환형동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
환형동물은 작은 고리라는 뜻의 라틴어에서 유래되었으며, 체절 구조를 가진 동물의 문이다. 다모류, 빈모류, 거머리류 등으로 분류되었으나, 1997년 이후 분자 계통 발생학적 연구를 통해 분류 체계가 변화하였다. 현재는 다모류, 유수동물, 의충동물, 선충류 등이 환형동물에 포함되며, 뇌, 복측 신경삭 등으로 구성된 신경계와 호흡, 소화, 배설 기관을 갖는다. 무성생식과 유성생식을 하며, 트로코포어 유생을 거쳐 발생한다. 토양 비옥도에 기여하고, 낚시 미끼나 양식 생물의 먹이로 사용되며, 의학적 활용 가치도 있다. 화석 기록은 드물지만, 캄브리아기 화석이 발견되었으며, 분자 계통 분석을 통해 계통 분류가 이루어지고 있다.
더 읽어볼만한 페이지
- 환형동물 - 지렁이
지렁이는 환형동물에 속하며, 체절로 구성된 몸으로 흙 속이나 물속에서 생활하며, 토양 생태계에서 중요한 역할을 하고 낚시 미끼, 동물의 먹이, 약재 등으로 활용되며, 암수한몸으로 교미를 통해 번식한다. - 환형동물 - 다모류
다모류는 환형동물문에 속하는 강으로, 몸은 체절로 나뉘고 각 체절에 측족과 강모가 있으며, 이동성인 유형류와 정착성인 좌형류로 나뉘는 등 형태와 생활 방식이 다양한 해양 생물이다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 환형동물 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Annelida |
| 명명자 | Lamarck, 1809 |
| 어원 | 라틴어 (작은 고리) |
| 생물학적 분류 | |
| 계 | 동물계 |
| 아계 | 진정후생동물아계 |
| 상문 | 촉수담륜동물상문 |
| 문 | 환형동물문 (Annelida) |
| 하위 분류 (계통 발생학적 관점) | |
| 플라이스토안넬리다 하위 분류 | |
| 에란티아 하위 분류 | |
| 아시쿨라타 하위 분류 | |
| 플라이스토안넬리다 하위 분류 | |
| 세덴타리아 하위 분류 | |
| 카피텔리다 하위 분류 | |
| 세덴타리아 하위 분류 | |
| 하위 분류 (전통적 관점) | |
| 클리텔라타 하위 분류 | |
| 고생물학적 정보 | |
| 화석 범위 | 캄브리아기 제3절 (5억 6700만 년 전) ~ 현세 |
2. 역사적 분류와 최근의 변화
1997년 이전까지 대부분의 교과서는 환형동물을 다모류, 유두류로 분류했고, 유두류는 다시 빈모류와 거머리류로 나뉘었다.[17][8] 그러나 1997년 이후 분자 계통 발생학적 분석을 통해 환형동물의 진화 계통수에 대한 새로운 견해가 제시되면서 기존 분류 체계에 큰 변화가 생겼다.[67][69]
궁형환충은 해양 퇴적물 입자 사이에 사는 작은 환형동물로, 단순한 신체 구조 때문에 별도의 강으로 취급되었지만, 현재는 다모류로 간주된다.[8]
유주동물(Myzostomida)은 바다나리와 다른 극피동물에 기생하며, 과거에는 흡충류 편형동물 또는 완보동물과 관련이 있는 것으로 여겨졌으나, 1998년에는 다모류의 하위 그룹으로 제안되었다.[7] 그러나 2002년의 다른 분석에서는 편형동물 또는 윤충류 및 극두동물과 더 가깝다고 제안되었다.[10]
중생동물에 속하는 기생충인 Orthonectida는 환형동물로 확인되었으며,[15] Nemertea는 환형동물이며, Oweniidae와 Magelonidae가 가장 가까운 친척이다.[16]
1997년, Greg Rouse와 Kristian Fauchald는 형태학적 특징을 기반으로 다모류를 Scolecida, Palpata로 분류했고, Palpata는 다시 Canalipalpata와 Aciculata로 나뉜다고 하였다.[62]
같은 해, Damhnait McHugh는 분자 계통 발생학적 분석을 통해 유두류가 다모류 계통에서 파생되었고, 선충류와 의충류가 다모류 계통에 속한다는 다른 견해를 제시했다.[65] 이후의 연구들도 비슷한 결론을 뒷받침했다.[66]
2007년, Torsten Struck과 동료들은 81개 분류군에서 3개의 유전자를 비교하여 유두류, 선충류, 의충류가 다모류 계통에 속하며, Scolecida, Canalipalpata, Aciculata 분류는 유효하지 않다고 결론지었다. 또한, 성구동물을 다모류 계통에 포함시키고, 거머리가 유두류가 아닌 빈모류의 하위 그룹이라고 결론지었다.[67]
Rouse는 분자 계통 발생학적 분석을 수용했으며,[8] 주요 결론은 과학적 합의를 얻었지만, 세부 사항은 여전히 불확실하다.[69]
2. 1. 전통적 분류
