호모 하빌리스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
호모 하빌리스는 '손재주 좋은 사람'이라는 뜻으로, 약 280만 년 전에서 165만 년 전까지 아프리카에 서식했던 초기 인류 종이다. 1959년 탄자니아 올두바이 협곡에서 화석이 발견되었으며, 도구를 사용하고 뇌 용량이 오스트랄로피테쿠스보다 컸다는 특징을 보인다. 호모 하빌리스는 초기 구석기 시대의 올두바이 석기 문화를 대표하며, 육상 생활에 적응한 신체적 특징과 잡식성 식단을 가졌을 것으로 추정된다.
더 읽어볼만한 페이지
- 1964년 기재된 화석 분류군 - 란톈 원인
란톈 원인은 1963년 중국 람톈현에서 발견된 호모 에렉투스 화석 인류로, 하악골 및 두개골 화석을 통해 약 165만 년에서 68만 4천 년 전에 생존했으며, 중국을 인류학 연구의 중심지로 부상시키고 아시아 호모 에렉투스 연구에 기여했다. - 1964년 기재된 화석 분류군 - 넓은이빨청상아리
넓은이빨청상아리는 마이오세에서 플라이오세 시기에 전 세계 바다에 서식했던 멸종된 악상어목 상어로, 현생 청상아리와 유사하나 톱니가 있는 이빨을 가졌으며 고래, 해양 포유류 등을 먹이로 삼았을 것으로 추정되고 넓은이빨 형태는 백상아리의 조상으로 여겨지지만 속의 분류는 불확실하다. - 전기 구석기 시대 - 호모 에렉투스
호모 에렉투스는 약 200만 년 전부터 10만 년 전까지 생존하며 직립 보행을 하고 아슐리안 석기를 제작했으며 불을 사용한 최초의 인류로 추정되는 종이다. - 전기 구석기 시대 - 올도완 문화
올도완 문화는 약 330만 년 전부터 25만 년 전까지 아프리카를 중심으로 아시아와 유럽에 걸쳐 간단한 석기를 제작하고 사용한 초기 석기 시대 문화로, 호미닌의 기술 발전과 사회적 행동을 보여주는 중요한 유물을 남겼다. - 1964년 기재된 포유류 - 작은붉은칼루타
작은붉은칼루타는 오스트레일리아 서부에 서식하며 붉은 갈색 털을 가진 주머니고양이과의 소형 육식 동물로, 스피니펙스 초원에서 곤충과 작은 척추동물을 먹고 수컷은 번식기 직후 사망하는 특징을 가진다. - 1964년 기재된 포유류 - 온두라스과일먹는박쥐
온두라스과일먹는박쥐는 중앙아메리카 열대 지역에 서식하며 무화과를 주식으로 하는 야행성 박쥐로, 반향정위를 통해 먹이를 찾고 씨앗을 퍼뜨려 생태계 유지에 기여하며, IUCN 적색 목록에서는 정보 부족 종으로 분류된다.
| 호모 하빌리스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Homo habilis |
| 명명자 | Leakey 외, 1964년 |
| 멸종 여부 | 멸종 |
| 이명 | Australopithecus habilis (우드 및 콜라드, 1999) Homo microcranous (Ferguson, 1995) Homo gautengensis? (커노, 2010) Homo rudolfensis? (알렉세프, 1986) |
| 뇌 용량 | 500~900 cm³ |
| 키 | 100–120 cm |
| 몸무게 | 20–37 kg |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 목 | 영장목 (Primates) |
| 아목 | 진원아목(Haplorhini) |
| 하목 | 협비원아목(Catarrhini) |
| 상과 | 사람상과(Hominoidea) |
| 과 | 사람과(Hominidae) |
| 아과 | 사람아과(Homininae) |
| 족 | 사람족(Hominini) |
| 아족 | 사람아족(Hominina) |
| 속 | 사람속(Homo) |
| 시간 척도 | |
| 화석 범위 | 신생대제4기홍적세 전기, 230만~165만 년 전 |
| 설명 | |
| 평균 키 (여성) | 약 148 cm |
| 평균 체중 (여성) | 약 35 kg |
2. 이름
호모 하빌리스는 '손재주 좋은 사람', '손을 쓸 줄 아는 사람', '도구를 사용하는 사람'이라는 뜻이다. 북한에서는 문화어로 '손쓴사람'으로도 부른다.
