T세포
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
T세포는 잭 밀러에 의해 흉선이 면역에 미치는 영향이 처음으로 제시된 림프구의 일종이다. T세포는 항원을 만나지 못한 미접촉 T세포, 항원을 만나 성숙한 효과 T세포, 기억 T세포로 분류되며, CD4 또는 CD8 표지자를 발현한다. T세포는 흉선에서 분화 및 성숙 과정을 거치며, 활성화는 항원 제시 세포의 MHC 분자에 결합된 T 세포 수용체 펩타이드의 인지에 의해 이루어진다. T세포 결핍, T세포 암, T세포 고갈 등 다양한 질환과 관련이 있으며, 자가 면역 질환, 이식 거부 반응에도 관여한다.
더 읽어볼만한 페이지
- T 세포 - CD4
CD4는 보조 T 세포 표면에서 T 세포 수용체의 공동수용체로 작용하여 항원제시세포와 상호작용을 돕고 T 세포 활성화 신호를 증폭시키는 면역계의 당단백질이며, HIV의 숙주 세포 침투 수용체로 작용하여 HIV 감염 및 AIDS 발병에 중요한 역할을 한다. - T 세포 - T세포 수용체
T세포 수용체(TCR)는 T 세포 표면의 이량체 단백질 복합체로, MHC 분자에 결합된 펩타이드를 인식하며, CD3 분자와 복합체를 이루어 신호 전달을 매개하고, V(D)J 재조합을 통해 다양한 항원 특이성을 갖는 T 세포 생성에 기여하여, CAR-T 치료 등 면역 요법 발전에 기여했다. - 백혈구 - 과립구
과립구는 골수에서 생성되어 식세포 작용과 독성 물질 방출을 통해 면역 반응에 기여하며, 호중구, 호산구, 호염기구, 비만 세포로 분류된다. - 백혈구 - 호중구
호중구는 혈액 내에서 가장 풍부한 백혈구로서 세균 감염에 대한 신속한 면역 반응에 중요한 역할을 하며, 식세포 작용, 탈과립, 호중구 세포외덫 형성 등의 항미생물 작용을 수행하고 염증 반응을 조절하며 사이토카인을 분비한다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.
| T세포 | |
|---|---|
| 개요 | |
 | |
| |
| 라틴어 | lymphocytus T |
| 시스템 | 면역계 |
| 설명 | |
| 정의 | 면역계의 백혈구 |
| 유래 | 기관에서 유래 |
| 기관 설명 | T세포는 흉선에서 성숙한다. |
| 역할 | CD4 양성 T세포: 분화 및 기능 |
| 추가 이미지 | |
| |
2. 역사
1961년, 런던의 체스터 비티 암 연구소의 잭 밀러는 흉선을 절제한 생쥐를 해부하여 림프절, 비장, 말초 혈액에서 림프구가 격감하고 면역 부전을 발병하며, 이식 시 거부 반응이 억제되는 것을 발견했다.[1] 1968년 G. F. 미첼 및 밀러는 생쥐의 흉관 림프에서 19S 용혈소(항 양 적혈구 항원 IgM 항체) 생성 세포 전구 세포(B 세포) 및 그 전구 세포를 항원 의존적으로 19S 용혈소 생성 세포로 분화시키는 세포(T 세포)에서 두 개의 림프구 아집단이 존재한다는 것을 발견했다.[1] 1975년에는 필리파 마락 및 존 캐플러가 한계 희석법을 통해 T 세포 클론 간의 명확한 기능적 차이에 대해 보고한 이후 다양한 T 세포 아집단, 나아가 그 하위 아집단의 존재가 제기되었다.[1] 1984년에는 맥덕화 및 Mark M. Davis가 각각 사람과 생쥐의 TCR을 코딩하는 cDNA 클론을 동정했다.[1]
T세포는 크게 흉선에서 유래하는 일반적인 적응면역 T세포와 선천면역에 관여하는 T세포로 나눌 수 있다.
3. 종류
일반적인 적응면역 T세포에는 항원을 아직 만나지 못한 미접촉 T세포, 항원을 만나 성숙한 효과 T세포 (보조 T세포, 세포독성 T세포), 기억 T세포가 있다.[94] NK 세포와 T 세포의 성질을 함께 갖는 NKT 세포나, 다른 T 세포의 활성을 억제하는 조절 T 세포 등도 여기에 포함된다.
말초에 존재하는 대부분의 성숙한 T 세포는 세포 표면 표지자 분자로 CD4 또는 CD8 중 하나를 발현한다. CD4 양성 T 세포는 B 세포의 분화 및 성숙, 항체 생산을 유도하는 등 다른 T 세포의 기능 발현을 돕는 헬퍼 T 세포로 기능하며, 후천성 면역 결핍 증후군(AIDS)의 병원 바이러스인 인간 면역 결핍 바이러스(HIV)와, 성인 T 세포 백혈병(ATL)의 병원 바이러스인 인간 T 세포 백혈병 바이러스(HTLV-1)에 감염되는 세포이다. CD8 양성 T 세포는 바이러스 감염 세포 등을 파괴하는 킬러 T 세포로 기능한다.[94]
최근에는 흉선을 거치지 않고 분화, 성숙하는 말초성 T 세포가 존재한다는 사실도 알려졌다.
