보조 T세포
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
보조 T 세포(Th 세포)는 면역 세포를 돕는 사이토카인을 생성하고 방출하는 면역 세포의 한 유형이다. CD4 표면 단백질을 발현하는 성숙한 Th 세포는 CD4+ T 세포로 불리며, 항원 제시 세포가 펩타이드 항원을 MHC 클래스 II 단백질에 제시할 때 세포 간 상호 작용과 사이토카인을 통해 해당 세포를 돕는다. Th 세포는 활성화, 생존 및 분화 신호를 통해 활성화되며, Th1, Th2, Th17, THαβ 세포 등 다양한 아형으로 분화하여 면역 반응을 조절한다. 이러한 Th 세포의 불균형은 알레르기, 자가면역 질환, HIV 감염, COVID-19 등 다양한 질병의 발병과 관련이 있다.
Th 세포는 사이토카인을 포함하고 방출하여 다른 면역 세포를 돕는 역할을 한다. 사이토카인은 해당 사이토카인에 대한 수용체를 가진 표적 세포의 행동을 변화시키는 작은 단백질 매개체이다. Th 세포는 면역학적 손상의 종류(예: 바이러스, 세균, 세포 내 세균, 기생충, 곰팡이, 원생생물 등)에 따라 면역 반응의 방향을 결정한다.
보조 T세포(Th 세포)는 다른 면역 세포의 활동을 조절하고 돕기 위해 사이토카인이라는 작은 단백질 신호 물질을 분비한다. 이 사이토카인은 특정 수용체를 가진 표적 세포에 작용하여 그 행동을 변화시킨다. Th 세포는 침입한 병원체의 종류(예: 바이러스 대 세균 대 기생충 등)에 따라 면역 반응의 방향을 결정하는 중요한 역할을 한다.
보조 T 세포는 다양한 면역 세포에 영향을 미치며, 어떤 사이토카인과 같은 세포외 신호를 생성할지 결정함으로써 면역 반응의 성격을 조절한다. 이는 감염에 성공적으로 대처하기 위해 숙주에게 가장 유익한 면역 체계를 구축하는 데 필수적이다.[18] 보조 T 세포가 면역 반응을 어떻게 조절하는지 정확히 이해하는 것은 현재 면역학에서 중요한 연구 주제인데, 이는 질병 치료법 개발이나 백신의 효과를 높이는 데 크게 기여할 수 있기 때문이다.
2. 구조 및 기능
성숙한 Th 세포는 표면에 CD4 단백질을 발현하며, 이 때문에 '''CD4+ T 세포'''라고도 불린다. CD4+ T 세포는 일반적으로 면역 체계 내에서 보조 T 세포로서 특정 역할을 수행하는 것으로 여겨진다. 예를 들어, 항원 제시 세포가 펩타이드 항원을 MHC 클래스 II 분자에 실어 제시하면, CD4+ 세포는 세포 간 상호작용(예: CD40 (단백질)과 CD40L의 결합)과 사이토카인 분비를 통해 해당 항원 제시 세포를 돕는다.
Th 세포는 기능이나 다른 세포와의 상호작용 방식이 다양하여 하나의 통일된 집단으로 보기 어렵다. 일반적으로 아직 분화되지 않은 성숙한 T 세포가 전문적인 항원 제시 세포에 의해 자극을 받으면 특정 기능을 수행하는 이펙터 세포로 분화한다. 이러한 분화는 특정 '계통 결정 전사 인자'(때로는 마스터 조절자라고도 불리지만, 이 용어는 지나치게 단순화되었다는 비판도 있다)의 발현 여부에 따라 결정된다.[2] 만약 특정 계통 결정 전사 인자가 없다면 해당 종류의 보조 T 세포가 만들어지지 않아 숙주의 건강에 심각한 영향을 미칠 수 있다.
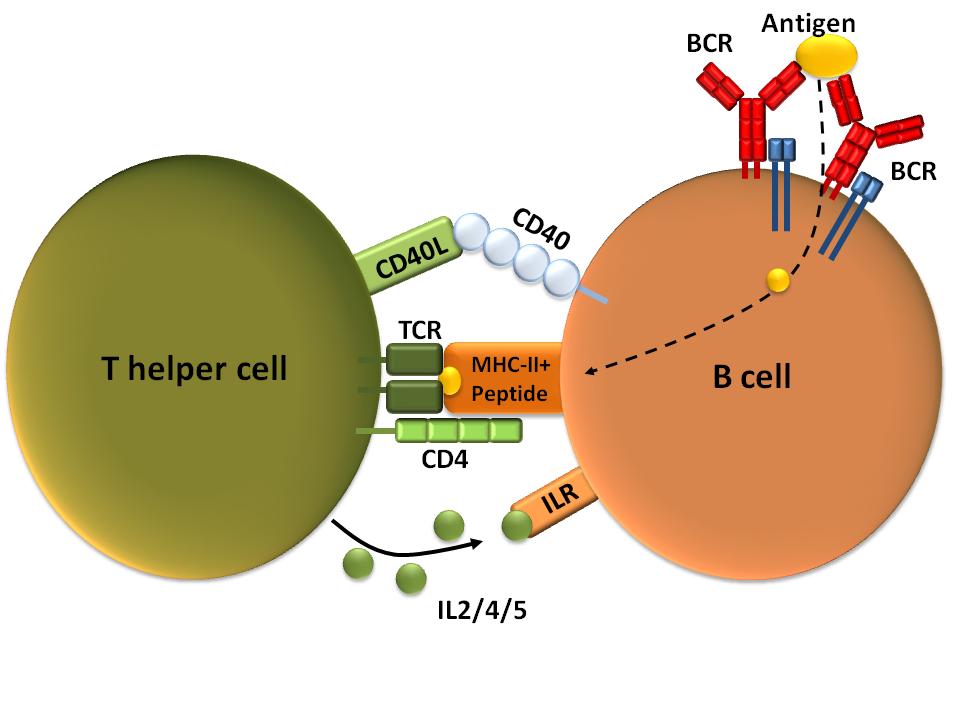
Th2 세포는 사이토카인 방출을 통해 액성 면역 반응을 활성화하는 주요 기능을 담당한다. 액성 면역은 항체 분자가 중요한 역할을 하는 면역 반응으로, 항체 단백질은 활성화된 B 세포에 의해 생산된다. B 세포는 표면에 B 세포 항원 수용체(BCR)를 가지고 있으며, 외부 항원을 세포 안으로 받아들여 분해한 뒤, 그 조각을 MHC 클래스 II 분자를 통해 세포 표면에 제시한다. Th2 세포는 자신의 T 세포 항원 수용체(TCR)를 이용해 B 세포가 제시한 항원 조각을 인식한다. 이후 Th2 세포는 표면에 CD40 리간드를 발현시키고 사이토카인을 방출하여 B 세포의 분화와 증식을 촉진한다. Th2 세포가 방출하는 사이토카인(Th2 사이토카인)에는 IL-4, IL-5, IL-6, IL-10, IL-13, GM-CSF 등이 있다. 특히 IL-4 자극은 B 세포에서 면역글로불린 E(IgE) 생산을 크게 늘린다. IgE는 비만 세포나 호염기구 표면에 결합하여 히스타민 방출을 촉진하는 등 중요한 역할을 한다. 또한, Th2 세포가 생산하는 IL-10 및 TGF-β는 Th1 세포의 반응을 억제하는 기능도 가지고 있다.
