사이토크롬 b6f 복합체
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
사이토크롬 b6f 복합체는 광합성에 관여하는 효소 복합체로, 두 개의 광계 사이에서 전자와 에너지를 전달하고 엽록체 스트로마에서 틸라코이드 내강으로 양성자를 이동시킨다. 이량체 구조로, 각 단량체는 8개의 소단위체로 구성되며, 사이토크롬 f, 사이토크롬 b6, 리에스케 철-황 단백질, 소단위체 IV를 포함한 4개의 큰 소단위체와 4개의 작은 소단위체로 이루어져 있다. 사이토크롬 b6f 복합체는 Q 회로를 통해 플라스토퀴놀에서 플라스토사이아닌으로 전자를 전달하며, ATP 합성을 위한 양성자 기울기를 생성하고 순환적 광인산화에도 관여한다.
더 읽어볼만한 페이지
- 철-황 단백질 - 페레독신
페레독신은 철-황 클러스터를 포함하며, 광합성, 수소 생산 등 다양한 생리적 역할에 관여하는 단백질이다. - 철-황 단백질 - 수소화효소
수소화효소는 활성 부위 금속 성분에 따라 니켈-철, 철-황, 철 수소화효소 등으로 나뉘며 미생물에서 발견되는 효소로, 수소 산화 환원 반응 촉매, 에너지 생성, 생물 정화 등 생물학적 기능 수행 및 재생 에너지 분야 응용 가능성으로 주목받는다. - 광반응 - 플라스토퀴논
플라스토퀴논은 광합성에서 전자 전달체로 작용하는 벤조퀴논 유도체로서, 광계 II에서 전자를 받아 사이토크롬 b6f 복합체로 전달하며 ATP 생성에 기여하고, 유도체들은 항산화 효과와 미토콘드리아 기능 개선을 통해 노화 및 뇌 질환 치료 연구에 활용된다. - 광반응 - 광계
광계는 틸라코이드 막에 위치하며 빛 에너지를 화학 에너지로 전환하는 단백질 복합체로, 광수집 복합체에 둘러싸인 반응 중심을 가지며 물 분해를 통해 전자를 얻고 산소를 방출하는 광계 II와 NADPH를 생성하는 광계 I 두 가지 주요 유형으로 나뉜다.
| 사이토크롬 b6f 복합체 | |
|---|---|
| 개요 | |
 | |
| 일반 정보 | |
| 명칭 | 시토크롬 b6f 복합체 |
| 다른 이름 | 플라스토퀴놀/플라스토시아닌 환원 효소 플라스토퀴놀/플라스토시아닌 산화환원 효소 |
| 식별자 | |
| CAS 등록번호 | 79079-13-3 |
| EC 번호 | 7.1.1.6 |
| TCDB | 3.D.3 |
| OPM 패밀리 | 92 |
| OPM 단백질 | 4pv1 |
| Pfam | PF05115 |
| 인터프로 | IPR007802 |
| 막단백질 슈퍼패밀리 | 258 |
2. 효소 구조
사이토크롬 *b*6*f* 복합체는 이량체이며, 각 단량체는 8개의 소단위체로 구성된다.[55] 복합체 내 단량체 간 공간은 지질로 채워져 있으며, 이는 단백질 내 유전 환경 조절을 통해 헴-헴 전자 전달에 방향성을 제공한다.[61][67]
2. 1. 주요 소단위체
사이토크롬 ''b''6''f'' 복합체는 이량체이며, 각 단량체는 8개의 소단위체로 구성된다.[55] 각 단량체는 4개의 큰 소단위체와 4개의 작은 소단위체(3~4 kDa)로 구성된다.[55][56] 총 분자량은 217 kDa이다.[55]- 4개의 큰 소단위체:
- 32 kDa 사이토크롬 ''f'' (''c''형 사이토크롬 포함)[55]
- 25 kDa 사이토크롬 ''b''6 (저전위 및 고전위 헴기 포함)[55]
- 19 kDa 리에스케 철-황 단백질 ([2Fe-2S] 클러스터 포함)[55]
- 17 kDa의 소단위체 IV[55]
2. 2. 분자량 및 결정 구조
사이토크롬 *b*6*f* 복합체의 총 분자량은 217 kDa이다.[55] 클라미도모나스 레인하르드티이(''Chlamydomonas reinhardtii''), 마스티고클라두스 라미노수스(''Mastigocladus laminosus''), 노스톡속(*Nostoc*) 종으로부터 사이토크롬 *b*6*f* 복합체의 결정 구조가 밝혀졌다.[54][57][58][59][60][61]2. 3. 사이토크롬 *bc*1 복합체와의 유사성
사이토크롬 *b*6*f* 복합체의 코어는 사이토크롬 *bc*1 복합체의 코어와 구조적으로 유사하다. 사이토크롬 *b*6와 소단위체 IV는 사이토크롬 *b*와 상동성이 있고,[62] 두 복합체의 리에스케 철-황 단백질은 상동성이다.[63] 그러나 사이토크롬 *f*와 사이토크롬 *c*1은 상동성이 없다.[64]사이토크롬 *b*6*f* 복합체에는 7개의 보결분자단이 포함되어 있다.[65][66] 이 중 4개는 사이토크롬 *b*6*f* 복합체와 사이토크롬 *bc*1 복합체 모두에서 발견되는데, 사이토크롬 *c*1과 *f*의 *c*형 헴, *bc*1과 *b*6*f*의 두 *b*형 헴(*b*p 및 *b*n), 리에스케 단백질의 [2Fe-2S] 클러스터이다. 사이토크롬 *b*6*f* 복합체에는 엽록소 *a*, β-카로틴, 헴 *c*n (헴 x라고도 함)이라는 3가지 독특한 보결분자단이 있다.[57]
사이토크롬 *b*6*f* 복합체 이량체의 코어 내의 단량체 간 공간은 지질에 의해 점유된다.[61]
2. 4. 보결분자단
사이토크롬 *b*6*f* 복합체는 7개의 보결분자단을 포함한다.[65][66] 이 중 4개는 사이토크롬 *b*6*f* 복합체와 사이토크롬 ''bc''1 복합체 모두에서 발견되는 사이토크롬 ''c''1과 사이토크롬 ''f''의 ''c''형 헴, ''bc''1과 *b*6*f*의 두 *b*형 헴(*b*p 및 *b*n), [2Fe-2S] 클러스터를 포함하는 리에스케 철-황 단백질이다. 사이토크롬 *b*6*f* 복합체에는 엽록소 ''a'', β-카로틴, 헴 *c*n (헴 x라고도 함)이라는 3가지 독특한 보결분자단이 추가로 존재한다.[57]2. 5. 단량체 간 공간
사이토크롬 *b*6*f* 복합체 이량체의 코어 내 단량체 간 공간은 지질로 채워져 있으며,[61] 이는 단백질 내 유전 환경 조절을 통해 헴-헴 전자 전달에 방향성을 제공한다.[67]3. 생물학적 기능