3. 1. 일반적인 적응면역 T세포
T세포는 항원을 아직 만나지 못한 미접촉 T세포와, 항원을 만나 성숙한 효과 T세포(보조 T세포, 세포독성 T세포, 자연살상 T세포), 기억 T세포로 분류된다.[94]
말초에 존재하는 대부분의 성숙한 T 세포는 세포 표면 표지자 분자로 CD4 또는 CD8 중 하나를 발현한다. CD4 양성 T 세포는 B 세포의 분화 및 성숙, 항체 생산을 유도하는 등 다른 T 세포의 기능 발현을 돕는 헬퍼 T 세포로 기능한다. CD8 양성 T 세포는 바이러스 감염 세포 등을 파괴하는 킬러 T 세포로 기능한다.[94]
NK 세포와 T 세포의 성질을 함께 갖는 NKT 세포나, 다른 T 세포의 활성을 억제하는 조절 T 세포 등도 있다.
3. 1. 1. 미접촉 T세포
분화와 성숙을 거쳤지만 아직 말초에서 항원을 만나지 못한 T세포이다. 항원전달세포에 제시된, 아직 인지되지 않은 MHC:항원 복합체를 만나면 T세포 항원 수용체 신호 전달 과정(T-cell receptor signaling pathway)을 통해 항원을 인식하고 효과 T세포로 활성화되어 적응 면역이 시작된다. 표면에는 세포 접착 분자(cell adhesion molecule)인 L-셀렉틴(CD62L)이 존재하는 반면, 효과 T세포의 특징인 CD25, CD44, CD69와 기억 T세포의 특징인 CD45 등은 거의 존재하지 않는다.
3. 1. 2. 보조 T세포 (헬퍼 T세포, CD4+ T세포)
도움 T세포(helper T cell, 또는 Th cell)는 체액성 면역을 촉진하는 효과 T세포의 일종으로, 다른 백혈구들의 분화 및 활성화를 조절한다. 세포 표면에 CD4 단백질을 가지고 있기 때문에 CD4+ T세포라고도 불린다.
CD4+ T세포는 세부 기능에 따라 Th1, Th2, Th17, Treg 등으로 나뉜다.
흉선 세포는 양성 선택 과정에서 MHC ''클래스 II'' 분자와 잘 상호작용하면 CD4+ "헬퍼" 세포가 된다. 이 과정에서 CD8 세포 표면 수용체의 발현을 하향 조절하여 CD4+ 세포가 되며, 이후에는 ''단일 양성'' 세포가 된다.[11]
1986년 T. R. Mosmann 등은 마우스 T세포 클론 간 사이토카인 분비 패턴 차이에 따라 Th1 세포와 Th2 세포라는 두 가지 헬퍼 T 세포 아집단 개념을 처음 제시했다. CD4 양성 T 세포에서 분화한 Th1 세포는 IFN-γ를, Th2 세포는 IL-4나 IL-5를, Th17 세포는 IL-17을 생산하여 다른 세포의 활성화와 기능 수행을 돕는다.
Th1 세포는 세포성 면역을 매개하며 자가면역 질환과 지연형 알레르기에, Th2 세포는 체액성 면역을 매개하며 즉시형 알레르기에 관여한다. Th17 세포는 여러 자가면역 질환 모델 마우스에서 증가하여 자가면역 질환과 관련이 있는 것으로 여겨진다.
Th1과 Th2 세포를 분화시키거나 분화 후 생산되는 사이토카인은 서로의 세포군을 억제하고 조절하는 성질을 보인다. 즉, Th1/Th2 균형이 서로 길항하며 유지된다고 볼 수 있다.
흉선에서 분화하는 조절 T 세포는 CD4, CD25, Foxp3 분자를 발현하여 다른 T 세포 활성을 억제한다. 말초에서 항원 특이적으로 유도되거나 CD8 양성 T 세포에서 분화하는 조절 T 세포도 있다.
3. 1. 3. 세포독성 T세포 (킬러 T세포, CD8+ T세포)
세포독성 T세포는 킬러 T세포라고도 불리며, 그랜자임(granzyme)이나 퍼포린(perforin)과 같은 세포독성물질을 분비하여 바이러스에 감염된 세포나 종양 세포 등을 죽이는 세포이다.[94] 세포 표면에 CD8 단백질을 가지고 있기 때문에 CD8 T세포라고도 한다. 세포성 면역을 매개하여 바이러스 및 암세포를 제거하는 역할을 한다.[94]
흉선 세포의 운명은 양성 선택 과정에서 결정된다. MHC ''클래스 I'' 분자와 잘 상호작용하는 흉선 세포는 CD8+ "킬러" 세포로 성숙한다.[11] 이 과정은 자가면역을 일으킬 수 있는 흉선 세포를 걸러내지는 않으며, 이러한 세포는 흉선 수질에서 발생하는 음성 선택 과정을 통해 제거된다.
