하체곱테릭스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
하체곱테릭스는 백악기 후기에 살았던 대형 익룡으로, 1899년 루마니아에서 화석이 처음 발견되었다. 날개폭이 최대 12m에 달하고 두개골 길이가 3m에 이르는 등, 케찰코아틀루스, 아람부르기아니아와 함께 가장 큰 익룡 중 하나로 여겨진다. 하체고프테릭스는 튼튼한 뼈 구조와 목뼈를 가지고 있었으며, 육상에서 먹이를 사냥하는 최상위 포식자였을 것으로 추정된다. 하체그 섬 생태계에서 섬 왜소증 공룡들을 포함한 다양한 동물들과 공존했으며, 하체고프테릭스가 발견된 지역은 현재 하체그 컨트리 공룡 지질공원 및 유네스코 세계 지질공원으로 지정되었다.
더 읽어볼만한 페이지
- 트란실바니아의 지리 - 트란실바니아알프스산맥
트란실바니아 알프스 산맥은 루마니아에 위치하며, 오스트리아 및 라라미아 조산 운동으로 형성되었고, 몰도베아누 봉을 포함하여 4개의 산악 그룹으로 구분되며 관광객과 과학자들에게 인기가 높다. - 마스트리흐트절의 생물 - 티라노사우루스
티라노사우루스는 백악기 후기 북아메리카에 서식했던 몸길이 15m에 달하는 거대한 육식 공룡으로, 강력한 턱힘과 짧은 앞다리, 긴 꼬리를 가진 최상위 포식자였으며, '수'와 '스카티' 등의 표본과 대중문화 속 인기 덕분에 그 생태와 행동에 대한 연구가 지속되고 있다. - 마스트리흐트절의 생물 - 아라리페수쿠스
아라리페수쿠스는 백악기 시대 남아메리카, 아프리카, 마다가스카르에 서식했고 곤드와나 기원을 시사하는 화석이 발견되었으며 튀어나온 주둥이, 큰 눈, 골배엽을 가진 육지 활동에 적합했을 것으로 추정되는 멸종된 악어형류 속이다. - 2002년 기재된 화석 분류군 - 호모 게오르기쿠스
호모 게오르기쿠스는 약 180만 년 전 조지아에서 발견된 멸종된 호미닌 종으로, 작은 뇌 용량과 원시적인 두개골을 가지며 아프리카 외 지역에서 가장 오래된 인류 중 하나로 여겨진다. - 2002년 기재된 화석 분류군 - 사헬란트로푸스
사헬란트로푸스는 약 700만 년 전에서 680만 년 전에 생존했던 화석 인류의 한 속으로, 2001년 차드에서 발견된 '투마이' 화석 표본을 통해 알려져 있으며, 작은 두개골과 턱뼈, 작은 뇌 용량을 특징으로 하고, 분류 및 직립 보행 여부에 대한 논쟁에도 불구하고 인류 진화 연구에 중요한 자료로 평가받는다.
| 하체곱테릭스 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Hatzegopteryx |
| 명명자 | Buffetaut, Grigorescu & Csiki, 2002 |
| 모식종 | Hatzegopteryx thambema |
| 모식종 명명자 | Buffetaut, Grigorescu & Csiki, 2002 |
| 화석 범위 | 마스트리흐트절, 6600만 년 전 |
| 분류 | |
| 상위 분류 | 아즈다르코과 |
| 특징 | |
| 추정 크기 | 날개폭 10~12m |
| 두개골 추정 길이 | 약 1.5m |
2. 발견
1899년 헝가리 출신의 고생물학자 프란츠 놉차가 루마니아에서 처음으로 익룡 화석을 발견했으며, 1970년대 후반 학생들의 발굴 활동을 통해 ''하체고프테릭스'' 화석이 처음으로 발견되었다.[8] 이 화석들은 루마니아 북서부 트란실바니아 서부 하체그 분지의 밸리오아라 지역에 있는, 후기 백악기 마스트리히트절(약 6,600만 년 전)에 속하는 덴슈-치울라 지층 상부에서 발견되었다. ''하체고프테릭스''의 모식표본인 FGGUB R 1083A는 두개골 후면 두 조각과 손상된 왼쪽 상완골 근위부로 구성되어 있다.[8] 이 조각 중 하나인 후두골은 처음 발표되었을 때 수각류 공룡으로 분류되기도 했다.[1][2] 인근에서 발견된 38.5cm 길이의 넙다리뼈 중간 부분(FGGUB R1625)도 ''하체고프테릭스''에 속할 수 있다.[5] FGGUB R1625는 날개 길이가 5m 에서 6m인 더 작은 개체의 ''하체고프테릭스''(해당 속에 속한다고 가정할 때)의 뼈일 수 있다. 이 지역에서 추가로 보고된 표본으로는 대형 개체의 미발표된 아래턱뼈가 있다.[10][3]
