오파비니아
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오파비니아는 캄브리아기에 살았던 멸종된 해양 절지동물로, 1911년 버제스 셰일에서 발견되었다. 몸길이는 4~7cm 정도이며, 5개의 눈, 코(앞 부속지), 지느러미와 아가미를 가진 몸통, 꼬리를 가지고 있다. 오파비니아는 처음에는 갑각류로 분류되었으나, 이후 절지동물도, 환형동물도 아닌 원시적인 계통으로 여겨졌다. 현재는 아노말로카리스 등과 함께 절지동물 줄기군으로 분류된다. 오파비니아의 독특한 생김새는 동물 진화 연구에 중요한 단서를 제공하며, 캄브리아기 대폭발 연구에 영향을 미쳤다.
더 읽어볼만한 페이지
- 고대 무척추동물 - 에스테리아 (갑각류)
에스테리아는 절지동물 무갑목 에스테리아과에 속하는 갑각류의 일종이다. - 고대 무척추동물 - 공하강
공하강은 캄브리아기 초기에 번성한 다족 연충 형태의 멸종 동물군으로, 특이한 소화샘 구조와 3엽 뇌를 가지며 탈피동물의 조상과 유사하고 절지동물 기원 연구에 중요한 위치를 차지한다. - 공하강 - 우타우로라
우타우로라는 유타주 휠러 지층에서 발견된 화석을 토대로 명명되었으며, 5개의 눈과 특이한 전방 부속지, 강모깃과 꼬리깃으로 이루어진 꼬리지느러미를 가진 오파비니아과의 새로운 속이자 종으로, 오파비니아와 구별되는 특징을 가지며 계통 분류 연구에 중요한 자료이다. - 공하강 - 팜브델루리온
팜브델루리온은 캄브리아기 중기 버제스 셰일에서 발견된 멸종된 엽족동물로, 커다란 머리 부속지, 가시돌기, 11쌍의 옆지느러미와 엽족, 그리고 얇은 큐티클을 가진 육식성 기엽족류이다. - 1912년 기재된 화석 분류군 - 사우롤로푸스
사우롤로푸스는 하드로사우루스과에 속하는 초식 공룡의 한 속으로, 머리 뒤쪽으로 솟아오른 볏이 특징이며 북아메리카와 아시아에서 발견되었고, 다양한 식물을 섭취하며 집단 생활을 했을 가능성이 있다. - 1912년 기재된 화석 분류군 - 나라오이아
나라오이아는 캄브리아기 후기부터 실루리아기 최후반까지 생존한 절멸된 절지동물로, 납작하고 넓은 몸체, 석회화되지 않은 머리방패와 꼬리방패를 가지며, 다양한 종으로 분류되고 바닥에서 굴을 파고 살았던 흙을 먹는 여과 섭식자이거나 포식자였을 가능성이 있다.
| 오파비니아 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
![국립 자연사 박물관에 전시된 화석 표본 ([[홀로타입]])](https://cdn.onul.works/wiki/noimage.png) | |
| 학명 | Opabinia regalis |
| 명명자 | 월콧, 1912 |
| 화석 범위 | 중기 캄브리아기 (울류안기), |
| 생물학적 분류 | |
| 계 | 동물계 |
| 상문 | 탈피동물상문 |
| 상문 계급 없음 | 범절지동물 |
| 문 | 스템그룹 절지동물문 |
| 강 | †공포동물강 |
| 과 | †오파비니아과 |
| 속 | †오파비니아속 |
| 종 | 오파비니아 레갈리스 |
| 기타 정보 | |
| 발견 장소 | 버지스 혈암 |
| 시기 | 캄브리아기 중기 (울류안기) |
2. 발견 및 연구사
1911년, 찰스 돌리틀 월콧은 버제스 셰일에서 거의 완전한 형태의 ''오파비니아 레갈리스'' 화석 9개와 ''오파비니아 ? 미디어''로 분류한 화석 몇 개를 발견하여 1912년에 발표했다.[7] 속명은 캐나다 브리티시컬럼비아주, 레이크 오하라 남동쪽에 위치한 오파빈 패스에서 유래했으며, 이 패스는 마운트 헝가비와 마운트 비들 사이에 있다.[7] 1966년~1967년, 해리 B. 휘팅턴은 또 다른 표본을 발견했으며,[8] 1975년에는 일부 표본의 해부와 다양한 각도에서 조명한 사진을 바탕으로 상세한 설명을 발표했다. 휘팅턴의 분석은 ''오파비니아 ? 미디어''는 다루지 않았는데, 월콧의 이 종 표본은 그의 컬렉션에서 확인할 수 없었다.[1] 1960년 러시아 고생물학자들은 시베리아의 노릴스크 지역에서 발견한 표본을 ''오파비니아 노릴리카''라고 명명했지만,[9] 보존 상태가 좋지 않아 휘팅턴은 속 ''오파비니아''로 분류할 만큼 충분한 정보를 제공한다고 생각하지 않았다.[1]
오파비니아는 캄브리아기에 살았던 기묘한 생김새의 동물이다. 머리에는 5개의 눈과 긴 코가 있고, 몸통은 15개의 마디로 나뉘어 있으며, 각 마디에는 부채꼴 모양의 지느러미와 아가미가 달려 있었다.