1959년 루이스 리키와 메리 리키는 탄자니아 올두바이 협곡에서 돌도끼와 사슴, 영양의 뼈를 인위적으로 간 흔적을 발견하였다. 처음에는 진잔트로푸스가 이 석기들을 사용했을 것이라 생각했지만, 더 큰 두개골이 발견되면서 리키 부부는 생각을 바꾸었다.[5]
1959년 루이스 리키와 메리 리키는 탄자니아 올두바이 협곡에서 돌도끼와 사슴, 영양의 뼈를 인위적으로 간 흔적을 발견하였다. 루이스 리키와 메리 리키는 처음에 그곳에서 발견된 진잔트로푸스가 이 석기들을 사용했으리라 예상했다. 그러나 1959년 올두바이 계곡에서 더 큰 두개골이 발견되면서, 리키 부부는 더 큰 두뇌를 가진 인간이 도구를 사용했다고 생각을 바꾸었다.[17]
호모 하빌리스는 다른 사람속과 비교했을 때, 눈에 흰자가 없고, 땀샘이 발달하지 않았다는 특징이 있었다. 남성과 여성 간의 신체 차이, 즉 성별에 따른 차이가 거의 없어서 남녀 모두 비슷한 체구를 가졌다. 여성의 질은 그리 좋지 않았고, 팔이 다리보다 길었다. 키는 약 130cm에서 160cm 사이였으며, 뇌 용량은 약 600cc에서 670cc 사이였다.[67][68][69]
일반적으로 ''호모 에르가스터''/''호모 에렉투스''는 최초로 일부일처제 사회에서 살았던 인류로 여겨지며, 그 이전의 모든 호미닌은 일부다처제였다고 여겨진다. 하지만 초기 호미닌의 집단 역학을 확신을 가지고 추측하기는 매우 어렵다.[41] 초기 호미닌의 해부학적 구조가 화석 기록에 잘 나타나 있지 않기 때문에 성적 이형성의 비율을 결정하기 어렵다. 짝짓기 시스템은 치아 해부학을 기반으로 하지만, 초기 호미닌은 현대 영장류에서 함께 나타나지 않는 다양한 특징을 가지고 있다.[41]
3. 발견 과정
1959년부터 올두바이 계곡에서 4명의 어린아이 두개골과 뼈 조각이 발견되었다. 이 화석들은 뇌 용량이 500~600cc로, 오스트랄로피테쿠스 아프리카누스와는 다른 종임을 보여주었다. 루이스 리키는 레이먼드 다트와 협의하여 이 종을 '호모 하빌리스'라고 명명했다. '손을 쓸 줄 아는 사람'이라는 뜻이다.[3]
그러나 발견된 화석이 어린아이의 것이었고, 오스트랄로피테쿠스 아프리카누스와 비슷하다는 비판이 있었다. 루이스 리키는 이를 반박하기 위해 노력했고, 케냐와 탄자니아에서 추가적인 화석들이 발견되었다.
1972년 케냐 투르카나 호수(루돌프 호수)에서 리처드 리키가 800cc 용량의 성인 남성 두개골 화석을 발견하면서 호모 하빌리스가 정식으로 인정받게 되었다.[7] 이후 투르카나 호수에서 발견된 화석들은 호모 루돌펜시스로 재분류되었다.[7]
이후 호모 하빌리스의 화석은 탄자니아, 케냐, 에티오피아, 남아프리카 등에서 발견되었다.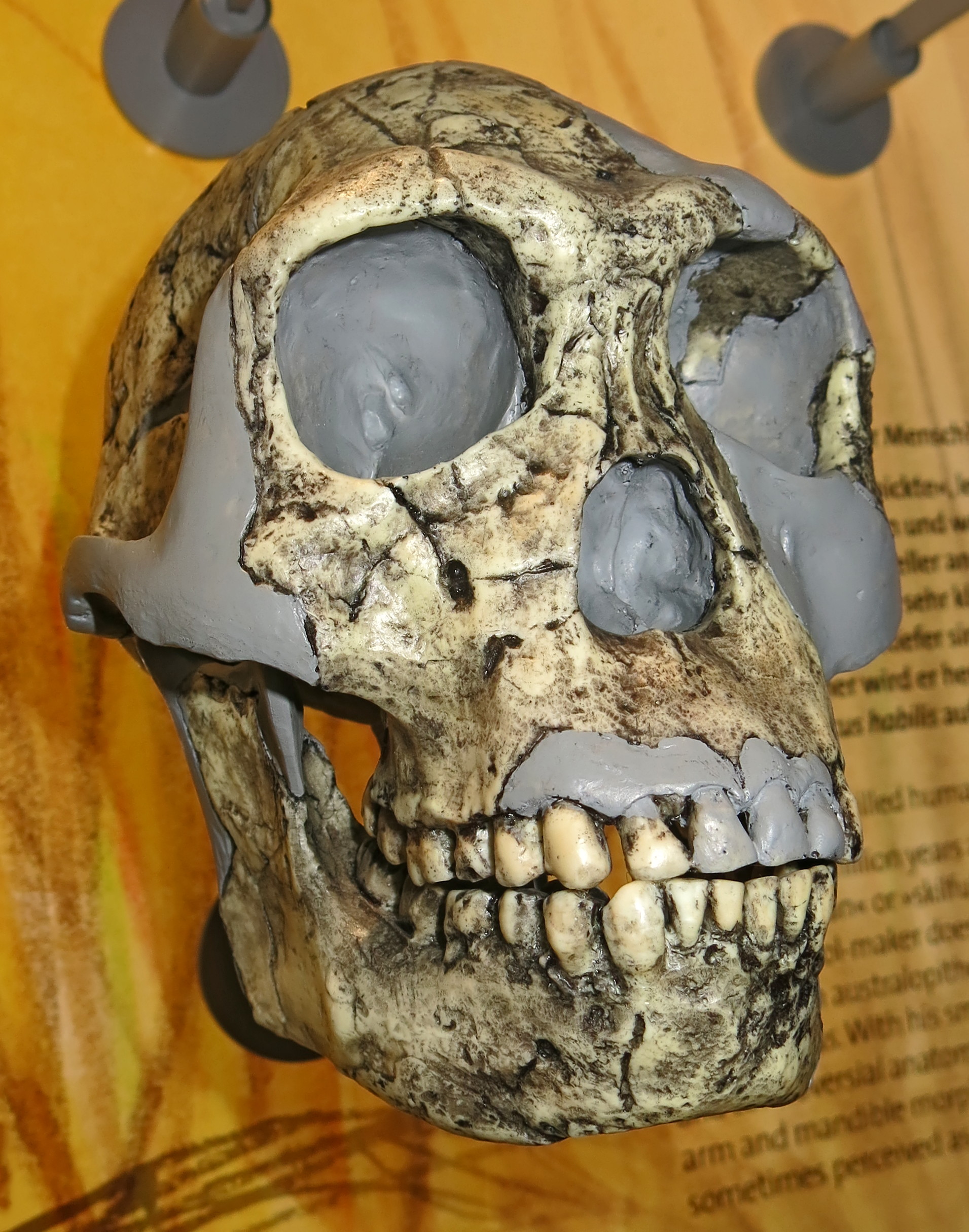
최초로 인정된 유해는 OH 7로, 1960년 탄자니아 올두바이 협곡에서 조나단 리키에 의해 발견되었다.[5] 1964년, 루이스 리키, 필립 V. 토비아스, 존 R. 네이피어는 공식적으로 이 유해를 호모 속에 할당하고, 레이먼드 다트의 권고에 따라 ''H. habilis''라는 종명을 부여했다.[3]
