메세노사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
메세노사우루스는 소형 단궁류의 일종으로, 주로 두개골 특징으로 분류된다. 가늘고 긴 전상악골이 좁고 직사각형의 주둥이를 형성하며, 콧구멍 선반, 콧구멍 경계, 안와후부 광대뼈 등 독특한 두개골 특징을 가진다. 굴을 파는 습성이 있었으며, 곤충이나 절지동물을 먹고 살았을 것으로 추정된다. 메세노사우루스 로메리와 메세노사우루스 에프레모비 두 종이 있으며, 페름기 시대에 유럽, 러시아, 미국 등지에서 번성했으나, 후기에 이궁류와의 경쟁으로 쇠퇴했다.
더 읽어볼만한 페이지
- 1938년 기재된 화석 분류군 - 파란트로푸스
파란트로푸스는 플라이오세 후기부터 홍적세 중기 아프리카에 살았던 멸종된 호미닌 속으로, 질긴 식물성 음식을 씹기에 적합한 튼튼한 두개골 구조를 지녔으며, 오스트랄로피테쿠스 및 초기 호모 속과 공존했고, 파란트로푸스 보이세이, 파란트로푸스 로부스투스, 파란트로푸스 에티오피쿠스 등의 종이 속한다. - 1938년 기재된 화석 분류군 - 파란트로푸스 로부스투스
파란트로푸스 로부스투스는 플라이스토세에 살았던 멸종된 호미닌의 일종으로, 남아프리카에서 화석이 발견되었으며, 튼튼한 턱과 큰 어금니를 가졌고, 잡식성이었으며 뼈 도구를 사용했을 가능성이 제기된다. - 러시아의 화석 - 글로소프테리스
글로소프테리스는 혀 모양의 잎과 그물맥을 가지는 멸종된 종자 식물로, 곤드와나 대륙에 널리 분포하며 대륙 이동설을 뒷받침하는 화석 증거가 되었다. - 러시아의 화석 - 큰뿔사슴
큰뿔사슴(Megaloceros giganteus)은 플라이스토세 후기부터 홀로세 초기까지 유라시아 대륙에 서식했던 멸종된 사슴으로, 수컷은 거대한 뿔을 가졌으며, 짝짓기, 사냥, 기후 변화 등의 요인으로 인해 멸종되었다. - 미국의 화석 - 트리케라톱스
트리케라톱스는 북아메리카에서 발견된 각룡류 공룡의 한 속으로, 거대한 두개골과 뿔이 특징이며 백악기 말기에 번성했고, 현재는 두 종으로 분류된다. - 미국의 화석 - 티라노사우루스
티라노사우루스는 백악기 후기 북아메리카에 서식했던 몸길이 15m에 달하는 거대한 육식 공룡으로, 강력한 턱힘과 짧은 앞다리, 긴 꼬리를 가진 최상위 포식자였으며, '수'와 '스카티' 등의 표본과 대중문화 속 인기 덕분에 그 생태와 행동에 대한 연구가 지속되고 있다.