''하체고프테릭스''는 2002년 프랑스 고생물학자 에릭 뷔페토와 루마니아 고생물학자 단 그리고레스쿠, 졸탄 치키가 명명했다. 속명은 뼈가 발견된 트란실바니아의 ''하체그'' 분지에서 유래했으며, 고대 그리스어 단어 ''프테릭스''(ἡ πτέρυξ, -υγος 또는 ἡ πτερύξ, -ῦγος, "날개"를 의미)에서 유래했다. 종명 ''탐베마''는 그리스어로 "공포" 또는 "괴물"(τό θάμβημα, -ήματος[4])을 의미하며, 거대한 크기를 나타낸다.[8]
이후 다른 지역에서도 ''하체고프테릭스''의 새로운 표본이 발견되었다. 산페트루 지층의 바두, 선터마리아-올레아 지역에서 날개 길이가 4.5m 에서 5m인 개체에서 유래했을 것으로 추정되는 중간 크기의 견갑골-까마귀뼈가 발견되었다. 덴슈-치울라 지층과 동시대이며 인접한 세베슈 지층의 라파 로시에 지역에서 "RR 표본" 또는 EME 215로 명명된 단일 대형 목뼈가 발견되었다.[10][3] 이 표본은 겹치는 부분이 없어 ''하체고프테릭스 탐베마''로 확실하게 분류할 수는 없지만, 독특한 내부 골격 구조와 해당 지역에서 두 번째 거대한 아즈다르크과 익룡의 증거가 없다는 점을 고려하여 최소한 ''H. sp.''로 분류되었다.[12]
3. 특징

1899년 헝가리 출신 고생물학자 프란츠 놉차에 의해 루마니아에서 익룡 화석이 처음 발견되었고, 1970년대 후반 학생들의 발굴 활동을 통해 ''할체고프테릭스'' 화석이 처음으로 발견되었다. 이 화석들은 트란실바니아 서부, 루마니아 북서부 하체그 분지의 밸리오아라 지역, 후기 백악기 마스트리히트절 (약 6,600만 년 전) 덴슈-치울라 지층 상부에서 발견되었다. ''할체고프테릭스''의 모식표본인 FGGUB R 1083A는 두개골 후면 두 조각과 손상된 왼쪽 상완골 근위부로 구성된다.[8] 이 조각 중 하나인 후두골은 처음 발표되었을 때 수각류 공룡으로 분류되었다.[1][2] 근처에서 발견된 38.5cm 길이의 넙다리뼈 중간 부분(FGGUB R1625)도 ''할체고프테릭스''의 것일 수 있다.[5] FGGUB R1625는 날개 길이가 5m에서 6m인 더 작은 개체의 ''할체고프테릭스''(해당 속에 속한다는 가정 하에) 뼈일 수 있다. 이 지역에서 추가로 보고된 표본에는 대형 개체의 미발표된 아래턱뼈가 있다.[10][3]
''할체고프테릭스''는 2002년 프랑스 고생물학자 에릭 뷔페토와 루마니아 고생물학자 단 그리고레스쿠, 졸탄 치키에 의해 명명되었다. 속명은 뼈가 발견된 트란실바니아의 ''하체그'' 분지에서 유래되었으며, 고대 그리스어 단어 ''프테릭스''(ἡ πτέρυξ, -υγος 또는 ἡ πτερύξ, -ῦγος이며 "날개"를 의미)에서 유래되었다. 종명 ''탐베마''는 그리스어로 "공포" 또는 "괴물"(τό θάμβημα, -ήματος[4])을 의미하며, 거대한 크기를 나타낸다.[8]
이후 다른 지역에서도 ''할체고프테릭스''의 새로운 표본이 발견되었다. 산페트루 지층의 바두, 선터마리아-올레아 지역에서 날개 길이가 4.5m에서 5m인 개체에서 유래했을 것으로 추정되는 중간 크기의 견갑골-까마귀뼈가 발견되었다. 덴슈-치울라 지층과 동시대이며 인접한 세베슈 지층의 라파 로시에 지역에서 "RR 표본" 또는 EME 215로 명명된 단일 대형 목뼈가 발견되었다.[10][3] 이 표본은 겹치는 부분이 없어 ''할체고프테릭스 탐베마''로 확실하게 분류할 수는 없지만, 독특한 내부 골격 구조와 해당 지역에서 두 번째 거대한 아즈다르크과 익룡의 증거가 없다는 점을 고려하여 최소한 ''H. sp.''로 분류되었다.[12]
