공룡
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
공룡은 중생대에 번성했던 지배 파충류로, 트리케라톱스와 조류의 가장 최근 공통 조상을 포함하는 분류군으로 정의된다. 공룡은 조반목과 용반목으로 나뉘며, 수각류, 용각류, 조반목 등 다양한 종류가 있다. 공룡은 트라이아스기 후기에 출현하여 중생대인 쥐라기와 백악기에 번성했으며, 깃털과 같은 특징을 가진 공룡도 존재했다. 공룡은 다양한 크기, 식성, 행동 양식을 보였으며, 육식 공룡과 초식 공룡이 모두 존재했다. 백악기 말의 소행성 충돌, 기후 변화, 화산 활동 등의 복합적인 요인으로 인해 비조류 공룡은 멸종되었지만, 조류는 살아남아 현재까지 존재한다. 공룡 화석은 뼈, 이, 알, 발자국 등 다양한 형태로 발견되며, 이를 통해 공룡의 생태와 진화에 대한 많은 정보를 얻을 수 있다. 공룡은 문화적으로도 큰 영향을 미쳐, 문학, 영화, 광고 등 다양한 매체에서 등장하며 대중의 상상력을 자극하고 있다.
더 읽어볼만한 페이지
- 1842년 기재된 화석 분류군 - 스밀로돈
스밀로돈은 플라이스토세 시대 아메리카 대륙에 살았던 멸종된 검치호 속으로, 긴 송곳니와 강력한 앞다리, 튼튼한 체격을 지녔으며 대형 초식동물을 사냥했던 최상위 포식자였으나 약 1만 년 전 멸종했다. - 1842년 기재된 화석 분류군 - 카메로케라스
카메로케라스는 고생대 오르도비스기에 서식한 다양한 크기의 두족류로, 촉수와 강력한 부리를 사용하여 먹이를 사냥했다. - 공룡 - 다이너소어 능선
다이너소어 능선은 콜로라도주 모리슨 근처에 위치한 백악기 지층 지역으로, 공룡 발자국과 화석이 발견되며, 골든 단층 등의 지질 구조와 함께 독특한 지형적 특징 및 석유, 가스 자원을 보유한 전시관이 있는 곳이다. - 공룡 - 우나이사우루스
우나이사우루스는 브라질에서 발견된 트라이아스기 후기의 작은 초식 공룡으로, 몸길이 약 2.5m이며 두 발로 걸었고, 브라질 최초의 용각류이다. - 리처드 오언이 명명한 분류군 - 극어류
극어류는 멸종된 어류의 한 무리로, 가시 상어라고도 불리며 실루리아기부터 페름기까지 번성했고, 비늘 장식으로 퇴적암 연대 추정에 사용되었으며, 연골어류에 속한다. - 리처드 오언이 명명한 분류군 - 악어
악어는 넓고 납작한 몸, 긴 주둥이, 짧은 목, 납작한 꼬리를 가진 파충류로 열대 및 아열대 지역에 서식하며 강력한 턱과 이빨로 다양한 먹이를 포식하고 인간과 연관이 깊지만 멸종 위기에 처해 보존 노력이 필요하다.
| 공룡 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Dinosauria |
| 학명 명명자 | Owen, 1842 |
| 한국어 이름 | 공룡류 |
| 영어 이름 | Dinosaur |
| 생물학적 분류 | |
| 도메인 | 진핵생물 |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 아문 | 척추동물아문 |
| 하문 | 유악하문 |
| 상강 | 사지상강 |
| 강 | 파충강 |
| 아강 | 이궁아강 |
| 하강 | 주룡형류 |
| 상목 | 공룡상목 |
| 시간 범위 | |
| 화석 범위 | 후기 삼첩기 - 홀로세 (약 2억 3140만 년 전 ~ 현재) |
| 비고 | (현생 조류 포함) |
| 하위 분류 | |
| 목 | 조반목 용반목 |
| 주요 그룹 | |
| 불확실한 친화 관계의 공룡 및 가능한 공룡 | |
2. 어원
공룡은 그리스어 δεινόσαυρος(데이노사우로스)의 번역어이다. 1842년 영국의 고생물학자 리처드 오웬은 파충류에 속하는 이 특별한 부류에 대해 처음으로 공룡이라는 이름을 붙였다.[311] δεινόσαυρος는 "무서운", "놀라운"을 뜻하는 그리스어 δεινός(데이노스)와 "도마뱀"을 뜻하는 σαύρα(사우라)가 합쳐진 단어이다.[312] 오웬은 공룡의 크기와 위엄있는 모습에 크게 감명받았으며, 이빨, 발톱 등의 생김새를 기준으로 분류하였다.[313]
분류학에서는 공룡을 "트리케라톱스와 근생 조류 및 이들에서 분기된 모든 자손을 아우르는 가장 최근의 공통 조상이 속해 있는 부류"로 보고 있다.[314] 이는 메갈로사우루스와 이구아노돈의 공통 조상이 곧 모든 공룡의 최근 공통 조상이라는 것을 의미한다. 메갈로사우루스와 이구아노돈은 리처드 오웬이 분류한 세 종류의 공룡 가운데 두 종류이다.[315]
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 공룡과 같은 시기에 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다. 어룡(어룡목), 모사사우루스(유린목), 플레시오사우루스(사경룡목)은 수중에서 서식하였으며 익룡(익룡목)은 날아다녔다.[318]
오늘날 다양한 발견이 이루어지면서 공룡의 특징을 일반화하는 것은 더욱 어려워졌다. 그러나 후기에 출현한 공룡의 특징이 초기 공룡의 그것과는 매우 다르게 변형되었다는 점을 감안하더라도 모든 공룡의 기본적 골격 구조는 지배파충류에서 파생되었다. 가장 초기에 출현한 이 공룡은 이후 다양하게 진화한 모든 공룡들의 기반이 되었다. 이와 같은 분류학적 특징을 파생공유형질이라 한다.
한국어 공룡(恐龍) 또한 무서운(恐) 용(龍)이란 뜻이다. 영어 dinosaur영어는 δεινός|deinos|ngrc "무시무시한" + σαῦρος|sauros|Ngrc "도마뱀"[216]의 합성어이다. 1842년 당시까지 발견되었던 3종의 화석 파충류(이구아노돈, 메갈로사우루스, 힐라에오사우루스)는 리처드 오웬에 의해 "공룡"이라는 새로운 분류명으로 통합, 명명되었다.[217]
오웬은 δεινόςgrc라는 단어를 영어로 fearfully great영어[218][219], 즉 무시무시할 정도로 큰[220]이라는 의미로 사용했다고 한다. 어근 -saur-에는 "용"을 붙이는 것이 통례이다.
3. 현대 생물학의 정의
계통분류학에서 공룡은 일반적으로 ''트리케라톱스''와 현생 조류(신조류)의 최근 공통 조상(MRCA)과 모든 후손으로 구성된 그룹으로 정의된다. 또한 리처드 오웬이 공룡을 인지했을 때 인용한 3개의 속 중 2개였던 ''메갈로사우루스''와 ''이구아노돈''의 MRCA와 관련하여 공룡강을 정의할 것을 제안하기도 했다.
조류는 유일하게 생존한 공룡이다. 전통적인 분류학에서 조류는 공룡으로부터 진화한 별도의 강으로 간주되었으며, 뚜렷한 상목이었다. 그러나 대부분의 현대 고생물학자들은 해부학적 유사성에 기초한 전통적인 분류 방식을 거부하고, 각 그룹을 주어진 창립 속의 모든 후손으로 정의하는 계통 발생학적 분류학을 선호한다.[10] 조류는 용반목인 코엘루로사우루스류인 공룡의 하위 그룹 마니랍토라에 속한다.
2017년 매튜 G. 배런, 데이비드 B. 노먼, 폴 M. 배럿의 연구는 공룡 계통학의 급진적인 수정을 제안했다. 배런 외의 계통 발생학적 분석 결과는 용각류와 수각류의 전통적인 결합과는 반대로 조반목이 용각류보다 수각류에 더 가깝다는 것을 밝혀냈다. 이것은 용각류와 그 친족을 전통적인 공룡 밖으로 떨어뜨릴 것이므로, 그들은 공룡을 ''트리케라톱스 호리두스'', ''집참새'' 및 ''디플로도쿠스 카르네기''의 마지막 공통 조상과 그 모든 후손으로 재정의하여 용각류와 그 친족이 공룡으로 계속 포함되도록 했다. 그들은 또한 조반목과 수각류를 포함하는 그룹을 지칭하기 위해 조류형류라는 분류군을 부활시켰다.[11][12]
4. 일반적 특징

공룡은 중생대에 크게 번성하였으며 특히 쥐라기와 백악기에 융성하였다. 다른 동물들은 공룡의 크기에 비할 수 없었는데, 설치류가 주를 이루었던 당시의 포유류는 지금의 고양이만한 크기가 일반적이었다.[319] 그러나 삼돌기치목에 속하는 레페노마무스와 같은 예외적인 동물도 있었다. 이 포유류는 몸무게가 12kg - 14kg에 달해 어린 프시타코사우루스와 같은 작은 공룡을 사냥할 정도였다.[320]
매우 다양한 종류의 공룡이 존재하였으며 2006년까지 밝혀진 바로는 500여 속에 이른다. 이 중에 골격 화석이 발견된 것은 약 75% 정도이다.[321] 초기 연구에서는 공룡을 3400여 속으로 추정하였기도 하였으나 오늘날 화석 기록과는 차이가 있다.[322] 2008년 10월 17일 현재 확인된 공룡의 종은 1047개이다.[323] 새를 포함하면 그 다양성은 증가한다.
공룡의 생김새, 크기, 식성, 행동 양식 등은 천차만별이었다. 초식 공룡과 육식 공룡이 모두 있었으며, 2족 보행을 하거나 4족 보행을 하였다. 플라테오사우루스나 이구아노돈 같은 공룡들은 2족 보행과 4족 보행을 모두 할 수 있었다. 거대한 크기의 공룡이 있었는가 하면 일반적으로 알려진 것과는 달리 많은 종류의 공룡이 인간과 유사한 크기이거나 그보다 작았다. 여러 종류의 공룡들이 자신의 몸을 보호하기 위한 뿔, 등딱지, 골갑, 볏 등을 갖고 있었다. 공룡은 지구 어디에서나 서식하였으며 남극에도 있었다.[324]
계통분류학에서 공룡은 일반적으로 ''트리케라톱스''와 현생 조류(신조류)의 최근 공통 조상 (MRCA)과 모든 후손으로 구성된 그룹으로 정의된다. 또한 리처드 오웬이 공룡을 인지했을 때 인용한 3개의 속 중 2개였던 ''메갈로사우루스''와 ''이구아노돈''의 MRCA와 관련하여 공룡강을 정의할 것을 제안하기도 했다. 두 정의 모두 동일한 알려진 속을 포함한다: 공룡강 = 조반목 + 용반목. 여기에는 안킬로사우루스류 (갑옷을 입은 초식 사족보행류), 스테고사우루스류 (판을 가진 초식 사족보행류), 케라톱스류 (목도리를 가진 이족 또는 사족보행 초식동물), 파키케팔로사우루스류 (두꺼운 두개골을 가진 이족 보행 초식동물), 조각류 (오리 주둥이를 포함한 이족 또는 사족보행 초식동물), 수각류 (주로 이족 보행 육식동물 및 조류), 용각류 (주로 긴 목과 꼬리를 가진 대형 초식 사족보행류)와 같은 주요 그룹이 포함된다.