4. 분류
1959년부터 올두바이 조지 계곡에서 4명의 어린아이의 두개골과 뼈조각이 발견되었다. 이 화석들은 뇌 용량이 500~600cc였는데, 이는 440~500cc의 뇌용량을 가진 오스트랄로피테쿠스 아프리카누스와는 다른 종임을 보여주었다. 루이스 리키는 레이먼드 다트와 협의하여 이 종을 호모 하빌리스라 명명했다. 이는 손을 쓸 줄 아는 사람, 도구를 사용하는 사람이라는 뜻이다.[17]
그러나 발견된 화석이 어린아이의 것이었고, 오스트랄로피테쿠스 아프리카누스와 생김새가 비슷하다는 비판이 있었다. 루이스 리키는 이를 반박하기 위해 노력했고, 케냐와 탄자니아에서 추가 화석이 발견되었다.[17]
1972년 케냐의 투르카나 호수(루돌프 호수) 동부에서 리처드 리키 등에 의해 800cc 용량의 성인 남성 두개골 화석이 발견되면서 호모 하빌리스가 정당한 인종으로 인정받게 되었다. 이후 투르카나 호수에서 발견된 화석들은 호모 루돌펜시스로 재분류되었다.[17]
이후 호모 하빌리스의 화석은 탄자니아, 케냐, 에티오피아, 남아프리카 등에서 발견되었다.[17]
''H. habilis''가 ''H. ergaster''/''H. erectus''의 조상인지, 아니면 인류 계통에서 갈라져 나온 종인지에 대해서는 아직 논란이 있다.[18] ''H. habilis''로 분류된 모든 표본이 정확하게 분류된 것인지, 아니면 이 종이 다양한 ''오스트랄로피테쿠스'' 및 ''호모'' 종의 집합인지에 대해서도 논란이 있다.[19] ''H. habilis''의 치아 형태는 후기 ''호모'' 종보다 ''오스트랄로피테쿠스''와 더 유사한 특징을 공유한다.[20]
현재 ''호모''가 ''오스트랄로피테쿠스''에서 진화했다는 데에는 대체로 동의하지만, LD 350-1의 발견은 아파르 지구에서 이루어졌으며, 이 시기에 속이 ''A. afarensis''에서 진화했을 수 있음을 시사한다. 이 표본은 처음에 ''호모'' 종으로 분류되었지만,[22] 후속 연구에 따르면 ''오스트랄로피테쿠스''와도 특징을 공유하며, ''H. habilis''와는 명확히 구별된다.[23][24] 가장 오래된 ''H. habilis'' 표본인 A.L. 666-1은 230만 년 전으로 거슬러 올라가지만, 더 어린 OH 7보다 해부학적으로 더 파생된 특징을 보여주어, ''H. habilis'' 계통이 230만 년 전 이전에 시작되었음을 시사한다.[28]
5. 신체적 특징
오스트랄로피테쿠스 아파렌시스를 발견하여 유명해진 도널드 요한슨은 1986년 올두바이 조지에서 10대 후반에서 20대 초반으로 추정되는 여성의 골격을 발견했는데, 이 골격은 팔이 길고 다리가 짧았으며 손가락과 발가락이 굽어 있었다. 그러나 이 개체군은 다른 지역에서 발견된 호모 하빌리스 화석들보다 유난히 작고 팔다리가 굽은 특징을 보였다.