Th1 세포와 Th2 세포는 서로의 기능을 억제하며 균형을 이루지만, 이 균형이 Th2 세포 쪽으로 기울어 Th2 사이토카인이 과도하게 생산되면 기관지 천식이나 아토피 피부염과 같은 알레르기성 질환(소위 Th2 질환)이 발생할 수 있는 것으로 알려져 있다.
3. 헬퍼 T 세포의 활성화
성숙한 Th 세포는 세포 표면에 CD4 단백질을 발현하며, 이 때문에 CD4+ T 세포라고도 불린다. 이 세포들은 주로 항원 제시 세포(APC)가 MHC 클래스 II 분자를 통해 제시하는 외부 항원 조각을 인식한다. CD4+ T 세포는 APC와 직접 접촉하거나(CD40 (단백질)와 CD40L 상호작용 등) 다양한 사이토카인을 분비하여 APC 및 다른 면역 세포들의 활성화를 유도한다.
Th 세포는 기능과 상호작용하는 세포에 따라 매우 다양하다. 일반적으로 흉선에서 발달을 마친 미성숙한 T 세포는 2차 림프 기관(비장, 림프절 등)으로 이동하여 성숙한 미분화 T 세포가 된다.[3] '미분화' 상태는 아직 특정 항원에 노출되어 활성화된 경험이 없음을 의미한다. 이 미분화 T 세포가 전문 항원 제시 세포(주로 수지상 세포, 대식세포, B 세포)로부터 특정 항원 자극을 받으면 활성화 과정을 거쳐 특정 기능을 수행하는 세포로 분화한다. 이 분화 과정은 특정 전사 인자(마스터 조절자라고도 불림)의 발현에 의해 결정되며, 특정 유형의 Th 세포가 부족하면 숙주의 면역 기능에 심각한 문제가 발생할 수 있다.[2]
미분화 T 세포의 활성화는 일반적으로 세 가지 주요 신호가 순차적으로 필요한 '3신호 모델'로 설명된다.[10] 첫 번째 신호는 T 세포의 T 세포 수용체(TCR)가 APC의 MHC 클래스 II 분자에 제시된 특정 항원 조각을 인식하고 결합하면서 시작된다. 이후 두 번째 신호(보조 자극 신호)와 세 번째 신호(사이토카인에 의한 분화 유도 신호)를 통해 T 세포는 완전히 활성화되고 증식하며, 특정 유형의 보조 T세포(예: Th1, Th2 등)로 분화하여 면역 반응을 효과적으로 조절하게 된다. 예를 들어, Th2 세포는 특정 사이토카인들을 분비하여 B 세포가 항체를 생산하도록 유도하는 액성 면역 반응을 촉진한다. Th1 세포와 Th2 세포는 서로의 기능을 억제하며 균형을 이루는데, 이 균형이 Th2 쪽으로 치우치면 기관지 천식이나 아토피 피부염과 같은 알레르기성 질환이 유발될 수 있다.
3. 1. 활성화 (신호 1)

흉선에서 발달을 마친 미성숙 보조 T세포는 2차 림프 기관(비장, 림프절 등)으로 이동하여 성숙한 미분화 T 세포가 된다. 미분화 상태란 아직 특정 항원에 노출되어 반응한 경험이 없음을 의미한다.[4][5] 이 미분화 CD4+ T 세포는 T 세포 수용체(TCR)-CD3 복합체와 CD4 보조 수용체를 세포 표면에 발현한다. TCR의 가변 영역은 T 세포가 인식하고 반응할 수 있는 특정 항원을 결정하며, CD4+ T 세포의 TCR은 II형 MHC 분자에 결합된 항원 조각을 인식하도록 만들어진다.
II형 MHC 분자는 주로 전문 항원 제시 세포(APC)의 표면에서 발견된다. 전문 APC에는 수지상 세포, 대식세포, B 세포 등이 있으며, 이 중 수지상 세포는 항상 II형 MHC를 발현하는 특징이 있다. 면역 반응이 시작되면, 전문 APC(주로 수지상 세포)는 외부에서 침입한 항원(예: 세균이나 바이러스)을 세포내이입하여 잘게 분해하는 항원 처리 과정을 거친다. 이후 APC는 감염 부위에서 가까운 림프절로 이동한다. 항원이 특정 분자 패턴을 가지면(이를 '신호 0'이라고도 함), 수지상 세포는 성숙하여 II형 MHC 발현을 늘리고, T 세포 활성화에 필요한 다른 분자들(보조 자극 분자, 신호 2에서 중요)의 발현도 증가시킨다.[11][12]
림프절에 도착한 APC는 처리된 항원 조각을 II형 MHC 분자에 실어 T 세포에게 보여준다(항원 제시). 미분화 CD4+ T 세포 중 특정 항원 조각-MHC 복합체를 인식할 수 있는 TCR을 가진 세포가 APC와 만나면 활성화 과정의 첫 번째 신호가 시작된다.
Th 세포가 APC 위의 항원-MHC II 복합체를 인식하면, T 세포의 TCR-CD3 복합체가 이 복합체에 강하게 결합한다. 동시에 T 세포의 CD4 보조 수용체도 MHC II 분자의 다른 부위에 결합하여 이 상호작용을 안정화시킨다. 이 결합은 T 세포 활성화의 첫 번째 신호로 작용한다. T 세포 활성화를 위해서는 약 50개의 이러한 상호작용이 필요하며, 충분한 결합이 이루어지면 CD4 분자는 Lck라는 키나아제 효소를 동원한다. Lck는 CD3 복합체의 특정 부위인 면역수용체 티로신 기반 활성 모티프(ITAM)를 인산화시킨다. 인산화된 ITAM에는 또 다른 키나아제인 ZAP-70이 결합하여 활성화되고, 이는 세포 내부로 활성화 신호를 전달하는 연쇄 반응을 일으킨다.[13] Lck의 활성은 CD45와 Csk라는 두 분자에 의해 조절된다. CD45는 Lck를 활성화시키는 역할을 하고, Csk는 Lck를 억제하는 역할을 한다.[13] 기억 T 세포는 미분화 T 세포보다 Lck를 더 많이 가지고 있으며 Csk의 억제 기능이 약화되어 있어 더 빠르게 반응할 수 있다.[14]
항원-MHC와 TCR/CD4의 결합 외에도, T 세포 표면의 인테그린 단백질인 LFA-1과 APC 표면의 ICAM 사이의 결합이 두 세포가 서로 단단히 붙어 있도록 하는 데 중요한 역할을 한다.