조류는 유일하게 생존한 공룡이다. 전통적인 분류학에서 조류는 공룡으로부터 진화한 별도의 강으로 간주되었으며, 뚜렷한 상목이었다. 그러나 대부분의 현대 고생물학자들은 해부학적 유사성에 기초한 전통적인 분류 방식을 거부하고, 각 그룹을 주어진 창립 속의 모든 후손으로 정의하는 계통 발생학적 분류학을 선호한다.[10] 조류는 용반목인 코엘루로사우루스류 공룡의 하위 그룹 마니랍토라에 속한다.
2017년 매튜 G. 배런, 데이비드 B. 노먼, 폴 M. 배럿의 연구는 공룡 계통학의 급진적인 수정을 제안했다. 배런 외의 계통 발생학적 분석 결과는 용각류와 수각류의 전통적인 결합과는 반대로 조반목이 용각류보다 수각류에 더 가깝다는 것을 밝혀냈다. 이것은 용각류와 그 친족을 전통적인 공룡 밖으로 떨어뜨릴 것이므로, 그들은 공룡을 ''트리케라톱스 호리두스'', ''집참새'' 및 ''디플로도쿠스 카르네기''의 마지막 공통 조상과 그 모든 후손으로 재정의하여 용각류와 그 친족이 공룡으로 계속 포함되도록 했다. 그들은 또한 조반목과 수각류를 포함하는 그룹을 지칭하기 위해 조류형류라는 분류군을 부활시켰다.[11][12]
위 정의 중 하나를 사용하여 공룡을 일반적으로 "다리"가 몸 아래에 세워진 뒷다리를 가진 아르코사우루스류로 묘사할 수 있다. 익룡, 모사사우루스, 어룡, 플레시오사우루스, 그리고 ''디메트로돈''을 포함한 다른 선사 시대 동물들은 종종 대중적으로 공룡으로 여겨지지만, 분류학적으로 공룡으로 분류되지 않는다. 익룡은 공룡과 멀리 관련된 종으로, 조반류에 속한다. 언급된 다른 그룹들은 공룡과 익룡과 마찬가지로 파충류에 속하지만, ''디메트로돈''(이는 단궁류)은 제외이다. 이들 중 어떤 것도 진정한 공룡의 특징인 똑바로 선 뒷다리 자세를 갖지 않았다.
공룡은 특히 쥐라기와 백악기 시대에 중생대의 지배적인 육상 척추 동물이었다. 다른 동물 그룹들은 크기와 생태적 지위가 제한적이었다. 예를 들어 포유류는 드물게 집 고양이의 크기를 초과했으며 일반적으로 설치류 크기의 작은 먹이를 잡아먹는 육식동물이었다. 공룡은 항상 매우 다양한 그룹으로 인식되어 왔다. 900개 이상의 비조류 공룡 속이 2018년에 확실하게 확인되었으며 1124종(2016년)이 있다. 화석 기록에 보존된 공룡 속의 총 수는 1850개로 추정되며, 거의 75%가 아직 발견되지 않았고,[13][14], 이전에 존재했던(화석 기록 내 또는 외부) 수는 3,400개로 추정된다. 2016년 추정치는 중생대에 살았던 공룡 종의 수를 1,543~2,468로 추정했다.[15][16], 이는 현대 조류(조류 공룡)의 수인 10,806종과 비교된다.[17]
멸종된 공룡과 현대 조류는 종자 식물, 어류, 곤충, 잡식성 동물을 포함하여 초식 및 육식성 동물을 포함한다. 공룡은 조상적으로 두 발로 걸었지만(모든 현대 조류와 마찬가지로), 일부는 네 발로 진화했고, ''안키사우루스''와 ''이구아노돈''과 같은 다른 종들은 두 다리 또는 네 다리로 쉽게 걸을 수 있었다. 뿔과 볏과 같은 두개골 수정은 일반적인 공룡 특성이며, 일부 멸종된 종은 골질 갑옷을 가지고 있었다. 가장 잘 알려진 속은 그 큰 크기로 유명하지만, 많은 중생대 공룡은 인간 크기 또는 그보다 작았으며, 현대 조류는 일반적으로 작은 크기이다. 오늘날 공룡은 모든 대륙에 살고 있으며, 화석은 그들이 늦어도 초기 쥐라기에 이르러 전 세계적으로 분포했음을 보여준다.[18] 현대 조류는 육상에서 해양에 이르기까지 이용 가능한 대부분의 서식지에 살고 있으며, 일부 비조류 공룡(예: ''마이크로랍토르'')이 날 수 있거나 적어도 활공할 수 있다는 증거가 있으며, 스피노사우루스과와 같은 다른 종은 반수생 습성을 가지고 있었다.
5. 해부학적 특징
공룡은 진화에 의한 종분화를 겪으면서 다양한 파생공유형질이 나타났다. 장골, 경골, 삼각근, 골반뼈 등 많은 해부학적 요소들이 변형을 겪었다.[314]
매우 다양한 공룡들 사이에서 공통적인 해부학적 특징을 찾을 수 있다. 대표적인 것으로는 공룡들이 모두 이궁류에 속한다는 점을 들 수 있는데, 공룡들은 단궁강에 속하는 동물들과는 달리 두개골에 있는 측두창이 둘로 나뉘어 있다.[325] 낮은 턱 뼈도 공룡의 공통적인 해부학적 특징으로 꼽을 수 있다.[326] 이외에도 초기 공룡으로부터 파생된 여러 골격 특징들이 있는데 길쭉한 견갑골, 세 개 이상의 척추뼈로 이루어진 만골[314], 하나의 구멍에 직결된 고관절을 이루는 관골구 등이 그것이다.[327]
스털링 네스빗(Sterling Nesbitt)의 아르코사우루스류 상호 관계에 대한 상세한 평가에서는 다음과 같은 열두 가지 명확한 공유파생형질을 확인하거나 발견했는데, 이 중 일부는 이전에 알려져 있었다.
네스빗은 몇 가지 추가적인 잠재적 공유파생형질을 발견했고, 이전에 제안된 공유파생형질의 수를 줄였다. 이러한 특징 중 일부는 실레사우루스과에도 존재하는데, 네스빗은 이들을 공룡류의 자매 그룹으로 복원했으며, 여기에는 큰 전방 전자, 길이가 비슷한 중족골 II와 IV, 좌골과 치골 사이의 접촉 감소, 경골의 경골능선과 거골의 상승 돌기의 존재 등이 포함된다.
공룡은 다양한 다른 골격 특징을 공유한다. 그러나 다른 아르코사우루스류 그룹에 공통적이거나 모든 초기 공룡에 존재하지 않았기 때문에, 이러한 특징은 공유파생형질로 간주되지 않는다. 예를 들어, 이궁류로서 공룡은 조상적으로 두 쌍의 관자놀이창 (눈 뒤의 두개골의 구멍)을 가지고 있었고, 이궁류 그룹 아르코사우루스류의 구성원으로서 주둥이와 아래턱에 추가적인 구멍을 가지고 있었다. 또한, 한때 공유파생형질로 여겨졌던 몇 가지 특징은 이제 공룡보다 먼저 나타났거나, 초기 공룡에는 없었고 서로 다른 공룡 그룹에 의해 독립적으로 진화한 것으로 알려져 있다. 여기에는 길쭉한 견갑골, 또는 어깨뼈; 세 개 이상의 융합된 척추로 구성된 천골 (다른 일부 아르코사우루스류에서는 세 개가 발견되지만, ''헤레라사우루스''에서는 두 개만 발견됨); 및 안쪽 표면 중심에 구멍이 있는 천공된 비구, 또는 엉덩이 소켓 (예를 들어, ''사투르날리아 투피니쿰''에서는 닫혀 있음)이 포함된다.[19] 공룡적인 특징을 결정하는 또 다른 어려움은 트라이아스기 후기의 초기 공룡과 다른 아르코사우루스류가 종종 잘 알려져 있지 않고 여러 면에서 유사하다는 점이다; 이 동물들은 때때로 문헌에서 잘못 식별되었다.
공룡은 대부분의 현대 포유류와 유사한 방식으로 뒷다리를 직립으로 세우지만, 다리가 양쪽으로 벌어진 대부분의 다른 파충류와는 다르다. 이러한 자세는 골반에 측면을 향하는 함몰부(일반적으로 열린 소켓)와 대퇴골에 해당되는 안쪽을 향하는 독특한 머리의 발달 때문이다. 그들의 직립 자세는 초기 공룡이 움직이는 동안 쉽게 호흡할 수 있게 하여, 캐리어의 제약에 따라 "벌어진" 파충류의 지구력과 활동 수준을 넘어설 수 있게 했다. 직립 다리는 또한 다리에 가해지는 굽힘 응력을 줄여 큰 크기의 진화를 지원하는 데 도움이 되었을 것이다. 라우이수치아를 포함한 일부 비공룡 아르코사우루스류도 직립 다리를 가지고 있었지만, 엉덩이 관절의 "기둥 직립" 구성을 통해 이를 달성했는데, 이는 대퇴골에서 돌출부가 엉덩이의 소켓에 삽입되는 대신, 상부 골반뼈가 회전하여 돌출된 선반을 형성한다.