광합성에서 사이토크롬 *b*6*f* 복합체는 두 가지 주요 기능을 수행한다. 광계 II와 광계 I 사이에서 전자와 에너지를 전달하고, 엽록체의 스트로마에서 틸라코이드 막을 가로질러 틸라코이드 내강으로 양성자(H+)를 수송하여 ATP 합성을 촉진하는 양성자 기울기를 생성한다. 또한 환원된 페레독신으로부터 전자를 받을 NADP+가 없을 때 순환적 광인산화에 관여하여 P700+의 에너지로 ATP 합성에 사용될 수 있는 양성자 기울기를 형성한다. 이 순환은 탄소 고정에 필요한 ATP/NADPH 생성 비율을 적절하게 유지하는 데 도움을 준다. 사이토크롬 *b*6*f* 복합체 내의 p쪽 퀴놀 탈양성자화-산화 반응은 활성 산소 생성을 유발하며, 퀴놀 산화 부위에 위치한 필수 엽록소 분자는 활성 산소 생성 속도를 높이는 구조적, 비광화학적 기능을 수행하고 세포 내 통신을 위한 산화환원 경로를 제공한다.[71][72]
3. 1. 전자 및 에너지 전달 매개
광합성에서 사이토크롬 ''b''6''f'' 복합체는 광계 II와 광계 I 사이에서 전자와 에너지를 전달하는 기능을 한다. 이와 동시에 양성자(H+)를 엽록체의 스트로마에서 틸라코이드 막을 가로질러 틸라코이드 내강으로 수송한다.[54] 사이토크롬 ''b''6''f'' 복합체를 통한 전자 전달은 양성자 기울기를 생성하여 엽록체에서 ATP 합성을 촉진한다.[56]별도의 반응에서 사이토크롬 ''b''6''f'' 복합체는 환원된 페레독신으로부터 전자를 받을 NADP+가 없을 때 순환적 광인산화에서 중심적인 역할을 한다.[53] P700+의 에너지로 작동되는 이 순환은 ATP 합성에 사용될 수 있는 양성자 기울기를 형성하는 데 기여한다. 이 순환은 광합성에 필수적이며,[68] 탄소 고정에 필요한 ATP/NADPH 생성 비율을 적절하게 유지하는 데 도움을 준다.[69][70]
사이토크롬 ''b''6''f'' 복합체 내의 p쪽 퀴놀 탈양성자화-산화 반응은 활성 산소 생성과 관련이 있다.[71] 퀴놀 산화 부위에 위치한 필수 엽록소 분자는 활성 산소 생성 속도를 높이는 구조적, 비광화학적 기능을 수행하고 세포 내 통신을 위한 산화환원 경로를 제공하는 것으로 제안되었다.[72]
3. 2. 순환적 광인산화
NADP+가 환원된 페레독신으로부터 전자를 수용할 수 없을 때 사이토크롬 ''b''6''f'' 복합체는 순환적 광인산화에서 중심적인 역할을 한다.[53] P700+의 에너지에 의해 구동되는 이 순환은 ATP 합성을 추진하는 데 사용할 수 있는 양성자 기울기의 형성에 기여한다. 이 순환은 광합성에 필수적이며,[68] 탄소 고정을 위한 ATP/NADPH 생산의 적절한 비율을 유지하는 데 도움을 주는 것으로 나타났다.[69][70]3. 3. 활성 산소 생성
사이토크롬 ''b''6''f'' 복합체 내의 p쪽 퀴놀 탈양성자화-산화 반응은 활성 산소 생성과 관련되어 있다.[71] 퀴놀 산화 부위에 위치한 필수 엽록소 분자는 활성 산소 생성 속도를 향상시키는 구조적, 비광화학적 기능을 수행하여 세포 내 통신을 위한 산화환원 경로를 제공하는 것으로 제안되었다.[72]4. 반응 메커니즘