3. 1. 크기
하체곱테릭스는 케찰코아틀루스, 아람부르기아니아와 함께 가장 큰 익룡 중 하나로 여겨졌다. 초기에는 날개폭이 12m 이상으로 추정되었으나[8][5], 최근 연구에서는 10~11m 정도로 추정한다.[6]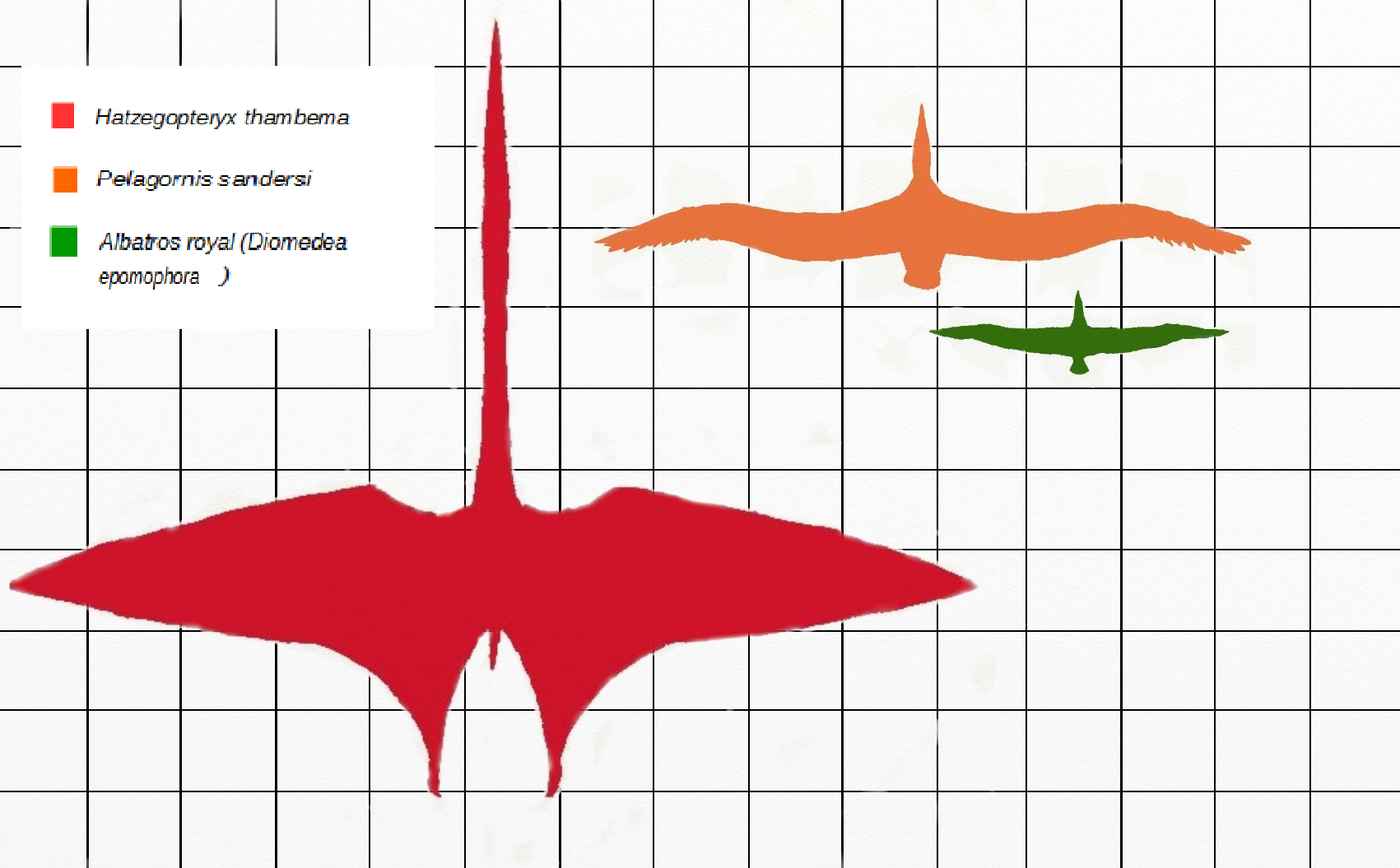
넓고 튼튼한 목뼈를 가졌다는 점을 근거로, 척주 전체가 유사하게 확장되었을 것이라는 주장이 제기되었다.[10] 그러나 익룡의 목뼈는 척추뼈보다 넓고 큰 경향이 있으므로, 척추뼈만으로 크기를 추정하는 것은 신뢰하기 어렵다.[12]
3. 2. 두개골
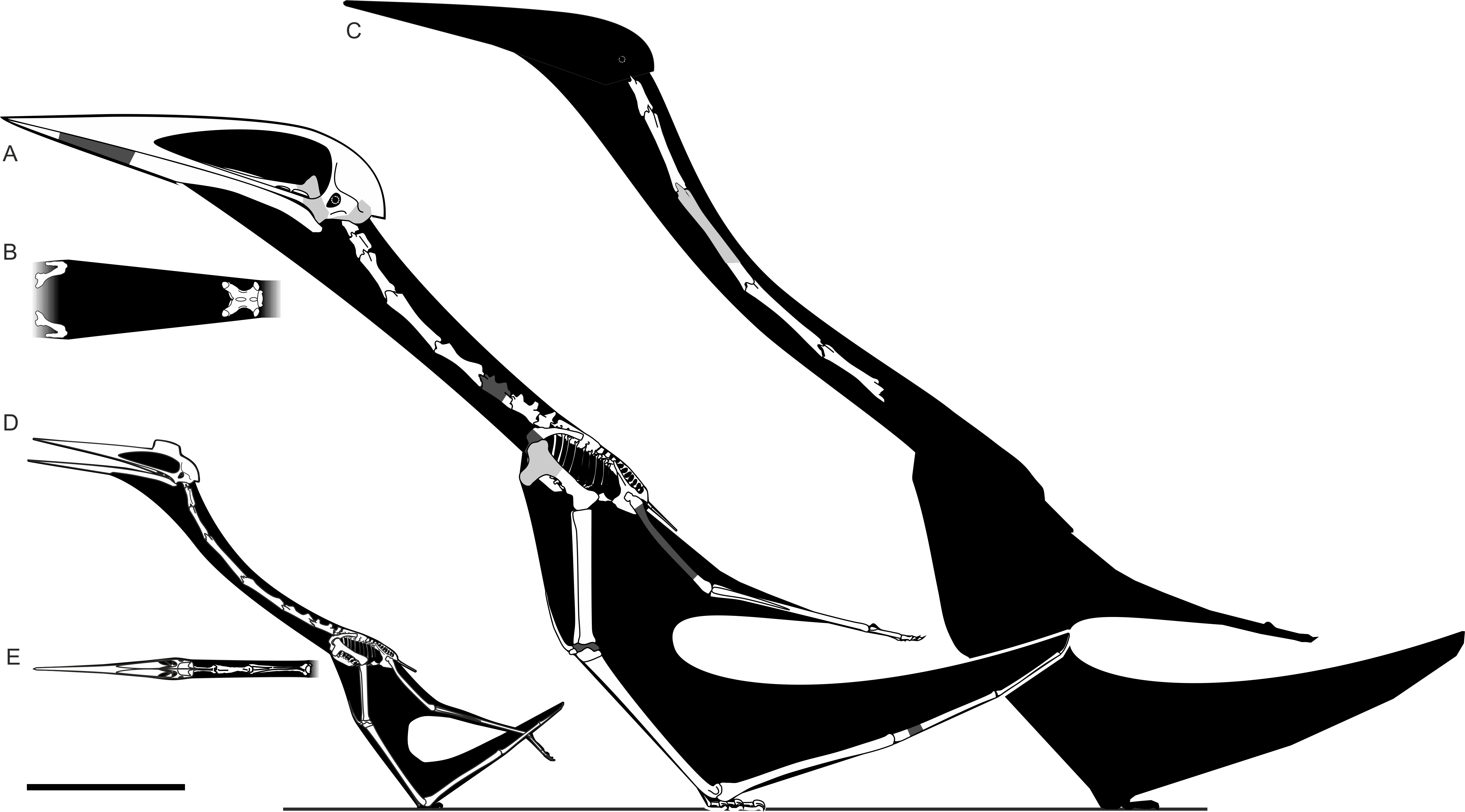
아즈다르코과 익룡인 하체곱테릭스의 두개골은 매우 컸으며, 닉토사우루스와 안항게라와 비교했을 때 추정 길이가 2.5m에 달해 비 해양 동물 중 가장 큰 두개골 중 하나였다.[5] 그러나 2018년 연구에서는 두개골 길이가 1.6m일 것으로 추정하기도 했다.[7] 두개골은 뒤쪽으로 넓어져 사각골을 가로질러 0.5m 너비였다.[5] 대부분의 익룡 두개골이 가늘고 가는 판과 버팀대로 구성된 반면, 하체곱테릭스의 두개골 뼈는 튼튼하고 견고하며 강한 근육 부착을 나타내는 큰 능선이 있었다.[8] 거대한 턱에는 프테라노돈 등 다른 익룡에서도 볼 수 있는 뚜렷한 홈이 있어 입을 매우 넓게 벌릴 수 있었다.[8] 미발표 유해를 통해 하체곱테릭스는 "가늘고 긴 부리"의 아즈다르코과보다는 "무딘 부리"의 아즈다르코과에 속할 가능성이 제기되었다.[9]
3. 3. 경추(목뼈)
하체곱테릭스의 목뼈는 짧고 유난히 튼튼하다. 보존된 부분은 240mm 길이이며, 전체 뼈는 생전에 300mm였을 것으로 보인다.[10] 익룡은 아홉 개의 경추를 가지고 있었다.[11] 회귀 분석에 따르면 셋째부터 일곱째 경추까지 합친 길이는 1.508m였을 것이고, 가장 긴 다섯 번째 뼈는 약 400mm였을 것이다. 반면, 이와 유사하게 거대한 아람보우르기아니아의 같은 뼈는 2.652m였다. 이는 하체곱테릭스의 목이 같은 크기의 거대한 아즈다르크과의 예상 길이의 약 50~60% 정도임을 나타낸다.[12]
목뼈의 아래쪽 표면은 4mm에서 6mm로 유난히 두꺼웠다. 아람보우르기아니아를 포함한 다른 대부분의 거대한 아즈다르크과의 경우, 이 표면의 두께는 2.6mm 미만이다. 뼈의 신경극이 완전히 보존되지는 않았지만, 보존된 부분의 너비는 다른 익룡에 비해 비교적 높고 튼튼했을 것으로 보인다. 뼈의 다른 측면은 더 작은 아즈다르코의 일곱 번째 목뼈와 가장 유사하다. 관절 소켓(cotyles)은 신경궁보다 훨씬 얕고 높이보다 너비가 네 배나 크며, 뼈의 아래쪽 부분에 있는 돌기인 하이파포피시스(hypapophysis)가 있고, 뼈 앞쪽의 돌기인 전관절돌기가 벌어져 있으며, 뼈의 중심 중간에 가늘어지는 "허리"가 있다.[12] 처음에는 세 번째 목뼈로 확인되었지만,[10] 이러한 특징은 뼈가 목 뒤쪽, 더 구체적으로는 일곱 번째 뼈에서 나온 것임을 뒷받침한다.[12]
4. 분류
''하체고프테릭스''의 상완골은 케찰코아틀루스 노스로피의 상완골과 유사하게 길고 매끄러운 삼각근능선과 두꺼운 상완골 머리를 가지고 있어 처음에는 아즈다르크과로 분류되었다.[8] 이 때문에 두 종이 동의어일 가능성도 제기되었으나,[13] 목과 턱의 해부학적 구조가 케찰코아틀루스와는 뚜렷하게 구별되어 별개의 분류군으로 유지되고 있다.[8][12][14]