6. 평균 크기
공룡의 지리적 분포는 물론 모양, 행동 및 크기는 급격하게 변화하였다. 따라서 "평균 크기"를 정확하게 정의하기는 쉽지 않아, 연구자마다 다양한 값을 제안하고 있다.[328]
빌 에릭슨에 따르면 무게가 500kg과 5톤 사이로 추정되는 공룡 가운데 바이오매스로 측정했을 때 80%는 미국 서부의 모리슨층에서 발견된다. 스테고사우루스와 용각류 등 평균 20톤 이상인 공룡은 오늘날의 일반적인 대형 포유류와 비교되지 않을 정도로 거대하다. 니콜라스 호튼은 공룡 63세대에 따라, 395.9 킬로그램(회색곰의 중량)을 초과하는 평균 질량, 그리고 일반 대중 중간값으로 2톤(기린과 비교)을 제시했다. 이는 평균 무게 631g인 멸종 포유류(788세대)와 크게 대조되며(작은 설치류), 현재까지 발견된 가장 작은 공룡의 3분의 2 이상의 크기였다. 현존하는 모든 포유동물과 비교했을 때, 공룡의 거의 대부분은 개인의 2%를 제외한 모든 살아있는 포유동물보다 컸다.[329]
평균적으로 크기가 컸던 공룡으로는 아르헨티노사우루스, 세이스모사우루스가 있고, 육식공룡 중에서는 스피노사우루스, 티라노사우루스, 기가노토사우루스, 수코미무스가 있다.
7. 공룡의 종류
분류학에서 공룡은 "트리케라톱스와 근생 조류 및 이들에서 분기된 모든 자손을 아우르는 가장 최근의 공통 조상이 속해 있는 부류"로 정의된다.[314] 이는 메갈로사우루스와 이구아노돈의 공통 조상이 곧 모든 공룡의 최근 공통 조상이라는 것을 의미한다.[315]
고생물학자들은 새가 공룡의 일종에서 진화한 것으로 보고 있으며, 새와 공룡을 같은 분지로 다룬다. 고생물학에서는 새가 용반목 수각아목 마니랍토라의 일종인 코에루로사우리아에 속한 조상으로부터 진화한 것으로 파악하고 있다.[316]
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 공룡과 같은 시기에 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다.
공룡은 매우 다양한 종류가 있었으며, 2006년까지 밝혀진 바로는 500여 속에 이른다. 이 중에 골격 화석이 발견된 것은 약 75% 정도이다.[321] 2008년 10월 17일 현재 확인된 공룡의 종은 1047개이다.[323] 새를 포함하면 그 다양성은 증가한다.
공룡은 생김새, 크기, 식성, 행동 양식 등이 매우 다양했다. 초식 공룡과 육식 공룡이 모두 있었으며, 2족 보행을 하거나 4족 보행을 하였다. 플라테오사우루스나 이구아노돈처럼 2족 보행과 4족 보행을 모두 할 수 있는 공룡도 있었다. 거대한 크기의 공룡도 있었지만, 많은 종류의 공룡은 인간과 유사한 크기이거나 그보다 작았다. 여러 종류의 공룡들이 뿔, 등딱지, 골갑, 볏 등을 몸을 보호하기 위한 수단으로 가지고 있었다. 공룡은 남극을 포함한 지구 어디에서나 서식하였다.[324]
전통적으로 공룡은 조반목(鳥盤目)과 용반목(龍盤目)의 두 종류로 분류되며, 세부적으로 용각아목, 수각아목, 곡룡류, 검룡류, 각룡류, 조각류 등으로 나뉜다.
7. 1. 조반류
조반목은 크게 각각아목과 장순아목[330]으로 나뉜다. 모든 조반목은 나무나 풀을 뜯어먹고 살았던 초식 공룡으로 입에는 부리처럼 생긴 뼈가 발달했으며, 대부분 몸에 골판(骨板)이 붙어 있었다. 허리는 조류처럼 치골이 좌골에 평행하며, 뒤쪽을 향하고 있었다. 잎 모양의 치관을 가진 초식동물로 어떤 것은 앞니가 없으나 대체로 강력한 어금니를 가졌다. 또한 척추를 뻣뻣하게 하는 힘줄도 가졌다. 백악기에는 조반목이 가장 중요한 초식공룡이었다. 현생 조류하고는 분류학상 무관하다.전통적으로 공룡은 조반목(鳥盤目)과 용반목(龍盤目)의 두 종류로 분류된다. 전문가들은 이들 공룡이 헤레라사우루스라는 훨씬 초기 공룡으로부터 진화했다고 믿고 있다. 시간이 지나면서 조반목의 동물이 용반목의 동물의 수를 넘어섰고 새로운 공룡들이 두 집단에서 진화되어 나왔다. 이들 새로운 공룡들은 생존에 보다 잘 적응하여 먹이를 얻고 적으로부터 도망치는 데 익숙했다. 용반류는 도마뱀의 골반을 닮았고, 조반류는 새의 골반을 닮았다.
7. 2. 용반류
용반목은 수각아목과 용각아목으로 나뉘며, 파충류처럼 장골, 치골, 좌골이 세 방향을 나타내고 있으며, 좌골에서 치골이 앞으로 향하는 것으로 식별된다.[292] 용반목에는 크기가 크거나 육식인 공룡들이 포함된다. 용반목에 속하는 수각류는 현생 조류의 진정한 조상이다.| 구분 | 속 | |
|---|---|---|
| 용반목 | 수각류 | 알로사우루스 티라노사우루스 데이노니쿠스 오르니토미무스 콤프소그나투스 |
| 용각류 | 플라테오사우루스 울트라사우루스 아파토사우루스 디플로도쿠스 브라키오사우루스 | |
분지학적 관점에서 "현생 조류와 트리케라톱스(''Triceratops'')를 포함하는 그룹의 가장 가까운 공통 조상에서 분기된 모든 자손"이 공룡의 정의로 자주 사용된다.[224][222]
현생 조류는 용반류의 수각류에 포함되고, 트리케라톱스는 조반류의 한 속이다. 즉, "용반류와 조반류 각 동물의 공통 조상에서 분기된 모든 것"이 공룡의 정의이며, "현생 조류"와 "트리케라톱스"는 각각 용반류와 조반류를 대표하는 예시로 언급된 것일 뿐이다.[223][225]
이러한 정의에 따르면, 수각류의 한 무리인 조류는 필연적으로 공룡(더 자세히는 용반류 중 수각류, 코엘루로사우리아에 속하는 마니랍토라)에 포함된다.
8. 공룡의 생활
트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 어룡(어룡목), 모사사우루스(유린목), 플레시오사우루스(사경룡목)는 수중에서, 익룡(익룡목)은 하늘에서 서식했다. 페름기에 살았던 디메트로돈은 단궁강 반룡목에 속하며 포유류와 연관이 깊다.[318] 쥐라기와 백악기에 융성하였으며, 당시 설치류가 주를 이루었던 포유류는 지금의 고양이만한 크기가 일반적이었다.[319] 삼돌기치목 레페노마무스는 예외적으로 몸무게가 12 - 14 kg에 달해 어린 프시타코사우루스를 사냥했다.[320]
2006년까지 500여 속, 2008년 10월 17일 현재 1047종이 확인되었다. 골격 화석이 발견된 것은 약 75% 정도이다.[321] 초기 연구의 3400여 속 추정은 오늘날 화석 기록과 차이가 있다.[322] 새를 포함하면 그 다양성은 증가한다.
생김새, 크기, 식성, 행동 양식은 천차만별이었다. 초식, 육식, 2족/4족 보행, 플라테오사우루스나 이구아노돈처럼 2족/4족 보행을 모두 하는 경우도 있었다. 대부분 인간과 유사하거나 작았지만 거대한 종류도 있었다. 뿔, 등딱지, 골갑, 볏 등으로 몸을 보호하기도 했다. 남극을 포함한 지구 어디에서나 서식했다.[324]
20종이 넘는 공룡이 같은 시기, 지역에서 함께 살았다. 대부분 목초지에서 무리를 이루었으며, 아파토사우루스나 티라노사우루스는 평생을 혼자 또는 작은 무리로 지냈다. 곡룡류·각룡류·검룡류는 바닷가나 탁 트인 곳에서, 알로사우루스와 티라노사우루스 등 커다란 수각류는 거대한 초식공룡을, 일부 작은 수각류는 곤충, 알, 포유류, 작은 공룡, 파충류를 잡아먹었다. 작은 수각류는 활동적이고 재빨랐다.
번식 방법은 확실하지 않으나, 공룡알 화석으로 보아 다른 파충류처럼 알을 낳았을 것으로 추측된다. 암컷은 땅에 둥지를 파고 12~24개의 알을 낳았으며, 새끼를 돌보거나, 스스로 살아가도록 내버려두는 경우도 있었다.
포유류와 달리 살아있는 동안 계속 자라는 것으로 짐작되었지만, 티라노사우루스의 생장곡선 등으로 보아 일정 기간 자란 다음 성장이 멈추는 것으로 알려진다. 수명은 공룡마다 다르지만, 대부분 온혈동물이었으므로 100년 이내로 살았을 것으로 보인다. 티라노사우루스 렉스의 수명은 30년이다.
9. 새를 제외한 공룡의 멸종
과학자들은 새를 제외한 모든 공룡의 멸종을 설명하는 많은 가설을 세웠다. 확실한 것은 백악기 말에 지구에 일어난 변화에 그들이 적응하지 못해 멸종했다는 것이다.
