트랜스로콘
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
트랜스로콘은 세균의 SecYEG, 진핵생물의 Sec61로 불리는 이종 삼량체 단백질 복합체이며, 단백질이 세포막을 통과하도록 돕는 채널이다. 이 채널은 단백질 전위 과정에서 펩타이드가 양방향으로 이동할 수 있게 하므로, 트랜스로콘 내에는 추가적인 시스템이 존재한다. 트랜스로케이션에는 리보솜에서 단백질이 합성되는 동안 일어나는 공동 번역 트랜스로케이션과 번역 후에 일어나는 번역 후 트랜스로케이션이 있으며, 진핵생물과 세균에서 각각 다른 방식으로 일어난다. 또한, 트랜스로콘은 소포체 내강에서 잘못 접히거나 손상된 단백질을 세포질로 이동시켜 분해를 돕는 역할도 수행하며, 트랜스로콘 품질 관리 시스템을 통해 기능 장애를 회복한다.
더 읽어볼만한 페이지
- 단백질 복합체 - 단백질-단백질 상호작용
단백질-단백질 상호작용은 단백질 간의 결합을 의미하며, 결합의 지속성, 결합력, 구성 단백질 종류에 따라 다양한 유형으로 분류되고, 물 분자에 의해 조절되며, 연구 방법과 데이터베이스를 통해 질병 연구 및 신약 개발에 응용된다. - 단백질 복합체 - 광계 I
광계 I(PSI)은 광합성 전자전달계의 효소로, 빛 에너지를 흡수하여 전자를 전달하며, P700을 포함한 다양한 색소, 조효소, 보조 인자로 구성되어 있다. - 내재성 막 단백질 - 광계
광계는 틸라코이드 막에 위치하며 빛 에너지를 화학 에너지로 전환하는 단백질 복합체로, 광수집 복합체에 둘러싸인 반응 중심을 가지며 물 분해를 통해 전자를 얻고 산소를 방출하는 광계 II와 NADPH를 생성하는 광계 I 두 가지 주요 유형으로 나뉜다. - 내재성 막 단백질 - 사이토크롬 b6f 복합체
사이토크롬 b6f 복합체는 광합성에서 광계 II와 광계 I 사이의 전자 및 에너지 전달을 매개하고 틸라코이드 막을 가로질러 양성자를 수송하여 ATP 합성을 촉진하는 단백질 복합체로서, 순환적 광인산화에도 중요한 역할을 하며 사이토크롬 bc1 복합체와 구조적 유사성을 갖는 8개의 소단위체로 구성된 이량체이다.
| 트랜스로콘 | |
|---|---|
| 일반 정보 | |
 | |
| 발견 연도 | 1970년대 |
| 발견자 | 귄터 블로벨 |
| 기능 | 소포체, 세균의 세포막, 고세균의 세포막을 가로지르는 폴리펩타이드 수송 막 단백질 삽입 |
| 구조 | |
| 진핵생물 | Sec61 복합체 (Sec61α, Sec61β, Sec61γ로 구성) TRAM TRAP 복합체 OST 수용체 |
| 세균 | SecYEG 복합체 SecA SecDF-YajC YidC |
| 메커니즘 | |
| 신호 인식 | 진핵생물: 신호 인식 입자 (SRP) 세균: SecA |
| 채널 개방 | 펩타이드의 소수성 신호 서열 또는 막 횡단 도메인에 의해 유도됨 |
| 펩타이드 수송 | 채널을 통한 폴리펩타이드의 통과; ATP 가수분해에 의해 구동될 수 있음 |
| 막 삽입 | 측면 방출 메커니즘을 통해 |
| 관련 과정 | |
| 단백질 타겟팅 | 세포 내 특정 구획으로 단백질을 전달하는 과정 |
| 단백질 접힘 | 샤페론의 도움으로 |
| 단백질 변형 | 글리코실화, 지질화 |
| 중요성 | |
| 세포 생존 | 세포가 생존하고 기능하는 데 필요한 많은 단백질이 translocon을 통해 수송됨 |
| 질병 | translocon 기능 장애는 다양한 질병과 관련됨 |
| 추가 정보 | |
| 다른 이름 | 단백질 translocon, Sec61 복합체, SecYEG 복합체 |
2. 중앙 채널
트랜스로콘의 핵심 구성 요소는 세균에서는 SecYEG, 진핵생물에서는 Sec61 복합체이다.[5] 이들은 막을 관통하는 단백질 통로를 형성하여 단백질이 세포막을 통과할 수 있도록 돕는다. 더 큰 7량체 복합체는 폴리펩티드 하위 집합을 소포체로 수송하는 역할을 한다.[7] 이 채널은 소포체 막에서 기능하며, 들어가는 요소의 선택성을 보장한다.
