코브라과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
코브라과는 전 세계 열대 및 아열대 지역에 분포하며, 신경독을 가진 독사들을 포함하는 뱀 과이다. 육상 및 해양 환경에 적응하여 다양한 형태와 크기를 가지며, 킹코브라가 가장 긴 종으로 알려져 있다. 코브라과는 코브라아과와 바다뱀아과로 나뉘며, 일부 종은 식용으로 이용되기도 하지만, 독성으로 인해 위험하다. IUCN 적색 목록에 위협받는 종이 등재되어 있으며, 일본에서는 특정동물로 지정되어 사육이 금지되었다.
더 읽어볼만한 페이지
| 코브라과 - [생물]에 관한 문서 | |
|---|---|
| 일반 정보 | |
 | |
 | |
 | |
 | |
| 학명 | Elapidae |
| 명명자 | F. Boie, 1827 |
| 어원 | 그리스어 ἔλαψ (élaps, "코브라") |
| 분류 | |
| 아과 및 속 | Calliophiinae Micrurinae Naja Bungarus Elapsoidea Hydrophiinae |
| 특징 | |
| 길이 | 18 cm ~ 5.85 m |
| 설명 | 코브라과는 독사의 한 과이다. 과거에는 육상 종으로 이루어진 Elapinae 아과와 해양 종으로 이루어진 Hydrophiinae 아과가 있는 것으로 여겨졌다. 1997년 슬로윈스키, 나이트, 루니는 독 단백질의 아미노산 서열을 사용한 계통 분석에서 오스트레일리아 육상 종이 Hydrophiinae 내에 중첩되어 있음을 발견했다. 이로 인해 오스트레일리아 육상 종을 제거하고 Hydrophiinae에 배치하게 되었다. 이후의 최근 유전체 분석에서도 이는 지지되었지만, 이 연구들은 Elapinae 아과가 Hydrophiinae에 대해 측계통군임을 발견했다. 이 연구들은 산호뱀, 코브라, 맘바, 크레이트, 아프리카 가터뱀이 Hydrophiinae에 대한 연속적인 외집단을 형성한다는 것을 발견했다. 이 그룹에 사용할 수 있는 분기군 이름이 있으므로 (Elapsoidea 제외), Calliophiinae, Micrurinae, Najinae 및 Bungarinae 아과를 다시 가져오는 것이 이상적이다. |
2. 분포
코브라과는 전 세계 열대 및 아열대 지역에 걸쳐 분포한다. 대부분 남반구의 습한 열대 환경을 선호하지만, 건조한 환경에서도 발견되는 종도 많다. 육상 코브라과는 아프리카, 오스트레일리아, 북아메리카, 남아메리카, 유라시아 대륙 남부, 인도네시아, 스리랑카, 일본 (난세이 제도), 파푸아뉴기니, 필리핀, 타이완 등지에 서식한다. 바다뱀류는 주로 인도양과 남서 태평양의 연안 해역과 얕은 곳에 서식하며, 산호초에서 흔히 발견된다.[22] ''Hydrophis platurus''는 태평양을 가로질러 중앙아메리카와 남아메리카 해안까지 분포한다.
코브라과는 다양한 크기를 가진다. 킹코브라는 최대 몸길이 5.85m에 달하는 세계에서 가장 긴 독사이다.[7] 육상 코브라과는 뱀과와 유사하게 길고 가느다란 몸, 큰 비늘로 덮인 머리, 둥근 동공을 가진 눈을 가진다.
코브라과는 육상, 수상, 굴을 파는 등 다양한 서식 환경에 적응했다. 대부분의 코브라과는 육상에 서식하지만, 일부는 나무 위에서 생활하는 경향이 강하다 (아프리카의 ''Pseudohaje''와 ''Dendroaspis'', 호주의 ''Hoplocephalus'').[7] 많은 종은 습하거나 건조한 환경에서 굴을 파는 데 특화되어 있다(예: ''Ogmodon'', ''Parapistocalamus'', ''Simoselaps'', ''Toxicocalamus'', ''Vermicella'').[7]
코브라과는 위턱 뒤쪽에 위치한 한 쌍의 전구치아 엄니를 통해 독을 주입한다. 이 엄니는 상악골에 있는 두 개의 이빨 중 확대되고 속이 빈 형태로, 입을 다물면 뺨 바닥의 홈에 맞물린다. 일부 코브라과는 움직이는 상악골에 긴 엄니를 가지고 있어 빠르게 공격할 수 있으며, 침뱉는 코브라처럼 방어를 위해 엄니 앞쪽 구멍에서 독을 분사하기도 한다.
