코리스토데라
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
코리스토데라는 파충류의 한 분류군으로, 쥐라기 중기부터 마이오세까지 생존했다. 코리스토데라는 머리뼈 구조, 특히 전전두골의 특징적인 배열과 측두린의 형태, 그리고 얕은 홈에 이빨 뿌리가 고정되는 아조생치 형태의 이빨을 가진다. 크기는 30cm에서 5m까지 다양하며, 신코리스토데라와 비신코리스토데라로 나뉜다. 신코리스토데라는 악어와 유사한 형태를 보이며, 비신코리스토데라는 도마뱀과 유사하거나 긴 목을 가진 형태를 보인다. 코리스토데라는 주로 민물에 서식하며, 다양한 생태적 특징과 번식 방식을 보여준다. 화석은 북아메리카, 유럽, 아시아 등 북반구에서 발견되며, 한국에서도 생흔 화석이 발견되었다. 코리스토데라의 계통 발생은 불확실하지만, 신디압시드의 일원으로 분류되며, 다른 파충류와의 관계에 대한 연구가 진행 중이다.
더 읽어볼만한 페이지
- 코리스토데라 - 캄프소사우루스
캄프소사우루스는 백악기 후기부터 팔레오세에 북아메리카와 유럽에 서식한 코리스토데라목 파충류로, 긴 주둥이와 유선형 몸체를 가진 수생 생물이며 1877년 코프에 의해 발견되었고 7종이 알려져 있다. - 에드워드 드링커 코프가 명명한 분류군 - 수염고래
수염고래는 이빨 대신 케라틴 수염판을 가진 고래류 아목으로, 플랑크톤 등을 걸러 먹고, 저주파음을 내며 장거리 회유를 하며 멸종 위기에 처해 보호받고 있다. - 에드워드 드링커 코프가 명명한 분류군 - 곡경아목
곡경아목은 머리를 옆으로 구부려 껍질 속으로 넣는 특징을 가지며, 잠경아목과 구분되고, 경추 구조, 두개골 절흔, 껍질의 각질판 배열 등에서도 차이를 보이며, 남미늪거북과를 포함한 일부 종은 흡입 섭식을 하고, 주로 강, 호수, 늪 등에 서식하며, 남아메리카, 아프리카, 오스트레일리아 등지에 분포한다.
| 코리스토데라 - [생물]에 관한 문서 | |
|---|---|
| 코리스토데라목 정보 | |
 | |
 | |
| 학명 | Choristodera |
| 명명자 | 코프, 1876년 |
| 화석 범위 | 쥐라기 중기 – 마이오세, 잠재적 트라이아스기 기록 |
| 하위 그룹 | |
| 멸종 | Coeruleodraco Cteniogenys Heishanosaurus Irenosaurus "Allochoristodera" Dong et al., 2020 Lazarussuchus Monjurosuchus Philydrosaurus Hyphalosaurus Khurendukhosaurus Shokawa 네오코리스토데라 Evans & Hecht, 1993 Champsosaurus Kosmodraco Ikechosaurus Liaoxisaurus Mengshanosaurus Simoedosaurus Tchoiria |
2. 특징