2. 1. SecYEG/Sec61 복합체의 구조
트랜스로콘 채널은 세균에서는 SecYEG, 진핵생물에서는 Sec61이라고 불리는 이종 삼량체 단백질 복합체이다.[5] SecY, SecE, SecG 소단위체로 구성된다. 이 채널의 비활성 상태 구조는 고세균에서 X선 결정학으로 밝혀졌다.[6] SecY는 큰 기공 소단위체이다. 측면에서 보면 채널은 양쪽 측면에 깔때기가 있는 모래시계 모양이다. 세포외 깔때기는 알파 나선으로 형성된 작은 "플러그"를 가지고 있다. 막 중앙에는 측쇄를 안쪽으로 투사하는 6개의 소수성 아미노산으로 이루어진 기공 링 구조가 있다. 단백질 전위 동안 플러그는 치워지고 폴리펩티드 사슬은 세포질 깔때기를 통해 기공 링을 통과하여 세포외 깔때기를 거쳐 세포외 공간으로 이동한다. 막 단백질의 소수성 부분은 측면 게이트를 통해 지질상으로 옆으로 나가 막 관통 세그먼트가 된다.[6]2. 2. 추가 구성 요소
세균에서 SecYEG는 SecDF, YajC, YidC와 복합체를 형성한다.[8][9] 진핵생물에서 Sec61은 올리고사카릴 전달 효소 복합체, TRAP 복합체 및 막 단백질 TRAM (샤프론일 가능성이 있음)과 복합체를 형성한다. 신호 펩티다제 복합체 및 SRP 수용체와 같은 추가 구성 요소는 트랜스로콘 복합체와 일시적으로만 연관되는 것으로 보인다.[10]3. 트랜스로케이션 (단백질 이동)
트랜스로콘은 펩타이드가 양방향으로 이동할 수 있게 하므로, 펩타이드를 특정 방향으로 이동시키기 위해서는 트랜스로콘 내에 추가적인 시스템이 필요하다. 트랜스로케이션에는 공동 번역 트랜스로케이션(번역과 동시에 일어남)과 번역 후 트랜스로케이션(번역 후에 일어남)의 두 가지 유형이 있다.[33]
공동 번역 트랜스로케이션은 리보솜에서 단백질이 합성되는 동안 신호 인식 입자(SRP)가 신생 단백질의 신호 서열을 인식하여 진행된다. 진핵생물에서 번역 후 트랜스로케이션은 BiP와 SEC62/SEC63 막 단백질 복합체를 포함한 다른 복합체에 의존하며, 세균에서는 SecA가 이 과정을 수행한다.