다모류는 약 12,000종으로,[29] 각 체절마다 여러 개의 강모(털)가 있다. 다모류는 다리 역할을 하는 측지를 가지고 있으며, 두정기는 화학수용체로 여겨진다.[17] 대부분 해양 동물이지만, 일부 종은 담수에서, 더 적은 수는 육지에서 서식한다.[7]유두류는 약 10,000종으로,[5] 각 체절마다 강모가 거의 없거나 전혀 없으며, 두정기나 측지가 없다. 그러나 유두류는 독특한 생식 기관인 몸 주위에 고리 모양의 환대(clitellum) ("안장")를 가지고 있는데, 이는 수정란이 부화할 때까지 저장하고 영양을 공급하는 고치를 생성하거나,[8][9] 몬닐리가스트리드에서는 배아에 영양을 공급하는 노른자 알을 생성한다. 유두류는 다음과 같이 세분된다.[17]
- 빈모류("털이 적음")에는 지렁이가 포함된다. 빈모류는 입 천장에 끈적끈적한 패드가 있다.[17] 대부분은 완전히 또는 부분적으로 분해된 유기 물질을 먹는 굴착 동물이다.[7]
- 거머리류는 "거머리 모양"을 의미하며, 가장 잘 알려진 구성원은 거머리이다.[17] 해양 종은 주로 어류를 빨아먹는 기생충인 반면, 대부분의 담수 종은 포식자이다.[7] 거머리류는 몸의 양쪽 끝에 흡반이 있으며, 이를 사용하여 자벌레처럼 움직인다.[36]
2. 2. 분자 계통 분석에 따른 변화
1997년 이후의 연구로 환형동물의 진화 계통수에 대한 과학자들의 견해가 근본적으로 바뀌었다.[67][69] 분자 계통 발생학을 이용한 연구 결과, 다모류는 여러 계통으로 나뉘는 다계통군으로 밝혀졌다.유수동물(Pogonophora / Siboglinidae)은 1914년에 처음 발견되었으며, 내장이 없어 분류하기 어려웠다. 처음에는 별도의 문(Pogonophora)으로 분류되거나, 두 개의 문(Pogonophora, Vestimentifera)으로 나뉘기도 했다. 그러나 최근에는 다모류 내의 시보글리누스과로 재분류되었다.[7][10]
의충동물(Echiura)은 19세기에 "Gephyrea" 문에 속했으나, 이 문은 현재 구성원들이 다른 문으로 옮겨지면서 비어 있다. 1940년대까지 환형동물로 여겨지다가 별도의 문으로 분류되었으나, 1997년 분자 계통 발생학 분석 결과 환형동물로 결론지어졌다.[29][10][11]
성구동물(Sipuncula)은 분절, 강모 등 환형동물의 특징이 없어 처음에는 환형동물로 분류되었다. 이후 연체동물과 연관 지어졌으나, 리보솜 단백질 기반 계통 발생학적 분석 결과 환형동물 내에 위치하는 것으로 나타났다.[12] 이후 미토콘드리아 DNA 분석에서도 유주동물 및 환형동물(의충동물, 선충류 포함)과의 연관성이 확인되었다.[13] 또한, 성체에서는 없지만 초기 유충 단계에서 환형동물과 유사한 신경 분절이 발생한다는 사실도 밝혀졌다.[14]
이러한 변화는 과거 무척추동물 분류에서 강조되었던 분절의 중요성을 약화시킨다. 분자 계통 발생학 분석에서 다모류는 완전히 분절된 몸체를 가지지만, 의충동물과 성구동물은 분절되지 않았고, 선충류는 몸 뒷부분만 분절되어, 분절이 진화 과정에서 쉽게 나타나고 사라질 수 있음을 보여준다.[67][65]
분자 계통 분석 결과, 다모강과 그 안의 분류군은 해체되었으며,[80] 업데이트된 계통수는 다음과 같다.
{{clade|style=font-size:80%;|label1='''Annelida'''
|1={{clade
|1={{clade|label1=Palaeoannelida
|1={{clade
|1=Oweniidae
|2=Magelonidae
}}}}
|2={{clade
|1={{clade|label1=Chaetopteriformia
|1={{clade
|1=''Apistobranchus''
|2={{clade
|1=Psammodrilidae
|2=Chaetopteridae
}}
}}}}
|2={{clade
|1={{clade
|1=Amphinomida
|2={{clade
|1=''Lobatocerebrum''
|2=Sipuncula
}}
}}
|2={{clade|label1=Pleistoannelida |1={{clade
|label1=Errantia|1={{clade
|label2=Protodriliformia|2={{clade
|1=Polygordiidae
|2=Protodrilida
}}
|label1=Aciculata|1={{clade
|1=Eunicida
|2=Phyllodocida
}}
}}
|label2=''?''|2=Myzostomida|state2=dashed
|label3=Sedentaria|3={{clade
|1=Orbiniida
|2={{clade
|1={{clade
|1=
|2=
}}
|2={{clade
|1=}}
|2={{clade
|1=
|2=}}
}}}}
}}
}}
}}
}}
}}
}}
3. 주요 특징
환형동물은 대부분 몸의 내부 기관과 외부 강모가 동일하게 구성되어 있으며, 체절 구조를 가진다.[17][8] 몸은 길고 얕은 고리 모양으로 외부가 분절되어 있으며, 이를 환절이라고 한다. 내부는 격막("분할")으로 나뉘어 있다.[17] 대부분의 분절은 동일한 기관을 가지지만, 내장, 순환계, 신경계를 공유하여 상호 의존적이다.[17][8]
환형동물은 다른 무척추동물 문과 구별되는 단일 특징은 없지만, 독특한 특징 조합을 가진다.