코리스토데라는 그 크기가 매우 다양하다. 가장 작은 속인 ''크테니오게니스''와 ''라자루스수쿠스''는 몸길이가 약 30cm에 불과했지만, 가장 큰 코리스토데라인 ''코스모드라코 다코텐시스''는 약 5m에 달했다.[5][1] ''샴포사우루스''와 같은 신코리스토데라는 현대의 크로커다일류, 특히 가비알과 유사하게 생겼으며, 작고 날카로운 원뿔형 이빨이 있는 길고 얇은 주둥이를 가졌다. 반면, ''쇼카와'', ''후렌두코사우루스'', ''히팔로사우루스''와 같이 "비신코리스토데라"로 통칭되는 코리스토데라들은 대부분 작은 도마뱀과 같은 형태를 보이지만, 긴 플레시오사우루스와 같은 목을 가진 경우도 있었다. "비신코리스토데라" 그룹은 분류학적 무리 (공통 조상의 모든 후손을 포함하지 않음)이며, 도마뱀과 같은 몸 형태는 그룹의 조상 형태를 나타낸다.[5]
대부분의 코리스토데라는 다소 단순하고 분화되지 않은 동형치를 가졌으며, 치아 에나멜의 줄무늬가 치관을 덮고 있지만 치아 기저부는 덮지 않았다. 신코리스토데라는 줄무늬 에나멜로 완전히 덮인 치아를 가졌으며, 치아 기저부에 에나멜이 접혀 들어가고, 설측으로 압축되고 갈고리 모양을 하고 있다. 예외적으로 이케코사우루스는 에나멜 접힘이 시작되는 것을 제외하면 여전히 다소 단순한 치아를 가졌다.[12]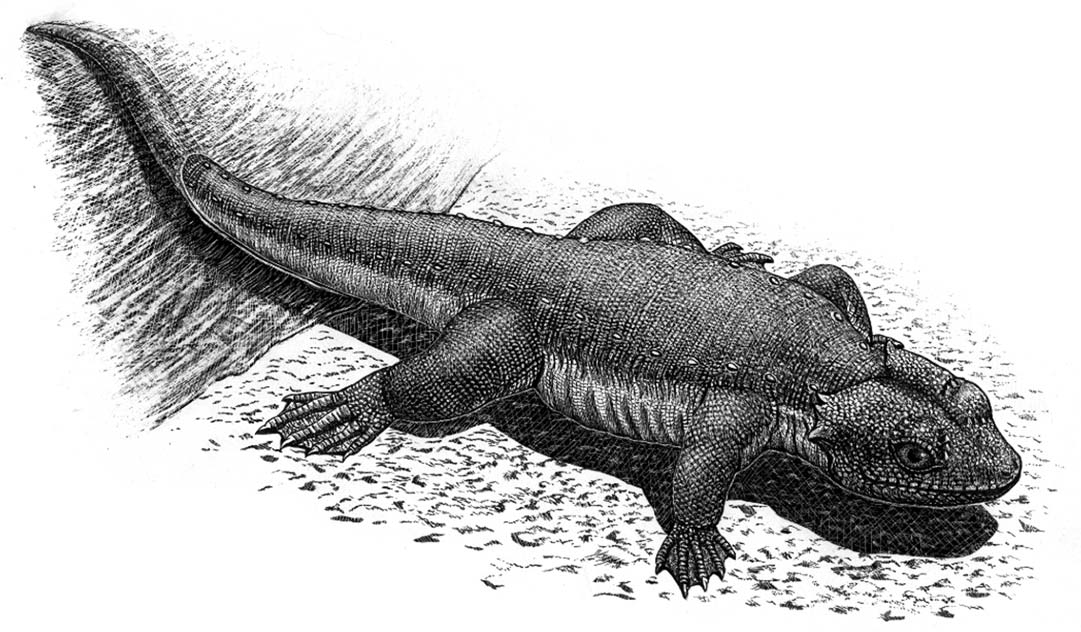

매우 잘 보존된 ''몬주로수쿠스'' 표본에서는 주름진 피부가 발견되었는데, 이는 살아있을 때 피부가 얇고 부드러웠을 가능성을 보여준다. 보존된 비늘은 작고 겹쳐져 있으며, 몸통의 아랫부분보다 등쪽에서 더 작다. 두 줄의 더 큰 타원형 비늘이 몸의 등쪽(상부 중앙선)을 따라 뻗어 있다. 이 화석에서는 물갈퀴 발도 발견되었다.[13]
2. 1. 두개골

코리스토데라의 머리뼈를 보면, 보통 파충류의 머리 양 옆에 있는 전전두골(prefrontal)이 머리 중앙에 모여 전두골(frontal)과 비골(nasal) 사이를 갈라놓는다. 두정골(parietal)에는 홈이 없다. 측두린(squamosal)이 후두돌기(occipital condyle)보다 뒤쪽으로 뻗어 나가, 머리뼈의 뒤쪽이 대개 하트 모양처럼 보인다. 이빨은 턱에 얕은 홈이 나 거기에 이빨 뿌리가 박혀 고정되는 아조생치(subthecodont) 형태이다.[46]
머리에는 코리스토데라 고유의 뼈(choristoderan neomorphic bone)도 발견된다. 이는 코리스토데라 외의 동물에는 없는 뼈로, 두정골 옆에 양쪽으로 하나씩 있다. 고유 뼈는 모든 네오코리스토데라에서 발견되었고, 원시적인 코리스토데라인 코에룰레오드라코(''Coeruleodraco'') 등에서도 발견되어 코리스토데라 모두의 보편적인 특징일 가능성이 있다. 그러나 코리스토데라 고유의 뼈에 대한 연구가 부족하여 이를 보편적 특징으로 정의하기에는 이르다.[47]