한편, CD45 분자는 T 세포의 활성화 상태에 따라 세포 바깥 부분의 크기가 변하는 여러 동형체(isoform)를 가진다. 예를 들어, 미분화 T 세포는 긴 형태의 CD45(CD45RA+)를 가지지만, 활성화되면 짧은 형태(CD45RO+)로 바뀐다. 긴 형태의 CD45RA가 TCR의 접근성을 낮춰 더 강한 결합(높은 친화력)을 요구함으로써 활성화의 문턱을 높일 수 있다는 가설이 있다. 일단 활성화되어 짧은 CD45RO 형태로 바뀌면, T 세포가 다른 세포들과 더 쉽게 상호작용하고 반응할 수 있게 된다.
3. 2. 생존 (신호 2)
첫 번째 신호인 TCR/CD3 신호를 받은 나이브 T 세포는 '신호 2'라고 불리는 두 번째 독립적인 생화학적 경로를 활성화해야 한다. 이 과정은 T 세포가 외부 항원(외래 항원)에 제대로 반응하는지 확인하는 일종의 검증 단계이며, 우리 몸을 보호하기 위한 중요한 조치이다. 만약 처음 항원에 노출되었을 때 이 두 번째 신호가 없다면, 해당 T 세포는 우리 몸의 일부를 공격할 수 있는 위험한 세포(자가 반응성 세포)로 간주된다. 그 결과, 이 T 세포는 무반응 상태에 빠지게 된다. 무반응 상태가 된 세포는 나중에 두 신호가 모두 주어지더라도 어떤 항원에도 반응하지 못하며, 결국 세포자멸사 과정을 통해 제거될 때까지 아무런 기능을 하지 못하고 몸속을 순환하는 것으로 여겨진다.[15]
두 번째 신호는 주로 보조 T 세포(CD4+ T 세포) 표면에 있는 CD28 단백질과 전문 항원제시세포(APC) 표면에 있는 CD80(B7.1) 또는 CD86(B7.2) 단백질 사이의 상호작용을 통해 전달된다. CD80과 CD86은 모두 CD28 수용체를 활성화시키는 역할을 하며, 이들을 공동 자극 분자라고 부르기도 한다.
이러한 검증 단계는 나이브 보조 T 세포를 활성화하는 데 필수적이지만, 그 중요성은 CD8+ 세포 독성 T 세포의 활성화 과정에서 더욱 두드러진다. 나이브 CD8+ T 세포는 특정 항원에 대한 사전 정보가 없기 때문에, CD28 활성화를 통해 자신이 인식한 항원이 실제로 외부 침입자임을 확인해야 한다. (CD80/CD86은 활성화된 APC에서만 발현된다.) 따라서 CD28은 T 세포가 우리 몸의 정상 세포를 공격하는 자가 면역 반응의 위험을 줄이는 데 핵심적인 역할을 한다.
나이브 T 세포가 첫 번째 신호와 두 번째 신호를 모두 성공적으로 받으면, 첫 번째 신호만 받았을 때 유도되는 무반응 상태 대신 세포가 활성화된다. 일단 활성화된 T 세포는 더 이상 두 번째 신호를 필요로 하지 않으며, 이후에는 첫 번째 신호만으로도 다시 활성화될 수 있다. 이는 적응 면역계의 중요한 특징 중 하나인 기억 T 세포의 경우에도 마찬가지이다. 기억 T 세포는 이미 과거에 항원 검증을 마쳤기 때문에, 동일한 항원에 다시 노출되면 훨씬 더 빠르게 반응하여 효과적인 면역 반응을 일으킬 수 있다.
3. 3. 분화 (신호 3)
두 신호 활성화가 완료되면 보조 T세포(Th)는 증식을 시작한다. 이는 강력한 T세포 성장 인자인 인터류킨 2 (IL-2)를 방출하여 자가분비 방식으로 스스로를 자극함으로써 이루어진다. 활성화된 T세포는 또한 IL-2 수용체의 알파 서브 유닛(CD25 또는 IL-2Rα)을 만들어 IL-2에 결합할 수 있는 완전한 수용체를 형성하고, 이는 T세포의 증식 경로를 활성화시킨다.
IL-2의 자가분비 또는 곁분비 작용은 동일한 Th 세포 또는 주변의 다른 Th 세포에 있는 IL-2 수용체에 결합하여 세포 증식과 클론 확장을 유도한다. 활성화 신호와 증식 신호를 모두 받은 Th 세포는 IL-2, IL-4, 인터페론 감마 (IFN-γ)를 분비하는 Th0 (보조 T세포 0) 세포가 된다. Th0 세포는 주변의 사이토카인 환경에 따라 Th1 또는 Th2 세포로 분화한다. IFN-γ는 Th1 세포 생성을 유도하는 반면, IL-10과 IL-4는 Th1 세포 생성을 억제한다. 반대로 IL-4는 Th2 세포 생성을 유도하고, IFN-γ는 Th2 세포 생성을 억제한다. 이러한 사이토카인들은 다면발현성을 가지며 면역 반응의 여러 다른 기능을 수행한다.
Th2 세포는 주로 사이토카인을 방출하여 액성 면역 반응을 활성화하는 역할을 한다. 액성 면역은 항체 분자가 관여하는 면역 체계로, 항체 단백질은 활성화된 B 세포에 의해 생산된다. B 세포 표면에는 B 세포 항원 수용체(BCR)가 있으며, 세포 안으로 항원을 받아들여 분해한 뒤 그 조각을 MHC 클래스 II 분자를 통해 세포 표면에 제시한다. Th2 세포는 T 세포 항원 수용체 (TCR)를 통해 B 세포가 제시한 항원 조각을 인식하고, Th2 세포 표면의 CD40 리간드 발현과 사이토카인 방출을 통해 B 세포를 자극하여 분화와 증식을 촉진한다. Th2 세포가 방출하는 사이토카인은 Th2 사이토카인이라고 하며, IL-4, IL-5, IL-6, IL-10, IL-13, GM-CSF 등이 있다. 특히 IL-4 자극은 B 세포에서 면역글로불린 E (IgE) 생산을 증가시킨다. IgE는 비만 세포나 호염기구 등의 세포 표면에 결합하여 히스타민 방출 촉진 등에 관여하는 중요한 분자이다. 또한, Th2 세포가 생산하는 IL-10 및 TGF-β는 Th1 반응을 억제하는 기능을 한다.
Th1 세포와 Th2 세포는 서로의 기능을 억제하며 균형을 유지하지만, 이 균형이 Th2 세포 쪽으로 기울어져 Th2 사이토카인이 과도하게 생산되면 기관지 천식이나 아토피 피부염과 같은 알레르기성 질환(소위 Th2 질환)이 발생할 수 있다.