3. 1. 공동 번역 트랜스로케이션 (Co-translational translocation)
리보솜에서 단백질이 합성되는 동안, 신호 인식 입자(SRP)가 신생 단백질의 신호 서열을 인식한다. SRP는 리보솜-mRNA-신생 단백질 복합체를 소포체 막의 트랜스로콘으로 유도한다. 리보솜이 트랜스로콘에 결합하면, 신생 단백질은 트랜스로콘 채널을 통과하여 소포체 내강으로 이동한다. 이 과정을 공동 번역 트랜스로케이션 (Co-translational translocation)이라고 한다.[12][13][14]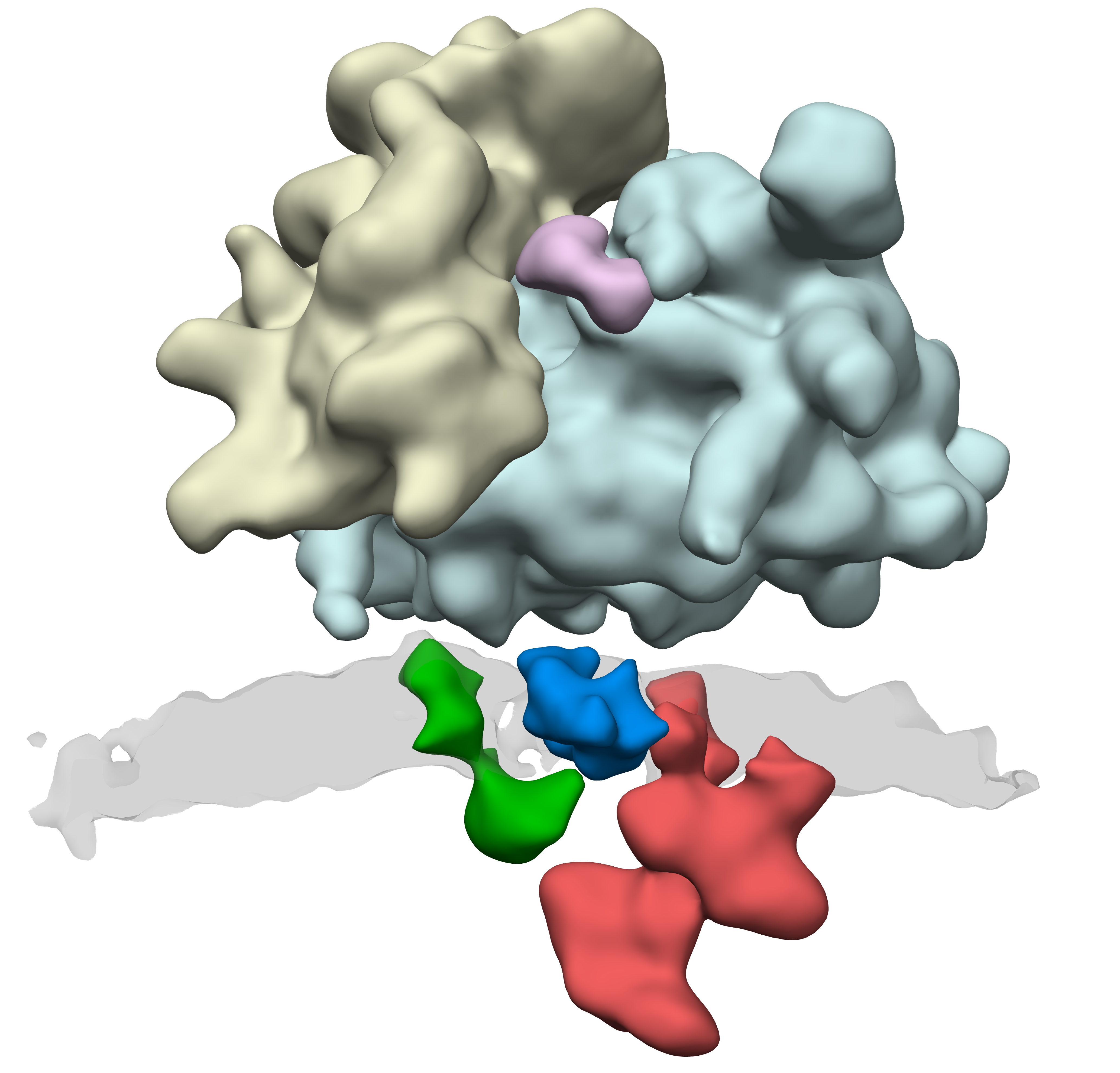
진핵생물에서 소포체로 전위될 단백질은 SRP에 의해 인식된다. SRP는 소포체에서 SRP 수용체에 리보솜을 부착하는 동안 번역을 일시 중단시킨다. 이러한 인식은 합성될 폴리펩타이드의 처음 몇 개 코돈에 있는 특정 N-말단 신호 서열을 기반으로 한다.[11] 박테리아는 진핵생물 TRAM과 유사한 샤페론 YidC와 함께 SRP를 사용한다.[15][11]
트랜스로콘은 막 단백질을 소포체 막에 올바른 방향으로 전위시키고 통합하는 역할도 한다. 이 과정은 완전히 이해되지는 않았지만, 막관통 나선이 되도록 운명 지어진 아미노산 서열의 소수성 부분을 트랜스로콘이 인식하고 처리하는 과정을 포함한다. 정지-전달 서열에 의해 닫히고 내장된 신호 서열에 의해 열리는 플러그는 열리고 닫힌 상태 사이에서 변경되어 나선을 다른 방향으로 배치한다.[11]
번역이 완료되면 신호 펩티드 분해 효소가 신호 서열을 제거하고, 단백질은 소포체 내강에 남거나 막에 삽입된 상태로 남는다.