마츠모토와 동료들(2019)에 따르면, 코리스토데라는 9개의 공유파생형질(그룹의 특징적인 공유 형질)로 묶인다. 여기에는 두개골의 길쭉한 전두정골이 중간에서 만나 비골을 전두골과 분리시키는 것, 상악골의 등쪽 플랜지가 내측으로 굽어지는 것(몸의 중앙선 방향), 송과공의 부재, 인두골이 후두융기 뒤쪽으로 확장된 것, 원뿔형이고 치아가 반치아 (얕은 소켓에 위치)인 것 등이 포함된다.[7] 알려진 모든 코리스토데라는 다른 파충류에서는 발견되지 않는 새로운 두개골 뼈, 즉 "신생골" 또는 신생 형태를 가지고 있거나, 이를 가지고 있을 것으로 추정되며, 이는 피부두개골의 구성 요소이다.[9][8] 조상 대대로 코리스토데라의 두개골은 길쭉한 위쪽 및 아래쪽 측두창(눈구멍 뒤 두개골의 구멍)을 가지고 있었으며, 이는 신코리스토데라에서 매우 확장되었으며, 특히 ''Champsosaurus''에서 두개골이 위에서 볼 때 심장 모양을 띠게 했다.[9]
2. 2. 치아
대부분의 코리스토데라는 다소 단순하고 분화되지 않은 동형치를 가지고 있으며, 치아 에나멜의 줄무늬가 치관을 덮고 있지만 치아 기저부는 덮지 않는다. 신코리스토데라는 줄무늬 에나멜로 완전히 덮인 치아를 가지고 있으며, 치아 기저부에 에나멜이 접혀 들어가고, 설측으로 압축되고 갈고리 모양을 하고 있는데, 예외적으로 이케코사우루스는 에나멜 접힘이 시작되는 것을 제외하면 여전히 다소 단순한 치아를 가지고 있다.[12] 치아의 착생은 반부착성으로, 치아 기저부의 설면(혀를 향하는 치아 측면)의 구멍이 침식되면서 치아가 교체된다. 신코리스토데라 사이에는 약간의 치아 분화가 있는데, 앞쪽 치아가 뒤쪽 치아보다 더 날카롭고 가늘다. 코리스토데라는 구개 치아(구개골에 존재하는 치아)를 유지한다. 구개 치아가 감소하거나 완전히 사라지는 대부분의 이궁류와는 달리, 코리스토데라의 구개 치아는 구강 내에서 음식 조작을 나타낼 정도로 광범위하게 발달되어 있으며, 아마도 혀와 함께 사용되었을 것이다. 대부분의 코리스토데라에서 구개 치아의 세로 열은 익상골, 구개골, 서골에 존재하며, 익상골 플랜지에도 한 줄이 있다. 일부 신코리스토데라에서는 구개 치아 열이 융기된 플랫폼 위에 치아 배터리로 변형된다. 구개 치아의 형태는 비신코리스토데라의 가장자리 치아의 형태와 동일하며, 구개 치아의 교체는 가장자리 치아의 교체와 거의 동일하다.[12]2. 3. 기타 골격
코리스토데라의 머리뼈는 전전두골(prefrontal)이 머리 중앙에 모여 전두골(frontal)과 비골(nasal)을 분리시키는 특징이 있다. 두정골(parietal)에는 홈이 없으며, 측두린(squamosal)은 후두돌기(occipital condyle)보다 뒤쪽으로 뻗어 머리뼈 뒤쪽이 하트 모양을 띤다. 이빨은 턱에 얕은 홈에 뿌리가 박혀 고정되는 아조생치(subthecodont) 형태이다.[46]코리스토데라만의 고유한 뼈(choristoderan neomorphic bone)가 발견되기도 하는데, 이는 두정골 옆에 양쪽으로 하나씩 나 있다. 이 고유 뼈는 모든 네오코리스토데라와 일부 원시 코리스토데라에서 발견되어 보편적인 특징일 가능성이 있지만, 추가 연구가 필요하다.[47]
마츠모토와 동료들(2019)의 연구에 따르면, 코리스토데라는 9개의 공유파생형질로 묶인다. 해당 형질은 다음과 같다.