사이토크롬 *b*6*f* 복합체는 플라스토퀴놀(QH2)에서 플라스토사이아닌(Pc)으로 전자를 전달하는 반응을 촉매하며, 이 과정에서 스트로마에서 틸라코이드 내강으로 양성자를 펌핑한다.[53] 이 반응은 복합체 III에서와 같이 Q 회로를 통해 일어난다.[73]
구체적인 반응식은 다음과 같다.[42]
:QH2 + 2Pc(Cu2+) + 2H+ (스트로마) → Q + 2Pc(Cu+) + 4H+ (내강)
플라스토퀴놀은 전자 분기라고 불리는 메커니즘을 통해 2개의 전자를 고전위 및 저전위 전자전달계로 전달한다.[74] 사이토크롬 *b*6*f* 복합체에는 Q 회로의 작동, 광합성에서 산화환원 감지 및 촉매 기능을 담당하는 전자 전달 네트워크를 형성하는 최대 3개의 플라스토퀴논 분자가 포함되어 있다.[75]
4. 1. 비순환적 전자 흐름
사이토크롬 ''b''6''f'' 복합체는 두 가지 이동형 산화환원 운반체인 플라스토퀴놀(QH2)과 플라스토사이아닌(Pc) 사이의 전자 전달을 담당한다. 사이토크롬 ''b''6''f'' 복합체는 플라스토퀴놀로부터 플라스토사이아닌으로 전자를 전달하면서, 스트로마에서 틸라코이드 내강으로 양성자를 펌핑한다.[53] 반응식은 다음과 같다.:QH2 + 2Pc(Cu2+) + 2H+ (스트로마) → Q + 2Pc(Cu+) + 4H+ (내강)[42]
이 반응은 Q 회로를 통해 일어난다.[73] 플라스토퀴놀은 전자 분기 메커니즘을 통해 2개의 전자를 고전위 및 저전위 전자전달계로 전달한다.[74] 이 복합체에는 Q 회로의 작동과 광합성에서 산화환원 감지 및 촉매 기능을 담당하는 전자 전달 네트워크를 형성하는 최대 3개의 플라스토퀴논 분자가 포함되어 있다.[75]
4. 2. 순환적 전자 흐름
사이토크롬 *b*6*f* 복합체는 순환적 광인산화에서 핵심적인 전자 전달 반응을 촉매한다. 페레독신(Fd)의 전자는 플라스토퀴논으로 전달된 다음 사이토크롬 *b*6*f* 복합체로 전달되어 플라스토사이아닌을 환원시키며, 이는 광계 I의 P700에 의해 재산화된다.[76] 페레독신이 플라스토퀴논을 환원시키는 정확한 메커니즘은 아직 연구 중이다. 한 가지 제안은 페레독신:플라스토퀴논 환원효소 또는 NADP 탈수소효소가 존재한다는 것이다.[76] 헴 *x*는 Q 회로에 필요하지 않고 복합체 III에서도 발견되지 않기 때문에, 다음 메커니즘에 의해 순환적 광인산화에 사용되는 것으로 제안되었다.[74][77]# Fd (환원형) + 헴 *x* (산화형) → Fd (산화형) + 헴 *x* (환원형)
# 헴 *x* (환원형) + Fd (환원형) + Q + 2H+ → 헴 *x* (산화형) + Fd (산화형) + QH2
4. 3. Q 회로
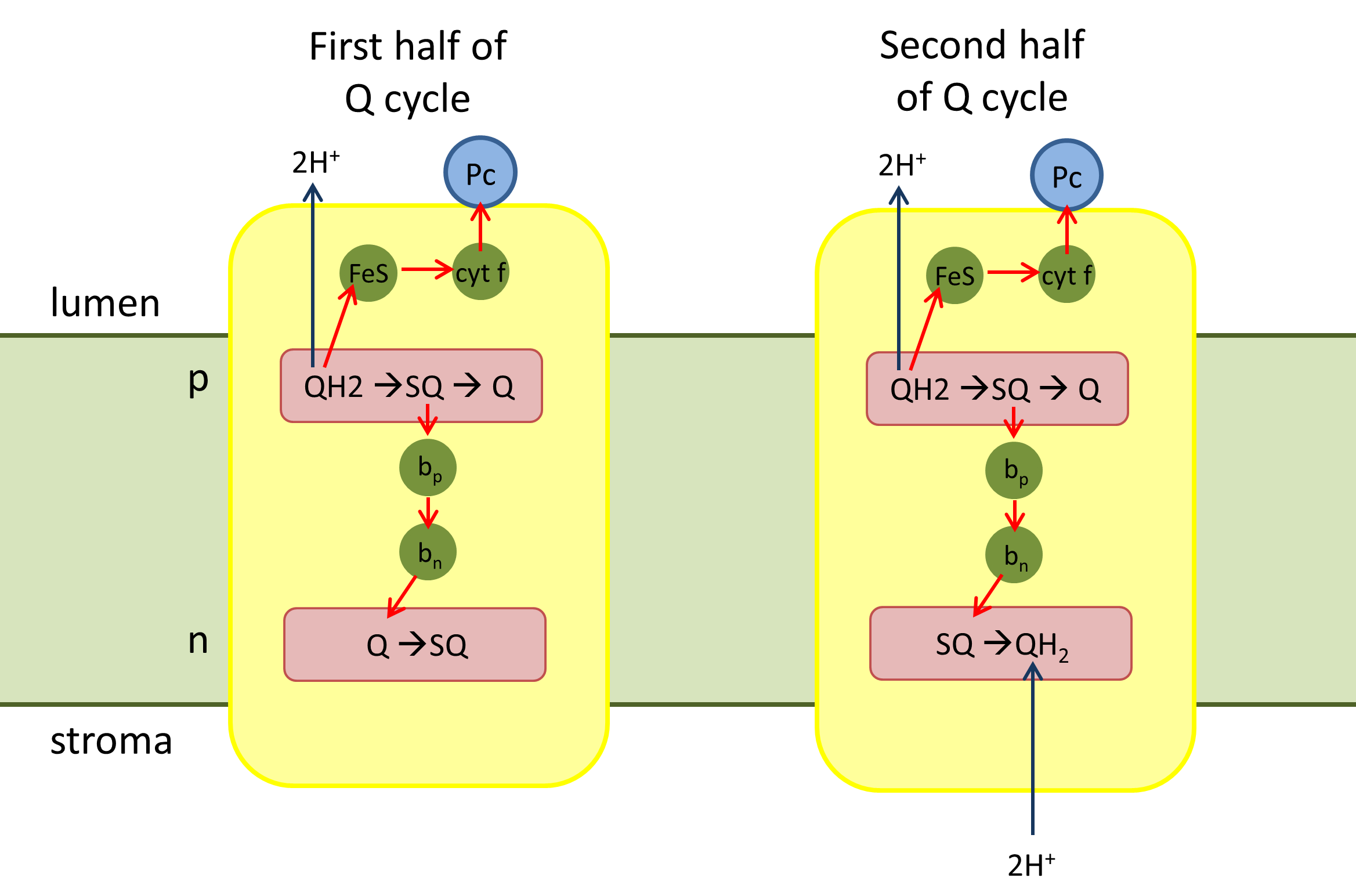
Q 회로는 사이토크롬 *b*6*f* 복합체에서 일어나는 전자와 양성자의 이동 경로를 설명하는 순환 과정이다. 이 과정은 크게 두 부분으로 나뉜다.
