stenurothripidae)
Pensoft Publishers
2024-02-21
[23]
논문
Higher-level phylogeny of paraneopteran insects inferred from mitochondrial genome sequences
[24]
논문
Evolution of Thrips (Thysanoptera) Phylogenetic Patterns and Mitochondrial Genome Evolution
[25]
논문
Phylogeny of thrips (Insecta: Thysanoptera) based on five molecular loci
[26]
논문
Order Thysanoptera Haliday, 1836 in Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness
http://www.mapress.c[...]
[27]
논문
Homologies and Host-Plant Specificity: Recurrent Problems in the Study of Thrips
[28]
논문
Molecular Identification Key for Pest Species of Scirtothrips (Thysanoptera: Thripidae)
https://www.research[...]
[29]
논문
Mesozoic Thrips and Early Evolution of the Order Thysanoptera (Insecta)
[30]
논문
Invasion Biology of Thrips
https://doi.org/10.1[...]
[31]
논문
Thysanoptera as predators: Their diversity and significance as biological control agents
https://doi.org/10.1[...]
[32]
논문
Two new ectoparasitic species of Aulacothrips Hood, 1952 (Thysanoptera: Heterothripidae) associated with ant-tended treehoppers (Hemiptera)
[33]
논문
Biology of a new species of socially parasitic thrips (Thysanoptera: Phlaeothripidae) inside Dunatothrips nests, with evolutionary implications for inquilinism in thrips: A NEW INQUILINE THRIPS
2012
[34]
논문
Natural history and behaviour of Dunatothrips aneurae Mound (Thysanoptera: Phlaeothripidae), a phyllode-gluing thrips with facultative pleometrosis: Natural History of Dunatothrips Aneurae
2013
[35]
논문
Egg predation within the nests of social wasps: a new genus and species of Phlaeothripidae, and evolutionary consequences of Thysanoptera invasive behaviour
[36]
서적
The Biology of the Codling Moth as the Basis for Its Control
Univerzitet U Beogradu
[37]
서적
Feeding behavior and nutritional requirements
Plenum Press
[38]
웹사이트
Onion Thrips
http://ipm.ncsu.edu/[...]
NCSU
2017-02-23
[39]
논문
The significance of prey in the diet of the phytophagous thrips, ''Frankliniella schultzei''
[40]
논문
Odor-Mediated Push-Pull Pollination in Cycads
2007
[41]
논문
Thrips: the primeval pollinators?
[42]
논문
Thrips pollination of Mesozoic gymnosperms
https://pnas.org/doi[...]
2012
[43]
논문
Thrips pollination of androdioecious ''Castilla elastica'' (Moraceae) in a seasonal tropical forest
[44]
논문
Pollination and gene flow in chillies with ''Scirtothrips dorsalis'' as pollen vectors
[45]
논문
Generalist flowers, biodiversity and florivory: implications for angiosperm origins
[46]
서적
The effects of cross and self fertilization in the vegetable kingdom
https://archive.org/[...]
D. Appleton & Company
[47]
논문
Thrips (Thysanoptera) pollination in Australian subtropical rainforests, with particular reference to pollination of ''Wilkiea huegeliana'' (Monimiaceae)
[48]
논문
Minute pollinators: The role of thrips (Thysanoptera) as pollinators of pointleaf manzanita, ''Arctostaphylos pungens'' (Ericaceae)
[49]
논문
The role of thrips in pollination of ''Arctostaphyllos uva-ursi''
[50]
서적
Feeding and oviposition injuries to plants
CAB International
[51]
논문
Thysanoptera: diversity and interactions
[52]
논문
The distribution of injurious thrips in the United States
[53]
논문
Bionomics of Thrips
[54]
논문
Problems with the Concept of "Pest" among the Diversity of Pestiferous Thrips
[55]
웹사이트
"Thrips tabaci" (onion thrips)
http://www.cabi.org/[...]
CABI
2017-02-24
[56]
논문
A new leaf-galling ''Holopothrips'' (Thysanoptera: Phlaeothripidae) and the structural alterations on ''Myrcia retorta'' (Myrtaceae)
[57]
논문
Austral Thysanoptera: 100 years of progress
[58]
논문
Towards understanding Thysanoptera: Chemical ecology of the Thysanoptera
[59]
논문
Defense in Thrips: Forbidding Fruitiness of a Lactone
[60]
논문
Chemical defence in thrips
[61]
논문
Mating Aggregations and Mating Success in the Flower Thrips, ''Frankliniella schultzei'' (Thysanoptera: Thripidae), and a Possible Role for Pheromones
[62]
서적
Encyclopedia of Insects
https://archive.org/[...]