- 두개골의 길쭉한 전두정골이 중간에서 만나 비골을 전두골과 분리
- 상악골의 등쪽 플랜지가 내측으로 굽어짐
- 송과공 부재
- 인두골이 후두융기 뒤쪽으로 확장
- 원뿔형이고 치아가 반치아(얕은 소켓에 위치)
- 하악골의 치골은 가늘고 뼈의 입술 표면을 따라 길쭉한 홈이 있음
- 추가적인 천골 존재
- 척추에 확장된 "가시 테이블" 존재
- 척추 중심의 양쪽 끝 표면이 평평(양평형)
2. 4. 피부
매우 잘 보존된 ''몬주로수쿠스''(Monjurosuchus) 표본에서는 주름진 피부가 발견되었는데, 이는 살아있을 때 피부가 얇고 부드러웠을 가능성을 보여준다. 보존된 비늘은 작고 겹쳐져 있으며, 몸통의 아랫부분보다 등쪽에서 더 작다. 두 줄의 더 큰 타원형 비늘이 몸의 등쪽(상부 중앙선)을 따라 뻗어 있다. 이 화석에서는 물갈퀴 발도 발견되었다.[13] ''히팔로사우루스''(Hyphalosaurus)는 몸의 위치에 따라 다양한 모양의 비늘로 덮여 있었으며, 적어도 한 줄, 어쩌면 여러 줄의 큰 타원형 비늘이 몸통과 꼬리 측면을 따라 이어졌다. 발에는 물갈퀴의 흔적이 있고, 꼬리 위아래에는 추가적인 조직이 있어서 ''히팔로사우루스''(Hyphalosaurus)가 물속에서 움직이는 데 사용되었을 것으로 보인다.[10][14]
''참포사우루스''(Champsosaurus)의 피부 흔적도 보고되었는데, 작고(0.6mm-0.1mm) 오돌토돌하고 마름모꼴인 비늘로 구성되어 있으며, 가장 큰 비늘은 몸의 측면에 있고 등쪽으로 갈수록 크기가 작아지며, 골판은 없었다.[15] ''라자루수쿠스''(Lazarussuchus)의 Menat 표본은 연조직의 잔해를 일부 보존하고 있지만 비늘은 없었고, 뒷발에 물갈퀴가 없었으며, 꼬리 척추뼈 위에는 물결 모양의 가장자리가 있는 어두운 반점이 있어 투아타라, 도마뱀, 악어와 같은 일부 현존 파충류에서 발견되는 것과 유사한 볏을 보여준다.[43]
3. 화석 기록
코리스토데라 중 가장 오래된 화석은 영국의 쥐라기 중기 지층에서 발견된 크테니오게니스(''Cteniogenys'')이다. 영국의 트라이아스기 후기 지층에서 발견된 파키스트로페우스(''Pachystropheus'')는 코리스토데라에 속하는지 불확실하다. 가장 최근 화석은 독일의 마이오세 후반 지층에서 발견된 라자루스수쿠스(''Lazarussuchus'')이다[48].
코리스토데라는 북아메리카, 유럽, 아시아 등 북반구에서 화석이 발견된다. 대한민국에서는 아직 체화석이 보고되지 않았으나, 울산 반구대에 있는 대구층에서 발자국 생흔화석이 발견되었다. 이 생흔화석에는 노바페스 울산엔시스(''Novapes ulsanensis'')라는 이름이 붙여졌다[49].
4. 생태
코리스토데라는 주로 민물에 살았던 수생 파충류이다. 캄프소사우루스와 같은 네오코리스토데라는 악어와 비슷하게 진화하여 매복하는 포식자였을 것이고, 히팔로사우루스나 쇼카와는 목이 길게 진화하였다.
코리스토데라는 담수 퇴적층에서만 발견되며, 종종 거북, 어류, 개구리, 도롱뇽 및 악어형류와 연관되어 있다. 코리스토데라는 거의 전적으로 따뜻한 온대 기후에서 발견된 것으로 보이며, 신코리스토데라의 분포는 극심한 온난기였던 백악기 후기(약 8,900만~8,300만 년 전)의 코냐크절-상토절 동안 캐나다 북부의 북극까지 확장되었다. 코리스토데라와 악어형류 사이의 형태학적 유사성 때문에, 이들이 경쟁 관계에 있었을 것으로 추정되어 왔다. 그러나 "신코리스토데라가 아닌" 종들은 성체 수생 악어형류보다 작았고 다른 분류군과의 경쟁이 더 컸을 가능성이 있다. 좀 더 악어와 비슷한 신코리스토데라의 경우, 생태적 지위 분화가 있었던 것으로 보이며, 가비알과 같은 신코리스토데란은 뭉툭한 주둥이의 악어형류와 함께 나타났지만 좁은 주둥이 형태와는 함께 나타나지 않았다.