2013년 조지아에서 완전한 형태의 두개골이 발견되어 Dmanisi(D4600)와 D2600 두개골들을 분석한 결과, 이들의 두개골 용적은 546cc로 나타났다. 이는 호모 하빌리스, 호모 에렉투스 등 다른 호모 화석들과의 연관성을 보여주며, 현생 인류까지의 진화를 직접적으로 보여주는 증거가 되었다.[67]
일반적으로 뇌의 크기는 종 간의 전환기에 빠르게 증가하는 경향을 보였다. ''H. habilis''의 뇌 크기는 ''H. ergaster''/''H. erectus''보다 작았고, ''H. habilis''에서 약 600cc에서 650cc였던 것이 ''H. ergaster''와 ''H. erectus''에서는 약 900cc에서 1000cc로 급증했다.[28][26] 2015년 연구에서는 OH 7의 뇌 용적을 729cc에서 824cc로 재평가하면서 ''H. habilis'', ''H. rudolfensis'', 그리고 ''H. ergaster''의 뇌 크기가 일반적으로 500cc에서 900cc 사이였다고 밝혔다.[28] 이는 오스트랄로피테쿠스의 뇌 크기(약 400cc에서 500cc)에서 크게 증가한 것이다.[26]
모든 ''호모''의 뇌는 오스트랄로피테쿠스에 비해 대뇌가 팽창된 특징을 보인다. OH 65의 치아에 있는 줄무늬 패턴은 오른쪽으로 기울어져 있는데, 이는 오른손으로 돌 도구를 사용하여 고기를 자르려고 하는 동안 왼손과 치아로 고기를 잡아당기면서 실수로 생긴 상처일 수 있다. 이것이 사실이라면, 오른손잡이였음을 나타낼 수 있으며, 이는 뇌의 주요 재구성과 좌우 반구 간의 뇌 기능의 측면화와 관련이 있다. 이러한 시나리오는 일부 네안데르탈인 표본에서도 가설로 제기되었다. 뇌의 측면화는 도구 사용과 관련이 있을 수 있으며, 현대 인류의 경우 언어와도 약하게 관련되어 있다.[27]
''H. habilis''의 치열은 나중에 ''호모''에서 U자형인 것과 달리 V자형이었고, 얼굴은 위에서 코까지 평평했지만 입은 바깥쪽으로 돌출되어 있었다(전안면).[28]
OH 62(암컷으로 추정)와 KNM-ER 3735(수컷으로 추정)의 골격을 바탕으로, ''H. habilis''는 초기 ''A. afarensis''보다도 유인원과 더 유사하며, 오스트랄로피테신처럼 적어도 부분적으로 수목 생활을 했을 것으로 보인다. OH 62를 기준으로 하고 오스트랄로피테신과 비슷한 신체 치수를 가정하면, ''H. habilis''는 일반적으로 오스트랄로피테신처럼 몸집이 작았을 것으로 해석되며, OH 62는 키가 100cm에서 120cm 사이, 몸무게가 20kg에서 37kg 사이로 추정된다. 그러나 더 길고 현대 인류와 같은 다리를 가정하면 OH 62는 약 148cm이고 35kg이며, KNM-ER 3735도 거의 같은 크기였을 것이다.[29] ''H. ergaster'' 이전의 호미닌( ''H. habilis'' 포함)은 수컷이 암컷보다 현저히 큰 성적 이형성을 보였다고 일반적으로 추정되지만, 이 종에서 암컷의 상대적 체질량은 알려져 있지 않다.[31]
''H. habilis''를 포함한 초기 호미닌은 현대의 비인간 유인원처럼 털이 두껍게 덮여 있었을 것으로 생각된다. 이는 이들이 더 추운 지역에 거주했을 것으로 보이며, (털이 없는 것으로 추정되는) ''ergaster'' 이후 종보다 덜 활동적인 생활 방식을 가졌을 것으로 여겨지기 때문이다. 따라서 따뜻함을 유지하기 위해 두꺼운 체모가 필요했을 것이다.[32] 치아 발달 속도를 기준으로, ''H. habilis''는 현대 인류보다 성장 속도가 빨랐으며, 현대의 비인간 유인원과 더 유사했을 것으로 추정된다.[33]