| 메세노사우루스 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Mesenosaurus |
| 명명자 | Efremov, 1938 |
| 속 분류 | M. romeri 이프레모프, 1938 M. efremovi 마호, Gee 외 라이즈, 2019 |
| 모식종 | Mesenosaurus romeri |
| 모식종 명명자 | 이프레모프, 1938 |
| 생물 분류 | |
| 과 | 바라놉스과 |
| 시대 | |
| 시기 | 페름기, |
2. 특징
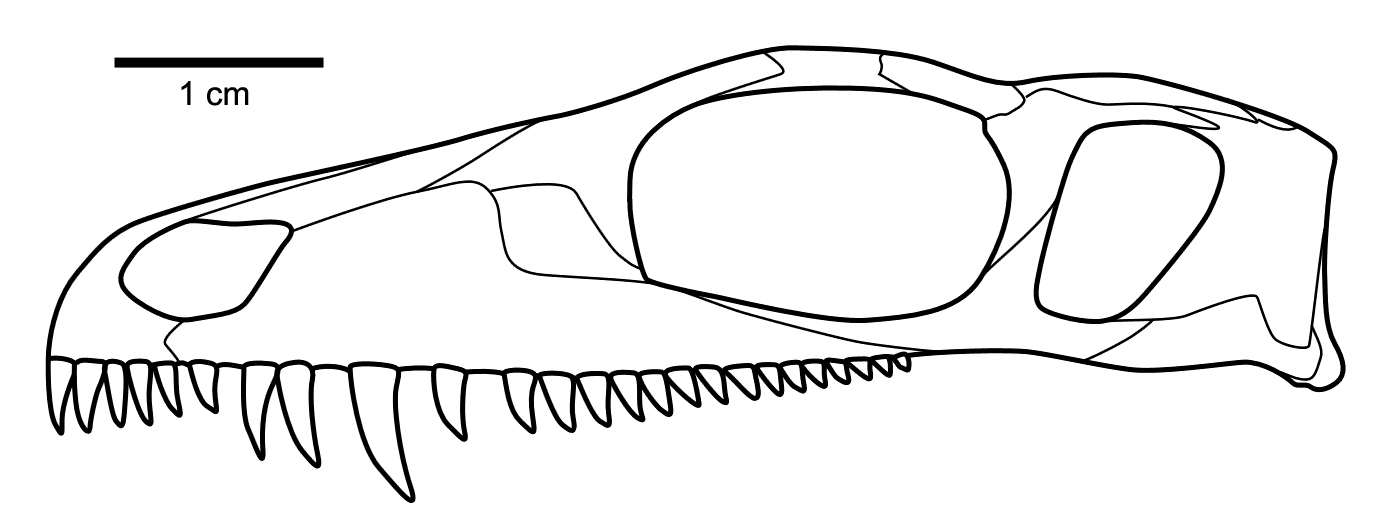
''메세노사우루스''는 소형 바라노피드(varanopid) 단궁류이다. 이들은 주로 두개골 특징으로 특징지어진다.[2]
''메세노사우루스''의 두개골 특징은 다음과 같다.[2]
# 가늘고 긴 전상악골이 좁고 직사각형의 주둥이를 형성한다(등쪽 및 배쪽 시야에서).
# 전상악골의 등쪽 돌기는 길고 외부 콧구멍의 등쪽 가장자리의 앞쪽 절반을 형성한다.
# 주둥이 끝까지 거의 뻗어 있는 확장된 콧구멍 선반.
# 전상악골의 구개 돌기는 길고 긴 정중 봉합을 갖는다.
# 콧구멍 경계에서 앞쪽 눈물뼈 앞쪽 끝까지 뻗어 있는 비골의 측면에 잘 발달된 함몰부.
# 송곳니 모양 이빨 수준에서 상악골의 약간의 측면 부풀어 오름.
# 상악골의 짧은 후방 돌기는 안와후부 막대 수준에 도달하지 못함.
# 첫 번째 전상악 이빨은 두 번째 및 세 번째 이빨보다 작다. 전상악골에는 최대 5개의 이빨이 있다. 첫 번째 이빨은 두 번째 및 세 번째 이빨보다 작지만 마지막 두 개보다 크다. 이 5개의 이빨은 간격이 좁고, 강하게 뒤로 휘어져 있으며, 날카롭게 뾰족하며, 뒤쪽 가장자리의 먼쪽 절반에 날카로운 절단면을 가지고 있다.
# 단일, 정중 서골 이빨 열.
# 두개골의 안와후부 광대뼈 부위는 비정상적으로 넓고 낮으며 거의 수직인 후방 가장자리를 갖는다.
# 익상골의 가로 플랜지의 후방 가장자리는 기저 관절에서 약간 앞쪽으로 각을 이룬다.
# 등골은 가늘고 짧으며 막대 모양이며 적당히 발달된 발판과 먼쪽으로 확장된 사골 돌기를 갖는다. ''메세노사우루스''의 등골은 페름기 초기 테랍시드와 더 유사하다.
# 전두골, 안와후부 및 협골의 안와 가장자리에 매우 두드러진 결절 장식이 존재한다. 이 특징은 잘 보존된 ''메세노사우루스''의 두개골에서 매우 두드러져 에프레모프는 원래 이를 ''메세노사우루스''의 고유 파생형으로 간주했다. 그러나 주의 깊은 검토와 분석을 통해 이 특징이 다른 바라노피드에도 존재한다는 것이 밝혀졌다.