모든 비조류 공룡과 대부분의 조류 계통은 백악기 말에 일어난 대량 멸종 사건인 백악기-고생대(K-Pg) 멸종 사건으로 멸종되었다. 6,603만 8천 ± 2만 5천 년 전으로 추정되는 백악기-고생대 경계 상층에서 비조류 공룡의 화석은 갑자기 사라진다.[160] 이러한 대량 멸종을 야기한 사건은 1970년대부터 광범위하게 연구되었으며, 유카탄 반도의 외계 충돌 사건과 인도에서의 홍수 현무암 화산 활동이 주요 원인으로 여겨진다. 그러나 멸종 사건의 구체적인 메커니즘과 공룡에 미치는 영향은 여전히 연구 중이다.[161] 공룡과 함께 익룡, 모사사우루스 및 플레시오사우루스와 같은 해양 파충류, 여러 그룹의 포유류, 암모나이트(앵무조개와 같은 연체동물), 루디스트(산호초를 만드는 이매패류), 다양한 해양 플랑크톤 그룹이 멸종되었다.[167] 총 47%의 속과 지구상의 종의 76%가 K-Pg 멸종 사건 동안 멸종되었다.[162] 대부분의 공룡이 비교적 큰 크기를 가지고 있었고 백악기 말에 소형 공룡 종의 다양성이 낮았던 점이 멸종에 기여했을 수 있다.[163]
볼라이드 충돌 가설은 1980년 월터 알바레즈, 루이스 알바레즈와 동료들에 의해 처음으로 널리 알려졌으며, K-Pg 멸종 사건을 볼라이드(외계 발사체) 충돌로 돌린다.[177] 알바레즈와 동료들은 백악기-고생대 경계 암석 퇴적물에서 전 세계적으로 기록된 이리듐 수치의 급격한 증가는 충돌의 직접적인 증거라고 제안했다. 앨런 힐데브랜드와 동료들의 1991년 논문에서 멕시코 남동부 유카탄 반도에 있는 충돌구가 발견, 공개되면서 충돌 지점이 확인되었다.[179] 현재 대부분의 증거는 너비 5km 에서 15km인 볼라이드가 6600만 년 전에 유카탄 반도에 충돌하여 이 충돌구를 형성하고, 멸종 사건을 촉발하는 "킬 메커니즘"을 생성했음을 시사한다.[180][181][182]
칙술루브 충돌은 몇 시간 안에 지진,[183] 쓰나미,[184] 전 세계적인 화재 폭풍과 같은 즉각적인 영향을 미쳤을 것이다.[185][186] 며칠 안에 충돌 지점 암석에서 방출된 황산염 에어로졸은 산성비와 해양 산성화에 기여했을 것이다.[187][190] 그을음 에어로졸은 수개월에서 수년에 걸쳐 전 세계로 퍼져, 열 복사를 반사하여 지구 표면을 냉각시키고, 햇빛을 차단하여 광합성을 크게 늦춰 충격 겨울을 초래했을 것이다.[161][188][189] 광합성 중단은 곡물을 먹는 새를 제외한 모든 공룡을 포함, 잎이 많은 식물에 의존하는 먹이 사슬의 붕괴로 이어졌을 것이다.[165]
K-Pg 대멸종 시기에 인도의 데칸 트랩(Deccan Traps) 대규모 현무암 분출이 활발하게 일어났다. 분출은 K-Pg 경계 부근을 기준으로 세 단계로 나눌 수 있는데, 두 단계는 경계 이전에, 한 단계는 이후에 발생했다. 경계와 매우 가까운 시기에 발생한 두 번째 단계는 이 분출량의 70~80%를 간헐적인 펄스로 분출했으며, 이는 약 10만 년 간격으로 발생했다.[191][192] 이 화산 활동으로 이산화 탄소와 이산화 황과 같은 온실 가스가 방출되었고,[193][194] 이로 인해 약 3°C의 온도 변화, 또는 최대 7°C에 달하는 기후 변화가 발생했을 것이다.[195] 칙술루브 충돌과 마찬가지로, 분출은 산성비를 유발하고 지구를 냉각시키는 황산염 에어로졸을 방출했을 수도 있다.[196] 그러나 분출 시기의 불확실성이 커서, K-Pg 대멸종에서 데칸 트랩의 역할은 여전히 불분명하다.[160][161]
월터 알바레스 자신도 칙술루브 충돌 외에 데칸 트랩과 다른 생태학적 요인들이 멸종에 기여했을 수 있음을 인정했다. 일부 추정에서는 데칸 트랩 분출의 두 번째 단계가 칙술루브 충돌 이후 5만 년 이내에 시작되었다고 보고 있다.[200]
비조류 공룡의 유해가 K-Pg 경계 위에서 가끔 발견된다. 2000년, 스펜서 루카스와 동료들은 뉴멕시코의 산 후안 분지에서 단일 하드로사우루스 오른쪽 대퇴골을 발견했다고 보고했으며, 이를 고생대 공룡의 증거로 묘사했다. 뼈가 발견된 암석층은 약 6480만 년 전의 초기 고생대 시기로 연대가 측정되었다. 그러나 이러한 고생대 공룡으로 추정되는 것들은 많은 다른 연구자들에 의해 재작업된, 즉 원래 위치에서 씻겨 나와 더 젊은 퇴적물에 다시 묻힌 것으로 여겨진다.
10. 공룡의 화석
과학자들은 공룡의 뼈, 이, 알, 발자국 등의 화석을 연구하여 공룡에 대해 많은 것을 알아냈다. 19세기 후반에서 20세기 초반 사이에 북아메리카 서부, 유럽, 아시아, 아프리카 등지에서 공룡 화석이 대규모로 발견되었다. 공룡 화석 산지로는 미국의 유타주, 동아시아의 몽골, 유럽의 벨기에 등이 유명하다.[85]
미국의 애리조나주나 유타주(특히 클리블랜드)에서는 대형 공룡 발자국 화석이 많이 발견되었다. 중국 북동부 지방에서는 소형이면서 두 다리로 걸어 다닌 열하공룡의 발자국이 64m2에 4,000개 이상 같은 방향을 향하고 있어, 이들이 당시에 동쪽으로 대이동을 하였음을 보여준다.[85]
공룡 화석은 수천 년 동안 알려져 왔지만, 그 진정한 정체는 인식되지 못했다. 중국인들은 그것들을 용의 뼈로 여기고 기록했다. 예를 들어, 서진 시대(265–316)에 창거가 편찬한 지리지인 화양국지에는 쓰촨성 우청에서 용의 뼈가 발견되었다고 보고했다. 중국 중부의 마을 사람들은 오랫동안 화석화된 "용의 뼈"를 전통 의학에 사용하기 위해 발굴해왔다. 유럽에서는 공룡 화석이 일반적으로 거인과 다른 성경 속 생물의 유해로 여겨졌다.[20]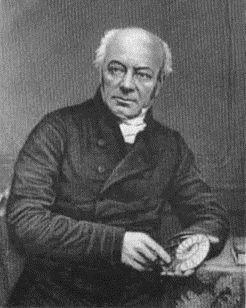
현재 공룡 뼈로 인정받는 것에 대한 학술적 설명은 17세기 후반 영국에서 처음 등장했다. 1676년 옥스퍼드셔주 콘웰 근처 치핑 노턴의 석회암 채석장에서 현재 ''메갈로사우루스''의 대퇴골로 알려진 뼈의 일부가 발견되었다. 애시몰린 박물관의 초대 큐레이터이자 옥스퍼드 대학교 화학 교수인 로버트 플롯은 자신의 저서 ''옥스퍼드셔의 자연사''(1677)에서 이 뼈가 큰 동물의 대퇴골 하단부라는 것을 정확히 확인했으며, 알려진 어떤 종에도 속하지 않을 정도로 크다고 설명했다.[21] 그는 이 뼈가 전설에 등장하는 티탄이나 다른 종류의 거인일 가능성이 있는 거대한 인간의 대퇴골이라고 결론지었다.[22][23] 아이작 뉴턴 경의 친구인 에드워드 류이드는 ''리소필락시 브리타니키 이크노그라피아''(1699)에서 옥스퍼드셔주 위트니 근처의 카스웰에서 발견된 용각류 이빨 "루텔룸 임피카툼"을 묘사하고 이름을 붙였다.
1815년에서 1824년 사이에 옥스퍼드 대학교 지질학 첫 번째 강사인 윌리엄 버클랜드 목사는 ''메갈로사우루스''의 화석화된 뼈를 더 수집했으며, 과학 저널에 조류가 아닌 공룡을 처음으로 묘사했다. 1822년 영국의 지질학자 기데온 맨텔의 아내인 메리 앤 맨텔이 ''이구아노돈''을 발견했다고 전해지지만, 기데온이 수년 전에 유해를 입수했다는 주장도 있다. 기데온 맨텔은 자신의 화석과 현대 이구아나의 뼈 사이의 유사성을 인식하고 1825년에 자신의 연구 결과를 발표했다.
이러한 "거대한 화석 도마뱀"에 대한 연구는 유럽과 미국의 과학자들에게 큰 관심사가 되었고, 1842년 영국의 고생물학자 리처드 오웬 경은 "사우리아 파충류의 특정한 종족 또는 하위 분류"를 지칭하기 위해 "공룡"이라는 용어를 만들었다.[7][8][9][24] 오웬은 ''이구아노돈'', ''메갈로사우루스'', ''힐라에오사우루스''가 뚜렷한 특징을 공유한다는 것을 인식하고 이를 별개의 분류군으로 제시했다. 그는 1841년 영국 과학 진흥 협회에서 화석 파충류에 대한 발표를 했지만, "공룡"이라는 단어를 언급하거나 공룡을 파충류의 별개의 그룹으로 인식하지 않았다. 그는 1842년 4월에 발표된 강연의 수정된 텍스트 버전에서만 공룡을 소개했다.[7][8] 앨버트 공과 빅토리아 여왕의 남편의 지원을 받아 오웬은 공룡 화석 및 기타 생물학적, 지질학적 전시물을 전시하기 위해 런던 자연사 박물관을 설립했다.[25]
10. 1. 대한민국의 공룡 화석
1973년 경상북도 의성군 금성면 탑리의 경상 누층군 사곡층에서 공룡 뼈 화석이 발견된 이후, 1977년부터 '공룡 골짜기'에 대한 본격적인 발굴이 시작되어 많은 공룡 화석이 발굴되었다.[85] 1983년 경상남도 하동군 금남면의 경상 누층군 하산동층에서는 조반류의 알껍질 화석이, 고성군 하이면 덕명리의 경상 누층군 진동층에서는 약 360개의 발자국 화석이 발견되었다. 이 발자국은 거의 대부분 조반류의 두 다리로 걸은 공룡의 것이나, 일부 용반류의 것과 네 다리로 걸은 공룡의 것도 있다.[85] 그 밖에 경남 고성의 해안 및 함안, 통영, 울산 등지에서 수천 개나 되는 공룡 발자국 화석이 발견되었으며 합천, 진주, 군위 등지에서도 크고 작은 초식공룡과 육식공룡 등의 화석이 발견되어, 앞으로 대한민국에서도 공룡의 완전한 뼈대 화석이 발견될 날이 있을 것으로 기대된다.[85]10. 2. 일본의 공룡 화석