학명 "''Opabinia''"는 발견지(캐나다 브리티시컬럼비아 주, 버제스 셰일, 월콧과 레이몬드 채석장) 근처 오하라 호(Lake O'Hara) 남동쪽에 위치한 요호 국립공원(Yoho National Park)의 헝가비 산(Mount Hungabee)과 비들 산(Mount Biddle) 사이 오파빈 고개(Opabin Pass)에서 유래했다("Opabin"은 현지 언어로 "바위"를 의미한다[63])[53]). 모식종(타입 종) 종소명 "''regalis''"는 라틴어로 "왕의"를 의미한다.[65]
오파비니아 화석표본은 희귀하며,[65] 캐나다 브리티시컬럼비아주 버제스 산 퇴적층 버제스 셰일(Burgess Shale, 캄브리아기 우류안기, 약 510Ma - 505Ma[64]) 월코트 앤 레이먼드 채석장(Walcott and Raymond Quarries)에서 유일하게 ''오파비니아 레갈리스(''Opabinia regalis'')''가 발견된다.[65] 이 속은 미국 고생물학자 찰스 왈콧(Charles Doolittle Walcott)에 의해 처음 발견되어 1912년에 기재되었다.[53] 그러나 독특한 모습이 해명되어 주목받게 된 것은 영국 고생물학자 해리 휘팅턴(Harry B. Whittington)에 의해 재기재가 이루어진 1970년대 이후이다(후술).[54][119] 선정 기준 표본(렉토타입) USNM 57683은 미국 국립 자연사 박물관(Smithsonian National Museum of Natural History)에 소장되어 있다.[65]
시베리아에서 발견되어 Miroshnikov & Krawzov 1960에 종()으로 기재된 화석 표본도 있었지만,[123] 이후 연구에서 이 속에서 유래된 것으로 인정되지 않았다.[54]
오파비니아는 캄브리아기 고생물 중 형태학상 복원과 분류학상 위치에 대해 많은 논쟁이 벌어진 생물이다.[119] 20세기 초에는 갑각류,[53] 20세기 후기에는 논쟁적인 미확인 화석(프로블레마티카),[56] 1990년대 이후에는 오파비니아류 일원으로서 아노말로카리스 등 라디오돈트류와 함께 기반적인 절지동물로 널리 인정받게 되었다.[124][67][125][126][119][58][59][60][61][62]
버제스 셰일 (버제스 동물군)에서 처음 발견되었을 때 그 특이성이 이해되지 않아 오파비니아는 갑각류 절지동물로 여겨졌다.[53] 미국 고생물학자 찰스 왈콧(Charles Doolittle Walcott) 1912년 기재(Walcott 1912)에서는 현존하는 과 무갑류 (새갑류로 분류되는 논병아리새우와 갯강구를 포함한 갑각류 분류군)와 비교되었으며, 원시적인 무갑류로 여겨졌다. 또한 이 종류 무갑류 수컷은 앞에 갈라진 관이 있기 때문에 관(전부 부속지)이 보이는 오파비니아 화석표본은 모두 수컷으로 해석되었다.[53] 오파비니아 화석 표본에서 무갑류에 있을 머리 부분 구조(2쌍 촉각・큰턱・작은턱)가 보이지 않고, 지느러미(당시 '가슴 다리')에 있는 빗 모양 구조 해석에 난점이 있다는 것도 명기되었다.[53]
이 견해는 동물학자 조지 이블린 허친슨(George Evelyn Hutchinson)에게도 계승되어, 허친슨 1930에서는 논병아리새우처럼 배영하는 갑각류로 해석된, 사상 최초[119] 오파비니아 복원도가 실렸다.[88] 시모네타 1970에서는 오파비니아 화석 표본에서는 보이지 않지만, 당시 오파비니아 근연으로 오해된 레안코일리야와 요호이아(후술) 특징을 바탕으로 등판・촉각・관절로 나뉜 다리 등 절지동물적 특징을 오파비니아 복원도에 많이 추가했다.[89]
1970년대, 영국 고생물학자 해리 위팅턴(Harry B. Whittington) 재기재(Whittington 1975)를 통해 오파비니아는 갑각류는 물론, 절지동물로 분류하기조차 어려울 정도로 독특한 특징을 지녔다는 것이 밝혀졌다.[54] 1972년 버제스 동물군 학회 발표 때 위팅턴 복원도가 비춰지자마자 회장 안이 폭소로 뒤덮였다는 일화가 남아있다.[54][119]
Whittington 1975에서는 오파비니아는 어떤 동물 문에도 속하지 않고, 절지동물과 환형동물 체절제를 지닌 원시적인 계통군 종류가 아닐까 해석되었다.[54] (절지동물과 환형동물은 당시에는 근연으로 취급되었으며, 서로 다른 계통이라는 것이 밝혀진 것은 21세기 이후이다. 절지동물#다른 동물문과의 관계성 참조[90]). 그 난해함은 스티븐 제이 굴드(Stephen Jay Gould) 저술 『풀 하우스』(1989)에서 강조되었으며, 캄브리아기 아노말로카리스 등과 함께 현생 동물문에 분류 불가능한 "프로블레마티카"(미상 화석)로 버제스 동물군 "기묘기천열 동물"(Weird Wonders) 대표적인 예로 언급되었다.[56]
3. 형태 및 생태
오파비니아의 가장 독특한 특징 중 하나는 진공 청소기 호스처럼 줄무늬가 있고 유연한 코인데, 몸길이의 약 1/3 정도이며 머리 아래에서 아래로 뻗어 있었다. 코 끝에는 안쪽과 앞으로 5개의 가시가 튀어나온 발톱 모양의 구조가 있었다.[1] 머리에는 5개의 눈이 달려 있었는데, 앞쪽에 가깝고 머리 중앙에 가까이 있는 2개는 위와 앞을 향하고, 머리 뒤쪽과 바깥쪽에 있는 더 큰 2개는 위쪽과 옆쪽을 향하며, 이들 사이에 단일 눈이 위를 향하고 있었다.[1] 입은 머리 아래 코 뒤에 위치하며 뒤쪽을 향하고 있어, 소화관은 동물의 뒤쪽으로 U자형 굴곡을 이룬다.[1]
몸통은 폭이 약 5mm이고, 15개의 마디와 각 마디에 아래쪽과 바깥쪽을 향하는 한 쌍의 플랩(엽)이 있었다. 각 플랩은 앞부분이 그 앞에 있는 플랩의 뒷 가장자리에 덮이는 형태로 겹쳐져 있었다. 몸체는 위와 바깥쪽으로 겹치는 세 쌍의 꼬리 부채 날을 가진 단일 원뿔형 마디처럼 보이며, V자형 이중 부채 모양의 꼬리를 형성했다.[1]
오파비니아는 해저 표면에 살면서 코를 이용해 퇴적물 속 먹이 입자를 찾아 입으로 전달했을 것으로 추정된다.[1] 턱이 없으므로 먹이는 작고 부드러웠을 것이며,[1] 쌍을 이룬 장 게실은 음식 소화와 영양 섭취 효율을 높였을 것이다.[40]
오파비니아의 각 부위별 특징은 하위 섹션에서 자세히 다루고 있으며, 특히 지느러미, 아가미, 다리(엽족)에 대해서는 학자들 간에 이견이 존재한다.
