장새류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
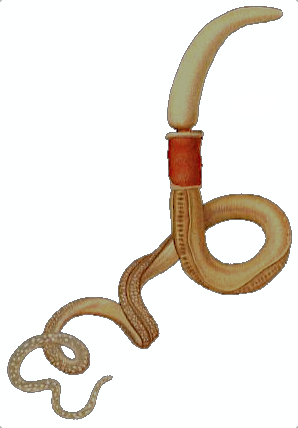
장새류는 몸길이 9~45cm의 해양 무척추동물로, 주둥이, 깃, 몸통의 세 부분으로 구성된다. 피부는 섬모와 점액을 분비하며, 일부 종은 브롬 화합물을 생성하여 포식자로부터 자신을 보호한다. 소화계는 입에서 항문까지 거의 직선으로 이어지며, 순환계는 개방형, 호흡은 아가미를 통해 이루어진다. 신경계는 등쪽과 배쪽 신경삭으로 집중되어 있으며, 굴을 파고 생활하며 유기물을 섭취하거나 물속 유기물을 걸러 먹는다. 암수딴몸으로, 체외 수정을 통해 번식하며, 토르나리아 유생 단계를 거치는 종도 있다. 반삭동물문 장새강에 속하며, 척삭동물과의 유사성 때문에 진화 연구에 중요한 모델 생물로 사용된다. 전 세계적으로 약 70종이 알려져 있으며, 한국에는 7종이 기록되어 있다.
장새류는 몸길이가 대략 9~45cm이며, 가장 큰 종은 1.5m에 달한다. 몸은 도토리 모양의 주둥이, 짧은 깃, 그리고 긴 몸통의 세 부분으로 구성된다. 입은 주둥이 뒤쪽 깃에 위치한다.[16]
장새류는 반삭동물문 내에서 깃벌레강과 자매군을 형성한다.[54][55] 분자 계통학 연구에 따르면, 장새류는 Harrimaniidae, Spengeliidae, Torquaratoridae, Ptychoderidae의 네 과로 나뉜다. 16S + 18S rRNA 서열 데이터와 계통유전학 연구를 기반으로 장새류 내의 관계가 밝혀지고 있다.[54][55]
장새류는 해안선에서 수심 3,050m까지의 해저에 U자형 굴을 파고 서식하며, 굴의 한쪽 구멍에서 주둥이를 내밀고 생활한다. 이들은 일반적으로 느리게 굴을 파고 이동한다.[56]
도토리 벌레는 암수가 따로 있는 자웅이체이며, 일부 종은 무성 생식을 할 수 있다. 한 쌍의 생식선을 가지고 있는데, 생식샘은 인두 가까이에 위치하며 아가미 구멍 근처의 작은 구멍을 통해 생식 세포를 방출한다. 암컷은 젤라틴 형태의 점액 덩어리에 많은 양의 알을 낳으며, 그 후 수컷이 체외 수정을 한다. 이후 물살이 덩어리를 깨고 각각의 알을 흩어지게 한다.[57]
2. 특징
피부는 섬모와 점액 분비선으로 덮여 있으며, 일부 종은 브롬 화합물을 생성하여 박테리아와 포식자로부터 자신을 보호한다. 장새류는 섬모의 움직임과 주둥이의 연동 운동을 이용해 느리게 움직인다.[16]
장새류는 척삭동물과 유사하게 아가미 구멍(새열)을 통해 호흡하며, 순환계는 심장과 신장의 기능을 함께 수행하는 독특한 구조를 가지고 있다.[52][53] 이러한 특징 때문에 고전적인 무척추동물과 척추동물 사이의 연결 고리로 여겨지기도 한다.[52][53] 과거에는 입맹낭이 척삭과 상동이라고 여겨져 원삭동물로 분류되기도 했지만, 현재는 이 견해가 부정되었다. 그러나 아가미 틈새의 존재는 장새류가 척삭동물과 유연 관계가 있음을 보여주는 증거로 여겨진다.[27]
2. 1. 외부 형태
몸은 가늘고 길며, 주둥이는 둥글고 형태 변화가 활발하다. 깃은 짧은 원통형이며, 몸통은 몸 전체 길이의 대부분을 차지한다.[28] 몸통 앞부분에는 새열(아가미 구멍)이 쌍으로 배열되어 있으며, 이 부분을 새역(branchial region)이라고 한다.[29] 생식선이 있는 부분을 생식역(genital region)이라고 하며, 일부 종은 생식돌기나 생식익을 가진다.