일본에서도 대한민국과 마찬가지로 공룡 화석이 많이 발굴되었다. 홋카이도(나카가와정, 오비라정, 유바리시), 이와테현 이와이즈미정, 후쿠시마현(미나미소마시, 이와키시, 히로노정), 군마현 간나정, 나가노현 오타리촌, 도야마현 도야마시, 기후현(시라카와정, 다카야마시), 이시카와현(하쿠산시), 후쿠이현(가쓰야마시, 오노시), 미에현 도바시, 효고현(단바시, 스모토시), 도쿠시마현 가쓰우라정, 야마구치현 시모노세키시, 후쿠오카현(기타큐슈시, 미야와카시), 구마모토현(미후네정, 아마쿠사시)에서 공룡 화석이 발견되었다.[296][297][298][299][300][301][302][303][304][305][306][307][308][309][310]| 발견 지역 | 공룡 종류 | 비고 |
|---|---|---|
| 홋카이도 나카가와정 | 파라리테리지노사우루스 자포니쿠스(테리지노사우루스류) | [297][298] |
| 홋카이도 아시베쓰시 | 티라노사우루스류 | |
| 홋카이도 고시로정 | 하드로사우루스류의 뒷다리 | |
| 홋카이도 유바리시 | 노도사우루스류의 두개골 일부 | |
| 홋카이도 무카와정 | 하드로사우루스과, 카무이 사우루스 자포니쿠스(무카와룡) | [299] |
| 이와테현 이와이즈미정 | 용각류(통칭 모시류) | |
| 후쿠시마현 미나미소마시 | 발자국(쥐라기) | |
| 후쿠시마현 히로노마치 | 조각류, 티라노사우루스류(통칭 후타바류) | [300] |
| 후쿠시마현 이와키시 | 용각류, 조각류 | |
| 군마현 간나정 | 스피노사우루스류의 이빨, 오르니토무스류(통칭 산추류), 수각류, 발자국 | |
| 나가노현 오타니무라 | 발자국(쥐라기) | |
| 도야마현 도야마시 | 이구아노돈류의 이빨, 수각류, 발자국 | |
| 기후현 시라카와정 | 용각류, 발자국 | |
| 기후현 다카야마시 | 이구아노돈류, 힙실로포돈류의 이빨, 조각류, 수각류 | |
| 이시카와현 하쿠산시 | 용각류(통칭 하쿠산류), 수각류(통칭 오비랍토르류), 이구아노돈류(통칭 시마류), 알바레즈사우루스 야마구치오로움 (각각류), 티라노사우루스류(통칭 카가류), 발자국 | [301][302] |
| 후쿠이현 가쓰야마시 | 후쿠이 사우루스 테트리엔시스(이구아노돈류), 후쿠이라프토르 키타다니엔시스(메가랩터류), 후쿠이티탄 닛포넨시스(용각류), 코시 사우루스 카츠야마(이구아노돈류), 후쿠이베나토르 파라독서스(코엘로사우루스류), 후쿠이프테릭스 프리마 (조류), 티라노미무스 후쿠이엔시스 (오르니토미모사우루스류), 수각류, 조각류, 발자국 | [303][304][305][306][307] |
| 후쿠이현 오노시 | 수각류의 이빨, 발자국 | |
| 미에현 토바시 | 티타노사우루스류(통칭 토바류), 발자국 | |
| 효고현 단바시.단바사사야마시 | 단바티타니스 아미키티애 (티타노사우루스형류), 힙노베나토르 마츠바라에토오에오룸 (코엘로사우루스류), 사사야마그노무스 사에구사이 (신각룡류), 각룡류, 티라노사우루스류, 검룡류 등 | |
| 효고현 스모토시 | 야마토사우루스 이자나기이(하드로사우루스류) 람베오사우루스류 | |
| 도쿠시마현 가쓰우라정 | 이구아노돈류의 이빨 | |
| 야마구치현 시모노세키시 요시모 | 발자국 | |
| 후쿠오카현 기타큐슈시 | 수각류 | |
| 후쿠오카현 미야와카시 | 수각류 (와키노사우루스 사토이) | [308] |
| 나가사키현 나가사키시 | 수각류 | [309] |
| 구마모토현 미후네마치 | 티라노사우루스류, 오르니토미무스류, 테리지노사우루스류, 드로마에오사우루스류, 안킬로사우루스류, 하드로사우루스류, 미후네류, 발자국 | |
| 구마모토현 아마쿠사시 | 수각류, 이구아노돈류, 조각류, 용각류, 발자국 | |
| 구마모토현 야츠시로시 | 수각류(알로사우루스형류), 늑골 | [310] |
| 가고시마현 사쓰마센다이시 | 수각류의 이빨, 이구아노돈류의 이빨, 각룡류의 이빨 |
11. 진화사
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다.[318] 공룡은 중생대에 크게 번성하였으며 특히 쥐라기와 백악기에 융성하였다.
매우 다양한 종류의 공룡이 존재하였으며 2006년까지 밝혀진 바로는 500여 속에 이른다.[321] 2008년 10월 17일 현재 확인된 공룡의 종은 1047개이다.[323] 새를 포함하면 그 다양성은 증가한다.
공룡의 생김새, 크기, 식성, 행동 양식 등은 천차만별이었다. 초식, 육식, 2족 보행, 4족 보행 등 다양한 형태가 있었으며, 플라테오사우루스나 이구아노돈처럼 2족 보행과 4족 보행을 모두 할 수 있는 공룡도 있었다. 거대한 크기의 공룡이 있었는가 하면 일반적으로 알려진 것과는 달리 많은 종류의 공룡이 인간과 유사한 크기이거나 그보다 작았다. 여러 종류의 공룡들이 자신의 몸을 보호하기 위한 뿔, 등딱지, 골갑, 볏 등을 갖고 있었다. 공룡은 지구 어디에서나 서식하였으며 남극에도 있었다.[324]
트라이아스기 이후 공룡의 진화는 식생의 변화와 대륙의 위치 변화에 따라 진행되었다. 후기 트라이아스기와 초기 쥐라기에는 대륙이 판게아로 연결되어 있었으며, 코엘로피시스상과 육식 공룡과 초기 용각류 초식 공룡으로 구성된 전 세계적인 공룡 동물군이 존재했다.
중기 및 후기 쥐라기까지 공룡 동물군의 일반적인 균일성은 계속되었으며, 대부분의 지역에는 세라토사우루스류, 메갈로사우루스상과, 알로사우루스상과로 구성된 포식자와 검룡류 조반류와 대형 용각류로 구성된 초식 공룡이 있었다. 중국의 공룡은 특화된 메트리아칸토사우루스과 수각류와 ''마멘키사우루스''와 같은 특이하고 목이 긴 용각류가 존재했다.
초기 백악기와 판게아의 계속되는 분열로 인해 공룡은 대륙별로 분화되었다. 아시아에서는 드로마에오사우루스과, 트로오돈과, 오비랍토르류와 같은 마니랍토라 코엘루로사우루스류가 흔한 수각류가 되었고, ''프시타코사우루스''와 같은 안킬로사우루스과와 초기 각룡류가 중요한 초식 공룡이 되었다. 호주에는 기저 안킬로사우루스류, 힙실로포돈류, 이구아노돈류의 동물군이 서식했다.
후기 백악기에는 세 가지 일반적인 공룡 동물군이 존재했다. 북아메리카와 아시아의 북부 대륙에서는 주요 수각류가 티라노사우루스과와 다양한 종류의 더 작은 마니랍토란 수각류였으며, 주로 하드로사우루스과, 각룡류, 안킬로사우루스과, 파키케팔로사우루스류로 구성된 조반류 초식 공룡 무리가 있었다. 현재 분열되고 있는 초대륙 곤드와나를 구성했던 남부 대륙에서는 아벨리사우루스과가 흔한 수각류였고, 티타노사우루스류 용각류가 흔한 초식 공룡이었다. 유럽에서는 드로마에오사우루스과, 랍도돈과 이구아노돈류, 노도사우루스과 안킬로사우루스류, 티타노사우루스류 용각류가 널리 퍼져 있었다.
약 6,600만 년 전 백악기 말에 발생한 백악기-고생대 멸종 사건으로 인해 조류를 제외한 모든 공룡 그룹이 멸종되었다.
1990년대 이후, 중국의 백악기 지층에서 깃털을 가진 공룡 화석이 잇따라 발견되어, 현재의 조류와 깃털이 없는 공룡 사이의 계통 관계가 밝혀졌다. 깃털을 가진 공룡에는 시노사우롭테릭스, 프로타르케오프테릭스, 카우디프테릭스, 미크로랍토르, 딜롱 등이 있다.
"조류는 공룡의 수각류의 일종이다"라는 설이 거의 정설이 되었다.[265]
공룡이 처음 발견되었을 때, 파충류로서 변온 동물로 여겨졌다. 그러나 존 오스트롬과 로버트 T. 바커 등은 공룡이 항온 동물이라고 주장했다. 하지만 뇌 발달 정도, 뼈에 나이테가 보이는 점 등으로 인해 항상성을 부정하는 설도 존재했다. 그러나 뼈의 나이테는 초식 포유류에게도 존재한다는 것이 밝혀졌고, 이 점에서도 항상성을 부정하는 근거가 될 수 없다.[267][268] 에드윈 해리스 콜버트 등은 아메리카 악어의 총배설강에 온도계를 꽂아 체온을 측정하는 실험을 한 결과, 대형 개체일수록 체온 변화가 완만하다는 결론을 얻었다.[269] 따라서 대형 용각류 등에서는 부피가 크기 때문에 결과적으로 체온을 체내에 유지할 수 있는 '관성 항온성'으로 체온을 유지했다는 주장도 있다.
토마스 홀츠가 제시한 주요 공룡 분류군의 시간 범위는 아래 표와 같다.