3. 1. 눈
오파비니아는 머리 앞쪽에 크고 작은 5개의 눈을 가지고 있는데, 이는 앞쪽 1쌍, 뒤쪽 양 옆 1쌍, 중앙 1개로 배열되어 있다. 앞뒤 2쌍의 눈은 모두 튼튼한 눈자루에 붙어 있는 겹눈이며, 뒤쪽 양 옆의 1쌍이 앞쪽 1쌍보다 크다.[54][124][69]
비슷한 눈을 가진 키린시아처럼, 이 눈들은 절지동물에서 흔히 볼 수 있는 두 종류의 눈, 즉 중앙의 작은 중안(median eye)과 좌우의 큰 측안(lateral eye)과 상동인 것으로 여겨진다.[69] 그러나 앞쪽 1쌍 눈의 상동성과 중앙 눈의 형태는 문헌에 따라 다르게 해석된다. 일반적인 학설에서는 모든 눈이 눈자루를 가지며, 뒤쪽 양 옆 1쌍만 측안이고 나머지 3개는 모두 약간 작은 중안이라고 본다.[69] 반면, Moysiuk & Caron 2022에서는 중앙의 1개 눈만 중안이며, 눈자루가 없고 기존 복원보다 크다고 해석한다.[70] 이러한 독특한 눈 구조는 오파비니아가 넓은 시야를 확보하고, 포식자나 먹이를 탐지하는 데 유리했을 것으로 보인다.
3. 2. 코 (전방 부속지)
오파비니아의 코는 몸길이의 1/3 정도이며, 진공 청소기의 호스처럼 줄무늬가 있고 유연했다. 코 끝에는 안쪽과 앞으로 5개의 가시가 튀어나온 발톱 모양의 구조, 즉 집게 모양의 부속지가 달려 있었다.[1] 이 부속지는 먹이를 잡는 데 사용했을 것으로 추정된다.[1] 코는 가동 범위가 넓어 정면으로 뻗거나 몸 아래로 접을 수 있었다.[54]
이 코는 일부 절지동물의 관절지 (예: 곤충의 아랫입술)처럼, 원래 한 쌍이었던 부속지가 기부를 좌우 융합하여 특화된 부분이며,[55][71][72][73] 전방 부속지의 기동성을 높이도록 진화한 것으로 생각된다.[66] 이는 라디오돈타류, 켈리그마켈라와 판부델리온의 좌우 독립적인 전방 부속지와는 크게 다르지만, 가시를 가지며, 눈과 입보다 앞에 특화된 부속지라는 공통점을 가지고 있다.[55][71][57][66][58]
3. 3. 지느러미와 아가미
오파비니아의 몸통에는 15개의 체절이 있었고, 각 체절에는 한 쌍의 지느러미가 달려 있었다.[54][55][124] 이 지느러미들은 아래쪽과 바깥쪽을 향하고 있었으며, 각 지느러미의 앞부분이 그 앞에 있는 지느러미의 뒷 가장자리에 덮이는 형태로 겹쳐져 있었다.[1]
각 지느러미에는 빗 모양의 구조물인 아가미가 붙어 있었는데,[126] 이 아가미와 지느러미의 구조에 대한 해석은 학자마다 조금씩 다르다.[119]
이러한 지느러미와 아가미는 오파비니아가 헤엄을 치거나 물속에서 산소를 얻는 데 중요한 역할을 했을 것으로 보인다.
3. 4. 다리 (엽족)
오파비니아의 다리(엽족) 존재 여부는 학계에서 논쟁 중인 বিষয়이다.휘팅턴(Whittington, 1975)은 몸통을 따라 거의 삼각 모양의 특징을 발견하고, 위장의 옆쪽 연장부(게실)일 가능성이 있다고 결론 내렸다.[1] 천(Chen) 등(1994)은 이를 엽 내에 포함된 것으로 해석했다.[18] 그러나 버드(1996)는 "삼각형"이 ''오파비니아''의 가늘고 긴 몸에 비해 너무 넓고, 단면에서 엽보다 낮고 분리되어 몸 아래로 확장된다고 생각했다.[17] 그는 다리의 외부 큐티클을 보존한 표본을 발견하고, "삼각형"을 짧고 살이 많고 원뿔형 다리(엽족)로 해석했다. 또한 끝 부분에서 작은 광물화된 패치를 발견하고 발톱으로 해석했다. 이 해석에 따르면, 아가미를 지닌 플랩과 엽족은 Marrella, 삼엽충, 갑각류에서 볼 수 있는 절지동물의 두 갈래 다리의 외부 아가미 가지와 내부 다리 가지에 해당한다.[17]
장과 브리그스(2007)는 "삼각형"의 화학적 조성을 분석하여 위장과 동일한 조성을 가지고 있음을 확인하고, 휘팅턴(Whittington)의 해석에 동의했다. 그들은 ''오파비니아''의 엽+아가미 배열을 절지동물 다리가 두 갈래 구조로 분리되기 전 초기 형태로 간주했다.[19] 그러나 버드와 데일리(2011)는 이러한 화학적 조성이 소화관뿐만 아니라 몸 내부의 액체로 채워진 공동 내에서 형성되는 광물화를 나타내며, 이는 속이 빈 엽족과 일치한다고 제안한다. 그들은 ''오파비니아''의 위장 게실이 "삼각형"에서 개별화된 일련의 원형 위장샘임을 명확히 한다. 터미널 발톱의 부재에는 동의하지만, ''오파비니아''에서 엽족의 존재는 여전히 타당한 해석으로 남아 있다.[16]