''H. habilis''와 오스트랄로피테쿠스의 팔은 비율상 길어 나무 타기 및 매달리기에 적합한 것으로 여겨져 왔다.[34][35][36] 2004년, 인류학자 마틴 하우스러(Martin Haeusler)와 헨리 맥헨리는 OH 62의 상완골 대 대퇴골 비율이 현생 인류의 변동 범위 내에 있고, KNM-ER 3735가 현생 인류 평균에 근접하기 때문에 유인원과 같은 비율로 추정하는 것은 안전하지 않다고 주장했다. 그럼에도 OH 62의 상완골은 258mm에서 270mm 사이였고, 척골(전완)은 245mm에서 255mm 사이로, 침팬지와 유사한 비율을 보였다. OH 7의 손뼈는 정밀한 잡기 능력을 시사하며, 이는 손재주에 중요하며, 나무 타기에 대한 적응을 보여준다. 대퇴골과 관련하여, 전통적으로 ''A. afarensis'' 표본 AL 288-1과의 비교를 통해 ''H. habilis''의 튼튼한 다리를 재구성해왔지만, 하우스러와 맥헨리는 더 가늘어진 OH 24 대퇴골 (''H. ergaster'' / ''H. erectus'' 또는 ''P. boisei''에 속할 수 있음)이 더 적절한 비교 대상일 수 있다고 제안했다. 이 경우 ''H. habilis''는 더 길고 인간적인 다리를 가지고, ''H. ergaster''처럼 장거리 이동에 효과적이었을 것이다.[37] 그러나 화석의 보존되지 않은 길이를 추정하는 것은 매우 문제가 많다. OH 62의 사지 뼈 두께는 ''H. ergaster'' / ''H. erectus'' 및 현생 인류보다 침팬지와 더 유사하며, 이는 ''H. habilis''의 수목 생활에 더 적합한 다른 하중 지지 능력을 나타낼 수 있다.[38] OH 35의 튼튼한 비골 (이것은 ''P. boisei''에 속할 수 있음)은 비인간 유인원과 더 유사하며, 수목 생활 및 수직 등반과 일치한다.[39]
OH 8 (발)은 ''A. afarensis''의 발보다 육상 이동에 더 적합하지만, 여전히 나무 타기와 일치하는 많은 유인원과 같은 특징을 유지하고 있다.[37] 그러나 발에는 돌출된 발가락뼈와 압축된 발 중앙 관절 구조가 있어 발과 발목 사이, 그리고 발 앞부분에서의 회전을 제한한다. 발의 안정성은 다리와 발 사이의 힘 전달 효율성을 높이고 그 반대의 경우도 마찬가지이며, 달리기를 할 때 에너지를 생성하는 족궁 탄성 스프링 메커니즘과 관련이 있다 (걷는 것은 아님). 이것은 ''H. habilis''가 어느 정도의 지구력 달리기 능력이 있었을 수 있음을 나타낼 수 있으며, 이는 일반적으로 ''H. ergaster'' / ''H. erectus''에서 더 나중에 진화한 것으로 여겨진다.[40]
6. 문화
올두바이 협곡의 FLK Zinj 및 PTK 유적지에서 도구와 가공된 동물 뼈의 공간적 분포는 거주자들이 이 지역을 현대 수렵 채집인의 핵가족 시스템과 달리, 공동 도축 및 식사 장소로 사용했음을 나타낸다.[42]
''호모 하빌리스''를 포함한 초기 ''호모''의 행동은 때때로 사바나 침팬지와 비비의 행동을 모델로 삼는다. 이러한 공동체는 위험하고 노출된 서식지에서 그룹을 방어하기 위해 여러 수컷으로 구성되며(하렘 사회와 반대), 때로는 적과 포식자를 상대로 돌과 막대기를 던지는 집단 시위를 벌이기도 한다.[43] 발 OH 8은 악어에 물린 것으로 보이며,[44] 아마도 ''Crocodylus anthropophagus''일 가능성이 있다.[45] 다리 OH 35는 ''호모 하빌리스'' 또는 ''파란트로푸스 보이세이''에 속하며 표범의 포식 흔적을 보인다.[44] ''호모 하빌리스''와 동시대의 호미닌은 (올두바이 협곡에서) 사냥 하이에나 ''Chasmaporthetes nitidula''와 검치호랑이 ''Dinofelis'' 및 ''Megantereon''과 같은 당시의 다른 대형 육식 동물에게 포식되었을 가능성이 높다.[46] 1993년, 미국 고인류학자 레슬리 C. 아이엘로와 영국 진화 심리학자 로빈 던바는 현대 비인간 영장류의 신피질 크기와 그룹 크기에서 보이는 경향을 바탕으로 ''호모 하빌리스''의 그룹 크기가 70~85명으로 추정했다. 이는 침팬지와 비비의 그룹 크기 상한선에 해당한다.[47]
''호모 하빌리스''는 ''호모 루돌펜시스'', ''호모 에르가스터''/''호모 에렉투스'', ''파란트로푸스 보이세이''와 공존했다. 이 모든 종이 어떻게 상호 작용했는지는 불분명하다.[5][48][49]