| 환형동물 | 별벌레[21] | 총채벌레[22] | 끈벌레[23] | 절지동물[24] | 유조동물[25] | |
|---|---|---|---|---|---|---|
| 외부 분절 | 예 | 아니오 | 아니오 | 소수 종 | 진드기 제외 예 | 아니오 |
| 내부 기관 반복 | 예 | 아니오 | 아니오 | 예 | 원시 형태 | 예 |
| 분절 사이 격막 | 대부분 | 아니오 | 아니오 | 아니오 | 아니오 | 아니오 |
| 큐티클 재료 | 콜라겐 | 콜라겐 | 콜라겐 | 없음 | α-키틴 | α-키틴 |
| 탈피 | 일부 다모류는 턱, 거머리는 피부 탈피[26], 그 외는 아님[18] | 아니오[27] | 아니오[27] | 아니오[27] | 예[20] | 예[20] |
| 체강 | 체강 (일부 감소/없음)[18] | 2개 (주 체강, 입술 내) | 2개 (주 체강, 촉수 내) | 입술 내 | 혈강 | 혈강 |
| 순환계 | 대부분 폐쇄형 | 개방형 (분지 정맥 귀환) | 개방형 | 폐쇄형 | 개방형 | 개방형 |
3. 1. 체절 구조
환형동물은 몸이 여러 개의 체절로 나뉘어 있으며, 각 체절은 '''격막'''(septa|세프타영어)으로 구분된다.[84] 가장 앞쪽의 '''구전엽'''(prostomium|프로스토미움영어)에는 뇌와 감각 기관이 있고,[86][85] 가장 뒤쪽의 '''미절'''(pygidium|피기디움영어)에는 항문이 있다.[84][85] 체절은 미절 바로 앞의 생장대에서 하나씩 발달하며, 가장 어린 체절은 생장대 바로 앞에, 구주는 가장 오래되었다.[17]
다모류는 기본 구조가 매우 유사한 다수의 환상 체절이 1열로 연결되어 있으며,[83] 성체의 몸통에는 체절이 이어지고, 몸통은 흉부와 복부로 분화되기도 한다.[84] 구전엽 및 구주절에 더하여 수 체절이 변형되어 머리 부분을 형성한다.[84]
거머리류는 몸의 전후단에 각각 흡반을 가짐으로써 체절이 현저하게 변형되어 있다.[84] 흡구충류에는 불완전한 체절이 있다.[85]
유설동물은 전체(protosoma|프로토소마영어), 중체(mesosoma|메소소마영어), 몸통(metasoma|메타소마영어), 후체(opistosoma|오피스토소마영어)의 4부분으로 나뉘며, 매우 짧은 후체에만 강모를 갖춘 복수의 체절(강모절)이 존재한다.[83][86]
3. 2. 몸의 구조
환형동물의 몸은 콜라겐으로 이루어진 큐티클 층으로 덮여 있다.[17] 큐티클은 세포를 포함하지 않지만, 그 아래 피부 세포에서 분비되며 질기고 유연하다.[17] 절지동물의 큐티클은 더 단단한 α-키틴으로 만들어져 있으며,[17][19] 성장하면서 탈피를 하지만,[20] 환형동물의 큐티클은 탈피하지 않는다.[18] 큐티클은 콜라겐 섬유로 이루어져 있으며, 일반적으로 섬유가 서로 교차하도록 교대로 나선형으로 배열된 층을 이룬다. 이는 단층의 표피(가장 바깥쪽 피부층)에서 분비된다.[17]표피 아래에는 결합 조직으로 구성된 진피가 있고, 그 아래에는 두 개의 근육층이 발달해 있다.[17] 바깥쪽의 환상근은 수축하면 마디를 더 길고 가늘게 만들고, 안쪽의 종주근은 수축하면 마디를 더 짧고 굵게 만든다.[17] 몇몇 환형동물은 몸의 아랫면 양쪽에 사선형 내부 근육을 가지고 있다.[18]

환형동물의 강모("털")는 표피에서 돌출되어 견인력 등의 기능을 제공한다.[17] 강모는 적당히 유연한 β-키틴으로 만들어지며, 각각 바닥에 강모모세포("털 형성")와 강모를 연장하거나 수축할 수 있는 근육을 가진 모낭에 의해 형성된다.[17]
거의 모든 다모류는 사지 역할을 하는 측각을 가지고 있는 반면, 다른 주요 환형동물 그룹은 측각이 없다. 측각은 몸통 벽의 관절이 없는 쌍을 이루는 확장부이며, 그들의 근육은 몸의 환상근에서 파생된다.[18]
뇌는 일반적으로 인두 (목구멍) 주위에 고리 모양을 형성하며, 인두 위와 앞쪽에 한 쌍의 신경절 (지역 제어 센터)로 구성되어 있다.[17] 다모류의 뇌는 일반적으로 전구절에 위치하며, 환대류의 뇌는 구절 또는 때로는 전구절 뒤의 첫 번째 마디에 위치한다.[31] 나머지 중추 신경계, 복측 신경삭은 일반적으로 "사다리 모양"이며, 몸의 아래쪽 부분을 통과하는 한 쌍의 신경삭으로 구성되어 있으며, 각 마디에는 가로 연결로 연결된 신경절 쌍이 있다.[17]
감각기는 주로 빛, 화학 물질, 압력파 및 접촉을 감지하는 단일 세포이며, 머리, 부속지 (있는 경우) 및 몸의 다른 부분에 존재한다.[17] 일부 다모류는 빛이 오는 방향을 감지하는 다양한 조합의 단안 ("작은 눈")과 이미지를 형성할 수 있는 카메라형 눈 또는 겹눈을 가지고 있다.[32][33]
대부분의 환형동물은 혈액이 혈관을 통해 전체 순환을 하는 폐쇄 순환계를 가지고 있다.[18] 잘 발달된 중격을 가진 종은 일반적으로 내장 위아래로 몸 전체에 혈관이 분포되어 있으며, 위쪽 혈관은 혈액을 앞으로 운반하고 아래쪽 혈관은 뒤로 운반한다. 체벽과 내장 주변의 모세 혈관 네트워크는 주요 혈관 사이와 산소와 영양분이 필요한 체절의 일부로 혈액을 전달한다.[17]