화석 표본의 각 체절과 지느러미 사이에는 다소 긴 삼각형 흔적이 한 쌍씩 나란히 존재하며, 이 부분에 대해서는 주로 '''소화관 가지'''와 '''부드러운 다리(엽족)'''라는 두 가지 가설이 있었다.[119] 이러한 해석 차이는 오파비니아의 분류학적 위치에 큰 영향을 미치지 않았다.[119][68]
- 휘팅턴(Whittington, 1975): 삼각형 흔적은 내부 구조이며, 소화관 가지 혹은 아가미에 연결되는 순환계의 일부로 해석했다.[54]
- 버드(Budd, 1996): 삼각형 흔적은 엽족의 체강에서 유래된 흔적이며, 각 체절 복측에 한 쌍의 짧은 엽족이 있고, 각 엽족 끝에 발톱이 있다고 추측했다.[124]
- 장 & 브리그스(Zhang & Briggs, 2007): 원소 매핑(elemental mapping)으로 삼각형 흔적의 원소 조성을 분석하여 소화관과 같은 성분임을 확인했다.[125] 버드(1996)의 엽족과 발톱 존재를 부정하고, 이 부분을 지느러미까지 삽입된 소화관 가지로 해석했다.[125]
- 버드 & 데일리(Budd & Daley, 2011): 엽족의 존재 가능성을 지지하며 버드(1996)의 해석을 거의 인정했다. 다만, 발톱의 존재는 부정했다.[126] 소화관 가지는 둥근 소화선이며, 삼각형 흔적의 원소 조성은 소화관에 국한되지 않는다고 밝혔다.[126] 엽족동물 아이셰아이아의 엽족 체강 흔적과 오파비니아의 삼각형 흔적 유사성을 제시하고, 일부 표본에서 엽족 외피 조직과 유사한 환형 근육 구조를 발견했다.[126]
3. 5. 꼬리
오파비니아의 몸체는 위와 바깥쪽을 향해 겹치는 세 쌍의 꼬리 부채 날을 가진 단일 원뿔형 분절처럼 보였고, V자형 이중 부채 모양의 꼬리를 형성했다.[1] 이는 오파비니아가 헤엄칠 때 방향을 조절하는 역할을 했을 것으로 보인다.3. 6. 내부 구조
오파비니아의 소화관은 머리에서 뒤쪽으로 U자 형태로 꺾여 있는데, 이는 입이 머리 아래쪽, 코 뒤에 위치하여 뒤쪽을 향하고 있기 때문이다.[1] 소화관의 앞쪽(인두)은 U턴하여 꺾여 있으며, 제2 체절부터 제13 체절에 걸쳐 중장의 좌우에 11쌍의 둥근 소화샘(digestive gland, 중장선 midgut gland)이 나란히 있다.[126][76] 이러한 소화샘은 몸통에 여러 쌍으로 존재하며, 먹이를 효율적으로 소화하는 데 도움을 주었을 것으로 보인다.[126][76] 이러한 소화계는 절지동물적이며, 여러 쌍의 소화선은 초기 절지동물에서 일반적으로 관찰되는 특징이다.[76][58][59][61]4. 분류 및 진화
오파비니아의 화석표본은 희귀하며[65], 캐나다브리티시컬럼비아주 버제스 산의 퇴적층인 버제스 셰일(캄브리아기우류안기, 약 5억 1,000만 - 5억 500만 년 전[64]) 월코트 앤 레이먼드 채석장에서 발견된 오파비니아 레갈리스(''Opabinia regalis'')가 유일한 종이다[65]。 찰스 왈콧(Charles Doolittle Walcott)이 1912년 처음 기재하였으나[53], 해리 휘팅턴(Harry B. Whittington)이 1970년대 재기재하면서 독특한 모습이 해명되어 주목받았다[54][119]。 선정 기준 표본(렉토타입) USNM 57683은 국립 자연사 박물관에 소장되어 있다[65]。
시베리아에서 발견된 표본이 오파비니아 속으로 기재되기도 했지만[123], 이후 연구에서 인정받지 못했다[54]。
소화관은 명확히 발견되었다[54][76]。 입이 머리 부분 복측에서 뒤쪽을 향해 열려 인두가 U턴하는 형태이다[54][58][59]。 제2~13 체절 중장 좌우에 11쌍의 둥근 소화선(digestive gland, 중장선 midgut gland)이 있다[126][76]。 이는 절지동물적이며, 여러 쌍의 소화선은 초기 절지동물(시베리온류[77][78], 켈리그마켈라[79], 판부델리온[80], 라디오돈타류[81][82][83], 키린시아[69], 이소크시스[84], 푸센후이아류[85], 메가케이라류[86], 삼엽충[87] 등)에서 흔히 보이며[76][58][59][61], 꺾인 인두는 진정 절지동물 공통 특징이다[58][59]。
소화관 외에 체강으로 추정되는, 각 체절에서 삼각형으로 갈라진 흔적이 있다[124][126]。
오파비니아는 캄브리아기 고생물 중 형태학적 복원과 분류학적 위치에 대해 많은 논쟁이 벌어진 생물이다[119]。
{{cladogram
|title=
|align=right
|caption=계통 분석에 의해 지지되는 오파비니아의 계통 위치[120]