1962년, 올두바이 협곡에서 화산암으로 만들어진 366cm 크기의 원이 발견되었다. 61cm 간격으로 암석들이 15cm 높이로 쌓여 있었다. 메리 리키는 암석 더미가 땅에 박힌 기둥을 지지하는 데 사용되었으며, 아마도 방풍벽이나 대충 지은 오두막을 지지하기 위한 것이라고 추측했다. 오늘날 일부 유목 부족들은 비슷한 낮은 암석 벽을 쌓아 임시 쉼터를 짓고, 기둥으로 곧게 뻗은 나뭇가지를 구부리고 풀이나 동물의 가죽을 스크린으로 사용한다.[61] 175만 년 전으로 거슬러 올라가며, 초기 ''호모''에게 기인하며, 건축의 가장 오래된 증거로 주장된다.[62]
6. 1. 석기
호모 하빌리스는 최초로 석기를 사용한 인류인 오스트랄로피테쿠스 가르히나 오스트랄로피테쿠스 아프리카누스보다 더욱 발달한 뗀석기를 만들었는데, 원시적인 찍개와 비슷한 도구를 사용하였을 것이다. 호모 하빌리스는 석기와 동물의 뼈를 이용해서 다양한 형태의 도구를 만들었다. 이것으로 직접 사냥을 하고 가죽과 뼈를 발라내고, 뼈를 깨트려 먹기 시작하였을 것이다.
''호모 하빌리스''는 초기 구석기 시대의 올두바이 석기 산업과 관련이 있다. 이들은 주로 동물을 도축하고 가죽을 벗기며 뼈를 부수는 데 이 도구들을 사용했을 것이지만, 때로는 나무를 톱질하고 긁거나 부드러운 식물을 자르기도 했을 것이다. 석공들은 돌을 깎아 만들었는데, 석핵을 신중하게 선택하고 특정 암석이 적절한 부위에 충분히 강하게 타격을 가하면 특정한 방식으로 깨진다는 것을 알고 있었다. 그들은 초퍼, 다면체, 원반형을 포함한 여러 종류의 도구를 제작했다. 하지만, 특정 모양을 미리 생각하지는 않았을 것이며, 이러한 도구를 생산하는 데 있어 표준화가 부족했고 석공들이 사용할 수 있는 원자재의 종류에서 비롯되었을 것이다.[2][57] 예를 들어, 구형은 올두바이에 흔하게 나타나는데, 이는 크고 부드러운 석영과 석영암 조각이 풍부하기 때문이다. 반면에 쿠비 포라에는 구형이 없고, 주로 단단한 현무암 용암 암석이 제공된다. 나중에 ''호모 에르가스터'' / ''호모 에렉투스''에 의해 발명된 아슐리안 문화와 달리, 올두바이 기술은 제조에 계획과 선견지명을 필요로 하지 않으므로, 올두바이 석공들의 높은 인지 능력을 나타내지는 않지만, 어느 정도의 협동과 기계에 대한 지식은 필요하다. 올두바이 도구는 재손질이 드물게 나타나며, 대부분 사용 직후에 버려졌을 것이다.[57]
올두바이는 1934년에 처음 보고되었지만, 1960년대에 이르러서야 180만 년 전으로 거슬러 올라가는 가장 초기 문화로, 그리고 ''호모 하빌리스''가 제작한 것으로 널리 받아들여졌다. 그 이후, 더 많은 발견들을 통해 물질 문화의 기원이 상당한 시간적으로 앞당겨졌는데,[2] 올두바이는 에티오피아의 레디-게라루와 고나에서 발견되었으며, 이는 260만 년 전으로 거슬러 올라가며, 아마도 속의 진화와 관련이 있을 것이다.[2][58] 오스트랄로피테신 또한 330만 년 전의 로메키 석기 산업과 같은 도구를 제작한 것으로 알려져 있으며,[59] 약 340만 년 전의 도축 증거도 있다.[60] 그럼에도 불구하고, 비교적 날카로운 가장자리를 가진 올두바이 문화는 오스트랄로피테신 기술에서 주요한 혁신이었으며, 변화하는 시대의 기후에서 유리했을 다양한 먹이 전략과 더 넓은 범위의 음식을 처리하는 능력을 허용했을 것이다.[58] 올두바이가 독립적으로 발명되었는지, 아니면 여러 종에 걸쳐 수십만 년 동안 호미닌이 암석을 가지고 실험한 결과인지는 불분명하다.[2]
6. 2. 사회
호모 하빌리스는 침팬지와 오스트랄로피테쿠스 등의 다른 유인원 및 영장류들보다 복잡한 사회 체제와 생각을 가졌다고 한다. 이들은 대체로 육식을 많이 했다. 루이스 리키 등은 호모 하빌리스가 적극적으로 사냥을 했다고 가정했으나, 이후 연구에서는 소형 동물을 사냥하거나 죽은 동물의 사체를 처리하는 청소부 역할을 하거나, 다른 동물의 사냥감을 빼앗기도 했던 것으로 확인되고 있다.