코브라과는 과거 뱀상과에 속했지만, 분자 계통 분석 결과 왕뱀과 등과 함께 코브라상과로 분류된다.[28][29] 코브라과는 크게 코브라아과와 바다뱀아과(광의)로 나뉜다.
3. 형태
바다뱀(Hydrophiinae)은 수생 생활에 적응하여 노와 같은 꼬리를 가지며, 몸이 옆으로 압축되어 있다. 복부 비늘 크기가 작아져 육상 이동성은 크게 감소했지만, 수영에는 도움이 된다.[7]
모든 코브라과는 위턱 앞쪽에 독을 주입하기 위한 한 쌍의 전구치아 엄니를 가지고 있다.[7] 일부 코브라과는 이 엄니를 통해 방어를 위해 독을 분사할 수 있다.[7]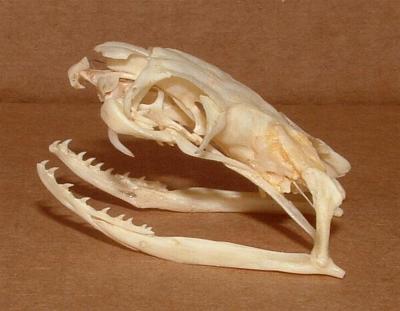
4. 생태
먹이는 종에 따라 다양하며, 양서류, 파충류, 조류, 포유류, 어류, 어란 등을 먹는다.[25] 일부 종은 매우 일반적인 먹이를 먹지만(광식성), 많은 종이 좁은 먹이 선호도(협식성)와 관련된 형태학적 특성을 가지고 있다. 예를 들어 다른 뱀, 특히 코브라와 해독사만을 거의 먹는 경우가 있다.[7]
번식 방식은 난생이 주를 이루지만, 오스트레일리아 주변에 분포하는 종이나 바다뱀류는 태생인 경우도 있다.[25]
위협을 받으면 코브라와 맘바는 몸을 들어 올리고 후드를 펼치며 쉭쉭거리는 소리를 낸다. 해독사는 종종 머리를 몸 안으로 숨기기 전에 몸을 웅크린다.[7]
일반적으로 바다뱀은 피부를 통해 호흡할 수 있다. 황복 바다뱀(''Hydrophis platurus'')을 대상으로 한 실험에 따르면, 이 종은 이러한 방식으로 산소 요구량의 약 20%를 충족할 수 있어 잠수를 오랫동안 할 수 있다.[7] 바다해독사(''Laticauda'' spp.)는 수생 생활에 가장 덜 적응된 바다뱀으로, 몸이 측면으로 덜 압축되어 있고, 더 두꺼운 몸과 복부 비늘을 가지고 있어 육상 이동이 가능하다. 이들은 알을 낳고 먹이를 소화하는 데 대부분의 시간을 육지에서 보낸다.[7]
5. 독
코브라과의 독은 주로 신경독으로 작용하여 먹이를 마비시키고 방어한다. 주요 독소군은 PLA2와 세 손가락 독소(3FTx)이다. 일부 종은 심장독, 세포독을 포함하며, 코브라 독에는 혈액독도 들어 있다. 맘바와 코브라 같은 대형 종은 한 번 물릴 때 많은 양의 독을 주입할 수 있어 위험하다.[9] 뱀에 물린 경우 즉시 항독소를 투여해야 하며, 특정 항독소는 뱀에 물린 상처를 치료하는 유일한 치료법이다.[10]
침을 뱉는 코브라의 독은 신경독보다 세포독성이 더 강하며, 눈에 닿으면 심한 통증과 실명을 유발할 수 있다. 하지만 상처가 없어 혈액으로 들어가지 않으면 피부에는 치명적이지 않다.[11] 코브라과 뱀의 독은 신경독으로, 먹이의 신경 방전을 막아 마비, 저림, 호흡 및 심장 정지를 일으켜 죽음에 이르게 한다. 타이판처럼 출혈독을 가진 종류나, 뱉는 코브라처럼 세포독을 가진 종도 있다. 사람이 물린 경우, 신속하게 의료기관에 연락하여 구급 이송을 받아야 한다.