3. 2. 번역 후 트랜스로케이션 (Post-translational translocation)
트랜스로콘은 펩타이드가 양방향으로 이동할 수 있게 하므로, 펩타이드를 특정 방향으로 이동시키기 위해 트랜스로콘 내에 추가 시스템이 필요하다. 번역 후 트랜스로케이션은 번역 후에 일어난다.진핵생물에서 번역 후 전위는 BiP와 SEC62/SEC63 막 단백질 복합체를 포함한 다른 복합체에 의존한다. 이 전위 방식에서 Sec63은 BiP가 ATP를 가수분해하도록 돕고, 이것은 펩타이드에 결합하여 펩타이드를 "끌어낸다". 이 과정은 전체 펩타이드가 통과될 때까지 다른 BiP 분자에 대해 반복된다.[11]
세균에서는 동일한 과정이 "미는" ATPase인 SecA에 의해 수행되며, 때로는 반대편에서 당기는 역할을 하는 SecDF 복합체의 도움을 받기도 한다.[16] SecA ATPase는 다중 펩타이드를 채널을 통해 이동시키기 위해 "밀고-미끄러지는" 메커니즘을 사용한다. ATP 결합 상태에서 SecA는 기질 내 아미노산의 부분 집합과 두 개의 나선형 손가락을 통해 상호 작용하여, ATP 가수분해와 함께 아미노산을 채널로 밀어 넣는다. 그런 다음 SecA가 ADP 결합 상태로 들어가면서 상호 작용이 약해지고, 폴리펩타이드 사슬이 어느 방향으로든 수동적으로 미끄러지도록 한다. 그 후 SecA는 펩타이드의 추가 부분을 잡아 이 과정을 반복한다.[11]
4. 소포체 역트랜스로콘 (ER-retrotranslocon)
트랜스로콘은 프로테아좀으로 보내지는 손상된 단백질과 같은 폴리펩타이드를 소포체 내강에서 세포질로 이동시킬 수 있다. 소포체 단백질은 세포질에서 26S 프로테아좀에 의해 분해되며, 이는 소포체 관련 단백질 분해로 알려진 과정이며, 따라서 적절한 채널을 통해 수송되어야 한다. 이 역트랜스로콘은 아직 수수께끼이다.
처음에는 Sec61 채널이 이 역행 수송을 담당하는 것으로 여겨졌으며, 이는 Sec61을 통한 수송이 항상 단방향이 아니라 양방향일 수도 있음을 의미했다.[17] 그러나 Sec61의 구조는 이러한 견해를 뒷받침하지 않으며, 소포체 내강에서 세포질로의 수송을 담당하는 여러 다른 단백질이 제안되었다.[18]
5. 트랜스로콘 품질 관리 (TQC)
트랜스로콘은 번역이 정지되거나 부적절하게 접힌 단백질이 복합체와 결합하여 막힐 수 있다. 이는 트랜스로콘이 기능 장애를 일으킬 수 있는 한 가지 방식이다. 예를 들어, 동시 번역적 이동(CTT)에서 트랜스로콘 막힘은 번역이 정지된 ER 표적 단백질로 인해 발생할 수 있다.[19] 번역 후 이동(PTT) 중 트랜스로콘 막힘은 단백질이 완전히 이동되기 전에 제대로 접히지 않거나 응집체를 형성할 때 발생할 수 있다.[20][21][22]
세포 내 트랜스로콘 품질 관리(TQC) 메커니즘은 단백질 이동 중 막힌 트랜스로콘 채널을 제거하여 트랜스로콘 기능을 회복시킨다.[21] 유비퀴틴 프로테아좀 시스템(UPS)은 TQC 하에 있는 여러 분해 메커니즘 중 하나이다. 이 과정에는 프로테아좀에 의한 분해를 위해 유비퀴틴 효소를 부착하여 막힌 단백질을 표적화하는 것이 포함된다.[23]
6. 관련 항목
템플릿은 제거해야 하는 대상이므로 제거하고, 내부 링크 문법에 맞게 수정한다.