대부분의 장새류는 길이가 9cm에서 45cm이며, 가장 큰 종인 Balanoglossus gigas는 1.5m 이상에 달한다. 몸은 도토리 모양의 전구강, 그 뒤에 있는 짧고 살이 많은 칼라, 그리고 길고 웜 모양의 몸통의 세 부분으로 구성된다. 이 생물의 입은 전구강 뒤의 칼라에 위치한다.[16]
피부는 섬모와 점액을 분비하는 선으로 덮여 있다. 일부는 약 냄새를 내는 브롬 화합물을 생성하여 박테리아와 포식자로부터 보호한다. 장새류는 섬모 작용과 전구강의 연동 운동을 사용하여 느리게 움직인다.[16]
미사키깃보시무시는 전체 길이 40cm에 달하는 대형종이지만, 주둥이와 깃의 길이는 각각 1cm 정도이며, 나머지는 체간이다. 이 중 전방의 새역이 약 4cm, 생식역이 6cm, 여기에서는 생식익이 양쪽에 있지만, 본 종에서는 배면을 덮는다. 그 후방에 14cm 정도 간맹낭이 늘어서 있으며, 그 이후는 매끄러운 원통형이 된다. 덧붙여 새역이나 생식역, 간맹낭부 등은 다소 중첩되어 있다.[29]
2. 2. 내부 구조
소화계는 입에서 항문까지 거의 직선으로 연결되며, 분기가 거의 없다.[30] 입 안쪽에는 입맹낭(stomochord)이라는 가느다란 관 모양의 맹관이 주둥이 안으로 뻗어 있다. 이 벽은 비후되어 있으며, 이전에는 척삭동물의 척삭과 상동 기관으로 여겨졌다. 소화관의 깃 뒷부분에는 측면 등쪽에 아가미 구멍이 쌍으로 배열되어 있다. 이 부분에서 소화관은 위쪽 호흡 수로와 배쪽 음식 수로로 나뉜다. 아가미 구멍은 적게는 12개, 많게는 700쌍까지 있다.[31] 소화관에 열리는 안아가미 구멍은 위로 열린 U자형이며, 등쪽에서 혀 모양 돌기가 나와 입 대부분을 덮고 있다. 안쪽에는 아가미실이 있어 바닷물이 소화관에서 이곳으로 들어간 후, 등쪽에 열리는 바깥아가미 구멍을 통해 밖으로 나온다. 그 이후 소화관에는 간맹낭이 다수 있는 종도 있지만, 그 이후로는 곧게 뻗어 있다.
혈관계는 개방혈관계이며, 혈액은 투명하다. 소화관 배면과 복면에는 뚜렷한 혈관이 있는데, 각각 등혈관, 복혈관이라 한다. 등혈관은 앞쪽에서 주둥이 부분으로 들어가 팽창하는데, 이를 중앙 혈동이라 한다. 바로 아래에는 심포가 있는데, 혈관과 연결되지 않지만 움직임을 통해 중앙 혈동 내 혈액을 흐르게 하는 심장과 같은 역할을 한다.
신경계는 복면과 배면을 세로로 지나는 복신경과 배신경을 가진다. 배신경의 깃 부분(깃신경삭)을 제외하면 신경은 표피에서 분리되지 않는다.