신코리스토데라는 어식 동물이었을 것으로 추정된다.[15] 특히, ''캄포사우루스''는 현대의 가비알처럼 무리를 지어 다니는 물고기 중에서 머리를 옆으로 휘저어 개별 물고기를 잡았을 것으로 생각되는 반면, ''시모에도사우루스''는 수생 및 육상 먹이를 모두 섭취할 수 있어 보다 잡식성이었을 것으로 생각된다.[16] ''크테니오게니스''와 ''라자루스수쿠스''는 무척추동물을 먹었을 것으로 추정된다.[6] ''몬주로수쿠스'' 표본의 보존된 내장 내용물에서는 절지동물 큐티클 조각이 발견되었다.[13] 또 다른 ''몬주로수쿠스'' 표본에서는 복강 내에 7마리의 어린 개체의 두개골이 보존된 채 발견되었다. 이것은 식인 풍습의 증거로 제시되었다.[17] 그러나, 이 주장은 다른 저자들에 의해 비판받았으며, 그들은 이것이 후기 단계의 배아를 나타낼 가능성이 더 높다고 주장한다.[18] ''히팔로사우루스'' 표본에서는 복강에 작은 갈비뼈가 발견되어, 적어도 가끔은 척추동물을 먹었다는 것을 시사한다.[10]
5. 분류 및 계통 발생
코리스토데라는 모두 신디압시드 파충류의 일종으로 인정받고 있다. 하지만 이들은 원시적인 특징과 파생된 특징이 섞여 있고, 다른 파충류와 분리된 이후 화석 기록이 부족한 긴 유령 계통을 가지고 있어, 정확히 어떤 종류의 파충류에 속하는지는 불분명하다.[28]
초기에는 두개류에 속하는 것으로 여겨졌으나, 이후 목뼈의 모양 때문에 도마뱀류에 속할 수 있다는 주장이 제기되었다. 루이 돌로는 1891년에 다시 코리스토데라를 두개류로 분류했지만, 1893년에는 파레이아사우루스와 가까운 관계라고 제안했다. 알프레드 로머는 1956년과 1968년 논문에서 코리스토데라를 "에오수키아"라는 분류군에 포함시켰는데, 이 분류는 널리 받아들여졌다. 그러나 1980년대에 컴퓨터를 이용한 분지학 연구 결과 "에오수키아"가 단일 계통군이 아니라는 것이 밝혀지면서 코리스토데라의 분류는 다시 불확실해졌다.[29]
이후 연구에서는 아르코사우로모르파, 렙토사우로모르파 등 다양한 분류군에 속할 수 있다는 주장이 제기되었다. 2016년 마르틴 에스쿠라의 연구에서는 코리스토데라가 사우리아에 속하며, 렙토사우로모르파 및 아르코사우로모르파와 함께 분지적 다분기를 이루는 것으로 나타났다. 또한, 코리스토데라가 이 두 그룹보다 먼저 분기했을 가능성도 제기되었다.[28] 코리스토데라가 기저 아르코사우로모르파에 속한다는 주장은 배아의 뼈가 형성되는 순서에 의해 뒷받침된다.[18]
5. 1. 내부 계통
과거에는 코리스토데라의 내부 계통 발생이 불분명했으며, 신코리스토데라가 지지도가 높은 분지군으로 회복되었지만, "비신코리스토데라"의 관계는 제대로 해결되지 않았다.[7] 그러나 2010년대에 아시아의 초기 백악기에서 발견된 "비신코리스토데라" (단, ''Heishanosaurus'')와 유럽의 신생대에서 발견된 ''Lazarussuchus''는 (약한 지지도로) 2020년 동과 동료들에 의해 "Allochoristoderes"라는 비공식적인 명칭으로 불린 단일 계통군에 속하는 것으로 밝혀졌으며, 공유된 특징으로 하부 측두창이 완전히 닫혀 있고, 유럽과 북미의 중후기 쥐라기에서 발견된 ''Cteniogenys''가 일관적으로 가장 기저적인 코리스토데라로 회복되었다.[25] 목이 긴 "비신코리스토데라"인 ''Shokawa''와 ''Hyphalosaurus''는 종종 2005년 가오와 폭스에 의해 Hyphalosauridae로 명명된 분지군으로 회복되었다.[26] 이전에 조각난 형태로만 알려졌던 ''Khurendukhosaurus''의 더 완전한 화석이 발견되면서, 이 종 역시 긴 목을 가지고 있으며 해당 분지군의 일부로 회복되었다는 것을 보여준다.[27]동과 동료들 (2020)의 분석에 따른 계통 발생은 다음과 같다.[25]
{| class="wikitable"
|-
! '''코리스토데라'''
|-
|
{| class="wikitable"
|-
| ''Cteniogenys'' sp.
|-
|
{| class="wikitable"
|-
| ''Heishanosaurus pygmaeus''
|-
| ''Coeruleodraco jurassicus''
|-
|
{| class="wikitable"
|-
! 신코리스토데라
|-
|
{| class="wikitable"
|-
| ''Ikechosaurus pijiagouensis''
|-
| ''Ikechosaurus sunailinae''
|-
| ''Tchoiria namsari''
|-
| ''Tchoiria klauseni''
|-
! ''Champsosaurus''
|-
|
| C. gigas |
| C. albertensis |
|-
! ''Simoedosaurus''
|-
|
| S. lemoinei |
| S. dakotensis |
|}
|-
! "Allochoristodera"
|-
|
{| class="wikitable"
|-
| ''Monjurosuchus splendens''
|-
| ''Philydrosaurus proseilus''
|-
|
{| class="wikitable"
|-
! ''Lazarussuchus''
|-
|
| L. inexpectatus |
| Lazarussuchus sp. |
| L. dvoraki |
|-
|
{| class="wikitable"
|-
| ''Khurendukhosaurus orlovi''
|-
| ''Hyphalosaurus'' sp.