1차 림프 조직인 흉선에서 생산된 미경험 T세포(naive T cell)는 말초 림프 조직을 순환하며 항원 침입에 대비한다. 미경험 T세포가 IL-4의 자극을 받으면 전사 인자인 STAT6을 통해 GATA-3의 전사 활성화가 유도된다.[44] GATA-3는 미경험 T세포에서도 어느 정도 발현되며, Th2 세포로 분화하는 과정에서 발현량이 증가하여 전사 인자로 기능한다. GATA-3는 Th2 사이토카인의 발현을 조절하는 동시에, Th1 분화에 관여하는 전사 인자인 STAT4의 발현을 억제하는 것으로 알려져 있다.[45]
4. 이펙터 T 세포 반응의 결정
T 세포의 보조 기능에 대한 분자적 이해는 1991년 세 연구 그룹이 CD154(CD40L)를 발견하면서 크게 진전되었다. CD154는 B 세포 표면의 CD40과 상호작용하여 B 세포 활성화에 중요한 역할을 하는 분자이다. 각 그룹의 주요 발견 내용은 다음과 같다.연구자 소속 주요 방법 발견 내용 세스 레더만 컬럼비아 대학교 의과대학 마우스 단일클론 항체 5c8 사용 인간 CD4+ T 세포에서 일시적으로 발현되는 32kDa 표면 단백질(CD154) 특징화, 접촉 의존적 T 세포 보조 기능 억제 확인 리처드 아미티지 암젠 CD40-Ig를 이용한 발현 라이브러리 스크리닝 CD154를 인코딩하는 cDNA 클론 랜돌프 노엘 다트머스 의과대학 마우스 T 세포의 39kDa 단백질에 결합하는 항체 생성 해당 항체가 T 세포 보조 기능을 억제함을 확인
4. 1. Th1/Th2 모델
증식하는 보조 T 세포는 이펙터 T 세포로 발달하여 Th1과 Th2 세포(각각 제1형 및 제2형 보조 T 세포)라는 두 가지 주요 아형으로 분화된다.
Th1 보조 세포는 주로 세포 내 세균이나 원생동물에 대항하여 세포 매개 면역 반응(주로 대식세포와 세포 독성 T 세포에 의해)을 강화시키는 역할을 한다.[19] 이 과정은 분극 사이토카인인 IL-12에 의해 시작되며, 주요 이펙터 사이토카인은 IFN-γ와 IL-2이다. Th1 면역 반응의 핵심적인 역할을 하는 세포는 대식세포, CD8 T 세포, IgG 항체를 생산하는 B 세포, 그리고 IFN-γ를 분비하는 CD4 T 세포이다. 주요 Th1 전사 인자로는 STAT4와 T-bet이 있다. CD4 T 세포에서 분비된 IFN-γ는 대식세포를 활성화시켜 세포 내 세균 및 원생동물을 식세포 작용으로 삼키고 소화하게 한다. 또한 IFN-γ는 유도성 산화 질소 합성 효소(iNOS)를 활성화시켜 산화 질소 자유 라디칼을 생성하게 하며, 이는 세포 내 세균 및 원생동물을 직접 사멸시킬 수 있다. 만약 Th1 세포가 자기 항원에 대해 과도하게 활성화되면 제IV형 과민 반응 또는 지연형 과민 반응을 일으킬 수 있다. 만투 반응과 제1형 당뇨병이 이러한 자가면역 질환의 예이다.[20]
Th2 보조 세포는 주로 선충과 같은 세포 외 기생충에 대항하여 체액성 면역 반응을 유도한다.[19] 이 분화는 분극 사이토카인인 IL-4와 IL-2에 의해 촉진되며, 주요 이펙터 사이토카인으로는 IL-4, IL-5, IL-9, IL-10, IL-13, IL-25 등이 있다. 주요 이펙터 세포는 호산구, 호염기구, 비만 세포뿐만 아니라 B 세포 및 IL-4/IL-5를 분비하는 CD4 T 세포이다. 주요 Th2 전사 인자로는 STAT6와 GATA3가 있다.[21] IL-4는 Th2 세포 분화를 촉진하는 양성 피드백 역할을 한다. 또한 IL-4는 B 세포를 자극하여 IgE 항체를 생산하도록 유도하는데, 이 IgE는 비만 세포를 자극하여 히스타민, 세로토닌, 류코트리엔 등을 방출하게 한다. 이러한 물질들은 기관지 수축, 장 연동 운동 촉진, 위액 분비 증가 등을 유발하여 기생충을 몸 밖으로 배출하는 데 도움을 준다. CD4 T 세포에서 분비되는 IL-5는 호산구를 활성화시켜 기생충을 공격하게 한다. IL-10은 Th1 세포의 분화와 수지상 세포의 기능을 억제한다. 항원에 대한 Th2 반응이 과도해지면 IgE가 매개하는 알레르기 반응인 제I형 과민 반응이 발생한다. 알레르기성 비염, 아토피성 피부염, 천식 등이 이러한 과민 반응에 해당한다.[20] 또한, Th2 세포는 서로 다른 사이토카인을 발현할 뿐만 아니라 세포 표면의 글리칸(올리고당) 구조에서도 Th1 세포와 차이를 보여, 일부 세포 사멸 유도 물질에 덜 민감한 특성을 보인다.[22][23]
| 제1형/ Th1 | 제2형/ Th2[20] | |
|---|---|---|
| 주요 파트너 세포 유형 | 대식세포, CD8+ T 세포 | B 세포, 호산구, 비만 세포 |
| 생성되는 사이토카인 | IFN-γ 및 TNF-β. 활성화된 Th1 세포에서 IL-2 및 IL-10 생성도 보고됨.[25] | IL-4, IL-5, IL-6, IL-9, IL-10, IL-13 |
| 촉진되는 면역 자극 | 세포 매개 면역. 대식세포의 살상 능력과 세포 독성 CD8+ T 세포의 증식을 극대화한다. 또한 옵소닌화 항체인 IgG 생성을 촉진한다. | 체액성 면역. B 세포 증식, 항체 클래스 전환 유도, 중화 항체(IgG, IgM, IgA 및 IgE) 생산을 증가시킨다. |
| 기타 기능 | 제1형 사이토카인 IFN-γ는 수지상 세포와 대식세포에 의한 IL-12 생산을 증가시킨다. IL-12는 양성 피드백을 통해 보조 T 세포에서 IFN-γ 생산을 자극하여 Th1 프로파일을 강화한다. IFN-γ는 또한 제2형 반응에 중요한 IL-4와 같은 사이토카인 생산을 억제하여 자체 반응을 유지하는 역할도 한다. | 제2형 반응은 두 가지 다른 사이토카인을 사용하여 자체 프로파일을 강화한다. IL-4는 보조 T 세포에 작용하여 Th2 사이토카인(자신 포함; 자기 조절) 생성을 촉진한다. 반면, IL-10은 보조 T 세포에서 IL-2 및 IFN-γ를 포함한 다양한 사이토카인과 수지상 세포 및 대식세포에서 IL-12 생산을 억제한다. 이 두 사이토카인의 조합된 작용은 T 세포가 특정 사이토카인을 생성하기로 결정하면 그 결정이 유지되고 다른 T 세포도 동일한 경로를 따르도록 유도함을 시사한다. |
보조 T 세포가 어떤 유형의 사이토카인 패턴을 생성하는지는 알려져 있지만, 그 패턴이 어떻게 결정되는지에 대해서는 아직 완전히 이해되지 않았다. 다양한 증거에 따르면 항원을 T 세포에 제시하는 APC의 유형이 T 세포의 분화 방향에 큰 영향을 미친다고 한다. 다른 연구에서는 1차 활성화 동안 T 세포에 제시되는 항원의 농도가 분화 선택에 영향을 미친다고 제시한다. 특정 사이토카인(위에 언급된 것들)의 존재 여부도 최종적으로 어떤 면역 반응이 유도될지에 영향을 미치지만, 이에 대한 이해는 아직 완전하지 않다.