6. 3. 식습관
호모 하빌리스는 침팬지나 오스트랄로피테쿠스보다 복잡한 사회 체제를 가졌으며, 육식을 많이 했다고 알려져 있다.[41] 초기에는 적극적인 사냥을 했을 것으로 추정되었으나, 이후 연구에 따르면 소형 동물을 사냥하거나 죽은 동물의 사체를 처리하고, 다른 동물의 사냥감을 빼앗기도 했던 것으로 보인다.
호모 하빌리스는 과일도 중요한 식량원으로 섭취했는데, 이는 치아 부식에서 나타나는 산성 노출 흔적으로 알 수 있다.[52] 치아 미세 마모 분석 결과, 질긴 음식을 নিয়মিত(정기적) 섭취하지는 않았으며, 잎을 먹는 동물과 질긴 음식을 먹는 동물 중간 정도의 식습관을 가졌던 것으로 보인다.[53] 이는 호모 하빌리스가 점차 다양한 음식을 섭취하는 잡식성 식단으로 변화했음을 보여준다.[54] 또한, 고고학 유적지에서 발견된 어류 유해를 통해 민물고기도 섭취했을 가능성이 제기되었다.[55]
일반적으로 호모 하빌리스와 초기 호모 속(屬)은 오스트랄로피테쿠스보다 고기를 더 많이 섭취했으며, 이는 뇌 성장과 관련이 있다고 여겨진다. 고기는 에너지와 영양소가 풍부하여 인지 능력 발달에 영향을 주었거나, 크고 칼로리 소모가 많은 내장 기관을 줄여 뇌 성장에 더 많은 에너지를 사용할 수 있게 했다는 가설이 있다. 또는 건조한 기후에서 식량 부족에 대응하기 위해 지하 저장 기관(덩이줄기 등)과 식량 공유에 의존하면서 사회적 유대가 강화되었다는 주장도 있다.[56]
호모 하빌리스는 큰 앞니를 가졌는데, 이는 앞니를 사용하여 음식을 섭취하는 데 더 의존했음을 시사한다. 아래턱 몸체는 현대인이나 다른 유인원보다 두꺼워 씹는 힘에 잘 견딜 수 있었으며, 이는 질긴 음식을 부수는 도구를 사용했음을 보여준다.[31]
호모 하빌리스는 주로 사냥보다는 다른 동물이 남긴 시체를 먹거나, 자칼이나 치타와 같은 작은 포식자로부터 먹이를 훔치는 방식으로 고기를 얻었을 것으로 추정된다.[51]
7. 기타
1980년대 중반 시베리아에서 호모 하빌리스가 사용하던 돌도구와 비슷한 형태의 석기들이 발견되었으며, 연대는 180만년 전이었다. 이 때문에 호모 하빌리스가 아프리카 밖으로 이주했을 것이라는 추론이 제기되었으나, 인골 화석이 발견되지 않아 확증은 어렵다. 이들의 언어 사용 여부는 미지수이다.
참조
[1]
논문
Early ''Homo'': Who, When, and Where
[2]
논문
The origins of stone tool technology in Africa: a historical perspective
[3]
논문
A New Species of the Genus ''Homo'' from Olduvai Gorge
http://users.clas.uf[...]
[4]
논문
Human evolution: taxonomy and paleobiology
[5]
서적
The First Humans – Origin and Early Evolution of the Genus ''Homo''
Springer, Dordrecht
2009
[6]
논문
Hominid evolution in Africa
https://capa-acap.ne[...]
[7]
서적
Ancestors: The hard evidence
Alan R. Liss
[8]
논문
''Homo rudolfensis'' Alexeev, 1986: Fact or phantom?
[9]
논문
A review of early ''Homo'' in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (''Homo gautengensis'' sp. nov.)
https://www.scienced[...]
[10]
논문
New partial skeleton of ''Homo habilis'' from Olduvai Gorge, Tanzania
[11]
논문
The Human Genus
https://pdfs.semanti[...]
[12]
논문
Craniofacial variation in ''Homo habilis'': an analysis of the evidence for multiple species
[13]
논문
The species ''Homo habilis'': example of a premature discovery
[14]
서적
Handbook of Paleoanthropology
[15]
논문
Tooth wear and dentoalveolar remodeling are key factors of morphological variation in the Dmanisi mandibles
[16]
논문
A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo
[17]
서적
Handbook of Paleoanthropology
Springer
2020-06-12
[18]
논문
Classification and phylogeny in human evolution
http://www.ludus-vit[...]