''메세노사우루스''는 소형 바라노피드(varanopid) 단궁류이다. 주로 두개골이 특징적이며, 화석 증거 부족으로 인해 두개골 외 특징들은 완전히 분석되지 않았다.
''메세노사우루스''는 구부러지지 않고 납작하게 만든 비구를 가지고 있었는데, 이것은 그들이 굴을 파는 동물이었음을 암시한다. 많은 바라노피드들이 나무 위에서 생활했지만, ''메세노사우루스''의 잘 발달된 척골의 주두는 상완삼두근과 주근의 존재를 나타내는데, 둘 다 강력한 팔뚝 확장을 제공했을 것이다. 팔뚝 확장력과 다소 작은 체격/중간 체격이 결합된 이 강도는 굴을 파는 생활 방식을 뒷받침한다. 또한 ''메세노사우루스''에서는 기능적 이족보행이 일어났다고 제안된다. 이것은 기능적 이족보행에 필요한 신체 질량 중심(가는 줄기, 길쭉한 뒷다리, 짧은 앞다리)의 후방 이동의 존재에 기초한다. 먹이로는 당대에 서식했던 곤충, 절지동물 등을 주로 잡아먹고 살았을 육식성 포식자로 추정된다.
2. 1. 두개골
메세노사우루스는 작은 크기의 바라놉세이드 시냅스이다. 주로 두개골이 특징적이다.[2]''메세노사우루스''의 두개골 특징은 다음과 같다.[2]
# 가늘고 긴 전상악골이 좁고 직사각형의 주둥이를 형성한다(등쪽 및 배쪽 시야에서).
# 전상악골의 등쪽 돌기는 길고 외부 콧구멍의 등쪽 가장자리의 앞쪽 절반을 형성한다.
# 주둥이 끝까지 거의 뻗어 있는 확장된 콧구멍 선반.
# 전상악골의 구개 돌기는 길고 긴 정중 봉합을 갖는다.
# 콧구멍 경계에서 앞쪽 눈물뼈 앞쪽 끝까지 뻗어 있는 비골의 측면에 잘 발달된 함몰부.
# 송곳니 모양 이빨 수준에서 상악골의 약간의 측면 부풀어 오름.
# 상악골의 짧은 후방 돌기는 안와후부 막대 수준에 도달하지 못함.
# 첫 번째 전상악 이빨은 두 번째 및 세 번째 이빨보다 작다. 전상악골에는 최대 5개의 이빨이 있다. 첫 번째 이빨은 두 번째 및 세 번째 이빨보다 작지만 마지막 두 개보다 크다. 이 5개의 이빨은 간격이 좁고, 강하게 뒤로 휘어져 있으며, 날카롭게 뾰족하며, 뒤쪽 가장자리의 먼쪽 절반에 날카로운 절단면을 가지고 있다.
# 단일, 정중 서골 이빨 열.
# 두개골의 안와후부 광대뼈 부위는 비정상적으로 넓고 낮으며 거의 수직인 후방 가장자리를 갖는다.
# 익상골의 가로 플랜지의 후방 가장자리는 기저 관절에서 약간 앞쪽으로 각을 이룬다.
# 등골은 가늘고 짧으며 막대 모양이며 적당히 발달된 발판과 먼쪽으로 확장된 사골 돌기를 갖는다. ''메세노사우루스''의 등골은 페름기 초기 테라시드와 더 유사하다.
# 전두골, 안와후부 및 협골의 안와 가장자리에 매우 두드러진 결절 장식이 존재한다. 이 특징은 잘 보존된 ''메세노사우루스''의 두개골에서 매우 두드러져 에프레모프는 원래 이를 ''메세노사우루스''의 고유 파생형으로 간주했다. 그러나 주의 깊은 검토와 분석을 통해 이 특징이 다른 바라노피드에도 존재한다는 것이 밝혀졌다.