- SecY 단백질
- 세균 분비 시스템
참조
[1]
논문
The translocon: a dynamic gateway at the ER membrane
[2]
논문
Structure and function of the bacterial Sec translocon
[3]
논문
Assembly of yeast Sec proteins involved in translocation into the endoplasmic reticulum into a membrane-bound multisubunit complex
1991-02
[4]
논문
The type III secretion system tip complex and translocon
2008-06
[5]
간행물
Academic Press
2016-01-01
[6]
논문
X-ray structure of a protein-conducting channel
2004-01
[7]
논문
Mammalian Sec61 Is Associated with Sec62 and Sec63
2000-05
[8]
논문
Distinct catalytic roles of the SecYE, SecG and SecDFyajC subunits of preprotein translocase holoenzyme
1997-05
[9]
논문
YidC, the Escherichia coli homologue of mitochondrial Oxa1p, is a component of the Sec translocase
2000-02
[10]
논문
Structure of the mammalian oligosaccharyl-transferase complex in the native ER protein translocon
[11]
논문
Protein translocation by the Sec61/SecY channel
2005
[12]
논문
A protein-conducting channel in the endoplasmic reticulum
1991-05
[13]
논문
Signal peptides open protein-conducting channels in E. coli
1992-05
[14]
논문
Delivering proteins for export from the cytosol
https://www.nature.c[...]
2009-04
[15]
논문
YidC protein, a molecular chaperone for LacY protein folding via the SecYEG protein machinery
2013-09
[16]
논문
The bacterial Sec-translocase: structure and mechanism
2012-04
[17]
논문
Surfing the Sec61 channel: bidirectional protein translocation across the ER membrane
1999-12
[18]
논문
Finding the will and the way of ERAD substrate retrotranslocation
2012-08
[19]
논문
Rkr1/Ltn1 Ubiquitin Ligase-mediated Degradation of Translationally Stalled Endoplasmic Reticulum Proteins
2015-07
[20]
논문
The Protease Ste24 Clears Clogged Translocons
https://linkinghub.e[...]
2016-01
[21]
논문
Overlapping function of Hrd1 and Ste24 in translocon quality control provides robust channel surveillance
2020-11
[22]
논문
Translocon Declogger Ste24 Protects against IAPP Oligomer-Induced Proteotoxicity
https://linkinghub.e[...]
2018-03
[23]
논문
ER Degradation of a Misfolded Luminal Protein by the Cytosolic Ubiquitin-Proteasome Pathway
https://www.science.[...]
1996-09-20
[24]
논문
The translocon: a dynamic gateway at the ER membrane
[25]
논문
Structure and function of the bacterial Sec translocon
[26]
논문
The type III secretion system tip complex and translocon
2008-06
[27]
논문
Assembly of yeast Sec proteins involved in translocation into the endoplasmic reticulum into a membrane-bound multisubunit complex
[28]
논문
X-ray structure of a protein-conducting channel
2004-01
[29]
서적
Biogenesis of Secretory Proteins
http://www.sciencedi[...]
Academic Press
2020-12-17
[30]
논문
X-ray structure of a protein-conducting channel
2004-01
[31]
논문
Three-dimensional structure of the bacterial protein-translocation complex SecYEG
2002-08
[32]
논문
Structure of the mammalian oligosaccharyl-transferase complex in the native ER protein translocon
[33]
논문
Protein translocation by the Sec61/SecY channel.
2005
[34]
논문
A protein-conducting channel in the endoplasmic reticulum
[35]
논문
Signal peptides open protein-conducting channels in E. coli
[36]
논문
Protein translocation by the Sec61/SecY channel.
2005
[37]
논문
YidC protein, a molecular chaperone for LacY protein folding via the SecYEG protein machinery
2013
[38]
논문
Protein translocation by the Sec61/SecY channel.
2005
[39]
논문
Protein translocation by the Sec61/SecY channel.
2005
[40]
논문
The bacterial Sec-translocase: structure and mechanism
2012
[41]
논문
Protein translocation by the Sec61/SecY channel.
2005
[42]
논문
Surfing the Sec61 channel: bidirectional protein translocation across the ER membrane
http://jcs.biologist[...]
1999-12
[43]
논문
Finding the will and the way of ERAD substrate retrotranslocation
[44]
논문
The translocon: a dynamic gateway at the ER membrane
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com