|-
|
| Hyphalosaurus lingyuanensis |
| Shokawa ikoi |
|}
|}
|}
|}
|}
|}
5. 2. 다른 파충류와의 관계
코리스토데라는 모두 신디압시드의 일원으로 인정받고 있지만, 기원적 특징과 파생된 특징이 혼합되어 있고, 다른 파충류와 분리된 이후의 긴 유령 계통(화석 기록 부재)으로 인해 계통 내 정확한 위치는 불확실하다.[28] 초기에는 두개류에 속했지만, 코프는 경추의 모양 때문에 나중에 도마뱀류에 속할 수 있다고 제안했다. 루이 돌로는 1891년에 코리스토데라를 두개류로 되돌렸지만, 1893년에는 ''파레이아사우루스''와 밀접한 관계가 있다고 제안했다. 알프레드 로머는 1956년과 1968년에 발표한 논문에서 코리스토데라를 분기도 또는 다계통군인 "에오수키아"에 포함시키면서, "기본적인 에오수키아 계통의 분파"라고 묘사했는데, 이 분류는 널리 받아들여졌다. 그러나 1980년대에 컴퓨터 기반 분지학을 사용하면서 "에오수키아"가 단일 계통군이 아니라는 것이 밝혀지면서 코리스토데라의 분류는 다시 불확실해졌다.[29] 이후 연구에서는 아르코사우로모르파, 렙토사우로모르파 또는 디압시드의 구성원 ''incertae sedis''로 위치를 제안했다. 마르틴 에스쿠라가 2016년에 수행한 신디압시드 관계 분석에서는 렙토사우로모르파 및 아르코사우로모르파와 함께 분지적 다분기에서 고급 신디압시드 그룹인 사우리아의 구성원으로 밝혀졌으며, 두 그룹 중 가장 먼저 분기된 구성원일 가능성도 있다.[28] 기저 아르코사우로모르파로서의 위치는 그들의 배아의 골화 순서에 의해 뒷받침된다.[18]6. 진화 역사
코리스토데라는 1876년 에드워드 드링커 코프가 린코세팔리아의 하위 목으로 만들었으며, 코프는 같은 논문에서 몬태나의 후기 백악기 지층에서 ''참포사우루스''를 기술했다.[3][4] 1877년 폴 제르베는 프랑스 랭스 근처 세르네-레-랭스의 상부 시원세 퇴적물에서 ''시모에도사우루스''를 기술했다. 이들은 20세기 후반까지 1세기 이상 코리스토데라로 인정받는 유일한 종이었다.[5] 1970년대 후반부터 소련-몽골 팀이 몽골의 하부 백악기 퇴적물에서 추가 분류군을 기술했다. 1989년부터 1991년까지 수잔 E. 에반스는 영국의 중기 쥐라기에서 ''크테니오게니스''의 새로운 표본을 기술했다. 찰스 W. 길모어가 1928년 미국의 후기 쥐라기에서 처음 기술한 이 속은 이전에는 수수께끼로 여겨졌으나, 이 연구를 통해 악어와 같은 형태와는 다른 작은 도마뱀과 같은 코리스토데라임이 밝혀졌다.[6]
코리스토데라는 2억 5천만 년 전 페름기 말 이전에 다른 모든 파충류 그룹에서 분기되었음이 분명하며, 이는 그들의 원시적인 계통 발생 위치에 근거한다.[5] 2015년 라이너 R. 쇼흐는 독일 남부의 중생대 트라이아스기 (라디니안) 하부 케이퍼에서 두개골과 몸통 골격을 모두 갖춘 새로운 소형(~20cm 길이) 이궁류를 보고했는데, 그는 이것이 가장 오래된 코리스토데레를 나타낸다고 주장했다.[30] 영국의 후기 트라이아스기 (레티안) ''파키스트로페우스''는 역사적으로 코리스토데레로 추정되었지만,[31] 탈라토사우루스류의 일원임이 밝혀졌다.[32] 가장 오래된 확실한 코리스토데란은 작은 도마뱀과 같은 ''크테니오게니스''로, 영국의 후기 쥐라기 중기 (바토니안 ~168Ma~166Ma) 포레스트 마블 및 킬말루아그 지층에서 가장 오래된 유해가 발견되었다. 포르투갈의 후기 쥐라기 알코바사 지층과 미국의 모리슨 지층, 키르기스스탄의 후기 쥐라기 중기 (칼로비안) 발라반사이 지층,[5][33] 서시베리아의 바토니안 이타트 지층,[34] 모로코의 바토니안 시대 아누알 지층에서도 유사한 유해가 발견되었다.[35]
코리스토데라는 백악기 초기 아시아에서 주요한 진화적 방사를 겪었고, 이는 코리스토데란 다양성의 정점을 나타낸다. 수생 신수키아 악어류가 지역적으로 부재한 상태에서 진화한 것으로 보이는 가비알과 유사한 네오코리스토데라의 첫 번째 기록도 이때 나타난다.[5] 불확실한 코리스토데레의 부분적인 대퇴골이 북아메리카 세다 마운틴 지층의 옐로우 캣 멤버에서 발견되었다.[36] 영국 베리아세스기 퍼벡 그룹과 스페인 바레미안 라 후에르기나 지층에서는 발견되지 않지만,[5] 프랑스 베리아세스기 앙제악-샤랑트 뼈대에서 작은 ''크테니오게니스''와 유사한 분류군이 기록되었다.[37] 후기 백악기 (캄파니안–마스트리히트기) 후반, 네오코리스토데레 ''샴포사우루스''는 라라미디아 섬 서부 내륙 해양의 서부 해안을 따라 유타, 와이오밍, 몬태나, 노스다코타, 앨버타, 서스캐처원에서 발견된다.[5] 불확실한 네오코리스토데레 유해는 캐나다 북극 고지대에서 후기 백악기 초기 (코니아시안–투로니안)로 거슬러 올라가며,[38] 애팔래치아의 분리된 섬을 형성한 뉴저지주 네이슨크 지층에서 최후기 백악기(마스트리히트기)에 발견된다.[39] 독일 세노마니안[40]과 오스트리아 캄파니안 시대 그룬바흐 지층에서 발견된 척추뼈는 이 기간 동안 유럽에 코리스토데레가 존재했음을 나타낸다. 후기 백악기 아시아에서 코리스토데레의 유일한 기록은 일본 투로니안에서 발견된 단일 척추뼈이다.[42] 캐나다 앨버타의 캄파니안 시대 올드만과 공룡 공원 지층에서 발견된 단편적인 유해는 후기 백악기 동안 북아메리카에 소형 "비 네오코리스토데레"가 존재했을 가능성을 시사한다.[5]