†: 멸종군, 파란색 테두리: 기저 절지동물, *: 엽족동물
|cladogram=
{{clade| style=width:35em;font-size:85%;line-height:70%;
|label1=탈피동물|1={{clade
|label1=환형동물|1=새예동물 --, 선형동물 
|label2=범절지동물|2={{clade
|1=유조동물(선구동물) --
|2=완보동물(물곰) --
|3=*†다양한 엽족동물(측계통군) --|state3=double
|4={{clade
|1=*†시베리온류 --|barbegin1=blue
|2={{clade
|1=*†팜브델리온 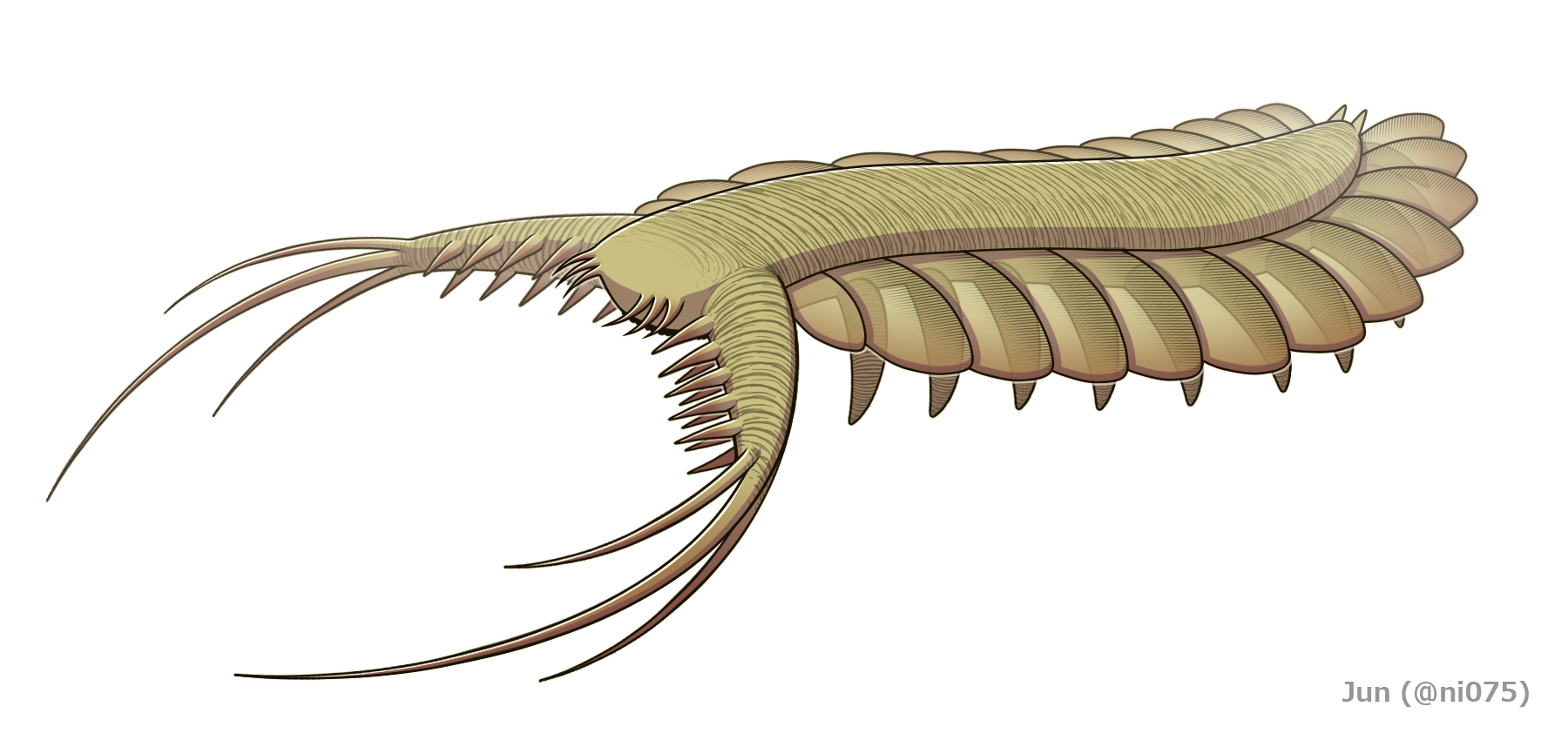
|2=*†케리그마케라 --|bar2=blue
|3={{clade
|label1=오파비니아류|1={{clade
|1=†'''오파비니아''' --|bar1=blue
|2=†유타우롤라 --|bar2=blue
}}
|2=†방사두류 
|3=진절지동물 --
}} }} }} }} }} }}
오파비니아는 '''오파비니아과'''( '''''', '''오파비니아류 opabiniid''' )의 모식속(타입 속)이다.[53] 한 개의 융합된 전구 부속지, 유연하지만 뚜렷한 경계를 가진 체절, 최소한 지느러미 일부를 덮는 아가미, 여러 쌍의 꼬리지느러미, 한 쌍의 짧은 가시 모양 꼬리털이 특징으로, 라디오돈타류, 팸부델리온, 케리그마켈라와 구별된다.[120] 현재 확실히 오파비니아과에 포함되는 속은 오파비니아와 유타우로라[121] (''Utaurora'')뿐이다.[120] 유타우로는 체절 등면까지 덮는 아가미와 7쌍 이상 꼬리지느러미로 오파비니아(아가미는 체절을 덮지 않고, 꼬리지느러미 3쌍)와 구별된다.[120]
4. 1. 진화 논쟁
오파비니아의 분류학적 위치는 오랫동안 논쟁의 대상이었다. 1970년대 해리 휘팅턴(Harry B. Whittington)의 재기재(Whittington 1975) 이후 오파비니아는 갑각류는 물론 절지동물로 분류하기조차 어려운 독특한 특징을 가진 것으로 알려졌다.[54] 당시 학회에서 휘팅턴의 복원도가 발표되자 회장이 폭소로 뒤덮였다는 일화가 있을 정도였다.[54][119]휘팅턴은 오파비니아가 어떤 동물 문에도 속하지 않으며, 절지동물과 환형동물의 체절제를 가진 원시적인 계통군일 수 있다고 해석했다.[54] 스티븐 제이 굴드(Stephen Jay Gould)는 오파비니아를 현생 동물문에 분류 불가능한 "프로블레마티카"(미상 화석)로 버제스 동물군의 "기묘기천열 동물"의 대표적인 예로 언급했다.[56]
그러나 이러한 견해에 대한 반발이 있었고, 오파비니아는 절지동물에 포함될 가능성이 제시되었다.[55][79][124][57][67] 1990년대 이후 새로운 발견과 계통분류학의 발전에 의해 오파비니아와 아노말로카리스의 공통점(특화된 전부 부속지, 눈자루에 돌출된 겹눈, 몸통의 지느러미와 아가미, 미부 등)이 주목받았다.[55][71] 오파비니아의 입 끝 부분의 "가위"는 가로로 열리는 좌우 대칭 구조이며, 융합된 한 쌍의 부속지이다.[55][71] 아가미는 절지동물의 외엽과 매우 유사한 구조를 가졌다.[124][125][126][74]
1990년대에는 케리그마케라와 팜브델리온이 발견되었는데, 이들은 오파비니아, 아노말로카리스, 엽족동물의 특징을 모두 갖춘 중간형 생물(미싱 링크)이었다.[79][124][80][67] 이를 통해 오파비니아는 절지동물의 멸종된 초기 계통(스템 그룹)에 속하는 "기저" 절지동물이라는 설과, 절지동물은 엽족동물로부터 진화했다는 설이 대두되었다.[79][124][80][67]
아노말로카리스는 방사두류의 일원으로 정리되었다.[57] 오파비니아는 오파비니아류 (오파비니아과)의 일원으로서 방사두류와 함께 기저 절지동물 중에서 케리그마케라와 팜브델리온보다 파생적이며, 나머지 "진정한" 절지동물 (진절지동물)과 가장 근연인 군으로 여겨지게 되었다.[124][67] 21세기 이후에도 많은 계통 분석[77][91][92][93][94][95][96][97][98][75][99][100][101][102][68][103][104][105][106][107][108][109][110][111][112][113][114][115][69][116][117][118][120][70]과 새로운 발견[125][126] (예: 중간형 생물 킬릭시아[69])에 의해 이러한 계통 관계가 널리 지지되고 있다.