메세노사우루스를 특징짓는 두개골의 특징은 가느다란 프리맥시야가 좁고 직사각형 모양의 주둥이를 형성하며(등측 및 복측 뷰에서 나타난다.) 전축의 등가공정은 길고 등가공성의 앞쪽 반을 형성한다. 코끝까지 거의 확장되는 확장된 내레이셜 쉘프이며 전축의 구개척 공정이 길고 중앙 봉합이 길다. 내경경계로부터 전전두엽의 거의 앞쪽 끝으로 후방으로 확장되는 코의 측면 표면에 잘 발달된 우울증이며 개복치 수준에서 맥실라의 약간 측면 부기이고 맥실라의 짧은 후공정에서 후두막대 수준에 도달하지 못한다. 첫 번째 전치아는 두 번째와 세 번째 치아보다 작다. 전치아에는 최대 5개의 치아가 있다. 첫 번째 치아는 두 번째 치아와 세 번째 치아보다 작지만 마지막 두 개의 치아보다 크다. 이 다섯 개의 치아는 간격이 촘촘하고 강하게 재발하며 날카롭게 뾰족하며 후두 가장자리 원위 반을 따라 날카롭고 절단을 이루고 있다. 단일 중앙분리대 보메라린 톱니 행이며 두개골의 후두부 광대 부위는 거의 수직 후방 여백이 있는 비정상적으로 넓고 낮다. 프테리고이드 가로 플랜지의 후측 가장자리는 기저 관절에서 약간 무테라터적으로 각이 져 있다. 가늘고 짧으며 로드처럼 생긴 스테이플로 중간 정도 발달된 발판과 멀리 확장된 사분율 공정을 갖추고 있다.(메세노사우루스의 모습은 초기 페미안의 치료법에 있는 것과 더 흡사하다.) 매우 두드러지고 결절형 장식이 전전, 후두, 주갈의 궤도 여백에 존재한다.(이 특성은 메세노사우루스의 잘 보존된 두개골에서 두드러지게 나타나므로 에프레모프는 원래 메세노사우루스의 오토포모피라고 여겼다.그러나 세심한 검사 및 분석을 통해 이 기능이 다른 바라놉스과에 존재한다는 것을 밝혀냈다.)
2. 2. 두개골 외 특징
메세노사우루스는 작은 크기의 바라놉세이드 시냅스이다. 주로 두개골이 특징적이며 화석 증거의 부족으로 인해 두개골 외 특징들은 완전히 분석되지 않았다.메세노사우루스는 구부러지지 않고 납작하게 만든 비구슬을 가지고 있었는데 이것은 그들이 발굴자인 것을 암시한다. 많은 바라놉세이드들이 수목형이었지만 메세노사우루스의 잘 발달된 올레크라논은 삼두근과 안코네우스근의 존재를 나타내는데 둘 다 강력한 팔뚝 확장을 제공할 것이다. 팔뚝 확장력과 다소 작은 체격/중간 체격이 결합된 이 강도는 파고드는 라이프스타일 사상을 뒷받침한다. 또한 메세노사우루스에서는 기능적 두발주의가 일어났다고 제안한다. 이것은 기능적 두발주의에 필요한 신체 질량 중심(슬렌더 줄기, 길쭉한 뒷줄기, 짧은 앞줄기)의 후방 이동의 존재에 기초한다. 먹이로는 당대에 서식했던 곤충, 절지동물 등을 주로 잡아먹고 살았을 육식성의 포식자로 추정되는 종이다.