말초에 존재하는 대부분의 성숙한 T 세포는 세포 표면의 표지자 분자로서 CD4 또는 CD8 중 하나를 발현하고 있다. CD8 양성 T 세포는 바이러스 감염 세포 등을 파괴하는 킬러 T 세포(CTL)로서 기능한다.[94]
3. 1. 4. 기억 T세포
기억 T세포는 항원을 인지한 T세포가 분화 및 선별 과정을 거친 뒤 장기간 생존하고 있다가 나중에 항원이 재차 침입하였을 때 빠르게 활성화되어 효과 T세포의 기능을 할 수 있는 잠재적 능력을 가진 세포이다.[94] 미접촉 T세포가 항원을 만나 활성화된 상태의 세포, 또는 효과 T세포가 인터루킨-7(IL-7)과 인터루킨-15(IL-15)의 영향을 받아 장기 생존 가능한 기억 T세포로 분화하게 된다.
3. 2. 선천면역에 관여하는 T세포
말초에 존재하는 대부분의 성숙한 T 세포는 세포 표면의 표지자 분자로서 CD4 또는 CD8 중 하나를 발현한다. CD4를 발현하는 T 세포는 헬퍼 T 세포로서, 다른 T 세포의 기능 발현을 유도하거나 B 세포의 분화 및 성숙, 항체 생산을 유도한다. CD4 양성 T 세포는 후천성 면역 결핍 증후군(AIDS)의 원인 바이러스인 인간 면역 결핍 바이러스(HIV)와, 성인 T 세포 백혈병(ATL)의 원인 바이러스인 인간 T 세포 백혈병 바이러스(HTLV-1)에 감염되는 세포이다. CD8 양성 T 세포는 킬러 T 세포로서 바이러스 감염 세포 등을 파괴한다. 이 외에도 CD25 분자를 발현하여 다른 T 세포의 활성을 억제하는 조절 T 세포 등이 있다. 최근에는 흉선을 거치지 않고 분화, 성숙하는 말초성 T 세포가 존재한다는 사실도 알려졌다.
3. 2. 1. 자연살상 T세포 (NKT세포)
NK 세포와 T 세포의 성질을 함께 갖는 효과 T세포의 일종이다. 표면에 T세포 항원수용체(T cell receptor, TCR)를 가지고 있으나, NK1.1과 같은 자연 살세포 특이적 분자도 가지고 있다.[94] CD1d 분자에 제시되는 당지질 항원을 인식한다. 감마인터페론, 인터루킨-4(IL-4), 인터루킨-10(IL-10) 등을 분비하여 면역 반응을 조절하고, 세포독성 작용을 통해 면역 반응에 관여한다.[94]
3. 2. 2. γδ T세포 (감마 델타 T세포)
T세포 수용체(TCR)가 αβ인 다른 T세포와 달리 γδ TCR을 발현하는 T세포 유형이다. NK 세포와 T세포의 성질을 함께 갖는 NKT 세포처럼, 선천 면역과 적응 면역 사이를 넘나드는 특징을 보인다.[92] γδT 세포는 TCR 유전자를 재배열하여 수용체의 다양성을 확보하고 기억 표현형을 발달시킬 수 있다는 점에서 적응 면역의 요소이지만, 제한된 TCR이나 NK 수용체가 패턴 인지에 사용될 수 있다는 점에서는 선천 면역계의 일부분으로 여겨지기도 한다.[92]
γδT 세포가 반응하는 조건은 완전히 밝혀지지 않았지만, 일부는 다음과 같이 알려져 있다.
3. 2. 3. 점막 관련 불변 T세포 (MAIT세포)
MR1 분자에 제시되는 세균 유래 비타민 B 대사 산물을 인식하는 T세포이다. 선천 면역과 유사한 특성을 가지며, 점막 면역에 중요한 역할을 한다.
4. 분화 및 성숙
모든 T 세포는 골수에 존재하는 조혈모세포에서 유래한다.[5] 조혈모세포는 다능선조세포, 초기 림프계 선조세포, 초기 T세포계 선조세포로 분화한다. 이 선조세포는 가슴샘(흉선)으로 이동하여 성숙한 T세포로 분화된다.
골수에서 흉선으로 이동한 T세포 전구체는 흉선 세포(thymocyte)가 되며, 이후 여러 단계를 거쳐 성숙한다. 흉선에 도착한 초기 세포는 CD4와 CD8 보조 수용체를 모두 발현하지 않아 '이중 음성' 세포라고 불린다. 이중 음성 단계에서 흉선 세포는 CD2, CD5, CD7을 발현하고, CD34 발현은 중단되며 CD1이 발현된다. 이후 T 세포 수용체 (TCR) 유전자 재배열을 통해 '이중 양성' 세포가 된다. 이중 양성 흉선 세포는 자신의 MHC 분자와 결합할 수 있는 능력을 평가하는 양성 선택과 자신의 항원에 강하게 반응하는 세포를 제거하는 음성 선택을 거친다.