오파비니아의 지느러미와 아가미 구조는 재기재될 때마다 해석이 달라졌다.[119] 화석 표본의 각 체절과 지느러미 사이에는 삼각형 흔적이 한 쌍씩 나란히 존재하는데, 이에 대해서는 "소화관 가지"와 "부드러운 다리 (엽족)"라는 두 가지 가설이 있었다.[119]
- Whittington 1975: 제1 체절에 아가미는 없고, 아가미는 지느러미로부터 독립된 돌출된 근육으로 여겨졌다.[54] 삼각형 흔적은 소화관 가지, 혹은 아가미에 연결되는 순환계의 일부로 해석되었다.[54]
- Bergström 1986: 아가미는 다수의 가늘고 긴 잎 모양 부속체로 이루어져 있고, 지느러미 복면에 연결되는 빗살 모양 구조체로 판명되었다.[55]
- Budd 1996: 아가미는 지느러미 배면에 연결되고, 제1 체절은 아가미가 없다고 여겨졌다.[124] 삼각형 흔적은 엽족의 체강에서 유래된 흔적으로, 각 체절 복측에 한 쌍의 짧은 엽족이 있으며, 각 엽족 끝에 발톱이 있다고 추측되었다.[124]
- Zhang & Briggs 2007: 지느러미 뒷 가장자리에 큰 절개가 있고, 거기에 아가미가 있다고 해석했다.[125] 삼각형 흔적은 지느러미까지 삽입된 소화관 가지라고 해석되었다.[125]
- Budd & Daley 2011: 엽족의 존재 가능성을 지지하고, Budd 1996의 해석을 거의 인정하며, Zhang & Briggs 2007의 많은 판단을 부정했다.[126] 지느러미 뒷 가장자리는 완전하고, 제1 체절 지느러미에도 아가미가 있다고 판명되었다.[126] 삼각형 흔적은 엽족의 외피 조직과 유사한 환형 근육으로 갈라지는 구조가 발견되었다.[126]
5. 오파비니아의 의의
오파비니아는 캄브리아기 고생물 중에서도 형태학적 복원과 분류학적 위치에 대해 많은 논쟁이 벌어진 생물이다.[119] 20세기 초에는 갑각류로 여겨졌으며,[53] 20세기 후기에는 논쟁적인 미확인 화석(프로블레마티카)으로 분류되었다.[56] 1990년대 이후에는 오파비니아류의 일원으로서 아노말로카리스 등의 라디오돈트류와 함께 기반적인 절지동물로 널리 인정받고 있다.[124][67][125][126][119][58][59][60][61][62]
찰스 돌리틀 월콧은 버제스 셰일에서 거의 완전한 형태의 ''오파비니아 레갈리스'' 화석을 발견했다.[7] 오파비니아의 속명은 캐나다 브리티시컬럼비아주 레이크 오하라 남동쪽에 위치한 오파빈 패스에서 유래했다.[7]
5. 1. 대중문화 속 오파비니아
오파비니아는 그 독특한 외형 덕분에 여러 대중문화 작품에서 영감을 주었다. 버제스 셰일(버제스 동물군)에서 처음 발견되었을 때, 오파비니아는 갑각류 절지동물로 여겨졌다.[53] 미국의 고생물학자 찰스 왈콧은 1912년 논문에서 오파비니아를 현존하는 무갑류(논병아리새우와 갯강구를 포함)와 비교하며, 원시적인 무갑류와 유사하다고 생각했다. 또한, 이 종류의 무갑류 수컷은 앞에 갈라진 관이 있는데, 이를 연상시키는 관(전부 부속지)을 가진 오파비니아 화석 표본은 모두 수컷으로 해석되었다.[53]동물학자 조지 이블린 허친슨은 1930년에 논병아리새우처럼 배영하는 갑각류로 해석한 최초의 오파비니아 복원도를 발표했다.[88] 시모네타는 1970년에 오파비니아의 화석 표본에서는 보이지 않지만, 레안코일리야와 요호이아의 특징을 바탕으로 등판, 촉각, 관절로 나뉜 다리 등 절지동물적인 특징을 복원도에 추가했다.[89]
참조
[1]
논문
The enigmatic animal ''Opabinia regalis'', Middle Cambrian Burgess Shale, British Columbia
1975-06
[2]
논문
Taphonomy of the Greater Phyllopod Bed community, Burgess Shale
2006-10
[3]
논문
The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland)
https://www.cambridg[...]
1998
[4]
논문
Extraordinary fossils reveal the nature of Cambrian life: a commentary on Whittington (1975) 'The enigmatic animal Opabinia regalis, Middle Cambrian, Burgess Shale, British Columbia'
2015-04-19
[5]
논문
Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848
http://dx.doi.org/10[...]
2016
[6]
논문
Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective
2017-05-01
[7]
간행물
Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata
https://repository.s[...]
Smithsonian Miscellaneous Collections
1912
[8]
서적
Wonderful Life
Hutchinson Radius
[9]
문서
Rare paleontological remains and traces of life in late Cambrian deposits of the northwestern Siberian platform.
Palaeontology and biostratigraphy of the Soviet Arctic
1960
[10]
논문
The Taphonomy and Affinities of the Problematic Fossil Myoscolex from the Lower Cambrian Emu Bay Shale of South Australia
1997-01
[11]
논문
Anatomy and relationships of the Early Cambrian worm ''Myoscolex''
[12]
서적
Originations, Radiations and Biodiversity Changes – evidences from the Chinese fossil record.
2006
[13]
논문
An embryological perspective on the early arthropod fossil record
2015-12-18
[14]
논문
Phylogeny of the Myriapoda-Crustacea-Insecta: a new attempt using photoreceptor structure*
[15]
논문
''Opabinia'' and ''Anomalocaris'', unique Cambrian arthropods
[16]
논문
The lobes and lobopods of ''Opabinia regalis'' from the middle Cambrian Burgess Shale
[17]
논문
The morphology of ''Opabinia regalis'' and the reconstruction of the arthropod stem-group
[18]
논문
Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators
1994-05
[19]
논문
The nature and significance of the appendages of ''Opabinia'' from the Middle Cambrian Burgess Shale
[20]
간행물
Restudy of some Burgess shale fossils
https://repository.s[...]
Proceedings of the United States National Museum
1930
[21]
문서
Studies on non trilobite arthropods of the Burgess Shale (Middle Cambrian).
Palaeontogr. Ital.
1970
[22]
논문
Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa
2009
[23]
논문
The largest Cambrian animal, Anomalocaris, Burgess Shale, British-Columbia
https://royalsociety[...]