3. 종
3. 1. 메세노사우루스 로메리 (''Mesenosaurus romeri'')

메젠강 유역은 러시아 북부에 위치해 있으며 메젠강의 지류를 따라 중기 페름기 퇴적층이 광범위하게 노출되어 있다. 이 퇴적층에서 다양한 양막류의 많은 골격 유해가 발견되었는데, 그중 가장 중요한 것은 "바라노프세이드(varanopseid) 친화성을 가진 작은 단궁류"인 ''Mesenosaurus romeri''의 두개골 일부이다.[2] ''M. romeri''는 1938년 이반 예프레모프에 의해 발견되었으며, 다른 종과의 두개골 유사성이 없어 최초의 ''Mesenosaurus'' 종이 되었다. 또한, 다른 알려진 펠리코사우루스류(가늘고 굽어져 있으며 확대된 앞니, 단일 송곳니 형태의 이빨)와 일치하는 윗턱 이빨을 가지고 있어 러시아 지역에서 "좋은 펠리코사우루스"로 간주된 최초의 단궁류이기도 하다. 로머와 프라이스는 ''M. romeri''가 바라노프세과(Varanopseidae)에 속한다고 가설을 세웠다.[3] 이 가설은 2001년에 다음과 같은 공유 파생 형질을 기반으로 확인되었다.
- 등배 방향으로 확장된 측두창이 측두 부위 높이의 대부분을 차지하여 좁은 하측두대를 형성한다.
- 변연 치열은 강하게 굽어져 있고, 측면으로 압축되어 있으며, 날카롭게 뾰족한 앞뒤 절단면이 이빨 원위부 절반으로 제한된 이빨로 구성된다.
- 잘 발달된 전상악골 비공하 선반은 외부 표면이 횡단면에서 넓게 둥글다.
- 두정골의 전방 중앙 돌기는 두개골 테이블의 상안와 부위까지 확장된다.
- 작은 후전두골은 전두골의 좁고 후방 돌기의 내측에 접한다.
- 판상골은 작아지고 좁은 요소가 되어 두정골의 후외측 날개의 내측 가장자리에 접한다.
- 사골의 후방 가장자리의 내측, 후두골 플랜지의 부재로 사골의 후방 가장자리를 덮는다.
- 부시상골판은 넓고, 기저접형골 결절은 날개 모양이며, 쟁기 모양 돌기의 기저에서 멀리 측면 및 후방으로 뻗어 있다.
- 현저한 결절 장식이 전두골, 후안와골 및 협골의 안와 가장자리에 존재한다.[2]
3. 2. 메세노사우루스 에프레모비 (''Mesenosaurus efremovi'')
메세노사우루스속의 두 번째 종은 ''메세노사우루스 에프레모비(''Mesenosaurus efremovi'')''이다. 거의 완전한 두개골과 아래턱뼈는 오르도비스기 아버클 석회암의 채워진 카르스트 균열에서 발견되었으며, 이곳은 초기 페름기 사지형류 화석이 가장 풍부하게 발견되는 곳 중 하나이다.[4]''M. efremovi''를 분류하는 측면에서, 이 종은 미크테로사우루스류(''Mycterosaurus longiceps''를 포함하고 ''Varanodon agilis''보다 더 밀접하게 관련된 모든 바라노프세이드의 줄기 기반 그룹)와 뚜렷한 두개골 특징을 공유한다. 예를 들어, "외측 비공으로부터의 눈물뼈의 배제, 그리고 코뼈 아래에 있고 이마뼈와 접촉하는 위아래로 넓은 위턱뼈의 등쪽 판"이 있다. 그러나 ''M. efremovi''는 러시아에서 발견된 ''M. romeri''와 더 많은 특징을 공유한다. 이러한 공유된 특징 중 일부는 측두창의 상대적인 크기와 모양, 송곳니 모양 영역의 위턱뼈의 측면 부풀림, 그리고 5개의 앞위턱뼈 치아 위치(다른 미크테로사우루스류에서는 보고되지 않음) 등이 있다.[1]
비록 ''M. efremovi''와 ''M. romeri''가 많은 뚜렷한 특징을 공유하지만, 종 수준에서 분류학적 구분을 정당화하는 이 표본 간의 4가지 주요 형태학적 차이점이 있다(종 수준 이상에서 구별하기에는 충분하지 않은 차이점).