3. 3. 신경계
환형동물의 신경계는 머리 부분 등쪽의 발달된 중추 신경절(머리 신경절[83]), 신경환, 배쪽 신경삭으로 구성된다.[85] 다모류에서는 대부분 각 체절에 있는 한 쌍의 신경절이 연락하여 배쪽 신경삭을 형성하지만, 환대류에서는 중앙에서 1개로 융합되어 있다.[84] 심해벌레류에서는 체단부의 2개의 신경삭이 털집부 후부에서 1개로 융합되어 있다.[84] 멍게벌레동물에서는 1개의 배쪽 신경삭이 뻗어 있으며, 그 전단은 2개로 분기하여 주둥이의 주연부를 환상으로 1바퀴 돌지만, 뇌나 신경절은 형성되지 않는다.[87] 별벌레동물의 신경계는 뇌, 배쪽 신경삭 및 그것들을 잇는 식도 신경환으로 구성된다.[88] 뇌는 소형이며 인두 등벽에 접하며, 배쪽 신경삭은 배의 정중앙을 따라 뻗어 있으며 1개로 분절되지 않는다.[88]3. 4. 호흡, 소화 및 배설
지렁이와 같은 일부 환형동물은 피부를 통해 호흡한다. 그러나 많은 다모류와 일부 환대류는 대부분의 체절에 아가미를 가지고 있으며, 다모류의 경우 측각의 연장으로 나타나는 경우가 많다. 관에 살거나 땅을 파고 사는 종의 아가미는 일반적으로 물의 흐름이 더 강한 쪽 끝 부분에 모여 있다.[18]
입 부위의 먹이 섭취 구조는 매우 다양하며, 동물의 식단과 거의 관련이 없다. 많은 다모류는 뒤집을 수 있는(안쪽으로 뒤집어 뻗을 수 있는) 근육질의 인두를 가지고 있다. Eunicidae와 Phyllodocidae의 두 과는 턱을 진화시켜 먹이를 잡거나, 식물 조각을 물어뜯거나, 죽거나 썩어가는 물질을 움켜잡는 데 사용할 수 있다. 반면에 일부 포식성 다모류는 턱도 뒤집을 수 있는 인두도 없다. 선택적 퇴적물 섭식자는 일반적으로 해저 튜브에서 살며 촉수를 사용하여 퇴적물에서 음식 입자를 찾아 입 안으로 닦아 넣는다. 여과 섭식 동물은 섬모로 덮인 촉수 "관"을 사용하여 음식 입자를 입으로 쓸어 넣는다. 비선택적 퇴적물 섭식자는 일반적으로 특화되지 않은 입을 통해 흙이나 해양 퇴적물을 섭취한다. 일부 환형동물은 입 천장에 끈적끈적한 패드를 가지고 있으며, 이 중 일부는 패드를 뒤집어 먹이를 잡을 수 있다. 거머리는 종종 뒤집을 수 있는 코나 두세 개의 이빨이 있는 근육질 인두를 가지고 있다.[18]
장은 일반적으로 장간막(마디 내의 수직 칸막이)에 의해 지지되는 거의 곧은 튜브이며, 항문으로 끝나는데, 이는 피지디움의 아랫면에 위치한다.[17] 튜브에 사는 Siboglinidae과의 구성원에서는 장이 부풀어 오른 내벽에 막혀 있는데, 이 내벽에는 웜 전체 무게의 15%를 차지할 수 있는 공생 세균이 있다. 박테리아는 무기 물질, 예를 들어 열수 분출구에서 나오는 황화 수소와 이산화 탄소, 또는 유전 분출구에서 나오는 메탄을 유기 물질로 전환하여 자신과 숙주에게 먹이를 공급하는 반면, 웜은 박테리아에 필요한 가스를 흡수하기 위해 가스 흐름으로 촉수를 뻗는다.[18]
혈관이 있는 환형동물은 후네프리디움을 사용하여 용해성 노폐물을 제거하는 반면, 혈관이 없는 동물은 전네프리디움을 사용한다.[17] 환형동물에서 체액이 전네프리디움 또는 후네프리디움으로 들어가는 지점은 격막의 앞쪽에 있으며, 두 번째 단계 여과기와 신장공(신체벽의 출구)은 다음 마디에 있다.[17]
4. 생식과 발생
다모류, 굴뚝벌레류, 갯지렁이, 별벌레는 연체동물처럼 나선 난할을 하여 트로코포어 유생을 거쳐 발생한다.[84][86][88][87] 시리스류, 고리벌레류는 수정된 알이 보육되어, 배(胚)는 정착 준비가 된 유생으로 모체에서 방출된다.[84] 별벌레 중에는 직접 발생을 하는 것도 있지만, 트로코포어 유생을 거치는 것 중에서도 트로코포어에서 성체가 되거나, 난황 영양성 펠라고사페라 유생 (pelagosphera larva|펠라고사페라 유생영어)을 거쳐 성체로 변태하거나, 섭식성 펠라고사페라 유생이 되어 장기간 유영하기도 한다.[88]
트로코포어 유생은 팽이 모양이며, 원구에서 유래하는 입을 사이에 두고 입앞 섬모환 (prototroch|프로토트로크영어), 입뒤 섬모환 (metatroch|메타트로크영어), 항문 바로 위에 말단 섬모환 (telotroch|텔로트로크영어)의 3개 섬모환을 가진다.[84] 후기 트로코포어 유생의 입앞 섬모환 앞쪽은 상체 (episphere|에피스피어영어)라고 하며, 성체의 입앞엽에 해당한다.[84] 성체의 꼬리마디에 해당하는 부분은 꼬리마디부라고 한다.[84]
환대류는 제한된 체절에만 생식소가 발달하며, 몸 앞쪽 몇 개 체절을 덮어 수정란의 난포를 분비하는 '''환대'''가 있다.[84] 암수 생식기를 모두 사용하여 교접하여 정포를 교환한다. 수정란은 난포에 싸여 발생하며, 유생을 거치지 않고 유충이 부화한다.[84]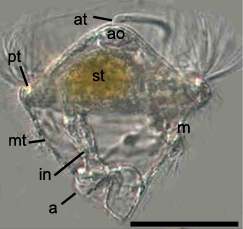