Th2 세포는 사이토카인을 방출하여 체액성 면역 반응을 활성화하는 기능을 한다. 체액성 면역은 항체 분자가 관여하는 면역 반응으로, 항체 단백질은 활성화된 B 세포에 의해 생산된다. B 세포 표면에는 B 세포 항원 수용체(BCR)가 있으며, 세포 내로 항원을 받아들여 분해한 후 그 조각을 MHC 클래스 II 분자를 통해 세포 표면에 제시한다. Th2 세포는 TCR을 통해 B 세포가 제시한 항원 조각을 인식하고, Th2 세포 표면의 CD40 리간드 발현과 사이토카인 방출을 통해 B 세포의 분화와 증식을 촉진한다. Th2 세포가 방출하는 사이토카인(Th2 사이토카인)에는 IL-4, IL-5, IL-6, IL-10, IL-13, GM-CSF 등이 있다. 특히 IL-4 자극은 B 세포의 IgE 생산을 증가시킨다. IgE는 비만 세포나 호염기구 표면에 결합하여 히스타민 방출 촉진 등에 관여하는 중요한 분자이다. 또한, Th2 세포가 생산하는 IL-10과 TGF-β는 Th1 반응을 억제하는 기능을 한다.
Th1 세포와 Th2 세포는 서로 억제하며 균형을 유지하지만, 이 균형이 Th2 쪽으로 기울어 Th2 사이토카인이 과잉 생산되면 기관지 천식이나 아토피성 피부염과 같은 알레르기성 질환(소위 Th2 질환)이 발생할 수 있다.
1차 림프 기관인 흉선에서 생산된 나이브 T 세포(미경험 T 세포)는 말초 림프 조직을 순환하며 항원 침입에 대비한다. 나이브 T 세포가 IL-4의 자극을 받으면 전사 인자인 STAT6를 통해 GATA-3의 전사 활성화가 유도된다.[44] GATA-3는 나이브 T 세포에서도 어느 정도 발현되지만, Th2 세포로 분화하는 과정에서 발현량이 더욱 증가하여 핵심 전사 인자로 기능한다. GATA-3는 Th2 사이토카인의 발현을 조절하는 동시에, Th1 분화에 관여하는 전사 인자인 STAT4의 발현을 억제하는 것으로 알려져 있다.[45]
4. 2. Th17, THαβ 헬퍼 세포
Th17 세포는 Th1 및 Th2 계통과는 발달적으로 구별되는 보조 T세포의 한 종류이다. Th17 세포는 인터루킨 17(IL-17), 인터루킨 21(IL-21), 인터루킨 22(IL-22)와 같은 염증 유발 물질을 만든다.[26] 이 때문에 Th17 세포는 특히 세포 밖의 병원체[26]나 진균, 특히 ''칸디다'' 속에 대한 점막 면역 반응에서 중요한 역할을 한다.[27]THαβ 보조 T 세포는 바이러스에 대한 숙주 면역을 제공한다. 이들의 분화는 IFN α/β 또는 IL-10에 의해 유발된다. 주요 효과기 사이토카인은 IL-10이다. 주요 효과기 세포는 NK 세포, CD8 T 세포, IgG B 세포, IL-10 CD4 T 세포 등이다. 주요 THαβ 전사 인자는 STAT1, STAT3, IRF 등이다. CD4 T 세포에서 나온 IL-10은 NK 세포의 ADCC를 활성화시켜 바이러스에 감염된 세포의 아폽토시스를 유도하고, 숙주와 바이러스의 DNA 조각화를 일으킨다. IFN 알파/베타는 바이러스 복제와 전파를 막기 위해 전사를 억제할 수 있다. 자가항원에 대한 THαβ의 과도한 활성화는 2형 항체 의존성 세포 독성 과민 반응을 일으킬 수 있다. 중증 근무력증 또는 그레이브스병이 이 범주에 속한다.[28]
4. 3. Th1/Th2 모델의 한계
Th1/Th2 모델에서 나타나는 사이토카인 간의 상호 작용은 일부 동물에서는 더 복잡한 양상을 보일 수 있다. 예를 들어, Th2 사이토카인으로 알려진 IL-10은 인간의 경우, Th1과 Th2 두 아형 모두의 사이토카인 생성을 억제한다. 인간의 IL-10(hIL-10)은 모든 T 세포의 증식과 사이토카인 생성을 억제하고 형질 세포의 활성도 억제하지만, 항체 생성이 지속되도록 자극하는 역할도 한다. 따라서 hIL-10은 인간에게서 단순히 Th2 반응을 촉진하기보다는, 항체 생성을 극대화하면서 동시에 헬퍼 T 세포의 과도한 자극을 방지하는 조절 기능을 수행하는 것으로 이해된다.또한, 자연 조절 T 세포(Treg)와 같이 헬퍼 T 세포의 발현과 활성에 영향을 미치는 다른 유형의 T 세포들이 존재한다. Th3 세포와 같이 덜 일반적인 사이토카인 분비 패턴을 보이는 아형도 있다. 특히, 헬퍼 CD4+ T 세포가 조절 T 세포와 별개로 스스로의 반응을 조절하고 억제할 수 있다는 사실이 밝혀지면서 '조절'이나 '억제'와 같은 용어의 의미가 다소 모호해졌다.
조절 T 세포와 이펙터 T 세포의 주요 차이점 중 하나는, 조절 T 세포는 주로 면역 반응을 조절하고 비활성화하는 역할을 하는 반면, 이펙터 T 세포 그룹은 일반적으로 면역을 촉진하는 사이토카인을 먼저 분비하고 이후 생애 주기의 후반부에 억제성 사이토카인을 분비하는 방향으로 전환한다는 점이다. 후자의 특징은 초기 활성화 및 사이토카인 생산 후 조절 아형으로 변환되는 Th3 세포에서 나타난다.