양성 선택과 음성 선택을 모두 통과한 흉선 세포는 CD4 또는 CD8 중 하나만 발현하는 '단일 양성' 세포로 성숙한다. 이후 성숙한 나이브 T 세포는 체순환에 들어가 이차 림프 조직에서 활성화되어 이펙터 T 세포가 된다.
4. 1. 흉선에서의 성숙 과정
조혈모세포에서 유래한 T세포 전구체는 흉선으로 이동하여 흉선 세포(thymocyte)가 된다. 흉선에 도착한 초기 세포는 CD4와 CD8 보조 수용체를 모두 발현하지 않아 '이중 음성'(double negative) 세포라고 불린다. 이 세포들은 CD4-CD8-CD44+CD25-ckit+ 세포이며 초기 흉선 전구 세포(ETP)라고 불린다.[6]이중 음성 단계에서 흉선 세포는 CD2, CD5, CD7을 발현하고, CD34 발현은 중단되며 CD1이 발현된다. 이후 T 세포 수용체 (TCR) 유전자 재배열이 시작된다. DN2 단계(CD44+CD25+)에서 세포는 재조합 유전자 RAG1, RAG2를 상향 조절하고 TCRβ 유전좌를 재배열하여 기능적인 TCRβ 사슬을 생성한다. DN3 단계(CD44-CD25+)에서는 TCRβ 유전자와 함께 pre-Tα라고 하는 불변 α 사슬을 발현한다. 재배열된 β 사슬이 불변 α 사슬과 성공적으로 쌍을 이루면 β 사슬 재배열이 중단되고(대립 유전자 침묵) 신호가 생성된다.[7] 이 신호는 세포 표면의 pre-TCR을 필요로 하지만, pre-TCR에 대한 리간드 결합과는 독립적이다. 사슬이 성공적으로 쌍을 이루면 pre-TCR이 형성되고 세포는 CD25를 하향 조절하여 DN4 세포(CD25-CD44-)가 된다. 이 세포들은 증식 과정을 거치고, '이중 양성' 단계에서 TCRα 유전좌를 재배열하기 시작한다.
CD4와 CD8을 모두 발현하면 '이중 양성'(double positive) 세포가 된다. 이중 양성 흉선 세포는 자신의 MHC 분자와 결합할 수 있는 능력을 평가하는 양성 선택을 거친다. 자신의 MHC 분자와 결합할 수 있는 흉선 세포는 생존하고, 그렇지 못한 세포는 세포자멸사로 제거된다.
양성 선택 후, 흉선 세포는 자신의 항원에 강하게 반응하는 세포를 제거하는 음성 선택을 거친다. 이는 자가면역 질환을 방지하기 위한 과정이다. 흉선 내 수지상 세포나 대식세포가 제시하는 자가 단백질과 강하게 반응하는 흉선 세포는 세포자멸사로 제거된다.
양성 선택과 음성 선택을 모두 통과한 흉선 세포는 CD4 또는 CD8 중 하나만 발현하는 '단일 양성'(single positive) 세포로 성숙한다. 이후 성숙한 나이브 T 세포는 체순환에 들어가 이차 림프 조직에서 활성화되어 이펙터 T 세포가 된다.
5. 활성화
항원전달세포에 제시된 MHC:항원 복합체를 만나면 T세포 항원 수용체 신호 전달 과정(T-cell receptor signaling pathway)을 통해 항원을 인식하고 효과 T세포로 활성화되어 적응 면역이 시작된다.[29] T세포 활성화는 미접촉 T세포가 MHC:항원 복합체를 만나면서 이루어진다. 미접촉 T세포의 T세포 항원 수용체가 MHC:항원 복합체와 결합하게 되면 T세포의 CD45 단백질이 세포 내부의 Fyn 단백질을 탈인산화한다.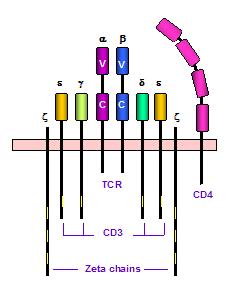
탈인산화되어 활성화된 Fyn은 T세포 항원 수용체의 보조 단백질 사슬인 CD3 감마(γ), 델타(δ), 엡실론(ε) 및 제타(ζ) 사슬의 아이탬(ITAM, Immunoreceptor Tyrosine-based Activation Motif)을 인산화한다. 아이탬이 인산화된 CD3 단백질의 제타 사슬에는 ZAP-70 단백질이 부착될 수 있게 되는데, 그 덕분에 T세포의 보조 수용체인 CD4 또는 CD8 단백질에 부착되어 있는 Lck 단백질이 ZAP-70 단백질을 인산화할 수 있게 된다. Lck 단백질에 의해 인산화되어 활성을 띠게 된 ZAP-70 단백질은 LAT 단백질과 SLP-76 단백질을 인산화하여 NFκB, AP-1 및 NFAT 등의 전사 인자를 활성화하는 신호 반응을 촉발시킨다. NFκB, AP-1 및 NFAT와 같은 전사 인자들은 인터루킨-2(IL-2)의 발현을 촉진함으로써 T세포가 분열 및 분화할 수 있게 한다.