''샴포사우루스''는 K-Pg 멸종에서 살아남았고, 동료 네오코리스토데레인 ''코스모드라코[1]'' 및 ''시모에도사우루스''와 함께 고생대 동안 유럽, 아시아, 북아메리카에서 발견되지만, 초기 에오세에 멸종되었다. 이들의 멸종은 상승된 온도와 관련된 주요 동물군 변화와 일치한다.[5] 초기 백악기 이후 화석 기록에서 발견되지 않는(가능한 북미 유해 제외) 소형 "비 네오코리스토데레"는 프랑스 후기 고생대에서 도마뱀과 같은 ''라자루수쿠스'' 형태로 다시 나타난다.[43] 유럽 고유종 ''라자루수쿠스''는 에오세 초기 네오코리스토데레 멸종에서 살아남은 마지막 코리스토데레로 알려져 있으며, 가장 젊은 유해는 체코 초기 마이오세에서 발견된 ''L. 드보라키''의 유해이다.[5][44] 독일 남부 후기 마이오세(~11.6Ma)에서 보고된 불확실한 ''라자루수쿠스'' 유해가 있을 수 있다.[45]
참조
[1]
논문
High morphological disparity in a bizarre Paleocene fauna of predatory freshwater reptiles
2022
[2]
서적
The Rise of Reptiles: 320 Million Years of Evolution
Johns Hopkins University Press
[3]
논문
On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana
https://www.biodiver[...]
[4]
논문
The Choristodera
https://www.biodiver[...]
[5]
논문
Choristoderes and the freshwater assemblages of Laurasia
[6]
서적
Evolutionary Biology
Springer US
[7]
논문
The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei province, People's Republic of China
https://discovery.uc[...]
[8]
논문
A neomorphic ossification connecting the braincase, squamosal, and quadrate in choristoderan reptiles: insights from µCT data
https://fr.pensoft.n[...]
2022-07-02
[9]
논문
Computed tomography analysis of the cranium of Champsosaurus lindoei and implications for the choristoderan neomorphic ossification
2020-04
[10]
논문
Osteology and taxonomic revision of Hyphalosaurus (Diapsida: Choristodera) from the Lower Cretaceous of Liaoning, China
2008-06
[11]
논문
The internal cranial anatomy of Champsosaurus (Choristodera: Champsosauridae): Implications for neurosensory function
2020-04-28
[12]
논문
Morphology and function of the palatal dentition in Choristodera
2016-03
[13]
논문
Exceptional fossil material of a semi-aquatic reptile from China: the resolution of an enigma
2000-09-25
[14]
논문
A new specimen of the Early Cretaceous long-necked choristodere Hyphalosaurus from Liaoning, China with exceptionally-preserved integument
https://linkinghub.e[...]
2023-04
[15]
논문
Aspects of some anatomical structures of ''Champsosaurus'' (Reptilia: Eosuchia)
1985-06
[16]
논문
Feeding behaviour and functional morphology of the neck in the long-snouted aquatic fossil reptile Champsosaurus (Reptilia: Diapsida) in comparison with the modern crocodilian Gavialis gangeticus
[17]
논문
Cannibalism in a semi-aquatic reptile from the Early Cretaceous of China
https://doi.org/10.1[...]