| 분류군 | 시기 (백만 년 전) |
|---|---|
| 헤레라사우루스과 | 233.23 ~ 210 |
| 과이바사우루스과 | 231.4 ~ 208 |
| 플라테오사우루스과 | 225 ~ 190 |
| 리오하사우루스과 | 228 ~ 213 |
| 마소스폰딜루스과 | 227 ~ 176 |
| 불카노돈과 | 183 ~ 175 |
| 투리아사우루스류 | 168 ~ 125 |
| 세티오사우루스과 | 175 ~ 150 |
| 디플로도쿠스상과 | 174 ~ 93 |
| 브라키오사우루스과 | 157 ~ 93 |
| 티타노사우루스류 | 140 ~ 66 |
| 코엘로피시스상과 | 221 ~ 183 |
| 케라토사우루스류 | 199.3 ~ 66 |
| 메갈로사우루스상과 | 170 ~ 85 |
| 카르노사우루스류 | 175.6 ~ 88 |
| 메가랍토르류 | 130 ~ 66 |
| 티라노사우루스상과 | 166 ~ 66 |
| 콤프소그나투스과 | 151.5 ~ 108 |
| 오르니토미무스류 | 140 ~ 66 |
| 알바레즈사우루스류 | 160 ~ 66 |
| 테리지노사우루스류 | 139 ~ 66 |
| 오비랍토르류 | 130 ~ 66 |
| 데이노니쿠스류 | 167 ~ 66 |
| 새무리 | 155 ~ 현재 |
| 헤테로돈토사우루스과 | 200 ~ 140 |
| 스테고사우루스류 | 169 ~ 100 |
| 곡룡류 | 170.3 ~ 66 |
| 후두류 | 92 ~ 66 |
| 각룡류 | 161 ~ 66 |
| 조각류 | 164 ~ 66 |
11. 1. 기원과 초기 진화
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 공룡과 같은 시기에 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다.공룡은 중기에서 후기 트라이아스기에 그들의 고룡 조상으로부터 분화되었는데, 이는 약 2억 5,200만 년 전 페름기-트라이아스기 대멸종으로 해양 종의 약 96%와 육상 척추 동물의 70%가 멸종된 후 약 2천만 년 후이다. 실질적인 유해가 발견된 가장 오래된 공룡 화석은 트라이아스기 카르니아 시대에 속하며, 주로 아르헨티나와 브라질의 이스치과라스토와 산타 마리아 형성과 짐바브웨의 페블리 아코스 형성에서 발견되었다.[67]

이스치과라스토 형성(2억 3,100만 년에서 2억 3,000만 년으로 방사성 연대 측정됨)[68]에서 초기 사우루스류인 ''에오랩터''가 발견되었는데, 원래는 에레라사우루스과에 속하는 것으로 여겨졌으나[69] 현재는 초기 용각류로 여겨진다. ''에레라사우루스''와 ''산후안사우루스'', 그리고 용각류인 ''크로모기사우루스'', ''에오드로마에우스'', ''판파기아''와 함께 발견되었다.[70] 모든 공룡의 공통 조상과의 ''에오랩터''의 유사성은 최초의 공룡이 작고, 두 발로 걷는 포식자였을 것이라는 것을 시사한다.[71]
산타 마리아 형성(2억 3,323만 년으로 방사성 연대 측정되어 더 오래됨)[72]에서 에레라사우루스과인 ''그나토보락스''와 ''스타우리고사우루스'', 용각류인 ''바구알로사우루스'', ''부리올레스테스'', ''과이바사우루스'', ''마크로콜룸'', ''은다미림'', ''팜파드로마에우스'', ''사투르날리아'', ''우나이사우루스''가 발견되었다.[70] 연대가 불확실하지만 다른 두 곳과 유사했을 것으로 보이는 페블리 아코스 형성에서는 용각류인 ''미비레사우루스''와 이름 없는 에레라사우루스과가 발견되었다.[67]
''자클라팔리사우루스''와 ''남발리아''의 보존 상태가 좋지 않은 용각류 유해와 초기 사우루스류인 ''알왈케리아''가 인도의 상부 말레리 및 하부 말레리 형성에서 알려져 있다.[73] 카르니아 시대의 아르헨티나 차냐레스 형성에서는 ''라고수쿠스''와 ''라게르페톤''과 같은 원시적인 공룡형 조류를 보존하고 있는데, 이는 공룡 진화를 이해하는 데 또 다른 중요한 장소가 된다. 이러한 조류는 초기 공룡이 작고, 두 발로 걷는 포식자였다는 모델을 뒷받침한다.[70][74]
공룡은 트라이아스기 아니시안 시대, 즉 약 2억 4,300만 년 전부터 나타났을 수 있는데, 이는 탄자니아 만다 형성의 ''니아사사우루스''의 연대이다. 그러나 알려진 화석은 공룡인지 아니면 가까운 친척인지 식별하기에는 너무 파편적이다.[75] 만다 형성을 아니시안으로 분류하는 것 또한 불확실하다. 어쨌든 공룡은 공룡이 아닌 조류와 일정 기간 공존했는데, 추정치는 500만 년에서 1천만 년[76]에서 2,100만 년에 이른다.[72]
공룡이 나타났을 때, 그들은 육상에서 지배적인 동물은 아니었다. 육상 서식지는 고룡류와 수궁류와 같은 다양한 종류의 고룡류와 키노돈트 및 린코사우루스와 같은 수궁류가 차지하고 있었다. 그들의 주요 경쟁자는 공룡보다 더 성공적인 의사수키아로, 에토사우루스, 오르니토수키아 및 라우이수키아 등이 있었다.[77] 이러한 다른 동물들 대부분은 트라이아스기 동안 두 가지 사건 중 하나로 멸종되었다.
먼저, 약 2억 1,500만 년 전, 원시 고룡류, 원룡류를 포함하여 멸종되었다. 이어서 트라이아스기-쥐라기 멸종 사건(약 2억 100만 년 전)으로 에토사우루스, 오르니토수키아, 피토사우루스, 라우이수키아 등 다른 초기 고룡류 그룹의 대부분이 멸종되었다. 린코사우루스와 이치류는 적어도 노리안 시대 초중반과 노리안 시대 후기 또는 레티아 단 초기까지 살아남았으며,[78][79] 멸종의 정확한 시기는 불확실하다. 이러한 손실로 인해 악어류, 공룡, 포유류, 익룡, 거북이 남겨졌다. 초기 공룡의 첫 번째 몇몇 계통은 트라이아스기의 카르니아 및 노리안 단계 동안 적응 방산을 거쳐 멸종된 그룹의 틈새를 차지했을 가능성이 있다. 또한 주목할 만한 점은 카르니아 홍수 사건 동안 멸종률이 높아졌다는 것이다.[80]
홀츠가 제시한 주요 공룡 분류군의 시간 범위는 아래 표와 같다.
| 분류군 | 시기 (백만 년 전) |
|---|---|
| 헤레라사우루스과 | 233.23 ~ 210 |
| 과이바사우루스과 | 231.4 ~ 208 |
| 플라테오사우루스과 | 225 ~ 190 |
| 리오하사우루스과 | 228 ~ 213 |
| 마소스폰딜루스과 | 227 ~ 176 |
| 불카노돈과 | 183 ~ 175 |
| 투리아사우루스류 | 168 ~ 125 |
| 세티오사우루스과 | 175 ~ 150 |
| 디플로도쿠스상과 | 174 ~ 93 |
| 브라키오사우루스과 | 157 ~ 93 |
| 티타노사우루스류 | 140 ~ 66 |
| 코엘로피시스상과 | 221 ~ 183 |
| 케라토사우루스류 | 199.3 ~ 66 |
| 메갈로사우루스상과 | 170 ~ 85 |
| 카르노사우루스류 | 175.6 ~ 88 |
| 메가랍토르류 | 130 ~ 66 |
| 티라노사우루스상과 | 166 ~ 66 |
| 콤프소그나투스과 | 151.5 ~ 108 |
| 오르니토미무스류 | 140 ~ 66 |
| 알바레즈사우루스류 | 160 ~ 66 |
| 테리지노사우루스류 | 139 ~ 66 |
| 오비랍토르류 | 130 ~ 66 |
| 데이노니쿠스류 | 167 ~ 66 |
| 참새류 | 155 ~ 현재 |
| 헤테로돈토사우루스과 | 200 ~ 140 |
| 스테고사우루스류 | 169 ~ 100 |
| 곡룡류 | 170.3 ~ 66 |
| 후두류 | 92 ~ 66 |
| 각룡류 | 161 ~ 66 |
| 조각류 | 164 ~ 66 |
11. 2. 진화와 고생물지리학
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다.[318] 공룡은 중생대에 크게 번성하였으며 특히 쥐라기와 백악기에 융성하였다.매우 다양한 종류의 공룡이 존재하였으며 2006년까지 밝혀진 바로는 500여 속에 이른다.[321] 2008년 10월 17일 현재 확인된 공룡의 종은 1047개이다.[323] 새를 포함하면 그 다양성은 증가한다.
공룡의 생김새, 크기, 식성, 행동 양식 등은 천차만별이었다. 초식, 육식, 2족 보행, 4족 보행 등 다양한 형태가 있었으며, 플라테오사우루스나 이구아노돈처럼 2족 보행과 4족 보행을 모두 할 수 있는 공룡도 있었다. 거대한 크기의 공룡이 있었는가 하면 일반적으로 알려진 것과는 달리 많은 종류의 공룡이 인간과 유사한 크기이거나 그보다 작았다. 여러 종류의 공룡들이 자신의 몸을 보호하기 위한 뿔, 등딱지, 골갑, 볏 등을 갖고 있었다. 공룡은 지구 어디에서나 서식하였으며 남극에도 있었다.[324]
익룡, 모사사우루스, 어룡, 플레시오사우루스, 디메트로돈은 공룡으로 오인되기도 하지만, 분류학적으로 공룡에 속하지 않는다.
공룡은 특히 쥐라기와 백악기 시대에 중생대의 지배적인 육상 척추 동물이었다. 2018년에는 900개 이상의 비조류 공룡 속이 확인되었으며, 2016년에는 1124종이 확인되었다.[13][14] 화석 기록에 보존된 공룡 속의 총 수는 1850개로 추정되며, 거의 75%가 아직 발견되지 않았다. 2016년 추정치는 중생대에 살았던 공룡 종의 수를 1,543~2,468종으로 추정했다.[15][16]
트라이아스기 이후 공룡의 진화는 식생의 변화와 대륙의 위치 변화에 따라 진행되었다. 후기 트라이아스기와 초기 쥐라기에는 대륙이 판게아로 연결되어 있었으며, 코엘로피시스상과 육식 공룡과 초기 용각류 초식 공룡으로 구성된 전 세계적인 공룡 동물군이 존재했다.
중기 및 후기 쥐라기까지 공룡 동물군의 일반적인 균일성은 계속되었으며, 대부분의 지역에는 세라토사우루스류, 메갈로사우루스상과, 알로사우루스상과로 구성된 포식자와 검룡류 조반류와 대형 용각류로 구성된 초식 공룡이 있었다. 중국의 공룡은 특화된 메트리아칸토사우루스과 수각류와 ''마멘키사우루스''와 같은 특이하고 목이 긴 용각류가 존재했다.
초기 백악기와 판게아의 계속되는 분열로 인해 공룡은 대륙별로 분화되었다. 아시아에서는 드로마에오사우루스과, 트로오돈과, 오비랍토르류와 같은 마니랍토란 코엘루로사우루스류가 흔한 수각류가 되었고, ''프시타코사우루스''와 같은 안킬로사우루스과와 초기 각룡류가 중요한 초식 공룡이 되었다. 호주에는 기저 안킬로사우루스류, 힙실로포돈류, 이구아노돈류의 동물군이 서식했다.
후기 백악기에는 세 가지 일반적인 공룡 동물군이 존재했다. 북아메리카와 아시아의 북부 대륙에서는 주요 수각류가 티라노사우루스과와 다양한 종류의 더 작은 마니랍토란 수각류였으며, 주로 하드로사우루스과, 각룡류, 안킬로사우루스과, 파키케팔로사우루스류로 구성된 조반류 초식 공룡 무리가 있었다. 현재 분열되고 있는 초대륙 곤드와나를 구성했던 남부 대륙에서는 아벨리사우루스과가 흔한 수각류였고, 티타노사우루스류 용각류가 흔한 초식 공룡이었다. 유럽에서는 드로마에오사우루스과, 랍도돈과 이구아노돈류, 노도사우루스과 안킬로사우루스류, 티타노사우루스류 용각류가 널리 퍼져 있었다.