# 짧은 등쪽 앞위턱뼈 돌기(뒤쪽 비공 가장자리 또는 코뼈에 의한 앞위턱뼈의 뒤쪽 분리까지 확장되지 않음)의 존재
# 더 뒤쪽으로 확장된 위턱뼈
# 위턱뼈의 더 적은 치아 위치
# 눈 뒤뼈와 위측두골의 접촉
''M. efremovi''는 또한 ''M. romeri''의 가장 큰 알려진 표본보다 크다.[1]
4. 계통 분류
분기도는 벤슨의 분석을 수정하여 만들어졌으며, ''Basicranodon''은 이 분기도에서 제외되었다.[5]
{| class="wikitable"
|-
! 양막류
|-
|
{| class="wikitable"
|-
! 단궁류
|-
|
{| class="wikitable"
|-
| 오피아코돈과
|-
|
{| class="wikitable"
|-
! 바라노피스과
|-
|
{| class="wikitable"
|-
| ''Archaeovenator hamiltonensis''
|-
|
{| class="wikitable"
|-
| ''Pyozia mesenensis''
|-
|
{| class="wikitable"
|-
! ''Mycterosaurinae''
|-
|
{| class="wikitable"
|-
| ''Mycterosaurus longiceps''
|-
|
| ?Elliotsmithia longiceps (BP/1/5678) |
| Heleosaurus scholtzi |
| '메세노사우루스(Mesenosaurus romeri |
|}
|-
! Varanopinae
|-
|
{| class="wikitable"
|-
|
{| class="wikitable"
|-
| ''Varanops brevirostris''
|-
|
| Watongia meieri |
| Varanodon agilis |
|}
|-
|
{| class="wikitable"
|-
| ''Ruthiromia elcobriensis''
|-
|
| Aerosaurus wellesi |
| Aerosaurus greenleorum |
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
5. 고생태학
메세노사우루스는 고생대 페름기인 지금으로부터 2억 9000만년전~2억 4500만년전에 생존했었던 종이다. 유럽, 러시아, 미국으로 이어지는 당시에 존재했었던 초원과 산맥에 형성된 밀림 및 열대우림에서 광범위하게 생존했었다. 화석은 1938년에 러시아의 페름기에 형성된 지층에서 러시아의 고생물학자인 이반 에프레모프에 의하여 처음으로 발굴되었으며 이후엔 유럽과 미국에서도 발견되었다.[6]
메세노사우루스는 바위나 나무를 오를 수 있는 작고 민첩한 생물이었다.[7] 이들은 작은 크기 덕분에 테랍시드가 대부분의 육상 환경을 지배하는 동안 작은 동물 식성 동물로서 틈새 시장을 점유하고 번성할 수 있었다. 늦은 페름기에 작은 이궁류가 나타나 경쟁을 벌이면서[8], ''메세노사우루스''는 쇠퇴하게 되었다. 크고 가늘고 구부러진 이빨을 가지고 있어 먹이를 꿰뚫을 때 심한 상처를 입힐 수 있었기 때문에 그들의 공동체에서 매우 민첩한 하위 포식자 길드를 대표한다.[1][9] 또한 다른 거의 모든 육상 양막류에 비해 매우 빠른 치아 발달 속도와 크게 감소된 치아 수명을 특징으로 했다.[11]
굽혀지지 않고 평평한 발톱을 가지고 있어 땅을 파는 동물이었음을 시사한다. 많은 바라노피드는 수목 생활을 했지만, 메세노사우루스의 잘 발달된 주두(팔꿈치의 뼈 돌출부)는 삼두근과 주근육의 존재를 나타내며,[12][13][14] 이 둘 다 강력한 전완부 신전을 제공한다. 이러한 전완부 신전 강도는 다소 작거나 중간 크기의 신체와 결합되어 굴을 파는 생활 방식이라는 생각을 뒷받침한다.[10] ''메세노사우루스''에서 임의적 이족 보행이 발생했다는 가설이 제기되었다. 이는 임의적 이족 보행에 필요한 신체 질량 중심의 후방 이동(가늘고 긴 몸통, 긴 뒷다리, 짧은 앞다리)을 기반으로 한다.[10]
6. 진화적 의의
''메세노사우루스''는 바위나 나무를 오를 수 있는 작고 민첩한 생물이었다.[7] 작은 크기 덕분에 테랍시드가 대부분의 육상 환경을 지배하는 동안 작은 동물 식성 동물로서 틈새 시장을 점유하고 번성할 수 있었다. 늦은 페름기에 작은 이궁류가 나타나 경쟁을 벌이면서[8], ''메세노사우루스''는 쇠퇴하게 되었다. 크고 가늘고 구부러진 이빨을 가지고 있어 먹이를 꿰뚫을 때 심한 상처를 입힐 수 있었기 때문에 그들의 공동체에서 매우 민첩한 하위 포식자 길드를 대표한다.[1][9] 또한 다른 거의 모든 육상 양막류에 비해 매우 빠른 치아 발달 속도와 크게 감소된 치아 수명을 특징으로 했다.[11]