6. 분류
육상 코브라과 뱀은 Colubridae와 유사하게 생겼다. 거의 모든 종은 매끄러운 비늘을 가진 길고 가느다란 몸, 큰 비늘로 덮인 머리(항상 목과 구별되지 않음), 둥근 동공을 가진 눈을 가지고 있다. 또한 뱀과와 마찬가지로, 대개 매우 활동적이고 빠르며, 대부분의 암컷은 난생이다.
바다뱀(Hydrophiinae)은 때때로 별도의 과로 간주되는데, 다양한 방식으로 해양 생활에 적응했다. 모든 종은 수영을 위한 노와 같은 꼬리를 진화시켰고, 염분을 배설하는 능력을 갖추었다. 대부분은 옆으로 압축된 몸을 가지고 있으며, 복부 비늘 크기가 훨씬 작고, 콧구멍이 등쪽에 위치하며(내비늘 없음), 새끼를 낳는다(난태생). 복부 비늘 감소는 육상 이동성을 크게 감소시켰지만, 수영에 도움이 된다.
이 과의 구성원은 다양한 크기를 가진다. ''Drysdalia'' 종은 일반적으로 18cm에서 50cm까지 작은 뱀이다. 코브라, 맘바, 타이판은 2m 이상에 도달할 수 있는 중간에서 큰 크기의 뱀이다. 킹코브라는 세계에서 가장 긴 독사로, 최대 길이 5.85m이고 평균 질량은 6kg이다.[7]
과거에는 코브라과에 대해 많은 아과가 인정되거나 제안되었는데, 여기에는 뱀과(Elapinae), 바다뱀과(Hydrophiinae) (바다뱀), 산호뱀과(Micrurinae) (산호뱀), Acanthophiinae (호주 코브라과), 그리고 Laticaudinae (바다 뱀) 등이 있었다. 현재는 어느 것도 보편적으로 인정되지 않는다. 핵형 분석, 단백질 전기영동 분석, 면역학적 거리 및 DNA 염기서열 분석과 같은 기술을 통한 분자적 증거는 아프리카, 아시아, 신세계의 Elapinae 대 호주 및 해양 Hydrophiinae의 두 그룹의 상호 단일계통성을 시사한다. 호주의 육상 코브라과는 기술적으로 'hydrophiines'이지만 바다뱀은 아니다. ''Laticauda''와 '진정한 바다뱀'은 호주 육상 뱀에서 별도로 진화한 것으로 여겨진다. 아시아 코브라, 산호뱀 및 아메리카 산호뱀도 단일계통군으로 보이며 아프리카 코브라는 그렇지 않다.[12][13]
6. 1. 코브라아과
| 속 | 분류군 저자 | 종 | 아종* | 일반 이름 | 지리적 범위 |
|---|---|---|---|---|---|
| 산호코브라속 (Aspidelaps) | 피칭거, 1843 | 2 | 4 | 방패 코 코브라 | 남아프리카 공화국 (케이프 주, 트란스발), 나미비아, 남부 앙골라, 보츠와나, 짐바브웨, 모잠비크 |
| 우산뱀속 (Bungarus) | 도댕, 1803 | 12 | 4 | 해구 | 인도 (안다만 제도 포함), 미얀마, 네팔, 베트남, 아프가니스탄, 파키스탄, 스리랑카, 방글라데시, 캄보디아, 인도네시아 (자바, 수마트라, 발리, 술라웨시), 말레이 반도, 싱가포르, 대만, 태국 |
| 둥근무늬산호뱀속 (Calliophis) | 그레이, 1834 | 15 | 11 | 동양 산호뱀 | 인도, 방글라데시, 스리랑카, 네팔, 인도네시아, 캄보디아, 말레이시아, 싱가포르, 태국, 버마, 브루나이, 필리핀, 베트남, 라오스, 중국 남부, 일본(류큐 열도), 대만 |