6. 임상적 의의
T 세포 결핍은 T 세포 림프구 감소증이나 개별 T 세포 기능 결함으로 발생한다. T 세포 기능이 완전히 부전되면 중증 복합 면역 결핍증(SCID) 같은 유전 질환이 생길 수 있고, 부분적 부전은 후천성 면역 결핍 증후군(AIDS) 등의 질환을 유발할 수 있다.[57] T 세포 결핍 시에는 단순 헤르페스 바이러스, 결핵균 같은 세포 내 병원체나 진균 감염에 취약해진다.[58][1]
T 세포 암은 T세포 림프종이라고 불리며, 비호지킨 림프종의 약 10%를 차지한다.[59] 주요 형태로는 결절외 T세포 림프종, 피부 T세포 림프종, 역형성 대형 세포 림프종, 혈관면역모구 T세포 림프종 등이 있다.
T 세포 고갈은 만성 감염이나 암과 같은 상황에서 T 세포 기능이 저하되는 상태이다.[60][61] 초기에는 IL-2와 TNFα 생성 능력을 상실하고, 이후 증식 능력과 세포 독성 잠재력을 잃게 되며, 결국에는 삭제(deletion)에 이른다. 만성 감염, 패혈증, 암 등에서 발생할 수 있다.[62]
Treg 세포는 다른 T 세포의 활성을 억제하는데, 이 세포의 기능 이상은 자가 면역 질환과 관련이 있다.[31] 또한 T 세포는 이식된 장기에 대한 면역 반응을 일으켜 이식 거부 반응을 유발할 수 있다. 장기 이식 과정에서 지속적인 항원 노출은 T 세포 고갈을 유발할 수 있으며, 이는 이식편 관용에 중요한 역할을 한다.[77]
6. 1. T세포 결핍
T 세포 결핍은 T 세포의 림프구 감소증 및/또는 개별 T 세포 기능의 결함 때문에 발생한다. T 세포 기능이 완전히 부전되면 중증 복합 면역 결핍증(SCID), 오멘 증후군, 연골-모발 형성 저하증과 같은 유전 질환이 생길 수 있다.[57] T 세포 기능의 부분적 부전은 후천성 면역 결핍 증후군(AIDS), 디조지 증후군(DGS), 염색체 파괴 증후군(CBS), 운동 실조증-모세 혈관 확장증(AT), 위스콧-앨드리치 증후군(WAS)과 같은 B 세포 및 T 세포 결합 질환을 포함한 유전 질환 때문에 발생한다.[57]T 세포 결핍 시 주요 관심 병원체는 ''단순 헤르페스 바이러스'', ''결핵균'', ''리스테리아''와 같은 세포 내 병원체이다.[58] 진균 감염 또한 T 세포 결핍에서 더 흔하고 심각하게 나타난다.[1]
6. 2. T세포 암
T세포의 암은 T세포 림프종이라고 하며, 비호지킨 림프종의 10% 정도를 차지한다.[59] T세포 림프종의 주요 형태는 다음과 같다.- 결절외 T세포 림프종
- 피부 T세포 림프종: 세자리 증후군 및 균상 식육종
- 역형성 대형 세포 림프종
- 혈관면역모구 T세포 림프종
6. 3. T세포 고갈 (T cell exhaustion)
T세포 고갈은 만성 감염이나 암과 같은 상황에서 T세포의 기능이 저하되는 상태를 의미한다.[60][61] T세포 고갈은 명확하게 정의되지는 않았지만, 일반적으로 세 가지 접근 방식으로 정의된다.[60] 첫째는 기능 장애(일반적으로 예상되는 이펙터 반응 부재)를 보이는 세포, 둘째는 특정 원인(주로 항원에 대한 만성 노출)에 의해 생성된 세포, 셋째는 동일한 분자 마커(일반적으로 프로그램된 세포 사멸 단백질 1 [PD-1])를 보이는 세포를 고갈된 것으로 정의한다.[60]기능이 저하된 T세포는 기능이 점진적으로 손실되고, 전사 프로필이 변화하며, 억제성 수용체 발현이 지속되는 특징을 보인다. 초기에는 IL-2와 TNFα 생성 능력을 상실하고, 이후에는 높은 증식 능력과 세포 독성 잠재력을 잃게 되며, 결국에는 삭제(deletion)에 이르게 된다. 고갈된 T세포는 일반적으로 CD43, CD69 수치가 높고 억제성 수용체의 발현이 증가하는 반면, CD62L과 CD127의 발현은 감소한다. 이러한 고갈 현상은 만성 감염, 패혈증, 암 등에서 발생할 수 있다.[62] 고갈된 T세포는 반복적인 항원 노출 후에도 기능적 고갈 상태를 유지한다.[63]