조절 T 세포와 Th3 세포 모두 사이토카인 형질 전환 성장 인자-베타(TGF-β)와 IL-10을 생성한다. 이 두 사이토카인은 모두 헬퍼 T 세포에 대해 억제적인 효과를 나타내며, 특히 TGF-β는 면역계 전반의 활성을 억제하는 강력한 기능을 가지고 있다. TGF-β가 활성화된 Th2 세포에 대해서는 미분화 T 세포(Naive cell)만큼 효과적으로 억제하지 못할 수 있다는 증거도 있지만, 일반적으로 Th2 사이토카인으로 분류되지는 않는다.
Th17 세포[29]라는 새로운 T 헬퍼 아형의 발견은 기존의 Th1/Th2 모델에 대한 의구심을 더욱 증폭시켰다. IL-17을 생성하는 이 세포는 처음에는 자가 면역 질환과 관련된 병적인 세포 집단으로 여겨졌으나, 현재는 독자적인 이펙터 기능과 조절 기능을 모두 갖춘 것으로 이해된다. 특히 일부 증거는 기능적 가소성(plasticity), 즉 환경에 따라 기능이나 특성이 변하는 능력이 헬퍼 T 세포의 고유한 특징일 수 있음을 시사한다. 실제로 쥐를 이용한 연구에서는 Th17 세포가 ''생체 내''에서 Th1 세포로 변환될 수 있음을 보여주었다.[30] 이후의 연구들은 인간에게서도 광범위한 T 헬퍼 세포 가소성이 뚜렷하게 나타난다는 것을 확인했다.[31]
이 글에서 언급된 많은 사이토카인들은 다른 종류의 면역 세포에서도 발현될 수 있다(자세한 내용은 각 사이토카인 항목 참조). 따라서 초기의 Th1/Th2 모델이 헬퍼 T 세포의 기능에 대한 중요한 통찰력을 제공하고 이해를 도왔던 것은 사실이지만, 헬퍼 T 세포의 전체 역할이나 작용 방식을 설명하기에는 너무 단순하다는 점이 분명해지고 있다. 일부 면역학자들은 이 모델 자체에 근본적인 의문을 제기하기도 하는데, 일부 생체 내 연구에서 개별 헬퍼 T 세포들이 Th 모델에서 제시하는 특정 사이토카인 프로파일과 항상 일치하지 않으며, 많은 세포가 Th1과 Th2 양쪽 프로파일에 속하는 사이토카인을 동시에 발현한다는 결과가 나왔기 때문이다.[32] 그럼에도 불구하고, Th 모델은 면역 반응 과정에서 헬퍼 T 세포와 이들이 생성하는 사이토카인의 역할 및 행동 양식을 이해하는 데 중요한 기여를 해왔다.
Stockinger 등의 연구에 따르면 또 다른 T 헬퍼 아형이 존재할 가능성이 있다. Th9 세포는 IL-9을 생성하는 T 세포 아형으로, 특히 기생충 감염에 대한 방어에 중요한 역할을 하는 것으로 알려져 있다.[33]
5. 기억 T 세포
역사적으로 기억 T 세포는 각각 고유한 세포 표면 마커 세트를 가진 효과기 또는 중심 기억 아형에 속하는 것으로 여겨졌다.[34] 중심 기억 T 세포는 림프절에 존재하고, 효과기 기억 T 세포는 C-C 케모카인 수용체 유형 7 (CCR7) 및 L-selectin (CD62L) 수용체가 없어 림프절로 이동하지 못한다.
이제 기억 T 세포의 추가적인 개체군이 존재하는 것으로 알려져 있다. 여기에는 조직 상주 기억 T (Trm) 세포와 가상 기억 T 세포가 포함된다.[35] 모든 기억 T 세포 아형에 대한 단일 통합 주제는 이들이 오래 살고 해당 항원을 만나면 효과기 T 세포의 큰 숫자로 빠르게 확장될 수 있다는 것이다. 이러한 메커니즘을 통해 그들은 이전에 만났던 병원체에 대한 "기억"을 면역계에 제공한다.
6. 질병에서의 역할
다양하고 중요한 보조 T 세포의 역할을 고려할 때, 이 세포들이 질병에 대한 면역 반응에 영향을 미치는 것은 놀라운 일이 아니다. 또한 때때로 유익하지 않은 반응을 생성하기도 한다. 매우 드물게 보조 T 세포 반응은 숙주의 죽음으로 이어질 수 있다.
6. 1. 항종양 면역
이 주제의 주요 내용은 CD4+ T 세포와 항종양 면역 문서에서 다룬다.6. 2. 과민성
면역 체계는 외부 항원에는 반응하면서도 자신의 신체 조직(숙주 항원)에는 반응하지 않도록 민감도의 균형을 유지해야 한다. 면역 체계가 정상적으로는 반응하지 않아야 할 매우 낮은 수준의 항원에 반응할 때 과민성 반응이 나타난다. 과민성은 알레르기 및 일부 자가면역 질환의 원인으로 여겨진다.과민 반응은 크게 네 가지 유형으로 나눌 수 있다.
- 제1형 과민성: 천식, 알레르기성 비염 (건초열), 습진, 두드러기, 아나필락시스와 같은 흔한 면역 질환이 여기에 속한다. 이러한 반응은 모두 IgE 항체와 관련이 있으며, 이 항체의 생성에는 보조 T 세포 중 Th2 세포로의 분화 과정이 필요하다. 코르티코스테로이드나 몬테루카스트와 같은 예방적 치료는 비만 세포나 다른 알레르기 관련 세포를 억제하는 데 초점을 맞춘다. 실제 염증 반응에서 T 세포가 주된 역할을 하지는 않는다. 과민성 반응의 '유형' 번호는 Th 세포 반응 모델의 번호(Th1, Th2 등)와는 직접적인 관련이 없다.
- 제2형 과민성과 제3형 과민성: 이 두 유형은 주로 자가 항체나 낮은 친화도의 항체로 인한 문제와 관련이 있다. 이 반응들에서 T 세포는 자가 항체를 만드는 데 관여할 수 있지만, 제2형 과민 반응 중 일부(예: 출산 시 Rh 인자 반응)는 건강한 면역 체계에서도 정상적인 면역 반응으로 간주될 수 있다. 이러한 반응에서 보조 T 세포의 정확한 역할은 아직 완전히 밝혀지지 않았지만, 일반적으로 Th2 세포에서 분비되는 사이토카인이 질환을 촉진할 것으로 생각된다. 예를 들어, 루푸스(SLE)와 같은 자가면역 질환은 Th2 사이토카인 생성과 관련이 있다는 연구 결과가 있다.
- 제4형 과민성: 지연형 과민성이라고도 불리며, 림프구나 대식세포와 같은 면역 세포가 과도하게 자극되어 만성적인 염증과 사이토카인 방출을 유발한다. 항체는 이 유형의 알레르기 반응에 직접 관여하지 않는다. T 세포는 이 과민 반응에서 중요한 역할을 하는데, 자극 자체에 대해 활성화될 뿐만 아니라, Th1 사이토카인을 통해 다른 세포, 특히 대식세포의 활성화를 촉진한다.