Q 회로 전반부에서는 플라스토퀴놀(QH2)이 산화되어 전자를 전달하고, 이 과정에서 양성자가 방출된다. 저전위 전자전달계에서는 전자가 순차적으로 이동하여 플라스토퀴논(Q)을 환원시킨다.
Q 회로 후반부에서는 두 번째 플라스토퀴놀(QH2)이 관여한다. 고전위 전자전달계에서는 전자가 플라스토사이아닌(Pc)을 환원시키고, 저전위 전자전달계에서는 전자가 이동하여 완전히 환원된 Q2-가 형성되고, 이는 다시 QH2로 전환된다. 최종적으로 산화된 Q와 환원된 QH2는 막으로 확산된다.[48][49]
4. 3. 1. Q 회로 전반부
# 플라스토퀴놀(QH2)이 복합체의 양성자 쪽('p' 쪽, 내강 쪽)에 결합한다.
# 플라스토퀴놀(QH2)은 철-황 중심에 의해 세미퀴논(SQ)으로 산화되고 2개의 양성자를 틸라코이드 내강으로 방출한다.
# 환원된 철-황 중심은 사이토크롬 ''f''를 통해 전자를 플라스토사이아닌(Pc)으로 전달한다.
# 저전위 전자전달계에서 세미퀴논(SQ)은 전자를 사이토크롬 ''b''6의 헴 ''b''p로 전달한다.
# 그런 다음 헴 ''b''p는 전자를 헴 ''b''n으로 전달한다.
# 헴 ''b''n은 하나의 전자로 플라스토퀴논(Q)을 환원하여 세미퀴논(SQ)을 형성한다.
4. 3. 2. Q 회로 후반부
# 두 번째 플라스토퀴놀(QH2)이 복합체에 결합한다.# 고전위 전자전달계에서 하나의 전자가 다른 산화된 플라스토사이아닌(Pc)을 환원시킨다.
# 저전위 전자전달계에서 헴 bn의 전자가 SQ로 전달되고, 완전히 환원된 Q2-는 스트로마로부터 두 개의 양성자를 받아 QH2를 형성한다.
# 산화된 Q와 재생된 환원형 QH2는 막 안으로 확산된다.
참조
[1]
웹사이트
ExplorEnz: EC 7.1.1.6
https://www.enzyme-d[...]