4. 1. 무성 생식

다모류는 두 개 이상의 조각으로 분열하거나 부모 개체가 완전한 상태로 유지되는 동안 새로운 개체를 출아하여 무성 생식을 할 수 있다.[17][40] 일부 빈모류, 예를 들어 ''Aulophorus furcatus''는 완전히 무성적으로 번식하는 것으로 보이며, 다른 종은 여름에는 무성적으로, 가을에는 유성적으로 번식한다. 빈모류의 무성 생식은 출아가 아니라 항상 두 개 이상의 조각으로 분열하는 방식으로 이루어진다.[8][38] 그러나 거머리는 무성 생식을 하는 모습이 관찰된 적이 없다.[8][39]
4. 2. 유성 생식
환형동물은 원래 두 개의 성별을 가진 동물로, 신관을 통해 물속으로 난자와 정자를 방출하는 것으로 생각된다.[17] 수정된 난자는 트로코포어 유생으로 발달하여 플랑크톤으로 살아간다.[43] 이후 이들은 해저로 가라앉아 미니어처 성체로 변태한다.그러나 대부분의 살아있는 다모류의 생활사는 알려져 있지 않으며, 이들은 거의 모두 해양 동물이다. 생활사가 알려진 300개 이상의 종 중 약 25%만이 이러한 패턴을 따른다. 약 14%는 유사한 체외 수정을 사용하지만 난황이 풍부한 난자를 생성하여 유생이 플랑크톤 사이에서 보내야 하는 시간을 줄이거나, 유생이 아닌 미니어처 성체가 나타나는 난자를 생산한다. 나머지는 수정된 알이 부화할 때까지 돌본다. 어떤 종은 알의 젤리 덩어리를 생성하여 돌보고, 어떤 종은 알을 몸에 부착하며, 일부 종은 부화할 때까지 몸 안에 알을 보관한다. 이 종들은 정자 전달을 위한 다양한 방법을 사용한다. 예를 들어, 일부 종의 암컷은 물에 방출된 정자를 수집하는 반면, 다른 종의 수컷은 암컷에게 정자를 주입하는 음경을 가지고 있다.[43]
일부 다모류는 일생에 한 번만 번식하는 반면, 다른 다모류는 거의 지속적으로 또는 여러 번의 번식기를 거쳐 번식한다. 대부분의 다모류는 평생 한쪽 성별로 남아 있지만, 상당한 비율의 종은 완전한 자웅동체이거나 일생 동안 성별을 바꾼다. 번식이 연구된 대부분의 다모류는 영구적인 생식선이 없으며, 어떻게 난자와 정자를 생성하는지 불확실하다. 몇몇 종에서는 몸의 뒷부분이 분리되어 적절한 환경, 일반적으로 표면 근처에서 유영하고 산란하기에 충분한 시간 동안만 사는 개체가 된다.[43]
대부분의 성숙한 환대류(지렁이와 거머리를 포함하는 그룹)는 완전한 자웅동체이지만, 몇몇 거머리 종에서는 어린 성체가 수컷으로 기능하다가 성숙기에 암컷이 된다. 모두 발달된 생식선을 가지고 있으며 모두 교미한다. 지렁이는 파트너의 정자를 정자낭("정자 저장소")에 저장한 다음 환대가 난소에서 난자를 수집한 다음 정자낭에서 정자를 수집하는 고치를 생성한다. 지렁이 알의 수정과 발달은 고치 내에서 일어난다. 거머리의 알은 난소에서 수정된 후 고치로 옮겨진다. 모든 환대류에서 고치는 알이 수정될 때 난황을 생성하거나 발달하는 동안 영양분을 공급한다. 모든 환대류는 유생이 아닌 미니어처 성체로 부화한다.[43]
다모류, 굴뚝벌레류, 갯지렁이, 별벌레에서는 연체동물과 마찬가지로 나선 난할을 하여 트로코포어 유생을 거쳐 발생이 진행된다.[84][86][88][87] 시리스류, 고리벌레류에서는 수정된 알이 보육되어, 배는 정착 준비가 갖춰진 유생으로 모체로부터 방출된다.[84] 또한, 별벌레 중에는 직접 발생을 하는 것도 있지만, 트로코포어 유생을 거치는 것 중에서도 트로코포어에서 성체로 되는 것, 트로코포어에서 난황 영양성 펠라고사페라 유생 (pelagosphera larva영어)이 되어 성체로 변태하는 것, 트로코포어에서 섭식성 펠라고사페라 유생이 되어 장기간 유영하는 것도 있다.[88]
환대류는 제한된 체절에만 생식소가 발달하며, 몸 앞쪽의 몇 개 체절을 덮어 수정란의 난포를 분비하는 '''환대'''를 가진다.[84] 암컷 생식기와 수컷 생식기를 동시에 사용하여 교접하여 정포를 교환한다. 수정란은 난포에 싸여 발생이 진행되고, 유생을 거치지 않고 유충이 부화한다.[84]
5. 생태적 중요성
지렁이는 토양 생태계에서 중요한 역할을 하며, 다양한 동물의 먹이가 된다. 또한, 해양 환형동물은 산호초와 조수 구역 생태계에서 중요한 구성원이다. 그러나 일부 육상 환형동물은 침입종으로 생태계를 교란시키기도 하며, 다른 동물에게 질병을 옮기기도 한다.
북미 빙하 지역에서는 토착 지렁이가 거의 사라지고 유럽과 아시아에서 유입된 지렁이가 서식한다. 특히 ''Amynthas agrestis''는 북부 경엽수림에 낙엽 부엽토 손실, 토양 비옥도 감소, 토양 화학 변화, 생태적 다양성 감소 등 부정적인 영향을 미쳐 우려되는 종이다. 위스콘신 주에서는 이 종을 금지종으로 지정했다. 지렁이는 낚시꾼, 차량 타이어, 신발 등에 묻어 빠르게 확산될 수 있다.