2005-02-01
[18]
논문
Extended embryo retention and viviparity in the first amniotes
2023-06-12
[19]
논문
Cretaceous choristoderan reptiles gave birth to live young
2010-04
[20]
논문
Implications of flexible-shelled eggs in a Cretaceous choristoderan reptile
2010-04
[21]
논문
Post-natal parental care in a Cretaceous diapsid from northeastern China
http://eprints.linco[...]
2014-10-20
[22]
논문
Fusion of sacrals and anatomy in ''Champsosaurus''(Diapsida, Choristodera)
[23]
논문
The first possible choristoderan trackway from the Lower Cretaceous Daegu Formation of South Korea and its implications on choristoderan locomotion
2020-09
[24]
논문
Ceratopsid tracks and associated ichnofauna from the Laramie Formation (Upper Cretaceous: Maastrichtian) of Colorado
http://www.tandfonli[...]
1995-09-14
[25]
논문
A new choristodere (Reptilia: Choristodera) from an Aptian–Albian coal deposit in China
https://www.tandfonl[...]
2020-08-02
[26]
논문
A new choristodere (Reptilia: Diapsida) from the Lower Cretaceous of western Liaoning Province, China, and phylogenetic relationships of Monjurosuchidae
2005-11-16
[27]
논문
Revealing body proportions of the enigmatic choristodere Khurendukhosaurus from Mongolia.
http://www.app.pan.p[...]
2019
[28]
논문
The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms
2016-04-28
[29]
논문
New choristoderes (Reptilia: Diapsida) from the Upper Cretaceous and Palaeocene, Alberta and Saskatchewan, Canada, and phylogenetic relationships of Choristodera
1998-12
[30]
서적
Der Lettenkeuper: ein Fenster in die Zeit vor den Dinosauriern
Staatliches Museum für Naturkunde Stuttgart
2015
[31]
논문
The earliest possible choristodere (Diapsida) and gaps in the fossil record of semi-aquatic Reptiles
http://jgs.lyellcoll[...]
1993-11-01
[32]
논문
The relationships and paleoecology of Pachystropheus rhaeticus , an enigmatic latest Triassic marine reptile (Diapsida: Thalattosauria)
https://www.tandfonl[...]
2024-06-04
[33]
논문
First Jurassic Choristodera from Asia
https://doi.org/10.1[...]
2006-01-01
[34]
논문
Middle Jurassic vertebrate assemblage of Berezovsk coal mine in western Siberia (Russia)
2016
[35]
논문
Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa
https://hal-insu.arc[...]
[36]
논문
A Barremian neochoristodere from the Cedar Mountain Formation, Utah, U.S.A.
http://dx.doi.org/10[...]
2006-12-11
[37]
간행물
Vertebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): implications for continental faunal turnover at the J/K boundary
https://hal.archives[...]
Museum National d’Histoire Naturelle Paris
[38]
논문
A fossil champsosaur population from the high Arctic: Implications for Late Cretaceous paleotemperatures
https://linkinghub.e[...]
2007-05
[39]
논문
An Appalachian population of neochoristoderes (Diapsida, Choristodera) elucidated using fossil evidence and ecological niche modelling
https://onlinelibrar[...]
[40]
논문
Filling the biostratigraphical gap: First choristoderan from the Lower - mid-Cretaceous interval of Europe
2018-12-13
[41]
논문
Erster nachweis von Choristodera (Reptilia, Diapsida) in der Oberkreide Europas: Champsosaurierwirbel aus den Gosau-Schichten (Campan) Niederösterreichs
https://www.zobodat.[...]
1989
[42]
논문
The first choristoderan record from the Upper Cretaceous of Asia, Tamagawa Formation, Kuji Group, Japan
https://linkinghub.e[...]
2021-08
[43]
논문
New material of the choristodere Lazarussuchus (Diapsida, Choristodera) from the Paleocene of France
[44]
논문
A choristoderan reptile (Reptilia: Diapsida) from the Lower Miocene of northwest Bohemia (Czech Republic)
[45]
논문
A biochronologic tie-point for the base of the Tortonian stage in European terrestrial settings: Magnetostratigraphy of the topmost Upper Freshwater Molasse sediments of the North Alpine Foreland Basin in Bavaria (Germany)
http://www.wahre-sta[...]
2016-08-01
[46]
저널
The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei Province, People’s Republic of China
https://www.tandfonl[...]
[47]
저널
Computed tomography analysis of the cranium of Champsosaurus lindoei and implications for the choristoderan neomorphic ossification
https://onlinelibrar[...]
[48]
저널
A new Miocene ape and locomotion in the ancestor of great apes and humans
http://www.nature.co[...]
[49]
저널
The first possible choristoderan trackway from the Lower Cretaceous Daegu Formation of South Korea and its implications on choristoderan locomotion
https://www.nature.c[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com