T세포 고갈은 지속적인 항원 노출, CD4 T 세포 도움 부족 등 여러 요인에 의해 유발될 수 있다.[64] 항원 노출 시간과 바이러스 부하(viral load)는 T세포 고갈의 심각성에 영향을 미치는데, 최소 2~4주 이상의 지속적인 노출이 고갈을 유발하는 데 필요한 것으로 알려져 있다.[65] PD-1, CTLA-4, T 세포 막 단백질-3(TIM3), 림프구 활성화 유전자 3 단백질(LAG3)과 같은 억제성 수용체도 T세포 고갈을 유도할 수 있다.[66][67] IL-10, TGF-β와 같은 가용성 분자 역시 T세포 고갈을 유발할 수 있다.[68][69] Treg 세포는 IL-10 및 TGF-β의 공급원이 될 수 있어 T세포 고갈에 기여할 수 있으며,[70] Treg 세포 고갈 및 PD1 차단 후 T세포 고갈이 회복될 수 있다는 연구 결과도 있다.[71]
패혈증의 경우, 사이토카인 폭풍의 결과로 T세포 고갈이 발생할 수 있다. 초기 패혈증 발생 후, 항염증성 사이토카인과 세포 사멸 촉진 단백질이 과도하게 발현되어 신체를 보호하려고 하지만, 높은 항원 부하와 염증으로 인해 T세포 고갈이 증가한다.[72][73] 현재 패혈증 치료에 억제 수용체 차단을 활용하는 연구가 진행 중이다.[74][75][76]
장기 이식 과정에서 지속적인 항원 노출은 T 세포 고갈을 유발할 수 있는데, 이는 이식 후 동종 항원(allogeneic antigen)이 존재하는 상황과 유사하다.[77] 신장 이식 후 시간이 지남에 따라 T 세포 반응이 감소하는 현상이 관찰되었으며,[78] 이는 T 세포 고갈이 동종 반응성 CD8 T 세포의 고갈을 통해 이식편 관용(transplant tolerance)에 중요한 역할을 한다는 것을 시사한다.[73][79] 만성 감염이 이식편 수용과 장기 생존에 긍정적인 영향을 미치며, 이는 부분적으로 T 세포 고갈에 의해 매개된다는 연구 결과도 있다.[80][81][82] 또한, 수혜자의 T 세포 고갈이 NK 세포 이식에 충분한 조건을 제공한다는 사실도 밝혀졌다.[83] 그러나 T 세포 고갈 유도는 감염 증가와 종양 발생 위험을 증가시킬 수 있다는 단점도 존재한다.[84]
암 발생 과정에서 T 세포 고갈은 종양 방어 기전을 약화시킨다. 일부 암 관련 세포와 종양 세포 자체가 종양 부위에서 T 세포 고갈을 유도할 수 있다는 연구 결과가 있다.[85][86][87] T 세포 고갈은 백혈병 재발에도 영향을 미칠 수 있으며,[88] 일부 연구에서는 T 세포의 억제 수용체 PD-1 및 TIM-3 발현을 통해 백혈병 재발을 예측할 수 있다고 제안한다.[89] 암 치료 분야에서는 면역 관문 억제제(immune checkpoint inhibitor)에 대한 연구가 활발히 진행되고 있으며, 일부는 이미 임상에서 사용되는 효과적인 치료법으로 승인되었다.[90] 이러한 치료법은 T 세포 고갈에 중요한 억제 수용체를 표적으로 하여, 억제 수용체 차단을 통해 T 세포 고갈을 회복시키는 기전을 이용한다.[91]
6. 4. 기타 질환
조절 T 세포(Treg)는 흉선에서 분화하며 CD4, CD25, Foxp3 분자를 발현하여 다른 T 세포의 활성을 억제한다. 말초에서 항원 특이적으로 유도되거나, CD8 양성 T 세포에서 분화하는 조절 T 세포도 있다. 이러한 조절 T 세포의 기능 이상은 자가 면역 질환과 관련이 있으며, 자가 반응성 T 세포의 활성화를 유발할 수 있다.[31]T 세포는 이식된 장기에 대한 면역 반응을 일으켜 이식 거부 반응을 유발할 수 있다. 이식 과정에서 T 세포 고갈은 지속적인 항원 노출 후에 발생할 수 있는데, 이는 이식편 이식 후 동종 항원 존재와 유사한 상황이다.[77] 신장 이식 후 T 세포 반응이 시간이 지남에 따라 감소하는 것으로 나타났다.[78] 이는 T 세포 고갈이 주로 동종 반응성 CD8 T 세포의 고갈을 통해 이식편 관용에 중요한 역할을 한다는 것을 시사한다.[73][79] 여러 연구에서 만성 감염이 이식편 수용과 장기 생존에 긍정적인 영향을 미치는 것이 부분적으로 T 세포 고갈에 의해 매개된다는 것을 보여주었다.[80][81][82] 또한 수혜자 T 세포 고갈이 NK 세포 이식에 충분한 조건을 제공한다는 것도 밝혀졌다.[83] T 세포 고갈 유도가 이식에 유익할 수 있지만, 감염 증가와 종양 발생 위험을 포함한 단점도 가지고 있다.[84]
참조
[1]
웹사이트
5. Hematopoietic Stem Cells
https://stemcells.ni[...]