5. 1. 토양 생태계에서의 역할
찰스 다윈의 저서 《지렁이의 활동을 통한 토양 형성》(1881)은 지렁이가 토양 비옥도에 기여하는 바를 최초로 과학적으로 분석한 것이다.[44] 지렁이는 굴을 파거나 땅 표면에서 생활하면서 토양을 생성하는 데 도움을 준다.[45][46] 굴을 파는 종은 토양을 느슨하게 하여 산소와 물이 침투할 수 있게 하고, 표면 및 굴을 파는 지렁이 모두 유기물과 무기물을 섞고, 유기물의 분해를 가속화하여 다른 유기체가 더 빨리 이용할 수 있게 하며, 광물을 농축하여 식물이 더 쉽게 사용할 수 있는 형태로 변환한다.[45][46]5. 2. 다른 생물과의 관계
환형동물은 다양한 동물의 먹이가 되며, 일부 종은 다른 동물에 기생하여 질병을 유발하기도 한다.지렁이는 울새에서 황새에 이르는 크기의 조류와 땃쥐에서 오소리에 이르는 포유류의 중요한 먹이이다.[47] 어떤 경우에는 멸종 위기에 처한 조류를 보존하기 위해 지렁이를 보존하는 것이 필수적일 수 있다.[47]
해양 환형동물은 산호초 주변과 조수 구역에서 저서 동물 종의 3분의 1 이상을 차지할 수 있다.[44] 굴을 파는 종은 물과 산소가 해저 퇴적물로 침투하는 것을 증가시켜 호기성 생물과 작은 동물의 굴 주변에서 개체군 성장을 촉진한다.[48]
피를 빨아먹는 거머리는 희생자에게 직접적인 피해를 거의 주지 않지만, 일부는 숙주에게 매우 위험할 수 있는 편모충을 전염시킨다. 일부 작은 관을 짓는 빈모류는 물고기에게 와동병을 유발하는 미포자충 기생충을 전염시킨다.[44]
6. 인간과의 관계
갯지렁이를 포함한 일부 환형동물은 낚시 미끼로 사용된다.[51] 과거에는 거머리가 의료 목적으로 널리 사용되었으나, 현대 의학에서는 그 사용이 줄었다. 하지만 최근에는 미세 수술 과정에서 혈액 순환을 돕거나 항응고제 개발과 같은 분야에서 다시 주목받고 있다.[52] 또한, 환형동물은 수질 오염의 정도를 나타내는 지표종으로 활용되기도 한다.[53]
7. 진화 역사와 계통 분류
1997년 이후의 연구들은 환형동물의 진화 계통수에 대한 과학자들의 견해를 근본적으로 바꾸어 놓았다.[67][69] 대부분의 교과서는 전통적으로 환형동물을 다모류와 환대류로 분류하며, 환대류는 다시 빈모류(지렁이 포함)와 거머리류(거머리 포함)로 나뉜다.[17][8]
궁형환충은 해양 퇴적물 입자 사이에 사는 작은 환형동물로, 단순한 신체 구조 때문에 별도의 강으로 취급되었지만, 현재는 다모류로 간주된다.[8] Pogonophora / Siboglinidae는 1914년에 처음 발견되었으며, 내장이 없어 분류하기 어려웠다. 처음에는 별도의 문(Pogonophora)으로 분류되거나 두 개의 문(Pogonophora, Vestimentifera)으로 분류되었으나, 최근에는 다모류 내의 과(Siboglinidae)로 재분류되었다.[7][10] Echiura는 19세기에 "Gephyrea" 문에 속했다가, 1940년대까지 환형동물로 간주된 후 자체 문으로 분류되었다. 그러나 1997년 분자 계통 발생학 분석 결과, Echiura는 환형동물로 결론지어졌다.[29][10][11]
Myzostomida는 바다나리 등 극피동물에 기생하며, 과거에는 흡충류 편형동물이나 완보동물과 관련이 있는 것으로 여겨졌으나, 1998년에는 다모류의 하위 그룹으로 제안되었다.[7] 그러나 2002년 분석에서는 편형동물이나 윤충류, 극두동물과 더 가깝다고 제안되었다.[10] Sipuncula는 분절, 강모 등 환형동물의 특징이 없어 처음에는 환형동물로 분류되었으나, 나중에 연체동물과 연합되었다. 그러나 79개 리보솜 단백질 기반 계통 발생학적 분석 결과, Sipuncula는 환형동물 내에 위치하는 것으로 나타났다.[12] 이후 미토콘드리아 DNA 분석에서도 Myzostomida, 환형동물(Echiurans, pogonophorans 포함)과의 관계가 확인되었다.[13] 또한, 성체에서는 없더라도 초기 유충 단계에서 환형동물과 유사한 신경 분절이 발생한다는 것이 밝혀졌다.[14]
미토게놈 및 계통 유전체 분석 결과, 중생동물에 속하는 기생충 그룹인 Orthonectida도 환형동물로 밝혀졌다.[15] 연구에 따르면 nemerteans는 환형동물이며, Oweniidae와 Magelonidae가 가장 가까운 친척이다.[16]
환형동물은 다른 무척추동물 문들과 구별되는 단일 특징은 없지만, 독특한 특징 조합을 가지고 있다. 몸은 길고, 얕은 고리 모양 수축으로 외부적으로 나뉘는 분절로 이루어져 있으며, 이를 환절이라고 한다. 또한 내부적으로는 동일 지점에 격막("분할")으로 나뉘어져 있다. 그러나 일부 종에서는 격막이 불완전하며, 몇몇 경우에는 존재하지 않는다. 대부분의 분절은 동일한 기관 세트를 포함하고 있지만, 공통적인 내장, 순환계, 신경계를 공유하여 상호 의존적이다.[17][8] 몸은 큐티클로 덮여 있는데, 세포를 포함하지 않지만, 그 아래 피부 세포에서 분비되며, 질기고 유연한 콜라겐으로 만들어져 있고,[17] 탈피하지 않는다.[18] 반면 절지동물의 큐티클은 더 단단한 α-키틴으로 만들어져 있으며,[17][19] 절지동물이 완전한 크기에 도달할 때까지 탈피한다.[20] 대부분의 환형동물은 혈액이 혈관을 통해 전체 순환하는 폐쇄 순환계를 가지고 있다.[18]
| 환형동물[17] | 최근 환형동물에 병합됨[67] | 가깝게 관련된 종 | 유사한 외형의 문 | |||
|---|---|---|---|---|---|---|
| 별벌레[21] | 총채벌레[22] | 끈벌레[23] | 절지동물[24] | 유조동물[25] | ||
| 외부 분절 | 예 | 아니오 | 소수의 종에서만 | 진드기를 제외하고 예 | 아니오 | |
| 내부 기관 반복 | 예 | 아니오 | 예 | 원시 형태에서 | 예 | |
| 분절 사이 격막 | 대부분의 종에서 | 아니오 | ||||
| 큐티클 재료 | 콜라겐 | 없음 | α-키틴 | |||
| 탈피 | 일반적으로 아니오;[18] 그러나 일부 다모류는 턱, 거머리는 피부 탈피.[26] | 아니오[27] | 예[20] | |||
| 체강 | 체강; 일부 거머리와 작은 다모류는 감소/미존재[18] | 주 체강, 입술 내 두 체강 | 주 체강, 촉수 내 두 체강 | 입술 내에서만 체강 | 혈강 | |
| 순환계 | 대부분 폐쇄형 | 개방형 유출, 분지 정맥 귀환 | 개방형 | 폐쇄형 | 개방형 | |