''하체고프테릭스 sp.''에 속하는 목 척추는 중심이 낮고, 지아포피시스가 크고 편평하며, 신경 극이 이분되어 있는 등 아즈다르크과의 척추 특징을 보인다.[12]
2018년 계통 분석 결과, ''하체고프테릭스''는 아즈다르크과 내에서 아람부르기아나와 케찰코아틀루스의 자매 분류군으로 나타났다.[15]
5. 고생물학
하체곱테릭스는 뼈 구조를 통해 비행 능력을 갖추었을 것으로 추정된다. 두개골은 크고 튼튼했지만, 뼈 내부는 10mm 길이의 작은 구멍(폐포)과 얇은 뼈 지지대(trabeculae)로 이루어져 스티로폼과 유사한 구조를 가졌다.[8] 이는 두개골을 가볍게 만들어 비행에 유리하게 작용했을 것이다.[8] 목뼈 또한 유사한 내부 구조를 가졌다.[12]
거대한 목뼈는 짧고 튼튼하며, 보존된 부분은 240mm 길이지만 생전에는 300mm였을 것으로 보인다.[10] 회귀 분석에 따르면 셋째에서 일곱째 경추는 합쳐서 1.508m였을 것이고, 가장 긴 다섯 번째 뼈는 약 400mm였을 것이다. 이는 유사한 크기의 아람부르기아니아의 목 길이(약 2.652m)의 50~60% 수준이다.[12]
목뼈 아래쪽 표면은 4mm에서 6mm로 두꺼웠는데, 이는 아람부르기아니아를 포함한 다른 거대 아즈다르크과 익룡(약 2.6mm 미만)보다 두꺼운 수치이다. 뼈의 신경극은 완전히 보존되지 않았지만, 다른 익룡에 비해 높고 튼튼했을 것으로 추정된다. 뼈의 다른 측면은 아즈다르코의 일곱 번째 목뼈와 유사하며, 관절 소켓이 얕고, 하이파포피시스(hypapophysis)가 있으며, 전관절돌기가 벌어져 있고, 척추 몸통 중간에 "허리"가 가늘어지는 특징을 보인다.[12]
하체곱테릭스의 목은 아람부르기아니아보다 훨씬 강했는데, 상대적 파괴력으로 비교하면 하체곱테릭스의 뒤쪽 목 척추뼈는 체중의 5~10배를 견딜 수 있었던 반면, 아람부르기아니아는 체중의 절반 정도에서 골절되었다.[12]
하체곱테릭스의 척추 몸통은 아람부르기아니아보다 튼튼하지만, 뼈 반경 대 뼈 두께의 비율(R/t)은 비슷하다(하체곱테릭스 9.45, 아람부르기아니아 9.9).[6] 이는 굴곡 강도와 좌굴 강도 사이의 절충을 보여주는 것으로, 하체곱테릭스는 뼈의 독특한 내부 구조와 척추뼈의 큰 관절을 통해 좌굴 강도를 향상시키는 적응을 보였다.[12]
튼튼한 머리를 지탱하기 위해 하체곱테릭스의 목은 근육이 발달했을 것으로 보인다. 후두골의 후두선은 발달되어 눈에 띄는 흉터를 가졌으며, 이는 머리와 목의 신전과 굴곡을 돕는 가로척추근을 지지했을 수 있다. 뒤통수 돌기, 신경 가시, 접합돌기 등도 크고 튼튼하며, 후두골 결절은 길어 다양한 근육 부착점 역할을 했을 것이다.[12]
5. 1. 뼈 구조
하체곱테릭스의 두개골은 크고 튼튼했지만, 날개뼈는 비행 익룡과 비교할 수 있어 비행 능력이 있었음을 시사한다.[8] 두개골 무게 감소는 뼈 내부 구조를 통해 이루어졌을 것으로 추정된다. 뼈는 10mm 길이의 작은 구멍(폐포)과 얇은 뼈 지지대(trabeculae) 매트릭스로 분리되어 스티로폼과 유사한 구조를 가졌다.[8] 이러한 구조는 두개골을 튼튼하고 스트레스에 강하게 만들면서도 가볍게 만들어 비행을 가능하게 했을 것이다.[8] 목뼈 또한 유사한 내부 구조를 가졌다.[12]하체곱테릭스의 거대한 목뼈는 짧고 튼튼하다. 보존된 부분은 길이가 240mm이고, 전체 뼈는 생전에 300mm였을 것으로 보인다.[10] 익룡은 아홉 개의 경추를 가지고 있었다.[11] 회귀 분석에 따르면 셋째에서 일곱째 경추는 합쳐서 길이가 1.508m였을 것이고, 가장 긴 뼈인 다섯 번째 뼈는 길이가 약 400mm에 불과했을 것이다. 반면에, 이와 유사하게 거대한 ''Arambourgiania''의 같은 뼈는 2.652m였다. 이는 할체고프테릭스의 목이 같은 크기의 거대한 아즈다르크과의 예상 길이의 약 50~60% 정도임을 나타낸다.[12]