약 6,600만 년 전 백악기 말에 발생한 백악기-고생대 대멸종 사건으로 인해 네오르니테스 조류를 제외한 모든 공룡 그룹이 멸종되었다.
1990년대 이후, 중국의 백악기 지층에서 깃털을 가진 공룡 화석이 잇따라 발견되어, 현재의 조류와 깃털이 없는 공룡 사이의 계통 관계가 밝혀졌다. 깃털을 가진 공룡에는 시노사우롭테릭스, 프로타르케오프테릭스, 카우디프테릭스, 미크로랍토르, 딜롱 등이 있다.
"조류는 공룡의 수각류의 일종이다"라는 설이 거의 정설이 되었다.[265]
공룡이 처음 발견되었을 때, 파충류로서 변온 동물로 여겨졌다. 그러나 존 오스트롬과 로버트 T. 바커 등은 공룡이 항온 동물이라고 주장했다.
하지만 뇌 발달 정도, 뼈에 나이테가 보이는 점 등으로 인해 항상성을 부정하는 설도 존재했다. 그러나 뼈의 나이테는 초식 포유류에게도 존재한다는 것이 밝혀졌고, 이 점에서도 항상성을 부정하는 근거가 될 수 없다.[267][268]
에드윈 해리스 콜버트 등은 아메리카 악어의 총배설강에 온도계를 꽂아 체온을 측정하는 실험을 한 결과, 대형 개체일수록 체온 변화가 완만하다는 결론을 얻었다.[269] 따라서 대형 용각류 등에서는 부피가 크기 때문에 결과적으로 체온을 체내에 유지할 수 있는 '관성 항온성'으로 체온을 유지했다는 주장도 있다.
토마스 홀츠의 주요 공룡 분류군의 시간 범위 표는 다음과 같다.
ImageSize = width:1000px height:auto barincrement:15px
PlotArea = left:10px bottom:50px top:10px right:10px
Period = from:-251 till:0
TimeAxis = orientation:horizontal
ScaleMajor = unit:year increment:10 start:-250
ScaleMinor = unit:year increment:1 start:-251
TimeAxis = orientation:hor
AlignBars = justify
Legend = orientation:vertical position:bottom columns:1
Colors =
#legends
id:CAR value:claret
id:ANK value:rgb(0.4,0.3,0.196)
id:HER value:teal
id:HAD value:green
id:OMN value:blue
id:black value:black
id:white value:white
id:mesozoic value:rgb(0.54,0.54,0.258)
id:triassic value:rgb(0.51,0.17,0.57)
id:earlytriassic value:rgb(0.6,0.22,0.61)
id:middletriassic value:rgb(0.73,0.53,0.71)
id:latetriassic value:rgb(0.78,0.65,0.8)
id:jurassic value:rgb(0.2,0.7,0.79)
id:earlyjurassic value:rgb(0,0.69,0.89)
id:middlejurassic value:rgb(0.52,0.81,0.91)
id:latejurassic value:rgb(0.74,0.89,0.97)
id:cretaceous value:rgb(0.5,0.78,0.31)
id:earlycretaceous value:rgb(0.63,0.78,0.65)
id:latecretaceous value:rgb(0.74,0.82,0.37)
id:cenozoic value:rgb(0.54,0.54,0.258)
id:paleogene value:rgb(0.99,0.6,0.32)
id:paleocene value:rgb(0.99,0.65,0.37)
id:eocene value:rgb(0.99,0.71,0.42)
id:oligocene value:rgb(0.99,0.75,0.48)
id:neogene value:rgb(0.999999,0.9,0.1)
id:miocene value:rgb(0.999999,0.999999,0)
id:pliocene value:rgb(0.97,0.98,0.68)
id:quaternary value:rgb(0.98,0.98,0.5)
id:pleistocene value:rgb(0.999999,0.95,0.68)
id:holocene value:rgb(0.999,0.95,0.88)
id:her value:red Legend:헤레라사우루스류
id:pur value:purple Legend:용각류
id:ther value:orange Legend:수각류
id:orn value:green Legend:조반류
Legend = columns:1 left:100 top:20 columnwidth:100
BarData=
bar:eratop
bar:space
bar:periodtop
bar:space
bar:NAM1
bar:NAM2
bar:NAM3
bar:NAM4
bar:NAM5
bar:NAM6
bar:NAM7
bar:NAM8
bar:NAM9
bar:NAM10
bar:NAM11
bar:NAM12
bar:NAM13
bar:NAM14
bar:NAM15
bar:NAM16
bar:NAM17
bar:NAM18
bar:NAM19
bar:NAM20
bar:NAM21
bar:NAM22
bar:NAM23
bar:NAM24
bar:NAM25
bar:NAM26
bar:NAM27
bar:NAM28
bar:NAM29
bar:NAM30
bar:space
bar:period
bar:space
bar:era
PlotData=
align:center textcolor:black fontsize:M mark:(line,black) width:25
shift:(2,-5)
bar:periodtop
from: -251 till: -245 color:earlytriassic text:초기
from: -245 till: -228 color:middletriassic text:중기
from: -228 till: -199.6 color:latetriassic text:후기
from: -199.6 till: -175.6 color:earlyjurassic text:초기
from: -175.6 till: -161.2 color:middlejurassic text:중기
from: -161.2 till: -145.5 color:latejurassic text:후기
from: -145.5 till: -99.6 color:earlycretaceous text:초기
from: -99.6 till: -65.5 color:latecretaceous text:후기
from: -65.5 till: -55.8 color:paleocene text:고신
from: -55.8 till: -33.9 color:eocene text:에오
from: -33.9 till: -23.03 color:oligocene text:올리고.
from: -23.03 till: -5.332 color:miocene text:마이오.
from: -5.332 till: -2.588 color:pliocene text:플.
from: -2.588 till: -0.0117 color:pleistocene text:플.
from: -0.0117 till: 0 color:holocene text:홀.
bar:eratop
from: -251 till: -199.6 color:triassic text:트라이아스기
from: -199.6 till: -145.5 color:jurassic text:쥐라기
from: -145.5 till: -65.5 color:cretaceous text:백악기
from: -65.5 till: -23.03 color:paleogene text:고생대
from: -23.03 till: -2.588 color:neogene text:신생대
from: -2.588 till: 0 color:quaternary text:제4기
PlotData=
align:left fontsize:M mark:(line,white) width:5 anchor:till shift:(5,-4)
color:her bar:NAM1 from:-233.23 till:-210 text:헤레라사우루스과
color:pur bar:NAM2 from:-231.4 till:-208 text:과이바사우루스과
color:pur bar:NAM3 from:-225 till:-190 text:플라테오사우루스과
color:pur bar:NAM4 from:-228 till:-213 text:리오하사우루스과
color:pur bar:NAM5 from:-227 till:-176 text:마소스폰딜루스과
color:pur bar:NAM6 from:-183 till:-175 text:불카노돈과
color:pur bar:NAM7 from:-168 till:-125 text:투리아사우루스류
color:pur bar:NAM8 from:-175 till:-150 text:세티오사우루스과
color:pur bar:NAM9 from:-174 till:-93 text:디플로도쿠스상과
color:pur bar:NAM10 from:-157 till:-93 text:브라키오사우루스과
color:pur bar:NAM11 from:-140 till:-66 text:티타노사우루스류
color:ther bar:NAM12 from:-221 till:-183 text:코엘로피시스상과
color:ther bar:NAM13 from:-199.3 till:-66 text:케라토사우루스류
color:ther bar:NAM14 from:-170 till:-85 text:메갈로사우루스상과
color:ther bar:NAM15 from:-175.6 till:-88 text:카르노사우루스류
color:ther bar:NAM16 from:-130 till:-66 text:메가랍토르류
color:ther bar:NAM17 from:-166 till:-66 text:티라노사우루스상과
color:ther bar:NAM18 from:-151.5 till:-108 text:콤프소그나투스과
color:ther bar:NAM19 from:-140 till:-66 text:오르니토미무스류
color:ther bar:NAM20 from:-160 till:-66 text:알바레즈사우루스류
color:ther bar:NAM21 from:-139 till:-66 text:테리지노사우루스류
color:ther bar:NAM22 from:-130 till:-66 text:오비랍토르류
color:ther bar:NAM23 from:-167 till:-66 text:데이노니쿠스류
color:ther bar:NAM24 from:-155 till:0 shift:(-45,5) text:새무리
color:orn bar:NAM25 from:-200 till:-140 text:헤테로돈토사우루스과
color:orn bar:NAM26 from:-169 till:-100 text:스테고사우루스류
color:orn bar:NAM27 from:-170.3 till:-66 text:곡룡류
color:orn bar:NAM28 from:-92 till:-66 text:후두류
color:orn bar:NAM29 from:-161 till:-66 text:각룡류
color:orn bar:NAM30 from:-164 till:-66 text:조각류
PlotData=
align:center textcolor:black fontsize:M mark:(line,black) width:25
shift:(2,-5)
bar:period
from: -251 till: -245 color:earlytriassic text:초기
from: -245 till: -228 color:middletriassic text:중기
from: -228 till: -199.6 color:latetriassic text:후기
from: -199.6 till: -175.6 color:earlyjurassic text:초기
from: -175.6 till: -161.2 color:middlejurassic text:중기
from: -161.2 till: -145.5 color:latejurassic text:후기
from: -145.5 till: -99.6 color:earlycretaceous text:초기
from: -99.6 till: -65.5 color:latecretaceous text:후기
from: -65.5 till: -55.8 color:paleocene text:고신
from: -55.8 till: -33.9 color:eocene text:에오
from: -33.9 till: -23.03 color:oligocene text:올리고.
from: -23.03 till: -5.332 color:miocene text:마이오.
from: -5.332 till: -2.588 color:pliocene text:플.
from: -2.588 till: -0.0117 color:pleistocene text:플.
from: -0.0117 till: 0 color:holocene text:홀.