배설계는 주둥이에 있는 사구체를 통해 이루어지며, 배출물은 주둥이체강을 통해 몸 밖으로 배출된다.[32]
진체강으로, 3개의 체강을 가진다. 주둥이 내부는 바깥쪽에 근육층이 있고, 안쪽에 주둥이체강이 있다. 이는 단일하며, 기부 배면 쪽에 작은 개구부가 있어 외부와 연결된다. 깃에는 깃체강이 있는데, 중앙에서 칸막이로 나뉘어 좌우 쌍을 이루고, 앞뒤로도 칸막이로 나뉘어 여러 개가 된다. 몸통 부분에서는 소화관 바깥쪽에 몸통체강이 있으며, 역시 정중선에서 좌우로 칸막이로 나뉘어 쌍을 이루고, 일부에서는 복배로도 칸막이로 나뉜다. 골격은 없지만, 주둥이와 깃 접합부에서 주둥이 기부 안쪽에 기저막이 비후된 부분이 있는데, 이를 주둥이골격이라 한다.[27]
2. 3. 소화계
장새류는 모래나 진흙 속 유기물을 섭취하거나, 물속 유기물을 걸러 먹는다. 구강은 관 모양이며, 좁은 게실 또는 구삭이 머리 쪽으로 뻗어 있다. 입은 인두 뒤쪽에서 열리며, 양쪽에 아가미 틈새가 있다. 소화계는 식도와 창자로 구성되며, 위는 없다.[16]
일부 종은 식도 등쪽에 구멍이 있어 음식물에서 나온 물을 짜내어 농축하는 데 도움을 준다. 소화는 창자에서 일어나며, 음식물은 근육 운동이 아닌 섬모에 의해 당겨진다.[16]
2. 4. 순환계
장새류는 개방 순환계를 가지며, 혈액은 조직 혈강을 통해 흐른다. 내장의 위에 있는 장간막의 등쪽 혈관은 심장 역할을 하는 근육 주머니가 들어 있는 돌기 내의 정맥동으로 혈액을 전달한다. 다른 대부분의 동물의 심장과는 달리, 이 구조는 내부가 혈액 시스템과 직접 연결되지 않는 닫힌 유체 채워진 소포이다. 그럼에도 불구하고, 규칙적으로 박동하여 주변의 정맥동을 통해 혈액을 밀어내는 데 도움이 된다.[16]
칼라의 중앙 정맥동에서 혈액은 일련의 복잡한 정맥동과 돌기의 복막 주름으로 흐른다. 이 일련의 구조를 '사구체'라고 하며, 도토리벌레는 별도의 배설계를 가지고 있지 않기 때문에 배설 기능을 가질 수 있다. 돌기에서 혈액은 소화관 아래로 흐르는 단일 혈관으로 흘러 들어가고, 이 혈관에서 더 작은 정맥동이 몸통에 혈액을 공급하고 다시 등쪽 혈관으로 들어간다.[16]
도토리벌레의 혈액은 무색이며 무세포이다.[16]
소화관의 배면과 복면에는 뚜렷한 혈관이 있으며, 각각 등혈관, 복혈관이라고 부른다. 등혈관은 앞쪽에서 吻부(주둥이 부분)로 들어가 거기에서 팽창한 부분을 갖는다. 이를 중앙 혈동이라고 한다. 그 바로 아래에는 심포가 있으며, 이는 혈관과 연락을 갖지 않지만 그 움직임에 의해 중앙 혈동 내의 혈액을 흐르게 하는 심장과 같은 역할을 한다.[32]
2. 5. 호흡계
장새류는 입을 통해 산소가 풍부한 물을 빨아들여 아가미를 통해 호흡한다. 아가미 구멍은 U자형 틈을 통해 인두와 연결되어 있으며, 섬모는 물을 지속적으로 흐르게 한다. 아가미 구멍 주변 조직에는 혈관동이 잘 발달되어 있어 효율적인 가스 교환을 돕는다.[16]
도로리벌레는 성장하면서 새로운 아가미 구멍을 지속적으로 형성하며, 일부 오래된 개체는 양쪽에 100개 이상의 아가미 구멍을 가지기도 한다. 미세한 종인 ''Meioglossus psammophilus''는 단 하나의 아가미 구멍만을 가진다. 일부 종은 아가미에 연골 지지 구조를 가지기도 한다.[19]