| 맘바속 (Dendroaspis) | 슐레겔, 1848 | 4 | 1 | 맘바 | 사하라 사막 이남 아프리카 |
| Elapsoidea | 보카주, 1866 | 10 | 7 | 아프리카 또는 독성 가터뱀 (인간에게 무해한 북미 가터뱀과는 관련 없음) | 사하라 사막 이남 아프리카 |
| Hemachatus | 플레밍, 1822 | 1 | 0 | 링칼스/목걸이 뱉는 코브라 | 남아프리카 공화국, 짐바브웨, 레소토, 에스와티니 |
| Hemibungarus | 페터스, 1862 | 3 | 0 | 줄무늬 산호뱀 | 필리핀 (루손, 파나이, 네그로스, 세부, 민도로, 카탄두아네스, 폴릴로 섬) |
| Micruroides | K.P. 슈미트, 1928 | 1 | 2 | 서부 산호뱀 | 미국 (애리조나, 뉴멕시코 남서부), 멕시코 (소노라, 시날로아) |
| 산호뱀속 (Micrurus) | 바글러, 1824 | 83 | 51 | 산호뱀 | 남부 북미, 남아메리카 |
| 코브라속 (Naja) | 로렌티, 1768 | 39 | 3 | 코브라 | 아프리카, 아시아 |
| 킹코브라속 (Ophiophagus) | 귄터, 1864 | 4[17] | 1 | 킹 코브라 | 방글라데시, 미얀마, 캄보디아, 중국, 인도, 안다만 제도, 인도네시아, 라오스, 태국, 베트남, 서 말레이시아, 필리핀 |
| Pseudohaje | 귄터, 1858 | 2 | 0 | 나무 코브라 | 앙골라, 부룬디, 카메룬, 중앙아프리카 공화국, 콩고 민주 공화국, 콩고, 가봉, 가나, 케냐, 나이지리아, 르완다, 우간다, 시에라리온, 라이베리아, 코트디부아르, 토고, 나이지리아 |
| Sinomicrurus | 슬로윈스키 외, 2001 | 8 | 6 | 아시아 | |
| Walterinnesia | 라타스트, 1887 | 2[18] | 0 | 검은 사막 코브라 | 이집트, 이스라엘, 레바논, 시리아, 요르단, 이라크, 이란, 쿠웨이트, 사우디 아라비아, 터키[19] |
''* 기준 아종은 포함하지 않음''
6. 2. 바다뱀아과 (광의)
바다뱀아과는 동남아시아에서 오스트레일리아에 분포하는 육상 서식종과 인도-태평양에 분포하는 해양 서식종을 포함한다. 전통적으로는 해양 서식종만을 바다뱀아과 또는 바다뱀과로 분류했다. 바다뱀아과는 난생종과 난태생종이 있으며, 해양 바다뱀은 난태생종에서 진화한 것으로 추정된다.[31] 다만, 에라브바다뱀속은 예외적으로 난생이다. 난태생종은 거의 대부분 오스트레일리아 고유종이다.이 무리의 특징은 입을 벌렸을 때 구개가 익상골에 대해 들리지 않는다는 점이다. 과거에는 에라브바다뱀속을 에라브바다뱀아과(Laticaudinae), 육상 서식종을 타이판아과(Oxyuraninae)로 구분하기도 했지만, 분자 계통 분석 결과 타이판아과가 측계통군으로 밝혀져 바다뱀아과에 포함되었다.[31]
다음은 바다뱀아과(광의)에 속하는 속(genus)들이다.