2023-12-24
[2]
논문
Quinone-dependent proton transfer pathways in the photosynthetic cytochrome b6f complex
2013-03-01
[3]
논문
Full subunit coverage liquid chromatography electrospray ionization mass spectrometry (LCMS+) of an oligomeric membrane protein: cytochrome b(6)f complex from spinach and the cyanobacterium Mastigocladus laminosus
2002-10-01
[4]
서적
Biochemistry
Wiley, J
[5]
논문
An atypical haem in the cytochrome b(6)f complex
2003-11-01
[6]
논문
Structure of the cytochrome b6f complex: quinone analogue inhibitors as ligands of heme cn
2007-06-01
[7]
논문
Structure-Function, Stability, and Chemical Modification of the Cyanobacterial Cytochrome b6f Complex from Nostoc sp. PCC 7120
2009-04-01
[8]
논문
Lipid-induced conformational changes within the cytochrome b6f complex of oxygenic photosynthesis
2013-04-01
[9]
논문
Internal lipid architecture of the hetero-oligomeric cytochrome b6f complex
2014-07-01
[10]
논문
Sequence homology and structural similarity between cytochrome b of mitochondrial complex III and the chloroplast b6-f complex: position of the cytochrome b hemes in the membrane
1984-02-01
[11]
논문
Biological identity and diversity in photosynthesis and respiration: structure of the lumen-side domain of the chloroplast Rieske protein
1997-12-01
[12]
논문
Crystal structure of chloroplast cytochrome f reveals a novel cytochrome fold and unexpected heme ligation
1994-02-01
[13]
논문
Structure-function of the cytochrome b6f complex
[14]
논문
Evolution of photosynthesis: time-independent structure of the cytochrome b6f complex
2004-05-01
[15]
논문
A map of dielectric heterogeneity in a membrane protein: the hetero-oligomeric cytochrome b6f complex
2014-06-01
[16]
서적
Biochemistry
https://archive.org/[...]
W.H. Freeman
[17]
논문
Cyclic electron flow around photosystem I is essential for photosynthesis
2004-06-01
[18]
서적
Molecular mechanisms of photosynthesis
Blackwell Science
[19]
논문
Cyclic photophosphorylation and electron transport
[20]
논문
Mechanism of enhanced superoxide production in the cytochrome b(6)f complex of oxygenic photosynthesis
2013-12-01
[21]
논문
Traffic within the cytochrome b6f lipoprotein complex: gating of the quinone portal
2014-10-01
[22]
논문
Some New Structural Aspects and Old Controversies Concerning the Cytochrome b6f Complex of Oxygenic Photosynthesis
1996-06-01
[23]
논문
Transmembrane traffic in the cytochrome b6f complex
[24]
논문
Cryo-EM Structure of the Spinach Cytochrome B 6 F Complex at 3.6 Å Resolution
http://eprints.white[...]
2019-11-01
[25]
논문
Cyclic electron transfer in plant leaf
2002-07-01
[26]
논문
Structure of the cytochrome b6f complex: new prosthetic groups, Q-space, and the 'hors d'oeuvres hypothesis' for assembly of the complex
[27]
웹사이트
ExplorEnz: EC 7.1.1.6
https://www.enzyme-d[...]
2023-12-24
[28]
논문
Quinone-dependent proton transfer pathways in the photosynthetic cytochrome b6f complex
2013-03-01
[29]
논문
Full subunit coverage liquid chromatography electrospray ionization mass spectrometry (LCMS+) of an oligomeric membrane protein: cytochrome b(6)f complex from spinach and the cyanobacterium Mastigocladus laminosus
2002-10-01
[30]
서적
Biochemistry
Wiley, J
[31]
논문
An atypical haem in the cytochrome b(6)f complex
2003-11-01
[32]
논문
Structure of the cytochrome b6f complex: quinone analogue inhibitors as ligands of heme cn
2007-06-01
[33]
논문
Structure-Function, Stability, and Chemical Modification of the Cyanobacterial Cytochrome b6f Complex from Nostoc sp. PCC 7120
2009-04-01
[34]
논문
Lipid-induced conformational changes within the cytochrome b6f complex of oxygenic photosynthesis
2013-04-01
[35]
논문
Internal lipid architecture of the hetero-oligomeric cytochrome b6f complex
2014-07-01
[36]
논문
Sequence homology and structural similarity between cytochrome b of mitochondrial complex III and the chloroplast b6-f complex: position of the cytochrome b hemes in the membrane
1984-02
[37]
논문
Biological identity and diversity in photosynthesis and respiration: structure of the lumen-side domain of the chloroplast Rieske protein
1997-12
[38]
논문
Crystal structure of chloroplast cytochrome f reveals a novel cytochrome fold and unexpected heme ligation
1994-02
[39]
논문
Structure-function of the cytochrome b6f complex
[40]
논문
Evolution of photosynthesis: time-independent structure of the cytochrome b6f complex
2004-05
[41]
논문
A map of dielectric heterogeneity in a membrane protein: the hetero-oligomeric cytochrome b6f complex
2014-06
[42]
서적
Biochemistry
https://archive.org/[...]