1985-05-14
[24]
논문
A Cambrian gilled lobopod from Greenland
[25]
서적
Arthropod Relationships – Special Volume Series 55
Systematics Association
[26]
논문
Anomalocaris and other large animals in the lower Cambrian Chengjiang fauna of southwest China
https://doi.org/10.1[...]
1995-09-01
[27]
논문
The "evolution" of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.)
https://www.cambridg[...]
1996
[28]
논문
Cambrian bivalved arthropod reveals origin of arthrodization
2012-12-07
[29]
논문
Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada)
https://www.cambridg[...]
2013
[30]
논문
The affinities of the cosmopolitan arthropod ''Isoxys'' and its implications for the origin of arthropods
https://onlinelibrar[...]
2013
[31]
논문
Arthropod fossil data increase congruence of morphological and molecular phylogenies
2013-09-30
[32]
논문
Hallucigenia's onychophoran-like claws and the case for Tactopoda
https://www.nature.c[...]
2014
[33]
논문
Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps
https://www.nature.c[...]
2015
[34]
논문
Hallucigenia's head and the pharyngeal armature of early ecdysozoans
https://www.nature.c[...]
2015
[35]
논문
Cambrian suspension-feeding lobopodians and the early radiation of panarthropods
2017-01-31
[36]
논문
A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK
2018
[37]
논문
An early Cambrian euarthropod with radiodont-like raptorial appendages
https://www.nature.c[...]
2020
[38]
논문
Stranger than a scorpion: a reassessment of ''Parioscorpio venator'', a problematic arthropod from the Llandoverian Waukesha Lagerstätte
https://onlinelibrar[...]
2021
[39]
논문
Exceptional multifunctionality in the feeding apparatus of a mid-Cambrian radiodont
2020
[40]
논문
Sophisticated digestive systems in early arthropods
2014-05-02
[41]
논문
New opabiniid diversifies the weirdest wonders of the euarthropod stem group
2022-02-09
[42]
논문
Middle Cambrian arthropods from Utah
https://www.cambridg[...]
2008
[43]
논문
Some problems and patterns of evolution exemplified by fossil invertebrates
[44]
서적
Time Frames: The Evolution of Punctuated Equilibria
1989
[45]
간행물
Neoproterozoic-Cambrian Biological Revolutions: Presented as a Paleontological Society Short Course at the Annual Meeting of the Geological Society of America, Denver, Colorado, November 6, 2004
Yale University Reprographics & Imaging Service; [[Paleontological Society]]
2015-02-06
[46]
논문
The fossil record and the origin of the Deuterostomata
[47]
서적
Wonderful Life
Hutchinson Radius
[48]
서적
The First Three Billion Years of Evolution on Earth
Princeton University Press
2009-04-22
[49]
논문
Affinities of ''Aysheaia'' (Onychophora), with Description of a New Cambrian Species
1985-01
[50]
논문
Molluscan engrailed Expression, Serial Organization, and Shell Evolution
[51]
논문
A new mitrate from the Upper Ordovician of Norway, and a new approach to subdividing a plesion
[52]
논문
The Cambrian Fossil Record and the Origin of the Phyla
[53]
간행물
Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata
https://hdl.handle.n[...]
[54]
논문
The enigmatic animal ''Opabinia regalis'', middle Cambrian, Burgess Shale, British Columbia
https://royalsociety[...]
1975-06-26
[55]
논문
''Opabinia'' and ''Anomalocaris'', unique Cambrian ‘arthropods’
https://onlinelibrar[...]
1986
[56]
서적
Wonderful life: the Burgess Shale and the nature of history
https://www.worldcat[...]
1989
[57]
논문
The “evolution” of ''Anomalocaris'' and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.)
https://www.cambridg[...]
1996-03
[58]
논문
Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848
http://eprints.esc.c[...]
[59]
논문
Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective
https://www.scienced[...]
2017-05-01
[60]
논문
Early fossil record of Euarthropoda and the Cambrian Explosion
https://www.pnas.org[...]
2018-05-22
[61]
논문
The Phylogeny and Evolutionary History of Arthropods
https://www.scienced[...]
2019-06-17
[62]
논문
Arthropod Origins: Integrating Paleontological and Molecular Evidence
https://www.annualre[...]
2020-11-02
[63]
웹사이트
Opabin Pass
http://www.peakfinde[...]
[64]
웹사이트
How Old is the Burgess Shale
https://burgess-shal[...]
2022-12-29
[65]
웹사이트
Opabinia regalis - The Burgess Shale
https://burgess-shal[...]
2011-06-10
[66]
논문
Dinocaridids – anomalous arthropods or arthropod-like worms?
http://citeseerx.ist[...]
2006
[67]
논문
The morphology and phylogenetic significance of ''Kerygmachela kierkegaardi'' Budd (Buen Formation, Lower Cambrian, N Greenland)
https://www.cambridg[...]
1998/ed
[68]
논문
Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps
https://www.research[...]
2015-03-11
[69]
논문
An early Cambrian euarthropod with radiodont-like raptorial appendages
https://www.nature.c[...]
2020-12
[70]
논문
A three-eyed radiodont with fossilized neuroanatomy informs the origin of the arthropod head and segmentation
https://linkinghub.e[...]
2022-07
[71]
논문
Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators
https://www.academia[...]
1994-05-27
[72]
논문
Cephalic and Limb Anatomy of a New Isoxyid from the Burgess Shale and the Role of “Stem Bivalved Arthropods” in the Disparity of the Frontalmost Appendage
https://journals.plo[...]
2015-06-03
[73]
논문
An embryological perspective on the early arthropod fossil record
https://www.research[...]
2015-12-18
[74]
논문
Exites in Cambrian arthropods and homology of arthropod limb branches
https://www.nature.c[...]
2021-07-30
[75]
논문
The affinities of the cosmopolitan arthropod ''Isoxys'' and its implications for the origin of arthropods
https://www.academia[...]
2013-09-16
[76]
논문
Sophisticated digestive systems in early arthropods
https://www.nature.c[...]
2014-05-02
[77]
논문
Morpho-anatomy of the lobopod ''Magadictyon'' cf. ''haikouensis'' from the Early Cambrian Chengjiang Lagerstätte, South China
https://www.research[...]
2007-04-19
[78]
논문
A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstätte
https://www.research[...]
[79]
논문
A Cambrian gilled lobopod from Greenland
https://www.nature.c[...]
1993-08
[80]
서적
"11. Stem group arthropods from the Lower Cambrian Sirius Passet fauna of North Greenland"
https://books.google[...]