National Institutes of Health, U.S. Department of Health and Human Services
2001-06-17
[2]
서적
Molecular Biology of the Cell
Garland Science
2002
[3]
논문
CD4⁺T cells: differentiation and functions
2012
[4]
서적
Molecular Biology of the Cell
Garland Science
2002
[5]
논문
One Niche to Rule Both Maintenance and Loss of Stemness in HSCs
2016-12
[6]
논문
Impaired CD8 T cell memory and CD4 T cell primary responses in IL-7R alpha mutant mice
2007-03
[7]
서적
Janeway's Immunobiology
Garland Science
[8]
논문
Distinct phases in the positive selection of CD8+ T cells distinguished by intrathymic migration and T-cell receptor signaling patterns
2014-06
[9]
논문
On the role of self-recognition in T cell responses to foreign antigen
https://onlinelibrar[...]
2003-02
[10]
논문
Positive and negative selection of T cells
2003-01-01
[11]
논문
The MHC reactivity of the T cell repertoire prior to positive and negative selection
1997-03
[12]
논문
Autonomous role of medullary thymic epithelial cells in central CD4(+) T cell tolerance
https://hal.archives[...]
2010-06
[13]
논문
Neonatal and adult recent thymic emigrants produce IL-8 and express complement receptors CR1 and CR2
2017-08
[14]
서적
Janeway's Immunobiology
Garland Science
[15]
논문
The role of the thymus in immune reconstitution in aging, bone marrow transplantation, and HIV-1 infection
[16]
논문
APC-derived cytokines and T cell polarization in autoimmune inflammation
[17]
논문
T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells
2020-08-18
[18]
논문
Helper T cell differentiation
2019-07
[19]
서적
T Helper Cell Differentiation and Their Function
[20]
논문
Two subsets of memory T lymphocytes with distinct homing potentials and effector functions.
[21]
논문
Loss of CD45R and gain of UCHL1 reactivity is a feature of primed T cells
1988-04
[22]
논문
Molecular signatures distinguish human central memory from effector memory CD8 T cell subsets.
https://www.pure.ed.[...]
[23]
논문
Multiparameter flow cytometric analysis of CD4 and CD8 T cell subsets in young and old people.
[24]
논문
Tissue-resident memory T cells
2013-09
[25]
논문
Alternative memory in the CD8 T cell lineage.
[26]
논문
CD4+ virtual memory: Antigen-inexperienced T cells reside in the naïve, regulatory, and memory T cell compartments at similar frequencies, implications for autoimmunity.
[27]
논문
Regulatory T cells: recommendations to simplify the nomenclature
http://www.escholars[...]
[28]
논문
Modulation of autoimmune diseases by interleukin (IL)-17 producing regulatory T helper (Th17) cells
[29]
논문
The burgeoning family of unconventional T cells
2015-11
[30]
논문
Translating Unconventional T Cells and Their Roles in Leukemia Antitumor Immunity
2021
[31]
논문
Invariant and noninvariant natural killer T cells exert opposite regulatory functions on the immune response during murine schistosomiasis
2007-05
[32]
논문
The Role of Mucosal Associated Invariant T Cells in Antimicrobial Immunity
2015-07-06
[33]
논문
Mucosal associated invariant T cells and the immune response to infection
2011-08
[34]
논문
Recognition of Vitamin B Precursors and Byproducts by Mucosal Associated Invariant T Cells
2015-12
[35]
논문
Mucosal-associated invariant T-cells: new players in anti-bacterial immunity
2014-10-08
[36]
논문
MR1-Restricted Mucosal-Associated Invariant T Cells and Their Activation during Infectious Diseases
2015-06-16
[37]
논문
Mucosal-associated invariant T cells in autoimmunity, immune-mediated diseases and airways disease
2016-05
[38]
논문
Invariant natural killer T cells and mucosal-associated invariant T cells in multiple sclerosis
2017-03
[39]
논문
Innate mucosal-associated invariant T (MAIT) cells are activated in inflammatory bowel diseases
2014-05
[40]
논문
MR1 antigen presentation to mucosal-associated invariant T cells was highly conserved in evolution
2009-05
[41]
논문
Bacteria, mucosal-associated invariant T cells and MR1
2010-11
[42]
논문
MR1 presents microbial vitamin B metabolites to MAIT cells
https://espace.libra[...]
2012-11
[43]
간행물
Understanding the Immune System (pdf)
https://www.niaid.ni[...]
NIAID
[44]
논문
Effector and memory CTL differentiation
2007-01-01
[45]
논문
CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes
2003-02
[46]
논문
Requirement for CD4 T cell help in generating functional CD8 T cell memory
2003-04
[47]
논문
CD4+T cells are required for the maintenance, not programming, of memory CD8+ T cells after acute infection
2004-09
[48]
문서
Turning off the T cells: Peptides for treatment of allergic Diseases
Today's life science publishing
1999
[49]
논문
Glycans in the immune system and The Altered Glycan Theory of Autoimmunity
[50]
논문
TRAF6, a molecular bridge spanning adaptive immunity, innate immunity and osteoimmunology
2003-11
[51]
서적
Signal transduction
Elsevier Academic Press
[52]
논문
CTLs respond with activation and granule secretion when serving as targets for T cell recognition
2011-01
[53]
뉴스
SpaceX ready for CRS-3 Dragon launch and new milestones
http://www.nasaspace[...]