목뼈의 아래쪽 표면 또한 4mm에서 6mm로 두꺼웠다. ''아람부르기아나''를 포함한 다른 대부분의 거대한 아즈다르크과의 경우, 이 표면의 두께는 2.6mm 미만이다. 뼈의 신경극이 완전히 보존되지는 않았지만, 보존된 부분의 너비는 다른 익룡에 비해 비교적 높고 튼튼했을 것으로 보인다. 뼈의 다른 측면은 더 작은 ''Azhdarcho''의 일곱 번째 목뼈와 가장 유사하다. 관절 소켓(cotyles)은 신경궁보다 훨씬 얕고 높이보다 너비가 네 배나 크며, 뼈의 아래쪽 부분에 있는 돌기인 하이파포피시스(hypapophysis)가 있고, 뼈 앞쪽의 돌기인 전관절돌기가 벌어져 있으며, 뼈의 중심 중간에 가늘어지는 "허리"가 있다.[12]
5. 2. 목의 생체 역학
하체곱테릭스의 목은 아람부르기아니아의 목보다 훨씬 강했는데, 이는 척추뼈의 골절력과 익룡의 체중을 나눈 값인 상대적 파괴력으로 정량화할 수 있다. 아람부르기아니아의 목 척추뼈는 체중의 약 절반에서 골절되는 반면, 하체곱테릭스의 뒤쪽 목 척추뼈는 뼈의 하중에 따라 체중의 5~10배를 견딜 수 있었다. 가상적으로 더 긴 하체곱테릭스의 앞쪽 목 척추뼈도 체중의 4~7배를 견딜 수 있었을 것이다.[12]하체곱테릭스의 척추 몸통은 아람부르기아니아보다 훨씬 튼튼하지만, 뼈 반경 대 뼈 두께의 비율(R/t)[6]은 거의 동일하다(하체곱테릭스는 9.45, 아람부르기아니아는 9.9). 이는 굴곡 강도와 좌굴 강도를 증가시키는 사이의 절충안을 나타낼 수 있다. R/t 비율이 높을수록 굴곡 강도가 개선되지만 좌굴 강도는 약해진다. 이를 보완하기 위해 하체곱테릭스는 좌굴 강도를 향상시키기 위한 여러 가지 적응을 보여주는데, 뼈의 독특한 내부 구조와 척추뼈의 큰 관절이 그것이다. 이들은 스트레스를 분산시키는 데 도움이 된다.[12]
튼튼한 머리를 지탱하기 위해 하체곱테릭스의 목은 근육이 발달했을 가능성이 높다. 후두골에서 근육 부착점 역할을 하는 후두선은 매우 발달되어 있으며 눈에 띄는 흉터를 가지고 있다. 이는 머리와 목의 신전과 굴곡을 돕는 가로척추근을 지지했을 수 있다. 마찬가지로, 뒤통수 돌기, 신경 가시 및 접합돌기는 모두 크고 튼튼해 보였고(후자는 근육 흉터를 나타낼 가능성이 있는 많은 구멍과 가장자리를 가지고 있음), 후두골 결절은 길었다. 이들은 모두 머리와 목의 다양한 근육 부착점 역할을 한다.[12]
6. 고생태

아즈다르크과 익룡인 ''하체고프테릭스''(Hatzegopteryx)는 육상에서 먹이를 찾는 잡식성 포식자였을 것으로 추정된다.[16] 마스트리히트절 유럽의 다른 육상 포식자들보다 훨씬 컸으며, 섬 왜소증 공룡이 지배적인 환경에서 큰 크기(이 지역에는 큰 육식 수각류가 없음) 때문에 하체그 섬(Hațeg Island) 생태계에서 최상위 포식자 역할을 했을 것으로 보인다. ''하체고프테릭스''의 튼튼한 해부학적 구조는 다른 아즈다르크과보다 더 큰 먹이를 다루었을 가능성을 시사하며, 이는 통째로 삼키기에는 너무 큰 동물까지 포함한다.[16] 반면, ''아람부르기아''(Arambourgiania)와 같은 다른 거대한 아즈다르크과는 부화하거나 작은 공룡과 알을 포함하여 작은 먹이(사람 크기까지)를 먹었을 것이다.[12]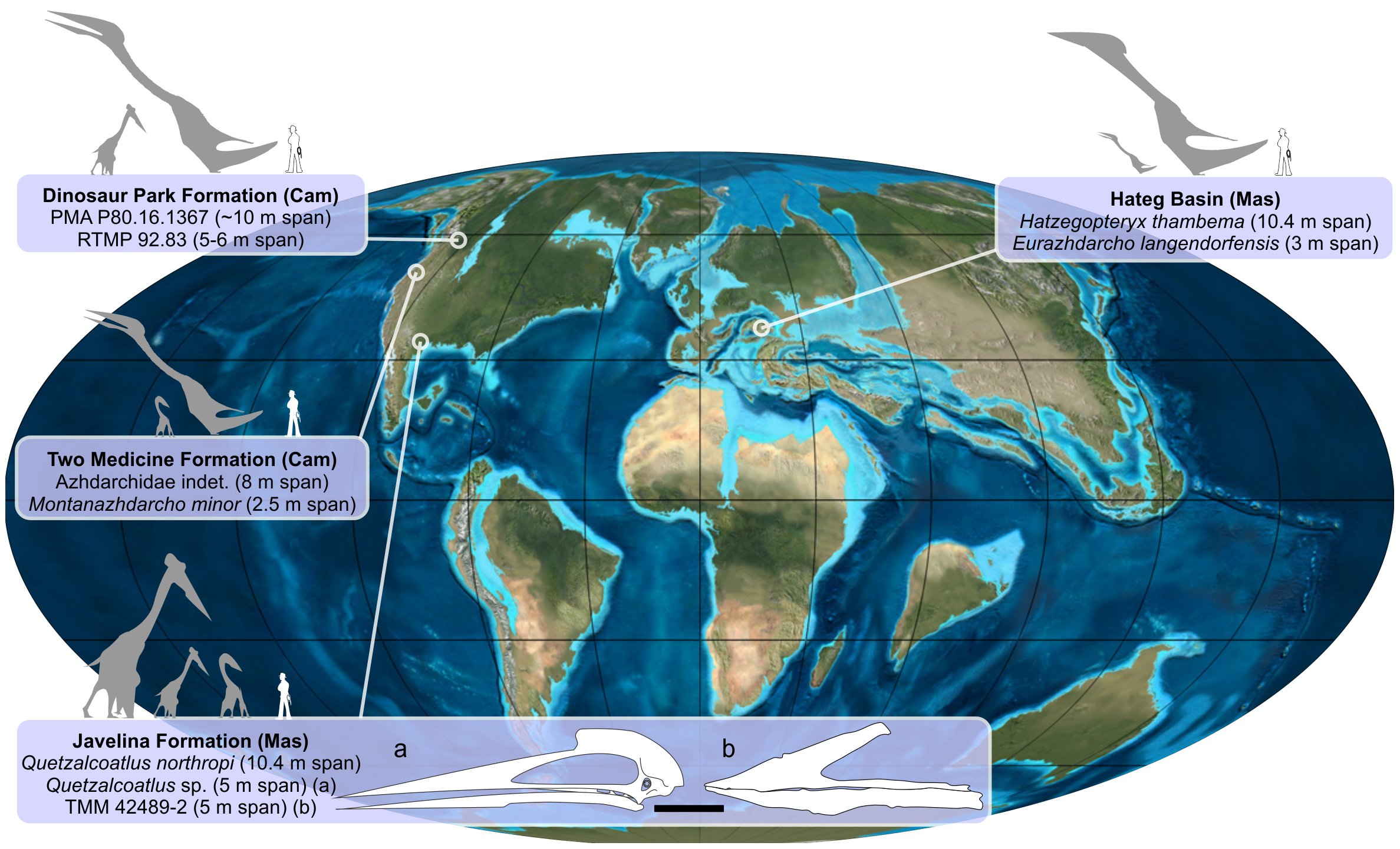