| 속 | 종 수 | 주요 서식지 및 특징 |
|---|---|---|
| 데스아더속 (Acanthophis) | 8 | 호주, 뉴기니, 인도네시아 (세람 섬 및 타님바르 제도). 코먼데스아더 |
| 크로우나지뱀속 (Antaioserpens) | 2 | |
| 아스프모도키속 (Aspidomorphus) | 3 | |
| 오스트레일리아구리머리뱀속 (Austrelaps) | 3 | |
| 고슈산호뱀속 (Brachyurophis) | 8 | |
| 관두꺼비뱀속 (Cacophis) | 4 | |
| 은밀뱀속 (Cryptophis) | 5 | |
| 채찍뱀속 (Demansia) | 14 | |
| 데니슨뱀속 (Denisonia) | 2 | |
| 히바카리모도키속 (Drysdalia) | 3 | |
| 버딕속 (Echiopsis) | 1 | |
| 히메차뱀속 (Elapognathus) | 2 | 히메차뱀 |
| 히우나지뱀속 (Furina) | 5 | |
| 그레이아스프속 (Hemiaspis) | 2 | 늪지에 서식하며, 진정한 바다뱀류와 가장 근연인 것으로 여겨진다. |
| 미나미오오즈뱀속 (Hoplocephalus) | 3 | |
| 에라브바다뱀속 (Laticauda) | 8 | 에라브바다뱀. 예외적으로 난생. |
| 모리노사메뱀속 (Loveridgelaps) | 1 | 모리노사메뱀 |
| 이카헤뱀속 (Micropechis) | 1 | 이카헤뱀 |
| 줄무늬복면뱀속 (Neelaps) | 1 | |
| 타이거뱀속 (Notechis) | 1 | |
| 피지뱀속 (Ogmodon) | 1 | 독아시뱀속과 마찬가지로 지렁이를 먹이로 하며, 이 두 속은 근연 관계이다. |
| 타이판속 (Oxyuranus) | 3 | 타이판 |
| 부겐빌뱀속 (Parapistocalamus) | 1 | 부겐빌뱀 |
| 검은두건뱀속 (Parasuta) | 6 | |
| 즈그로버딕속 (Paroplocephalus) | 1 | |
| 블랙스네이크속 (Pseudechis) | 9 | |
| 브라운스네이크속 (Pseudonaja) | 9 | |
| 후타이로은밀뱀속 (Rhinoplocephalus) | 1 | |
| 솔로몬뱀속 (Salomonelaps) | 1 | 솔로몬뱀 |
| 복면뱀속 (Simoselaps) | 5 | |
| 칼뱀속 (Suta) | 4 | |
| 독아시뱀속 (Toxicocalamus) | 15 | 코브라과에서는 예외적으로 지렁이를 먹는다. |
| 자라하다뱀속 (Tropidechis) | 1 | |
| 반디반디속 (Vermicella) | 6 |
협의의 바다뱀아과는 70종 정도의 그룹으로, 수생 생활에 적응하고 있다.
7. 인간과의 관계
코브라과는 독이 있어 위험하지만, 일부 지역에서는 식용으로 사용되기도 한다. 특히 심장은 갓 잡은 직후에 먹으면 맛있다고 알려져 있다. 하지만, 적어도 일본에서는 별미로 취급하는 것이 일반적이다.[32]
IUCN 적색 목록에는 일부 코브라과 종들이 위협받거나 취약 근접종으로 등재되어 있다. 예를 들어, 킹코브라(`Ophiophagus hannah`)는 취약종으로, 짧은코뱀 (`Elapognathus minor`)은 취약 근접종으로 분류된다.[20] 하지만, 이는 위협받는 뱀과류 전체 숫자를 보여주는 것은 아니다. 많은 종들이 외딴 지역이나 넓은 바다에 서식하여 개체수 조사가 어렵기 때문이다.[20]
2020년 기준으로 일본에서는 코브라과 전체가 특정동물로 지정되어 애완 목적의 사육이 금지되었다.[32]
참조
[1]
서적
The New Encyclopedia of Snakes
Princeton University Press
[2]
논문
Inferring species trees from gene trees: a phylogenetic analysis of the Elapidae (Serpentes) based on the amino acid sequences of venom proteins
1997
[3]
논문
A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes
2013
[4]
논문
Diversification rates and phenotypic evolution in venomous snakes (Elapidae)
2016
[5]
논문
A Species-Level Phylogeny of Extant Snakes with Description of a New Colubrid Subfamily and Genus
2016
[6]
웹사이트
Definition of 'elapid'.
http://dictionary.re[...]
dictionary.com
2009-07-13
[7]
논문
Characterization of venomous snakes of Thailand.