2014-04-14
[54]
논문
T cells and reactive oxygen species
2015-10
[55]
논문
Quantitative challenges in understanding ligand discrimination by alphabeta T cells
[56]
논문
An induced rebinding model of antigen discrimination
[57]
EMedicine
T-Cell Disorders
[58]
서적
Infection: Microbiology and Management
https://books.google[...]
Wiley-Blackwell
[59]
웹사이트
The Lymphomas
http://www.leukemia-[...]
The Leukemia & Lymphoma Society
2006-05
[60]
논문
Immunological exhaustion: How to make a disparate concept operational?
2021-09
[61]
논문
Defining 'T cell exhaustion'
2019-11
[62]
논문
T-cell exhaustion: characteristics, causes and conversion
2010-04
[63]
논문
Hepatitis B Virus-Specific CD8+ T Cells Maintain Functional Exhaustion after Antigen Reexposure in an Acute Activation Immune Environment
2018
[64]
논문
CD4+ T cells are required to sustain CD8+ cytotoxic T-cell responses during chronic viral infection
1994-12
[65]
논문
Progressive loss of memory T cell potential and commitment to exhaustion during chronic viral infection
2012-08
[66]
논문
T cell exhaustion
2011-06
[67]
논문
+ T cells during bovine leukemia virus infection
2018-06
[68]
논문
Interleukin-10 determines viral clearance or persistence in vivo
2006-11
[69]
논문
Cell-intrinsic transforming growth factor-beta signaling mediates virus-specific CD8+ T cell deletion and viral persistence in vivo
2009-07
[70]
논문
Role of regulatory T cells during virus infection
2013-09
[71]
논문
Interplay between regulatory T cells and PD-1 in modulating T cell exhaustion and viral control during chronic LCMV infection
2014-08
[72]
논문
The late phase of sepsis is characterized by an increased microbiological burden and death rate
2011-07
[73]
논문
Immunosuppression in patients who die of sepsis and multiple organ failure
2011-12
[74]
논문
Anti-PD-L1 peptide improves survival in sepsis
2017-02
[75]
논문
Frontline Science: Defects in immune function in patients with sepsis are associated with PD-1 or PD-L1 expression and can be restored by antibodies targeting PD-1 or PD-L1
2016-12
[76]
논문
Alpha-lactose reverses liver injury via blockade of Tim-3-mediated CD8 apoptosis in sepsis
2018-07
[77]
논문
The role of peripheral T-cell deletion in transplantation tolerance
2001-05
[78]
논문
Disappearance of T Cell-Mediated Rejection Despite Continued Antibody-Mediated Rejection in Late Kidney Transplant Recipients
2015-07
[79]
논문
Exhaustive differentiation of alloreactive CD8+ T cells: critical for determination of graft acceptance or rejection
http://pure-oai.bham[...]
2008-05
[80]
논문
Cytomegalovirus-Induced Expression of CD244 after Liver Transplantation Is Associated with CD8+ T Cell Hyporesponsiveness to Alloantigen
2015-08
[81]
논문
IL-10 Induces T Cell Exhaustion During Transplantation of Virus Infected Hearts
2016
[82]
논문
CMV Primary Infection Is Associated With Donor-Specific T Cell Hyporesponsiveness and Fewer Late Acute Rejections After Liver Transplantation
2015-09
[83]
논문
Recipient T Cell Exhaustion and Successful Adoptive Transfer of Haploidentical Natural Killer Cells
2018-03
[84]
논문
Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate T-cell function to promote tumoral immune escape
2012-02
[85]
논문
T cells in multiple myeloma display features of exhaustion and senescence at the tumor site
2016-11
[86]
논문
Cancer-associated fibroblasts induce antigen-specific deletion of CD8 + T Cells to protect tumour cells
2018-03
[87]
논문
The ion channel network in T lymphocytes, a target for immunotherapy
2012-02
[88]
논문
T cell exhaustion characterized by compromised MHC class I and II restricted cytotoxic activity associates with acute B lymphoblastic leukemia relapse after allogeneic hematopoietic stem cell transplantation
2018-05
[89]
논문
PD-1(hi)TIM-3(+) T cells associate with and predict leukemia relapse in AML patients post allogeneic stem cell transplantation
2015-07
[90]
뉴스
U.S. FDA Approved Immune-Checkpoint Inhibitors and Immunotherapies
https://medi-paper.c[...]
Medical Writer Agency {{!}} 香港醫學作家 {{!}} MediPR {{!}} MediPaper Hong Kong
2018-08-21
[91]
논문
Control of Toxoplasma reactivation by rescue of dysfunctional CD8+ T-cell response via PD-1-PDL-1 blockade
2011-05
[92]
논문
Immunosurveillance and immunoregulation by γδ T cells
[93]
논문
γδ T cells link innate and adaptive immune responses
[94]
서적
Janeway’s immunobiology
https://archive.org/[...]
Garland Science
[95]
저널
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com