굽혀지지 않고 평평한 발톱을 가지고 있어 땅을 파는 동물이었음을 시사한다. 땅을 파는 발톱이 굴을 파는 데 적합했는지, 아니면 단순히 먹이를 파는 데만 사용되었는지는 불확실하다.[10] 많은 바라노피드는 수목 생활을 했지만, 메세노사우루스의 잘 발달된 주두(팔꿈치의 뼈 돌출부)는 삼두근과 주근육의 존재를 나타내며,[12][13][14] 이 둘 다 강력한 전완부 신전을 제공한다. 이러한 전완부 신전 강도는 다소 작거나 중간 크기의 신체와 결합되어 굴을 파는 생활 방식이라는 생각을 뒷받침한다.[10] ''메세노사우루스''에서 임의적 이족 보행이 발생했다는 가설이 제기되었다. 이는 임의적 이족 보행에 필요한 신체 질량 중심의 후방 이동(가늘고 긴 몸통, 긴 뒷다리, 짧은 앞다리)을 기반으로 한다.[10]
참조
[1]
논문
A new varanopid synapsid from the early Permian of Oklahoma and the evolutionary stasis in this clade
[2]
서적
The skull of ''Mesenosaurus'' romeri, a small varanopseid (Synapsida: Eupelycosauria) from the Upper Permian of the Mezen River Basin, northern Russia
2001
[3]
간행물
28 : Review of the Pelycosauria
Geological Society of America
1940
[4]
논문
The unique preservational environment of the Early Permian (Cisuralian) fossiliferous cave deposits of the Richards Spur locality, Oklahoma
2017-06
[5]
논문
Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies
[6]
논문
Speleothem climate records from deep time? Exploring the potential with an example from the Permian
2010-05
[7]
논문
First arboreal 'pelycosaurs' (Synapsida: Varanopidae) from the early Permian Chemnitz Fossil Lagerstätte, SE Germany, with a review of varanopid phylogeny
2018-03-15
[8]
논문
Pyozia mesenensis, a new, small varanopid (Synapsida, Eupelycosauria) from Russia: "pelycosaur" diversity in the Middle Permian
2004-03-25
[9]
논문
A second varanopseid skull from the Upper Permian of South Africa: implications for Late Permian 'pelycosaur' evolution
2001-12-01
[10]
논문
A new mesenosaurine from the lower Permian of Germany and the postcrania of Mesenosaurus: implications for early amniote comparative osteology
2019-01-19
[11]
논문
Permian hypercarnivore suggests dental complexity among early amniotes
2022-08-19
[12]
논문
The flexor muscles of the forearm and hand in lizards and mammals
1950
[13]
논문
A revision of the extensor muscles in the forearm in tetrapods.
1939
[14]
논문
Osteology of the Reptiles
1957-07-15
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com