2011-06
[8]
간행물
2008-11-03
[9]
웹사이트
LD50 Scores for various snakes
http://www.seanthoma[...]
1999-12
[10]
논문
Horse immunization with short-chain consensus α-neurotoxin generates antibodies against broad spectrum of elapid venomous species
2019-08
[11]
웹사이트
Guidelines for the Prevention and Clinical Management of Snakebite in Africa
https://digicollecti[...]
2010
[12]
논문
Phylogenetic relationships of elapid snakes based on cytochrome b mtDNA sequences
https://biology-asse[...]
2000-04
[13]
논문
The good, the bad and the ugly: Australian snake taxonomists and a history of the taxonomy of Australia's venomous snakes
http://www.kingsnake[...]
2006-12
[14]
간행물
2006-11-27
[15]
웹사이트
The Hydrophiidae
http://www.cyberliza[...]
2007-08-12
[16]
간행물
Scanlon J,Lee M, Archer M, 2002, Mid-Tertiary elapid snakes (Squamata, Colubroidea) from Riversleigh, northern Australia: early steps in a continent-wide adaptive radiation, Geobios 36 (2003) 573–601
http://www.create.un[...]
2019-03-12
[17]
논문
Taxonomic revision of the king cobra ''Ophiophagus hannah'' (Cantor, 1836) species complex (Reptilia: Serpentes: Elapidae), with the description of two new species
https://europeanjour[...]
[18]
논문
"Walterinnesia aegyptia Lataste, 1887 (Ophidia: Elapidae) and the status of Naja morgani Mocquard, 1905"
2007
[19]
논문
"New record of Walterinnesia aegyptia Lataste, 1887 (Ophidia: Elapidae: Bungarinae) in Turkey"
2001
[20]
문서
2013
[21]
웹사이트
日本産爬虫両生類標準和名リスト(2024年3月11日版)
http://herpetology.j[...]
2024
[22]
서적
陸から海への適応 ウミヘビ
朝日新聞社
1993
[23]
서적
特異な威嚇行動と防御行動 コブラ、サンゴヘビ
朝日新聞社
1993
[24]
서적
全世界に分布する毒ヘビ類は、獲物を鎮圧するために毒と注毒機構を発達させてきた。
朝日新聞社
1993
[25]
서적
ヘビ類
平凡社
1986
[26]
서적
最も強力な毒を持つ タイパン亜科
朝日新聞社
1993
[27]
서적
爬虫綱分類表
裳華房
2006
[28]
논문
The phylogeny and classification of caenophidian snakes inferred from seven nuclear protein-coding genes
Académie des sciences
2007
[29]
서적
ヘビ大図鑑 ナミヘビ上科、他編:分類ほか改良品種と生態・飼育・繁殖を解説
誠文堂新光社
2021
[30]
서적
毒ヘビ全書
グラフィック社
2020-02-25
[31]
논문
2007年以降の日本産爬虫類の分類の変遷について
日本爬虫両棲類学会
[32]
웹사이트
特定動物リスト (動物の愛護と適切な管理)
https://www.env.go.j[...]
環境省
[33]
웹인용
Definition of 'elapid'.
http://dictionary.re[...]
dictionary.com
2009-07-13
[34]
저널
A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes.
http://www.biomedcen[...]
[35]
저널
A Taxonomic Revision of Boas (Serpentes: Boidae)
http://home.gwu.edu/[...]
[36]
저널
Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species
http://www.reptile-d[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com