2. 6. 신경계
신경총은 피부 아래에 위치하며, 등쪽 및 배쪽 신경삭으로 집중된다. 배쪽 신경삭은 깃까지 뻗어 있지만, 등쪽 신경삭은 주둥이까지 뻗어 있으며, 그 부분에서는 표피로부터 부분적으로 분리되어 있다. 등쪽 신경삭의 이 부분은 종종 속이 비어 있으며, 척추동물의 뇌와 상동성을 가질 수 있다. 갯지렁이에서는 굴을 파고 기어가는 동안 몸의 근육 활동을 조절하는 데 주로 관여하는 것으로 보인다.[16]
갯지렁이는 눈, 귀 또는 기타 특수한 감각 기관이 없지만, 입 앞쪽에 섬모 기관이 있어 여과 섭식과 아마도 미각에 관여하는 것으로 보인다.[16] 그러나 피부 전체에 수많은 신경 종말이 있다.[16]
2. 7. 골격계
도토리벌레는 Y자 모양의 목뼈(누찰골격)를 가지고 있으며, 이는 복부 쪽에 있는 주둥이와 깃 사이에서 시작된다. 목뼈의 뿔 길이는 종에 따라 다르다.[20] 주둥이 기부에는 기저막이 비후된 부분인 吻골격(입골격)이 있다.[27]
2. 8. 척삭동물과의 유사성
도토리벌레는 원시적인 물고기의 아가미와 비슷한 구조를 사용하여 호흡한다. 이러한 아가미 구조는 척삭동물과의 유사성을 보여주는 특징 중 하나이다. 순환계는 심장과 신장의 기능을 함께 수행하는 독특한 구조를 가지고 있다.[52][53] 또한, 일부 종은 척추동물의 항문 후 꼬리와 상동일 수 있는 항문 후 꼬리를 가지고 있다.[52][53]
장새류의 몸은 세 부분으로 나뉘는데, 이는 초기 척삭동물 조상의 해부학적 구조를 반영하는 것으로 여겨진다. 특히, 전두 신경관은 척추동물의 뇌로 발달하는 세 부분으로 나뉘는 구조와 유사하며, 이는 척삭동물 조상의 해부학적 특징이 여전히 남아있음을 시사한다.[52][53]
이러한 특징들 때문에 장새류는 고전적인 무척추동물과 척추동물 사이의 연결 고리로 여겨지기도 한다.[52][53]
한 이론에 따르면, 세 부분으로 구성된 몸체는 모든 후구동물의 초기 공통 조상에서 유래했을 수 있으며, 심지어 후구동물과 전구동물의 공통 조상에서 기원했을 가능성도 제기된다. 연구에 따르면 배아의 유전자 발현은 모든 척추동물의 뇌를 형성하는 세 개의 신호 전달 중심을 공유하지만, 신경계 형성 대신 다른 신체 부위의 발달을 제어한다.[21][22]
과거에는 입맹낭이 척삭과 상동이라고 여겨져 원삭동물로 분류되기도 했지만, 현재는 이 견해가 부정되었다. 그러나 아가미 틈새의 존재는 장새류가 척삭동물과 유연 관계가 있음을 보여주는 증거로 여겨진다.[27]
3. 계통 발생론
{| class="wikitable"
|-
! '''반삭동물'''
|-
|
{| class="wikitable"
|-
| 깃벌레강
|-
! 장새강
|-
|
{| class="wikitable"
|-
| ''Stereobalanus''
|-
|
{| class="wikitable"
|-
| Harrimaniidae
|-
|
{| class="wikitable"
|-
| Spengeliidae
|-