W.H. Freeman
[43]
논문
Cyclic electron flow around photosystem I is essential for photosynthesis
2004-06
[44]
서적
Molecular mechanisms of photosynthesis
Blackwell Science
[45]
논문
Cyclic photophosphorylation and electron transport
[46]
논문
Mechanism of enhanced superoxide production in the cytochrome b(6)f complex of oxygenic photosynthesis
2013-12
[47]
논문
Traffic within the cytochrome b6f lipoprotein complex: gating of the quinone portal
2014-10
[48]
논문
Some New Structural Aspects and Old Controversies Concerning the Cytochrome b6f Complex of Oxygenic Photosynthesis
1996-06
[49]
논문
Transmembrane traffic in the cytochrome b6f complex
[50]
논문
Cryo-EM Structure of the Spinach Cytochrome B 6 F Complex at 3.6 Å Resolution
http://eprints.white[...]
2019-11
[51]
논문
Cyclic electron transfer in plant leaf
2002-07
[52]
논문
Structure of the cytochrome b6f complex: new prosthetic groups, Q-space, and the 'hors d'oeuvres hypothesis' for assembly of the complex
[53]
서적
Biochemistry
https://archive.org/[...]
W.H. Freeman
[54]
논문
Quinone-dependent proton transfer pathways in the photosynthetic cytochrome b6f complex
2013-03
[55]
논문
Full subunit coverage liquid chromatography electrospray ionization mass spectrometry (LCMS+) of an oligomeric membrane protein: cytochrome b(6)f complex from spinach and the cyanobacterium Mastigocladus laminosus
2002-10
[56]
서적
Biochemistry
Wiley, J
[57]
논문
An atypical haem in the cytochrome b(6)f complex
2003-11
[58]
논문
Structure of the cytochrome b6f complex: quinone analogue inhibitors as ligands of heme cn
2007-06
[59]
논문
Structure-Function, Stability, and Chemical Modification of the Cyanobacterial Cytochrome b6f Complex from Nostoc sp. PCC 7120
2009-04
[60]
논문
Lipid-induced conformational changes within the cytochrome b6f complex of oxygenic photosynthesis
2013-04
[61]
논문
Internal lipid architecture of the hetero-oligomeric cytochrome b6f complex
2014-07
[62]
논문
Sequence homology and structural similarity between cytochrome b of mitochondrial complex III and the chloroplast b6-f complex: position of the cytochrome b hemes in the membrane
1984-02
[63]
논문
Biological identity and diversity in photosynthesis and respiration: structure of the lumen-side domain of the chloroplast Rieske protein
1997-12
[64]
논문
Crystal structure of chloroplast cytochrome f reveals a novel cytochrome fold and unexpected heme ligation
1994-02
[65]
논문
Structure-function of the cytochrome b6f complex
[66]
논문
Evolution of photosynthesis: time-independent structure of the cytochrome b6f complex
2004-05
[67]
논문
A map of dielectric heterogeneity in a membrane protein: the hetero-oligomeric cytochrome b6f complex
2014-06
[68]
논문
Cyclic electron flow around photosystem I is essential for photosynthesis
2004-06
[69]
서적
Molecular mechanisms of photosynthesis
https://archive.org/[...]
Blackwell Science
[70]
논문
Cyclic photophosphorylation and electron transport
[71]
저널
Mechanism of enhanced superoxide production in the cytochrome b(6)f complex of oxygenic photosynthesis
2013-12-01
[72]
저널
Traffic within the cytochrome b6f lipoprotein complex: gating of the quinone portal
2014-10-01
[73]
저널
Some New Structural Aspects and Old Controversies Concerning the Cytochrome b6f Complex of Oxygenic Photosynthesis
1996-06-01
[74]
저널
Transmembrane traffic in the cytochrome b6f complex
https://archive.org/[...]
[75]
저널
Cryo-EM Structure of the Spinach Cytochrome B 6 F Complex at 3.6 Å Resolution
http://eprints.white[...]
2019-11-01
[76]
저널
Cyclic electron transfer in plant leaf
2002-07-01
[77]
저널
Structure of the cytochrome b6f complex: new prosthetic groups, Q-space, and the 'hors d'oeuvres hypothesis' for assembly of the complex
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com