하체그 섬 생태계에는 ''하체고프테릭스'' 외에도 다양한 생물들이 서식했다. 함께 발견된 익룡에는 날개 길이가 3.8m인 작은 아즈다르크과인 ''유라즈다르코''(Eurazhdarcho)[3], 날개 길이가 3.5m에서 4m인 이름 없는 작은 크기의 짧은 목 아즈다르크과, 날개 길이가 5m인 다소 크고 이름 없는 아즈다르크과, 그리고 작은 프테라노돈과(pteranodontidae) 익룡이 있었다.[18] 튼튼하고 날지 못하며 초식성이었을 가능성이 있는 조류(avialae)[19] 또는 드로마에오사우루스과(dromaeosauridae)[20] ''발라우르 본독''(Balaur bondoc)''는 각 발에 두 개의 확대된 발톱을 가지고 있었는데,[20] 이는 이 동물군에서 매우 특이한 요소였다.
이 생태계에는 섬 왜소증을 겪은 티타노사우루스(titanosaur) ''마자르사우루스''(Magyarosaurus)[22]와 ''팔루디티탄''(Paludititan)[21], 하드로사우루스과(hadrosaurid) ''텔마토사우루스''(Telmatosaurus)''와 이구아노돈류(iguanodontia) ''잘목세스''(Zalmoxes)''[22], 노도사우루스과(nodosauridae) ''스트루티오사우루스''(Struthiosaurus)'', ''브라디크네메''(Bradycneme)'', ''엘롭테릭스''(Elopteryx)'', ''헵타스테오르니스''(Heptasteornis)''를 포함한 다양한 작고 파편적인 마니랍토라(maniraptora)도 서식했다.[22] ''알로다포수쿠스''(Allodaposuchus)'', ''도라토돈''(Doratodon)'', ''아키노돈''(Acynodon)'' 속의 악어 유해도 발견되었다.[23] 비archosaur(archosaur) 구성 요소에는 코가이오니아과(kogaionidae) 다지결절류(multituberculata) 포유류인 ''코가이오논''(Kogaionon)'', ''바르바토돈''(Barbatodon)'', ''리토보이 톨로세팔로스''(Litovoi tholocephalos)'', ''하이니나''(Hainina)''[24][25], 테이아과(teiidae) ''비쿠스피돈''(Bicuspidon)''과 파라마켈로두스과(paramacellodid) ''베클레시우스''(Becklesius)''와 같은 도마뱀, 이름 없는 마드토이아과(madtsoiidae) 뱀, 양서류(lissamphibia)인 ''알바네르페톤''(Albanerpeton)'', ''에오디스코글로수스''(Eodiscoglossus)'', ''파라디스코글로수스''(Paradiscoglossus)''가 포함되었다.[26]
이 동물군의 중요성은 2004년부터 2005년까지 이 지역을 유럽 지질공원 네트워크(European Geoparks Network)의 초기 멤버 중 하나인 하체그 컨트리 공룡 지질공원(Hațeg Country Dinosaurs Geopark)으로, 그리고 2015년 유네스코 세계 지질공원(UNESCO Global Geoparks) 지정이 비준되었을 때 하체그 유네스코 세계 지질공원으로 지정하는 주요 지질학적 근거가 되었다.[27]
마스트리히트절 동안 남유럽은 군도였다. 하체그 섬 생태계의 구성원들은 하체그 분지가 작은 부분이었던 티시아-다키아 블록(Tisia–Dacia Block)이라고 알려진 육지에 살았다. 이 육지는 약 80000km2 면적이었으며, 200km에서 300km 거리의 깊은 대양에 의해 모든 방향으로 다른 육지와 분리되어 있었다.[22] 위도 27°N에 위치하여,[28] 현재 위도 45°N보다 더 남쪽에 있었다. 따라서 기후는 아열대성으로 건기와 우기가 뚜렷했으며, 평균 기온은 약 25°C였다.[29] 환경은 다양한 충적 평야(alluvial plain), 습지(wetland), 강(river)으로 구성되었으며,[30] 양치류와 피자식물(angiosperms)이 지배적인 숲으로 둘러싸여 있었다.[22] 고토양(Paleosol)은 연 강수량이 1000mm 미만인 비교적 건조한 백악기 기후를 나타낸다.[29]