|Torquaratoridae Ptychoderidae
|}
|}
|}
|}
|}
4. 생활 방식
장새류는 먹이를 얻는 방식에 따라 두 가지로 나뉜다.섭식 방법 설명 퇴적물 섭식 지렁이처럼 유기물과 미생물이 포함된 모래나 흙을 삼키고, 썰물 때 뒤쪽 끝을 표면에 내밀어 처리된 퇴적물 덩어리를 배설한다.[56] 현탁물 섭식 물에서 유기물과 미생물의 부유 입자를 모아 섭취한다.[17]
일부 장새류는 브롬을 포함하는 브로모페놀을 생성하여 배출하는데, 이는 항균 작용을 하여 피부 감염 방어와 치유 효과를 가진다.[36][37]
5. 생식과 발생
대부분의 종에서 알은 섬모로 덮인 길쭉한 몸체를 가진 플랑크톤 유충으로 부화한다. 일부 종에서는 성충으로 직접 발전하지만, 다른 종에서는 ''토르나리아'' 유충이라고 불리는 자유 수영 중간 단계가 있다. 이들은 불가사리의 바이핀나리아 유생과 외관이 매우 유사하며, 꼬불꼬불한 섬모의 띠가 몸 주위를 돌아 다니고 있다. 난자 내의 포배의 배아 발달은 또한 극피동물의 배아 발달과 매우 유사하기 때문에, 이것은 두 그룹 사이의 밀접한 계통 발생적인 연관성을 시사한다.[16][57]
며칠 또는 몇 주가 지나면 유충의 중간 부분 주위에 홈이 형성되기 시작하며, 앞쪽 부분은 결국 코가 되고, 나머지는 목과 몸통을 형성한다. 유충은 결국 정착하여 작은 성체로 변하여 굴을 파는 생활 방식을 취한다. ''Saccoglossus_kowalevskii''와 같은 몇몇 종은 플랑크톤 유생 단계조차 없이 작은 성체로 직접 부화한다.[16][57]
장새류는 자웅이체이지만, 일부 종에서는 무성 생식의 한 형태인 분열 외에 작은 싹이 체간부에서 잘려나가는 증식도 알려져 있다.[25] [33]
left
right
토르나리아 유생은 거의 원추형의 외형을 가지며, 측면에 입이 있고, 장관은 중앙에서 아래로 굽어, 하단에 항문이 있다. 입을 둘러싸는 입앞·입뒤 섬모대를 가지며, 또한 하부를 둘러싸는 섬모관이 있다. 상반부 입쪽 내측에는 吻(문, 코)내강이 있으며, 그 개구부는 정단에 있고, 그 근처에 안점이 있다. 변태 시에는 吻내강이라는 부분이 吻(문, 코)으로, 그 후방이 襟(옷깃)으로, 그보다 하방이 체간이 된다. 즉, 토르나리아에 있는 부분은 성체에서는 거의 전단 부분만이 된다. 입앞 및 입뒤 섬모대가 물결치는 듯한 형상이 되어, 다수의 돌출부를 생기는 것도 있으며, 이를 유촉수 토르나리아라고 부른다.[34]
토르나리아 단계에서 앞·가운데·뒤의 3개의 체강이 형성되지만, 그 형성 과정은 불명확한 부분이 많다.
6. 주요 종
7. 인간과의 관계
장새류는 특별한 이용 가치는 없지만, 진화생물학, 특히 신구동물과 척삭동물의 진화 및 기원 연구에 중요한 모델 생물로 여겨진다.[26] 성체는 좌우대칭 구조와 새열을 가지고 있어 척삭동물과의 계통 관계가 논의되며, 과거에는 척삭이 있다고 판단되어 멍게 등과 함께 원삭동물에 포함되기도 했다. 유생은 토르나리아라고 불리며, 극피동물에서도 유사한 유생이 발견된다. 또한, 연체동물 등의 트로코포라와 기본 구성이 유사하여 계통 연구에서 비교 대상이 된다.