2020-06-10
[19]
서적
Extinct Humans
Basic Books
[20]
논문
Dental morphology in Homo habilis and its implications for the evolution of early Homo
https://www.nature.c[...]
2024-11-20
[21]
서적
Handbook of Paleoanthropology
Springer
[22]
논문
Early ''Homo'' at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia
[23]
논문
Comment on “Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia”
[24]
논문
Ledi-Geraru strikes again: Morphological affinities of the LD 350-1 mandible with early ''Homo''
[25]
논문
Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago
[26]
논문
The brain of ''Homo habilis'': A new level of organization in cerebral evolution
[27]
논문
OH-65: The earliest evidence for right-handedness in the fossil record
[28]
논문
Reconstructed ''Homo habilis'' type OH 7 suggests deep-rooted species diversity in early ''Homo''
[29]
논문
Spatial and temporal variation of body size among early ''Homo''
[30]
논문
Human Height
https://ourworldinda[...]
2020-06-16
[31]
논문
Diet in Early ''Homo'': A Review of the Evidence and a New Model of Adaptive Versatility
[32]
논문
Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling
[33]
논문
Growth, Development, and Life History throughout the Evolution of ''Homo''
[34]
논문
Evolutionary reversals of limb proportions in early hominids? evidence from KNM-ER 3735 (''Homo habilis'')
[35]
논문
New partial skeleton of ''Homo habilis'' from Olduvai Gorge, Tanzania
1987-05-21
[36]
논문
Who is the 'real' ''Homo habilis''?
[37]
논문
Body Proportions of ''Homo Habilis''
[38]
논문
Relative Limb Strength and Locomotion in ''Homo habilis''
[39]
논문
Relative fibular strength and locomotor behavior in KNM-WT 15000 and OH 35
[40]
논문
Endurance running and the evolution of Homo
http://doc.rero.ch/r[...]
[41]
논문
Mating Behavior in ''Australopithecus'' and Early ''Homo'': A Review of the Diagnostic Potential of Dental Dimorphism
https://ir.lib.uwo.c[...]
[42]
논문
A spatial analysis of stone tools and fossil bones at FLK Zinj 22 and PTK I (Bed I, Olduvai Gorge, Tanzania) and its bearing on the social organization of early humans
[43]
논문
The social organization of ''Homo ergaster'': Inferences from anti-predator responses in extant primates
https://www.research[...]
2017
[44]
논문
Crocodylian and mammalian carnivore feeding traces on hominid fossils from FLK 22 and FLK NN 3, Plio-Pleistocene, Olduvai Gorge, Tanzania
[45]
논문
A New Horned Crocodile from the Plio-Pleistocene Hominid Sites at Olduvai Gorge, Tanzania
[46]
논문
The hunters and the hunted revisited
[47]
논문
Neocortex Size, Group Size, and the Evolution of Language
1993
[48]
논문
New fossils from Koobi Fora in northern Kenya confirm taxonomic diversity in early ''Homo''
[49]
논문
Implications of new early ''Homo'' fossils from Ileret, east of Lake Turkana, Kenya
[50]
논문
A new fossil skull from Olduvai
[51]
논문
Tree-stored leopard kills: expanding the hominid scavenging niche
[52]
논문
Acidic-Food Choice in ''Homo habilis'' at Olduvai
[53]
논문
Dental Evidence for the Reconstruction of Diet in African Early ''Homo''
2012-02-09
[54]
논문
Dental Microwear and Diets of African Early ''Homo''
2006-01-01
[55]
논문
Early hominid utilisation of fish resources and implications for seasonality and behaviour
https://www.scienced[...]
2024-02-04
[56]
논문
Ecological Energetics in Early ''Homo''
[57]
논문
The Oldowan reassessed: A close look at early stone artifacts
[58]
논문
Earliest known Oldowan artifacts at >2.58 Ma from Ledi-Geraru, Ethiopia, highlight early technological diversity
[59]
논문
3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya
https://www.research[...]
[60]
논문
Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia
[61]
서적
Olduvai Gorge: Volume 3, Excavations in Beds I and II, 1960–1963
https://books.google[...]
Cambridge University Press
[62]
서적
The Perception of the Environment: Essays on Livelihood, Dwelling and Skill
https://books.google[...]
Psychology Press
[63]
웹사이트
Homo habilis
http://humanorigins.[...]
Smithsonian Institution
2015-12-19
[64]
논문
Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya
http://salamander.uk[...]
2015-12-19
[65]
논문
Fifty years after Homo habilis
http://www.nature.co[...]
2015-12-19
[66]
문서
[67]
저널
A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo
https://www.science.[...]
[68]
저널
https://www.sciencedirect.com/science/article/abs/pii/0047248488900395
https://www.scienced[...]
[69]
저널
Technical Note: Virtual reconstruction of KNM-ER 1813 Homo habilis cranium
https://onlinelibrar[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com