참조
[1]
서적
The Dinosauria
University of California Press
2004
[2]
웹사이트
New paper: when the short-necked, giant azhdarchid pterosaur ''Hatzegopteryx'' ruled Late Cretaceous Romania
http://markwitton-co[...]
2017
[3]
논문
A New Azhdarchid Pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for Azhdarchid Diversity and Distribution
2013
[4]
웹사이트
θάμβ-ημα
https://www.perseus.[...]
Tufts University
1940
[5]
논문
Giant azhdarchid pterosaurs from the terminal Cretaceous of Transylvania (western Romania)
http://sp.lyellcolle[...]
2003
[6]
논문
On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness
2010
[7]
간행물
Partial mandible of a giant pterosaur from the uppermost Cretaceous (Maastrichtian) of the Hațeg Basin, Romania
2018
[8]
논문
A new giant pterosaur with a robust skull from the latest Cretaceous of Romania
http://doc.rero.ch/r[...]
2002
[9]
학술회의
Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania
http://svpca.org/abs[...]
2013
[10]
논문
New faunal elements from the Late Cretaceous (Maastrichtian) continental deposits of Sebeș area (Transylvania)
http://www.cclbsebes[...]
2010
[11]
논문
A new specimen of the pterosaur ''Scaphognathus crassirostris'', with comments on constraint of cervical vertebrae number in pterosaurs
2014
[12]
논문
Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators
2017
[13]
학술회의
Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity
http://cstm.cnki.net[...]
2010
[14]
웹사이트
''Quetzalcoatlus'': the media concept vs the science
http://markwitton-co[...]
2016
[15]
논문
Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary
2018
[16]
논문
Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"?
2015
[17]
서적
Pterosaurs: Natural History, Evolution, Anatomy