기타 세포 매개 과민성으로는 세포독성 T 세포가 매개하는 자가면역 질환과 유사한 현상인 이식 거부가 있다. 보조 T 세포는 이러한 질환의 발달을 촉진하는 데 필수적이다. 충분한 수의 자가면역 반응성 살해 T 세포(세포독성 T 세포)를 만들기 위해서는 인터루킨-2(IL-2)가 필요한데, 이는 주로 CD4+ 보조 T 세포에 의해 공급된다. 또한 CD4+ T 세포는 인터페론-감마(IFN-γ)와 같은 사이토카인을 통해 자연 살해 세포나 대식세포를 자극하여, 특정 상황에서 이 세포들이 숙주 세포를 공격하도록 유도할 수 있다.
자가면역 질환에서 살해 T 세포가 사용하는 공격 메커니즘은 바이러스 감염에 대한 반응과 매우 유사하다. 실제로 일부 바이러스는 제1형 당뇨병과 같은 자가면역 질환의 유발 요인으로 지목되기도 한다. 세포 매개 자가면역 질환은 우리 몸의 항원 인식 시스템에 오류가 생겨 면역 체계가 자신의 항원을 외부 침입자로 잘못 인식하기 때문에 발생한다. 결과적으로, CD8+ T 세포는 해당 항원을 발현하는 자신의 세포를 감염된 세포로 간주하고 파괴하게 된다(이식 거부의 경우 이식된 장기 세포를 파괴한다).
Th2 세포는 사이토카인을 방출하여 액성 면역 반응을 활성화하는 기능을 한다. 액성 면역은 항체 분자가 관여하는 면역 반응으로, 항체 단백질은 활성화된 B 세포에 의해 생산된다. B 세포 표면에는 B 세포 수용체(BCR)가 있어 항원을 인식하고 세포 안으로 받아들여 분해한 뒤, 그 조각을 MHC 클래스 II 분자를 통해 세포 표면에 제시한다. Th2 세포는 자신의 T 세포 항원 수용체(TCR)를 통해 B 세포가 제시한 항원 조각을 인식한다. 이후 Th2 세포는 표면에 CD40 리간드를 발현시키고 사이토카인을 방출하여 B 세포의 분화와 증식을 촉진한다. Th2 세포가 방출하는 사이토카인(Th2 사이토카인)에는 IL-4, IL-5, IL-6, IL-10, IL-13, GM-CSF 등이 있다. 특히 IL-4 자극은 B 세포에서 면역글로불린 E(IgE) 생산을 크게 늘린다. IgE는 비만 세포나 호염기구 표면에 결합하여 히스타민 방출을 촉진하는 등 알레르기 반응에서 중요한 역할을 한다. 또한, Th2 세포가 생산하는 IL-10 및 TGF-β는 Th1 반응을 억제하는 기능도 가지고 있다.
앞서 언급했듯이 Th1 세포와 Th2 세포는 서로의 기능을 억제하며 균형을 이루지만, 이 균형이 Th2 쪽으로 기울어 Th2 사이토카인이 과도하게 생산되면 기관지 천식이나 아토피 피부염과 같은 소위 'Th2 질환'이라 불리는 알레르기성 질환이 발생할 수 있다.
이 설명의 일부는 단순화된 내용이다. 많은 자가면역 질환은 훨씬 더 복잡하며, 예를 들어 류마티스 관절염의 경우 항체와 면역 세포 모두 질병 과정에 관여하는 것으로 알려져 있다. 일반적으로 대부분의 자가면역 질환에 대한 면역학적 기전은 아직 완전히 밝혀지지 않았다.
6. 3. HIV 감염
보조 T 세포의 중요성은 인간 면역 결핍 바이러스(HIV) 감염 시 CD4+ T 세포가 파괴되는 현상을 통해 명확히 드러난다. HIV는 주로 림프구인 CD4+ T 세포를 표적으로 삼지만, 대식세포나 수지상 세포처럼 CD4를 발현하는 다른 세포들도 감염시킬 수 있다. 이들 세포는 CD4를 낮은 수준으로 발현한다.HIV 감염의 초기 무증상 단계에서는 바이러스가 T 세포보다는 대식세포에 대한 친화력이 더 높으며, 이로 인해 CD4+ T 세포의 사멸 속도가 비교적 느리다고 알려져 있다. 이때 면역계는 흉선(원래는 골수에서 생성됨)에서 새로운 보조 T 세포를 만들어 손실을 보충한다. 그러나 바이러스가 림프친화성(T 세포 친화성)으로 변하면, CD4+ T 세포를 훨씬 효율적으로 감염시키기 시작한다. 이는 감염 시 바이러스가 결합하는 공동 수용체의 변화 때문일 가능성이 높으며, 이 단계에 이르면 면역 체계는 감당하기 어려워진다. 연구에 따르면 HIV가 표적으로 하는 림프구 유래 CD4 T 세포 중 약 5%만이 바이러스를 생산적으로 감염시키는 데 허용적이다. 나머지 95% 이상의 사멸하는 CD4 T 세포는 휴지 상태이며 생산적인 감염을 지원할 수 없다. 이 세포들은 HIV에 의한 불완전한 감염을 겪는다.[36] 세포는 HIV의 외래 DNA 중간체를 감지하면 숙주를 보호하기 위해 자살적인 사멸 경로를 개시한다. 이 과정에서 인플라마솜 내 caspase-1이 활성화되어 파이롭토시스(매우 염증적인 형태의 프로그램 세포 사멸)라는 세포 사멸이 유발된다.[37][38][39]
이 시점부터 만성 염증이 시작되고 기능적인 CD4+ T 세포 수치가 감소하여, 결국 감지 가능한 모든 항원을 인식하기에는 CD4+ T 세포 집단이 너무 작아진다. CD4 T 세포 고갈과 만성 염증 발생은 후천성 면역 결핍 증후군(AIDS)으로 진행되는 HIV 발병 기전의 핵심 특징이다. AIDS 상태에서는 혈중 CD4 T 세포 수가 마이크로리터당 200개 미만으로 고갈되어, 다양한 병원체가 T 세포의 감시를 피할 수 있게 된다. 이로 인해 일반적으로 보조 T 세포 반응을 유도하는 기회 감염이 면역 체계를 쉽게 우회하게 된다.[40] 이러한 완전한 우회는 감염 제거에 보조 T 세포 반응이 필수적인 경우에만 발생하지만, 대부분의 감염은 보조 T 세포가 부족하여 면역 반응이 덜 효율적이 되므로 감염의 심각성이나 지속 기간이 증가한다.
면역 체계의 두 가지 주요 구성 요소는 CD4+ T 세포 의존성 때문에 AIDS에서 특히 큰 영향을 받는다.
# CD8+ T 세포: AIDS 단계에서는 효과적으로 자극받지 못하여, 환자는 HIV 자체를 포함한 대부분의 바이러스에 매우 취약해진다. CD4+ T 세포의 사멸 감소는 바이러스가 더 오랫동안 생성되게 하여(감염된 CD4+ T 세포가 더 빨리 사멸되지 않음) 바이러스 증식을 증가시키고 질병 진행을 가속화한다.