Academic Press
[63]
논문
Social biology and sex ratios of the eusocial gall-inducing thrips ''Kladothrips hamiltoni''
[64]
논문
A fully reproductive fighting morph in a soldier clade of gall-inducing thrips (''Oncothrips morrisi'')
[65]
서적
The evolution of social behaviour of insects and arachnids
Cambridge University Press
[66]
논문
High relatedness and inbreeding in two species of haplodiploid eusocial thrips (Insecta: Thysanoptera) revealed by microsatellite analysis
[67]
논문
Life history of ''Kladothrips ellobus'' and ''Oncothrips rodwayi'': insight into the origin and loss of soldiers in gall-inducing thrips
[68]
논문
Quick estimates of flight fitness in hovering animals, including novel mechanisms for lift production
http://jeb.biologist[...]
[69]
논문
The aerodynamics of insect flight
http://www.cs.washin[...]
[70]
논문
Dissecting Insect Flight
http://www.physics.e[...]
[71]
논문
On the Weis-Fogh mechanism of lift generation
[72]
논문
Cereal pests among nest parasites – the story of barley thrips, Limothrips denticornis Haliday (Thysanoptera: Thripidae)
https://journal.fi/e[...]
2019-08-15
[73]
논문
Meniscus ascent by thrips (Thysanoptera)
[74]
서적
The Insects: An Outline of Entomology
https://archive.org/[...]
Wiley
[75]
논문
Facultative viviparity in a thrips
https://www.nature.c[...]
1989
[76]
논문
Subsociality and female reproductive success in a mycophagous thrips: An observational and experimental analysis
http://link.springer[...]
1990
[77]
논문
The ultrastructure of malpighian tubules and the chemical composition of the cocoon of Aeolothrips intermedius Bagnall (Thysanoptera)
https://onlinelibrar[...]
2009
[78]
서적
Handbook of Vegetable Pests
https://books.google[...]
Gulf
[79]
논문
Evolution of asexuality via different mechanisms in grass thrips (Thysanoptera: Aptinothrips)
http://www.rug.nl/re[...]
[80]
논문
Egg size-mediated sex allocation and mating-regulated reproductive investment in a haplodiploid thrips species
2020
[81]
논문
First detection of Wolbachia in arrhenotokous populations of thrips species (Thysanoptera: Thripidae and Phlaeothripidae) and its role in reproduction
[82]
논문
The thrips, or Thysanoptera, of Illinois
[83]
논문
Arthropod transmission of plant viruses: a new synthesis
[84]
논문
So many thrips – so few tospoviruses?
2001
[85]
논문
Invasion Biology of Thrips
[86]
논문
Plant Viruses Transmitted by Thrips
http://link.springer[...]
2005
[87]
웹사이트
Western Flower Thrips, Management and Tospoviruses
https://ag.umass.edu[...]
University of Massachusetts Amherst
2017-02-21
[88]
서적
Invasive Species: Vectors And Management Strategies
https://books.google[...]
Island Press
[89]
논문
A review of thrips species biting man including records in Florida and Georgia between 1986–1997
2005
[90]
논문
Insecticide rotation programs with entomopathogenic organisms for suppression of western flower thrips (Thysanoptera: Thripidae) adult populations under greenhouse conditions
[91]
웹사이트
Western flower thrips in greenhouses: a review of its biological control and other methods
http://biocontrol.uc[...]
University of California, Riverside
2017-02-21
[92]
논문
Efficacy of red LED irradiation for controlling ''Thrips palmi'' in greenhouse melon cultivation
[93]
서적
バイオディバーシティ・シリーズ 節足動物の多様性と系統
裳華房
2008-04-05
[94]
논문
アリガタシマアザミウマの生態と防除効果
http://jppa.or.jp/ar[...]
[95]
PDF
アリガタシマアザミウマの生態
https://www.maff.go.[...]
農林水産省
[96]
웹사이트
アリガタシマアザミウマの生物農薬としての特徴
http://www.jppa.or.j[...]
日本植物防疫協会
[97]
간행물
Mesozoic Thrips and Early Evolution of the Order Thysanoptera (Insecta)
2004-09
[98]
간행물
Biological diversity.
CAB International, Wallingford, UK
[99]
웹인용
총채벌레아목의 분류학적 연구
http://www.naas.go.k[...]
2012-02-08
[100]
웹사이트
Strepsiptera. Twisted-wing parasites. Version 24 September 2002
http://tolweb.org/St[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com