1997년 Greg Rouse와 Kristian Fauchald는 해부학적 구조를 기반으로 다모류를 Scolecida, Palpata(Canalipalpata, Aciculata)로 분류했다.[62] 2007년 Torsten Struck과 동료들은 유전자 비교를 통해 환대류, 선충류, 치설류가 다모류 계통수 내에 위치하며, sipunculans 또한 다모류 계통수에, 거머리는 빈모류의 하위 그룹이라는 결론을 내렸다.[67]
분자 계통 발생학적 분석은 분절의 중요성을 훼손한다. 다모류는 분절된 몸체를 가지지만, 치설류와 sipunculans는 분절되지 않았고, 선충류는 몸 뒷부분만 분절된다. 분절은 이전 생각보다 진화 과정에서 더 쉽게 나타나고 사라지는 것으로 보인다.[67][65]
7. 1. 화석 기록
환형동물의 화석 기록은 드물지만, 캄브리아기 초기부터 다양한 화석이 발견되고 있다. 화석 발견을 통해 환형동물과 유촉수동물이 다른 어떤 문보다 서로 더 밀접하게 관련되어 있다는 가설이 제기되었다. lophotrochozoan 화석의 체제 때문에, 계통 발생 분석은 유촉수동물을 환형동물의 자매군으로 간주한다. 두 그룹은 미세융모에 의해 분비되는 강모(chaetae)의 존재, 쌍을 이루고 분절된 체강 구획, 그리고 유사한 신장(metanephridium) 구조와 같은 공통점을 가지고 있다.[79]7. 2. 계통 분류
분자 계통 분석 결과, 환형동물문의 계통 관계는 크게 변화하고 있으며, 새로운 분류 체계가 정립되고 있다. 1997년 이후의 연구들은 환형동물의 진화 계통수에 대한 과학자들의 견해를 근본적으로 바꾸어 놓았다.[67][69]현재 환형동물문은 크게 다음과 같이 분류된다:
- Palaeoannelida
- Chaetopteriformia
- Amphinomida/Sipuncula/''Lobatocerebrum'' 클레이드
- Pleistoannelida (Errantia와 Sedentaria 포함)
{{clade|style=font-size:80%;|label1='''Annelida'''
|1={{clade
|1={{clade|label1=Palaeoannelida
|1={{clade
|1=Oweniidae
|2=Magelonidae
}}
}}
|2={{clade
|1={{clade|label1=Chaetopteriformia
|1={{clade
|1=''Apistobranchus''
|2={{clade
|1=Psammodrilidae
|2=Chaetopteridae
}}
}}
}}
|2={{clade
|1={{clade
|1=Amphinomida
|2={{clade
|1=''Lobatocerebrum''
|2=Sipuncula
}}
}}
|2={{clade|label1=Pleistoannelida
|1={{clade
|label1=Errantia
|1={{clade
|label2=Protodriliformia
|2={{clade
|1=Polygordiidae
|2=Protodrilida
}}
|label1=Aciculata
|1={{clade
|1=Eunicida
|2=Phyllodocida
}}
}}
|label2=''?''
|2=Myzostomida|state2=dashed
|label3=Sedentaria
|3={{clade
|1=Orbiniida
|2={{clade
|1={{clade
|1=
|2=
}}
|2={{clade
|1=}}
|2={{clade
|1=
|2=
}}
}}
}}
}}
}}
}}
}}
}}
}}
분자 계통 발생학적 분석은 환대류, 선충류, 치설류가 다모류 계통수의 다양한 분기에 있다는 것에 동의한다.[67] 또한, sipunculans를 다모류 계통수의 다른 분기에 배치했으며, 거머리가 환대류에서 그들의 자매 그룹이라기보다는 빈모류의 하위 그룹이라는 결론을 내렸다.[67]
이러한 분석 결과는 수십 년 동안 무척추동물 분류에서 분절의 중요성을 강조했던 이전의 저술들을 훼손한다. 다모류는 완전히 분절된 몸체를 가지고 있는 반면, 다모류의 치설류와 sipunculans 분지들은 분절되지 않았고, 선충류는 몸의 뒷부분에서만 분절된다. 이제 분절은 이전에 생각했던 것보다 진화 과정에서 훨씬 더 쉽게 나타나고 사라질 수 있는 것으로 보인다.[67][65]
참조
[1]
논문
Guts, gut contents, and feeding strategies of Ediacaran animals
[2]
서적
[3]
서적
[4]
논문
Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary
http://www.cugb.edu.[...]
2013-02-07
[5]
서적
Cosmopolitan Earthworms
VermEcology, Yokohama.
[6]
논문
Diversity of Soil Fauna and Ecosystem Function
http://horizon.docum[...]
2009-04-20
[7]
서적
Invertebrate Zoology
Oxford University Press
[8]
서적
Invertebrate Zoology
Oxford University Press
[9]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[10]
논문
Unsegmented Annelids? Possible Origins of Four Lophotrochozoan Worm Taxa
[11]
논문
Molecular evidence that echiurans and pogonophorans are derived annelids
1997-07
[12]
논문
Spiralian Phylogenomics Supports the Resurrection of Bryozoa Comprising Ectoprocta and Entoprocta
[13]
논문
A close phylogenetic relationship between Sipuncula and Annelida evidenced from the complete mitochondrial genome sequence of Phascolosoma esculenta
[14]
논문
Sipunculans and segmentation
2009-01
[15]
서적
Annelida
https://books.google[...]