일본어 이름은 주둥이 부분을 불교 사찰 지붕이나 난간 끝 장식인 의보주(擬宝珠, 기보시)에 비유하여 붙여졌으며, 영어 이름 "Acorn worm"은 주둥이를 도토리에 비유하여 붙여졌다.[27]
참조
[1]
논문
An early Cambrian pelago-benthic acorn worm and the origin of the hemichordate larva
[2]
간행물
Enteropneusta
2017-11-20
[3]
논문
Evolution of the chordate body plan: New insights from phylogenetic analyses of deuterostome phyla
[4]
기타
Biogeography and adaptations of torquaratorid acorn worms (Hemichordata: Enteropneusta) including two new species from the Canadian Arctic - Research Proposal - Papyrus - Université de Montréal
https://papyrus.bib.[...]
[5]
논문
Hemichordate genomes and deuterostome origins
2015-11
[6]
논문
Enteropneust production of spiral fecal trails on the deep-sea floor observed with time-lapse photography
2005-07
[7]
논문
'Lophenteropneust' hypothesis refuted by collection and photos of new deep-sea hemichordates
[8]
기타
A new deep-sea species of epibenthic acorn worm (Hemichordata, Enteropneusta)
[9]
기타
Diversication of acorn worms (Hemichordata, Enteropneusta) revealed in the deep sea
[10]
기타
Observations on torquaratorid acorn worms (Hemichordata, Enteropneusta) from the North Atlantic with descriptions of a new genus and three new species
[11]
기타
The secret to an Oesia life: Prehistoric worm built tube-like 'houses' on sea floor
https://phys.org/new[...]
[12]
기타
Encyclopedia of Paleontology
https://books.google[...]
[13]
논문
An Anatomical Description of a Miniaturized Acorn Worm (Hemichordata, Enteropneusta) with Asexual Reproduction by Paratomy
2012-11-07
[14]
서적
Deuterostomians, Cyclostomes, and Fishes
https://books.google[...]
Elsevier
[15]
서적
Balanoglossus
[16]
서적
Invertebrate Zoology
Holt-Saunders International
[17]
논문
Particle retention and flow in the pharynx of the enteropneust worm Harrimania planktophilus: The filter-feeding pharynx may have evolved before the chordates
[18]
논문
Effect of flow and sediment transport on feeding rate of the surface-deposit feeder Saccoglossus kowalevskii
https://www.int-res.[...]
1996
[19]
기타
Invertebrate Zoology: A Tree of Life Approach
https://books.google[...]
[20]
논문
A phylogeny of the hemichordates based on morphological characters
https://cdnsciencepu[...]
2011-02-15
[21]
웹사이트
Secondary organizers of the early brain and the location of the meso-diencephalic dopaminergic precursor cells
http://discovery.lif[...]
2014-03-10
[22]
웹사이트
The Secret Origin of the Vertebrate Brain
http://sciencelife.u[...]
ScienceLife
2012-03-15
[23]
논문
The Global Diversity of Hemichordata
2016-10-04
[24]
논문
Mitogenomics Reveals a Novel Genetic Code in Hemichordata
2019-01-01
[25]
논문
Morphological characterization of the asexual reproduction in the acorn worm Balanoglossus simodensis: Asexual reproduction in B. simodensis
2010-09
[26]
서적
「研究者が教える動物飼育 第3巻 -ウニ,ナマコから脊椎動物へ」
[27]
기타
西村編著(1995),p.494
[28]
기타
以下、主として西村編著(1995),p.494-495
[29]
기타
岡田他(1961),p.100
[30]
기타
이하、主として岡田他(1961),p.100、ただし用語は岩槻、馬渡編著(2000)などに合わせてある
[31]
기타
西村編著(1995),p.495
[32]
기타
岩槻・馬渡編著(2000),p.254
[33]
기타
ここまで西村編著(1995),p.495
[34]
기타
この項は冨山他(1958),p.391
[35]
기타
Holland et al.(2012)
[36]
간행물
国際化学物質簡潔評価文書 2,4,6-トリブロモフェノールや他の単純臭素化フェノール
https://www.nihs.go.[...]