http://press.princet[...]
Princeton University Press
2013
[18]
논문
A medium-sized robust-necked azhdarchid pterosaur (Pterodactyloidea: Azhdarchidae) from the Maastrichtian of Pui (Hațeg Basin, Transylvania, Romania)
https://www.biodiver[...]
2015
[19]
논문
The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod ''Balaur bondoc'' (Dinosauria, Maniraptora): dromaeosaurid or flightless bird?
[20]
논문
An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania
2010
[21]
논문
A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nălaţ-Vad, Hațeg Basin, Romania
2010
[22]
논문
Dinosaurs and the island rule: The dwarfed dinosaurs from Hațeg Island
https://www.academia[...]
2010
[23]
논문
Late Cretaceous crocodilian diversity in Hațeg Basin, Romania
https://www.research[...]
2006
[24]
논문
Teeth of multituberculate mammals from the Late Cretaceous of Romania
https://www.app.pan.[...]
2000
[25]
논문
Première identification d'une espèce de mammifère du Crétacé supérieur du bassin de Rusca Montană (Transylvanie, Roumanie)
2017
[26]
논문
New lissamphibians and squamates from the Maastrichtian of Hațeg Basin, Romania
https://www.app.pan.[...]
2005
[27]
웹사이트
Haţeg UNESCO Global Geopark
https://en.unesco.or[...]
2023-03-25
[28]
논문
Palaeomagnetism of the Upper Cretaceous Sânpetru Formation (Hațeg Basin, South Carpathians)
2010
[29]
논문
Palaeoenvironments of the latest Cretaceous (Maastrichtian) dinosaurs of Romania: insights from fluvial deposits and paleosols of the Transylvanian and Hațeg basins
http://doc.rero.ch/r[...]
2005
[30]
논문
Palaeoenvironmental reconstruction of the Late Cretaceous Sânpetru Formation (Hațeg Basin, Romania) using paleosols and implications for the "disappearance" of dinosaurs
2009
[31]
문서
分類学上、現在のところ未整理の階級。階級未定の分類群。以下同様。
[32]
서적
世界最大の翼竜展
朝日新聞社
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com