[16]
웹사이트
Mitochondrial Genome Evolution in Annelida—A Systematic Study on Conservative and Variable Gene Orders and the Factors Influencing its Evolution
https://academic.oup[...]
[17]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[18]
서적
Invertebrate Zoology
Oxford University Press
[19]
논문
Arthropod cuticle features and arthropod monophyly
1980-08
[20]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[21]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[22]
서적
Invertebrate Zoology
Oxford University Press
[23]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[24]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[25]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[26]
논문
Molting polychaete jaws—ecdysozoans are not the only molting animals
2005-06
[27]
논문
Proposing a solution to the Articulata–Ecdysozoa controversy
http://www.museunaci[...]
2009-03-11
[28]
서적
The Invertebrate Tree of Life
https://books.google[...]
[29]
서적
Encyclopedia of Life Sciences
John Wiley & Sons
[30]
간행물
Perspectives in Animal Phylogeny and Evolution
https://books.google[...]
Oxford University Press
2009
[31]
논문
Challenging received wisdoms: Some contributions of the new microscopy to the new animal phylogeny
[32]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[33]
논문
High-resolution vision in pelagic polychaetes
2024-04-08
[34]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[35]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[36]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[37]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[38]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[39]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[40]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[41]
서적
Animal Diversity
Mc Graw Hill
[42]
서적
Invertebrate Zoology
Brooks / Cole
2004
[43]
서적
Invertebrate Zoology
Oxford University Press
[44]
서적
Assembling the tree of life
Oxford University Press
[45]
서적
Invertebrate conservation and agricultural ecosystems
https://books.google[...]
Cambridge University Press
[46]
서적
The worm book
https://books.google[...]
Ten Speed Press
2009-04-02
[47]
서적
Biology and ecology of earthworms
Springer
2009-04-12
[48]
논문
A review of the biology, ecology and potential use of the common ragworm ''Hediste diversicolor''
2002-02
[49]
서적
Practical Fresh Water Fishing
Read Books
2008
[50]
뉴스
Rags to riches
http://www.economist[...]
2009-04-20
[51]
서적
Invertebrate Zoology
Oxford University Press
[52]
논문
Decay and preservation of polychaetes; taphonomic thresholds in soft-bodied organisms
http://paleobiol.geo[...]
[53]
논문
The earliest annelids: Lower Cambrian polychaetes from the Sirius Passet Lagerstätte, Peary Land, North Greenland
http://www.app.pan.p[...]
[54]
웹사이트
A Revised Morphology of Cloudina with Ecological and Phylogenetic Implications
http://ajm.pioneerin[...]
2009-04-12
[55]
논문
Inconsistencies in proposed annelid affinities of early biomineralized organism Cloudina (Ediacaran): structural and ontogenetic evidences
http://paleopolis.re[...]
2012-08-29
[56]
논문
Anatomy and relationships of the Early Cambrian worm ''Myoscolex''
[57]
논문
A critical reappraisal of the fossil record of the bilaterian phyla
2000-05
[58]
논문
Calcareous tubeworms of the Phanerozoic
http://www.eap.ee/pu[...]
[59]
서적
Advances in Regolith
CRC LEME
2009-04-13
[60]
논문
Palaeosols in the upper Narrabeen Group of New South Wales as evidence of Early Triassic palaeoenvironments without exact modern analogues
http://www.uoregon.e[...]
2009-04-13
[61]
논문
A possible annelid from the Trenton Limestone (Ordovician) of Quebec, with a review of fossil oligochaetes and other annulate worms
[62]
논문
Cladistics and polychaetes
[63]
웹사이트
Annelida. Annelida. Segmented worms: bristleworms, ragworms, earthworms, leeches and their allies
http://tolweb.org/An[...]
Tree of Life Project
2009-04-13
[64]
문서
[65]
논문
Molecular evidence that echiurans and pogonophorans are derived annelids
[66]
논문
The new view of animal phylogeny
http://www-fourier.u[...]
2009-04-17
[67]
논문
Annelid phylogeny and the status of Sipuncula and Echiura
2007-04-05
[68]
웹사이트
Reading trees: A quick review
http://evolution.ber[...]
University of California Museum of Paleontology
2009-04-13
[69]
논문
Book Review: Reproductive Biology and Phylogeny of Annelida
[70]
논문
The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm
[71]
논문
Current status of annelid phylogeny
[72]
논문
Combined-data phylogenetics and character evolution of Clitellata (Annelida) using 18S rDNA and morphology
[73]
논문
Phylogeny of basal descendants of cocoon-forming annelids (Clitellata)
https://journals.tub[...]
2012-01-01
[74]
서적
Handbook of Zoology: Annelida
De Gruyter
2019
[75]
논문
Broad phylogenomic sampling improves resolution of the animal tree of life
[76]
논문
Evidence for a clade of nematodes, arthropods and other moulting animals
[77]
논문
Evolution of the bilaterian body plan: What have we learned from annelids?
2000-04
[78]
서적
Environment, Development, and Evolution
MIT Press
2009-07-03
[79]
논문
A Cambrian tommotiid preserving soft tissues reveals the metameric ancestry of lophophorates
2022
[80]
논문
Current status of annelid phylogeny
2016
[81]
논문
Phylogenomic analyses unravel annelid evolution
https://www.nature.c[...]
2011-03
[82]
논문
Current status of annelid phylogeny
http://link.springer[...]
2016-06
[83]
간행물
日本動物学会 2018
[84]
간행물
三浦・白山 2000
[85]
간행물
藤田 2010
[86]
간행물
三浦・白山 2000
[87]
간행물
西川 2000-23
[88]
간행물
西川 2000-22
[89]
간행물
岩波生物学辞典 第5版
2013
[90]
논문
Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa
https://academic.oup[...]
2015-11
[91]
논문
The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm
https://linkinghub.e[...]
2015-08
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com