Arthropod relationships
[81]
논문
Morphology of ''Anomalocaris canadensis'' from the Burgess Shale
https://www.academia[...]
2014-01
[82]
논문
Morphology of the radiodontan ''Lyrarapax'' from the early Cambrian Chengjiang biota
https://www.research[...]
2016-07-01
[83]
논문
Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan
https://academic.oup[...]
2018-06-01
[84]
논문
Arthropod visual predators in the early pelagic ecosystem: evidence from the Burgess Shale and Chengjiang biotas
https://www.research[...]
2009-07-22
[85]
논문
Gut glands illuminate trunk segmentation in Cambrian fuxianhuiids
https://www.research[...]
2018-02
[86]
논문
"Leanchoilia Guts and the Interpretation of Three-Dimensional Structures in Burgess Shale-Type Fossils"
https://www.jstor.or[...]
2002
[87]
논문
Controls on Gut Phosphatisation: The Trilobites from the Weeks Formation Lagerstätte (Cambrian; Utah)
https://journals.plo[...]
2012-03-14
[88]
논문
Restudy of some Burgess shale fossils
https://hdl.handle.n[...]
1930
[89]
간행물
Studies on non trilobite arthropods of the Burgess Shale (Middle Cambrian)
Palaeontogr. Ital.
1970
[90]
논문
Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa
https://evolution-ou[...]
2009-06
[91]
논문
A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany
https://www.research[...]
2009-02-06
[92]
논문
The Burgess Shale Anomalocaridid ''Hurdia'' and Its Significance for Early Euarthropod Evolution
https://www.research[...]
2009-03-20
[93]
논문
Morphology of ''Luolishania longicruris'' (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians
https://www.academia[...]
2009-07
[94]
논문
Phylogenetic position of ''Diania'' challenged
https://www.nature.c[...]
2011-08
[95]
논문
Lobopodian phylogeny reanalysed
https://www.nature.c[...]
2011-08
[96]
논문
Cambrian bivalved arthropod reveals origin of arthrodization
https://www.research[...]
2012-12-07
[97]
논문
Multi-Segmented Arthropods from the Middle Cambrian of British Columbia (Canada)
https://bioone.org/j[...]
2013-05-01
[98]
논문
The morphology and phylogenetic position of the Cambrian lobopodian ''Diania cactiformis''
https://www.research[...]
2013-05-29
[99]
논문
Arthropod fossil data increase congruence of morphological and molecular phylogenies
https://www.research[...]
2013-12
[100]
논문
A suspension-feeding anomalocarid from the Early Cambrian
https://research-inf[...]
2014-03
[101]
논문
"Hallucigenia 's onychophoran-like claws and the case for Tactopoda"
https://pubmed.ncbi.[...]
[102]
논문
Brain structure resolves the segmental affinity of anomalocaridid appendages
https://www.research[...]
2014-09
[103]
논문
"Hallucigenia’s head and the pharyngeal armature of early ecdysozoans"
https://pubmed.ncbi.[...]
2015-06-24
[104]
논문
A superarmored lobopodian from the Cambrian of China and early disparity in the evolution of Onychophora
https://www.research[...]
2015-07-14
[105]
논문
Onychophoran-like musculature in a phosphatized Cambrian lobopodian
https://www.ncbi.nlm[...]
2016-09
[106]
논문
Cambrian suspension-feeding lobopodians and the early radiation of panarthropods
https://bmcecolevol.[...]
2017-01-31
[107]
논문
Burgess Shale fossils illustrate the origin of the mandibulate body plan
https://www.nature.c[...]
2017-05
[108]
논문
Mandibulate convergence in an armoured Cambrian stem chelicerate
https://bmcecolevol.[...]
2017-12-21
[109]
논문
A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK
https://royalsociety[...]
2018-08-08
[110]
논문
New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton
https://www.nature.c[...]
2018-09-14
[111]
논문
Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan
https://academic.oup[...]
2018-11-01
[112]
논문
A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources
https://royalsociety[...]
2019-08-14
[113]
논문
Fossils from South China redefine the ancestral euarthropod body plan
https://doi.org/10.1[...]
2020-01-08
[114]
논문
A Tube-Dwelling Early Cambrian Lobopodian
https://www.cell.com[...]
2020-04-20
[115]
논문
The Collins’ monster, a spinous suspension-feeding lobopodian from the Cambrian Burgess Shale of British Columbia
https://onlinelibrar[...]
2020-07-10
[116]
논문
Stranger than a scorpion: a reassessment of ''Parioscorpio venator'', a problematic arthropod from the Llandoverian Waukesha Lagerstätte
https://onlinelibrar[...]
2021-04-19
[117]
논문
Exceptional multifunctionality in the feeding apparatus of a mid-Cambrian radiodont
https://www.cambridg[...]
2021-05-17
[118]
논문
"''Tabelliscolex'' (Cricocosmiidae: Palaeoscolecidomorpha) from the early Cambrian Chengjiang Biota, and the evolution of seriation in Ecdysozoa"
https://jgs.lyellcol[...]
2021-08-12
[119]
논문
Extraordinary fossils reveal the nature of Cambrian life: a commentary on Whittington (1975) ‘The enigmatic animal ''Opabinia regalis'', Middle Cambrian, Burgess Shale, British Columbia’
http://rstb.royalsoc[...]
2015-04-19
[120]
논문
New opabiniid diversifies the weirdest wonders of the euarthropod stem group
https://royalsociety[...]
2022-02-09
[121]
서적
カラー図説 生命の大進化40億年史 古生代編 生命はいかに誕生し、多様化したのか
講談社
2022-06-16
[122]
논문
A Cambrian unarmoured lobopodian, †''Lenisambulatrix humboldti'' gen. et sp. nov., compared with new material of †Diania cactiformis
https://www.nature.c[...]
2018-09-20
[123]
문서
Rare paleontological remains and traces of life in late Cambrian deposits of the northwestern Siberian platform
Palaeontology and biostratigraphy of the Soviet Arctic
1960
[124]
논문
The morphology of ''Opabinia regalis'' and the reconstruction of the arthropod stem-group
https://onlinelibrar[...]
1996
[125]
논문
The nature and significance of the appendages of ''Opabinia'' from the Middle Cambrian Burgess Shale
https://onlinelibrar[...]
2007
[126]
논문
The lobes and lobopods of ''Opabinia regalis'' from the middle Cambrian Burgess Shale
https://www.academia[...]
2011
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com