# 항체 클래스 전환: 보조 T 세포 기능 부전으로 인해 현저히 감소한다. 면역계는 항체의 친화력을 개선하는 능력을 상실하고, IgG나 IgA와 같은 중요한 항체를 생산하는 B 림프구를 만들 수 없게 된다. 이는 주로 보조 T 세포가 B 림프구와 제대로 상호작용하지 못하기 때문이다. AIDS의 또 다른 증상은 Th2 사이토카인 감소(및 보조 T 세포와의 상호작용 감소)로 인한 항체 수치 감소이다. 이러한 문제들은 IgM 항체가 도달하기 어려운 신체 부위에서 공격적인 세균 감염에 대한 감수성을 높인다.
환자가 HIV 치료에 반응하지 않거나 치료를 받지 못하면, 면역 체계가 질병을 처리하기에 더 이상 적절히 조절되거나 충분히 자극되지 못하는 지경에 이르러 결국 암이나 감염으로 사망하게 된다.
HIV 감염 중 CD4 T 세포 확장이 억제되는 것은 미생물 전위(microbial translocation)에 의해 IL-10 의존적인 방식으로 일어날 수 있다. 활성화된 단핵구에서 PD-L1 리간드에 의해 발현되는 PD-1을 활성화하면 IL-10 생성이 유도되어 CD4 T 세포 기능을 억제한다.[41]
6. 4. COVID-19
코로나19(COVID-19) 감염 시 B 세포, 자연 살해 세포 및 전체 림프구 수가 감소하지만, CD4+ T세포와 CD8+ T세포는 훨씬 더 큰 폭으로 감소하는 경향을 보인다.[42] 이는 SARS-CoV-2가 감염 과정에서 CD4+ T세포를 직접 공격할 수 있음을 시사한다. 실제로 낮은 CD4+ 수치는 환자가 중환자실에 입원할 가능성이 더 높다는 것을 예측하는 지표가 되었으며, CD4+ T세포 수는 바이러스 RNA가 체내에서 제거되는 기간을 예측하는 유일한 요인으로 밝혀졌다.[42] 흥미롭게도, 전체적인 CD4+ 수치가 감소했음에도 불구하고, 중증 코로나19 환자는 중등도 질환을 앓는 환자보다 특정 유형인 Th1 CD4+ T세포 수치가 더 높은 것으로 나타났다.[43]참조
[1]
논문
Chromosome contacts in activated T cells identify autoimmune disease candidate genes
2017-09
[2]
논문
Master regulators or lineage-specifying? Changing views on CD4+ T cell transcription factors
2012-11
[3]
논문
The biology of recent thymic emigrants
2013-03-21
[4]
논문
The full spectrum of human naive T cells
2018-06
[5]
논문
Neonatal thymectomy reveals differentiation and plasticity within human naive T cells
2016-03
[6]
논문
The ins and outs of MHC class II-mediated antigen processing and presentation
2015-04
[7]
논문
Variations in MHC Class II Antigen Processing and Presentation in Health and Disease
2016-05
[8]
논문
Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation
2017-03-17
[9]
논문
The Length Distribution of Class I-Restricted T Cell Epitopes Is Determined by Both Peptide Supply and MHC Allele-Specific Binding Preference
2016-02
[10]
서적
Janeway's immunobiology
Garland Science
2017
[11]
논문
The perfect mix: recent progress in adjuvant research
2007-07
[12]
논문
Molecular control of steady-state dendritic cell maturation and immune homeostasis
2013-03-21
[13]
논문
Why is there so much CD45 on T cells?
2007-09
[14]
논문
CD45 functions as a signaling gatekeeper in T cells
2019-10
[15]
논문
Apoptosis: a review of programmed cell death
2007-06
[16]
논문
Identification of a novel surface protein on activated CD4+ T cells that induces contact-dependent B cell differentiation (help)
1992-04
[17]
논문
Molecular and biological characterization of a murine ligand for CD40
1992-05
[18]
논문
A 39-kDa protein on activated helper T cells binds CD40 and transduces the signal for cognate activation of B cells
1992-07
[19]
논문
Thymic and Postthymic Regulation of Naïve CD4(+) T-Cell Lineage Fates in Humans and Mice Models
2016
[20]
논문
CD4 T cells: fates, functions, and faults
2008-09
[21]
논문
GATA3: a master of many trades in immune regulation
2014-06
[22]
논문
Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: a critical review
2015-02
[23]
논문
Differential glycosylation of TH1, TH2 and TH-17 effector cells selectively regulates susceptibility to cell death
2007-08
[24]
서적
Pharmacology
Churchill Livingstone
[25]
논문
Interleukin-10 production by Th1 cells requires interleukin-12-induced STAT4 transcription factor and ERK MAP kinase activation by high antigen dose
2009-08
[26]
논문
Th17 cytokines in mucosal immunity and inflammation
[27]
논문
The Th17/IL-17 Axis and Host Defense Against Fungal Infections
[28]
간행물
Microarray analysis of PBMC gene expression profiles after Plasmodium falciparum malarial infection
Johns Hopkins University
[29]
논문
Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages
2005-11
[30]
논문
Fate mapping of IL-17-producing T cells in inflammatory responses
2011-03
[31]
논문
Multiparameter grouping delineates heterogeneous populations of human IL-17 and/or IL-22 T-cell producers that share antigen specificities with other T-cell subsets
UPMC Paris 06 Institut National de la Santé et de la Recherche Médicale (Inserm) UMR-S 945
2011-09
[32]
논문
Helper T cell diversity and plasticity
2012-06
[33]
논문
Alternative lifestyles of T cells
2008-12
[34]
논문
Two subsets of memory T lymphocytes with distinct homing potentials and effector functions
1999-10
[35]
논문
CD4+ virtual memory: Antigen-inexperienced T cells reside in the naïve, regulatory, and memory T cell compartments at similar frequencies, implications for autoimmunity
2017-02
[36]
논문
Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue
2010-11
[37]
논문
Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection
2014-01
[38]
논문
IFI16 DNA sensor is required for death of lymphoid CD4 T cells abortively infected with HIV
2014-01
[39]
논문
NLRP3 inflammasome induces CD4+ T cell loss in chronically HIV-1-infected patients
2021-03
[40]
웹사이트
CD4 Count
https://www.aids.gov[...]
2015-04-30
[41]
논문
Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection
2010-04
[42]
논문
Lymphocyte Subset Counts in COVID-19 Patients: A Meta-Analysis
2020-08
[43]
논문
COVID-19 poses a riddle for the immune system
2020-08
[44]
간행물
STAT6-independent GATA-3 autoactivation directs IL-4 independent Th2 development and commitment.
2000
[45]
간행물
GATA-3 suppresses Th1 development by downregulation of Stat4 and not through effects on IL-12Rbeta2 chain or T-bet.
2003
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com