国立医薬品食品衛生研究所
[37]
Wikipedia
Acorn_worm
[38]
서적
岡田他(1961),p.8
[39]
서적
岩槻・馬渡監修(2000),p.23
[40]
서적
西村編著(1995),p.495-498
[41]
웹인용
Enteropneusta
https://www.marinesp[...]
World Register of Marine Species
2017-11-20
[42]
웹인용
Evolution of the chordate body plan: New insights from phylogenetic analyses of deuterostome phyla
https://www.ncbi.nlm[...]
Proceedings of the National Academy of Sciences of the United States of America.
2000
[43]
웹인용
Biogeography and adaptations of torquaratorid acorn worms (Hemichordata:Enteropneusta) including two new species from the Canadian Arctic -Researcher Proposal - Papyrus- Universite de Montreal
https://papyrus.bib.[...]
[44]
웹인용
Hemichordate genomes and deuterostome origins
https://www.nature.c[...]
[45]
웹인용
Enteropneust production of spiral fecal trails on the deep-sea floor observed with time-lapse photography
null
2005-07
[46]
웹인용
"'Lophenteropneust' hypothesis refuted by collection and photos of new deep-sea hemichordates"
null
2005
[47]
웹인용
null
null
A new deep-sea species of epibenthic acorn worm (Hemichordata, Enteropneusta)
2009
[48]
웹인용
null
null
Diversication of acorn worms (Hemichordata, Enteropneusta) revealed in the deep sea
2012
[49]
웹인용
null
null
Observations on torquaratorid acorn worms (Hemichordata, Enteropneusta) from the North Atlantic with descriptions of a new genus and three new species
2012
[50]
웹인용
The secret to an Oesia life: Prehistoric worm built tube-like 'houses' on sea floor
https://phys.org/new[...]
[51]
웹인용
null
https://books.google[...]
2014
[52]
저널
Secondary organizers of the early brain and the location of the meso-diencephalic dopaminergic precursor cells Archived 2014-03-10 at the Wayback Machine Retrieved March 10, 2014
https://en.wikipedia[...]
2022-01-03
[53]
저널
Rob Mitchum (March 15, 2012). "The Secret Origin of the Vertebrate Brain". ScienceLife. Retrieved February 18, 2014.
https://en.wikipedia[...]
2022-01-03
[54]
저널
Tassia, Michael G.; Cannon, Johanna T.; Konikoff, Charlotte E.; Shenkar, Noa; Halanych, Kenneth M.; Swalla, Billie J. (2016-10-04). "The Global Diversity of Hemichordata". PLOS ONE. 11 (10): e0162564. Bibcode:2016PLoSO..1162564T. doi:10.1371/journal.pone.0162564. PMC 5049775. PMID 27701429.
https://en.wikipedia[...]
2022-01-03
[55]
저널
Halanych, Kenneth M.; Bernt, Matthias; Cannon, Johanna T.; Tassia, Michael G.; Kocot, Kevin M.; Li, Yuanning (2019-01-01). "Mitogenomics Reveals a Novel Genetic Code in Hemichordata". Genome Biology and Evolution. 11 (1): 29–40. doi:10.1093/gbe/evy254. PMC 6319601. PMID 30476024.
https://en.wikipedia[...]
2022-01-03
[56]
저널
Cameron, C. (2002). "Particle retention and flow in the pharynx of the enteropneust worm Harrimania planktophilus: The filter-feeding pharynx may have evolved before the chordates". The Biological Bulletin. 202 (2): 192–200. doi:10.2307/1543655. JSTOR 1543655. PMID 11971814. S2CID 10556637.
https://en.wikipedia[...]
2022-01-03
[57]
저널
Barnes, Robert D. (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 1018–1026. ISBN 978-0-03-056747-6.
https://en.wikipedia[...]
2022-01-03
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com