bar:era
from: -251 till: -199.6 color:triassic text:트라이아스기
from: -199.6 till: -145.5 color:jurassic text:쥐라기
from: -145.5 till: -65.5 color:cretaceous text:백악기
from: -65.5 till: -23.03 color:paleogene text:고생대
from: -23.03 till: -2.588 color:neogene text:신생대
from: -2.588 till: 0 color:quaternary text:제4기
11. 3. 분류
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 존재하였던 지배파충류에 속하는 동물이다.[317] 공룡과 같은 시기에 어룡, 모사사우루스, 플레시오사우루스, 익룡과 같은 다른 고생물들도 있었으나 이들은 공룡과는 다른 목으로 분류된다.공룡은 진화에 의한 종분화를 겪으면서 다양한 해부학적 요소들이 변형을 겪었다.[314]
매우 다양한 공룡들 사이에서 공통적인 해부학적 특징을 찾을 수 있다. 대표적인 것으로는 공룡들이 모두 이궁류에 속한다는 점인데, 공룡들은 두개골에 있는 측두창이 둘로 나뉘어 있다.[325] 낮은 턱 뼈도 공룡의 공통적인 해부학적 특징으로 꼽을 수 있다.[326]
계통분류학에서 공룡은 일반적으로 ''트리케라톱스''와 현생 조류(신조류)의 최근 공통 조상 (MRCA)과 모든 후손으로 구성된 그룹으로 정의된다. 또한 리처드 오웬이 공룡을 인지했을 때 인용한 3개의 속 중 2개였던 ''메갈로사우루스''와 ''이구아노돈''의 MRCA와 관련하여 공룡강을 정의할 것을 제안하기도 했다. 두 정의 모두 동일한 알려진 속을 포함한다: 공룡강 = 조반목 + 용반목. 여기에는 안킬로사우루스류 (갑옷을 입은 초식 사족보행류), 스테고사우루스류 (판을 가진 초식 사족보행류), 케라톱스류 (목도리(neck frill)를 가진 이족 또는 사족보행 초식동물), 파키케팔로사우루스류 (두꺼운 두개골을 가진 이족 보행 초식동물), 조각류 (오리 주둥이를 포함한 이족 또는 사족보행 초식동물), 수각류 (주로 이족 보행 육식동물 및 조류), 용각류 (주로 긴 목과 꼬리를 가진 대형 초식 사족보행류)와 같은 주요 그룹이 포함된다.
조류는 유일하게 생존한 공룡이다. 전통적인 분류학에서 조류는 공룡으로부터 진화한 별도의 강으로 간주되었으며, 뚜렷한 상목이었다. 그러나 대부분의 현대 고생물학자들은 해부학적 유사성에 기초한 전통적인 분류 방식을 거부하고, 각 그룹을 주어진 창립 속의 모든 후손으로 정의하는 계통 발생학적 분류학을 선호한다.[10] 조류는 용반목인 코엘루로사우루스류인 공룡의 하위 그룹 마니랍토라에 속한다.
2017년 매튜 G. 배런, 데이비드 B. 노먼, 폴 M. 배럿의 연구는 공룡 계통학의 급진적인 수정을 제안했다. 배런 외의 계통 발생학적 분석 결과는 용각류와 수각류의 전통적인 결합과는 반대로 조반목이 용각류보다 수각류에 더 가깝다는 것을 밝혀냈다. 이것은 용각류와 그 친족을 전통적인 공룡 밖으로 떨어뜨릴 것이므로, 그들은 공룡을 ''트리케라톱스 호리두스'', ''집참새'' 및 ''디플로도쿠스 카르네기''의 마지막 공통 조상과 그 모든 후손으로 재정의하여 용각류와 그 친족이 공룡으로 계속 포함되도록 했다. 그들은 또한 조반목과 수각류를 포함하는 그룹을 지칭하기 위해 조류형류라는 분류군을 부활시켰다.[11][12]
제2차 세계 대전은 고생물학 연구에 일시적인 중단을 가져왔고, 전쟁 이후 연구의 초점은 공룡보다는 화석 포유류에 점차 쏠리게 되었는데, 이는 공룡이 느리고 냉혈동물로 여겨졌기 때문이다.[27][28] 그러나 1960년대 말에 공룡 연구 분야는 현재까지 이어지는 활동의 급증을 경험했다.[29] 몇몇 획기적인 연구들이 이러한 활동을 이끌었다. 첫째, 존 오스트롬은 조류와 같은 드라메오사우루스과 수각류인 ''데이노니쿠스''를 발견하여 1969년에 발표했다. 그 해부학적 구조는 당시 지배적이던 공룡의 이미지와는 대조적으로, 활동적인 포식자였으며 온혈 동물일 가능성을 시사했다.[27]
새로운 발견들은 공룡 발견의 증가에 의해 뒷받침되었다. 인도, 남아메리카, 마다가스카르, 남극 대륙, 그리고 가장 중요한 것은 중국을 포함하여 이전에 탐험되지 않은 지역에서 활동하는 고생물학자들에 의해 주요한 새로운 공룡 발견이 이루어졌다. 수각류, 용각류, 조반목 공룡에 걸쳐 명명된 속의 수가 1990년대에 기하급수적으로 증가하기 시작했다.[13]
공룡 르네상스 이전에 공룡은 주로 린네식 분류법의 전통적인 계급 기반 시스템을 사용하여 분류되었다. 르네상스 시대에는 계통 발생과 공유된 특성에 기초한 보다 객관적인 분류 방법인 분기도의 광범위한 적용이 동반되었으며, 이는 공룡 체계 및 진화 연구에 매우 유용하다는 것이 입증되었다. 분기도 분석은 다른 기술과 함께 종종 불완전하고 단편적인 화석 기록을 보완하는 데 도움이 된다.[34][35]

공룡 화석은 뼈에만 국한되지 않고, 피부 피복, 기관 및 기타 조직의 흔적이나 광물화된 잔해도 포함한다. 이 중 케라틴 단백질을 기반으로 한 피부 피복은 가교 결합된, 소수성 분자 구조로 인해 가장 쉽게 보존된다.[38] 케라틴 기반 피부 피복 또는 뼈로 된 피부 피복의 화석은 대부분의 주요 공룡 그룹에서 알려져 있다. 비늘 피부 흔적이 있는 공룡 화석은 19세기부터 발견되었다.
1990년대부터, 보존 라거슈테테로 알려진 퇴적물에서 발견된 예외적으로 잘 보존된 화석의 주요 발견은 공룡 연조직에 대한 연구에 기여했다.[42][43] 이 중 주요 발견은 제홀 (초기 백악기)과 얀랴오 (중기-후기 쥐라기) 생물군의 암석으로, 여기서 쉬싱과 동료들이 깃털과 유사한 구조의 흔적을 가진 수백 개의 공룡 표본 (조류와 밀접한 관련이 있거나 그렇지 않은 경우)이 기술되었다.[44][45] 살아있는 파충류와 포유류에서, 멜라노솜으로 알려진 색소 저장 세포 구조는 부분적으로 색상 생성을 담당한다.[50][46] 멜라닌의 화학적 흔적과 특징적으로 모양이 있는 멜라노솜은 제홀과 얀랴오 공룡의 깃털과 비늘에서 보고되었으며, 수각류와 조반류를 모두 포함한다.[47] 이를 통해 야코프 빈터와 동료들이 ''시노사우롭테릭스''[48]와 ''프시타코사우루스''[49]와 같은 여러 공룡의 전신 복원이 가능해졌으며, 유사한 기술은 다른 지역의 공룡 화석에도 확장되었다.[50]

동시에, 메리 히그비 슈바이처, 잭 호너와 동료들이 이끄는 연구에서는 공룡 뼈 화석 내에서 보존된 다양한 연조직과 단백질의 발생을 보고했다. 슈바이처 등이 1991년 초부터 티라노사우루스과 뼈에서 적혈구와 콜라겐 섬유를 나타낼 가능성이 있는 다양한 광물화 구조가 발견되었다.[55][56][57] 그러나 2005년, 슈바이처와 동료들은 ''티라노사우루스''의 대퇴골에서 유연한 연조직이 보존되어 혈관, 골질 및 미세 구조를 유지한 결합 조직(뼈 섬유)을 포함한다고 보고했다. 이러한 발견은 원래의 연조직이 지질학적 시간 동안 보존될 수 있음을 시사했으며,[38] 여러 메커니즘이 제안되었다.[58] 이후, 2009년에 슈바이처와 동료들은 ''브라킬로포사우루스'' 대퇴골에서 유사한 미세 구조가 보존되어 있으며, 면역조직화학적 기술(항체 결합 기반)을 통해 콜라겐, 엘라스틴 및 라미닌과 같은 단백질의 존재가 입증되었다고 보고했다.[59]
공룡 주요 그룹 연대표(Holtz, 2007)
ImageSize = width:1000px height:auto barincrement:15px
PlotArea = left:10px bottom:50px top:10px right:10px
Period = from:-251 till:0
TimeAxis = orientation:horizontal
ScaleMajor = unit:year increment:10 start:-250
ScaleMinor = unit:year increment:1 start:-251
TimeAxis = orientation:hor
AlignBars = justify
Legend = orientation:vertical position:bottom columns:1
Colors =
#legends
id:CAR value:claret
id:ANK value:rgb(0.4,0.3,0.196)
id:HER value:teal
id:HAD value:green
id:OMN value:blue
id:black value:black
id:white value:white
id:mesozoic value:rgb(0.54,0.54,0.258)
id:triassic value:rgb(0.51,0.17,0.57)
id:earlytriassic value:rgb(0.6,0.22,0.61)
id:middletriassic value:rgb(0.73,0.53,0.71)
id:latetriassic value:rgb(0.78,0.65,0.8)
id:jurassic value:rgb(0.2,0.7,0.79)
id:earlyjurassic value:rgb(0,0.69,0.89)
id:middlejurassic value:rgb(0.52,0.81,0.91)
id:latejurassic value:rgb(0.74,0.89,0.97)
id:cretaceous value:rgb(0.5,0.78,0.31)
id:earlycretaceous value:rgb(0.63,0.78,0.65)
id:latecretaceous value:rgb(0.74,0.82,0.37)
id:cenozoic value:rgb(0.54,0.54,0.258)
id:paleogene value:rgb(0.99,0.6,0.32)
id:paleocene value:rgb(0.99,0.65,0.37)
id:eocene value:rgb(0.99,0.71,0.42)
id:oligocene value:rgb(0.99,0.75,0.48)
id:neogene value:rgb(0.999999,0.9,0.1)
id:miocene value:rgb(0.999999,0.999999,0)
id:pliocene value:rgb(0.97,0.98,0.68)
id:quaternary value:rgb(0.98,0.98,0.5)
id:pleistocene value:rgb(0.999999,0.95,0.68)
id:holocene value:rgb(0.999,0.95,0.88)
id:her value:red Legend:헤레라사우루스류
id:pur value:purple Legend:용각아목
id:ther value:orange Legend:수각류
id:orn value:green Legend:조반목
Legend = columns:1 left:100 top:20 columnwidth:100
BarData=
bar:eratop
bar:space
bar:periodtop
bar:space
bar:NAM1
bar:NAM2
bar:NAM3
bar:NAM4
bar:NAM5
bar:NAM6
bar:NAM7
bar:NAM8
bar:NAM9
bar:NAM10
bar:NAM11
bar:NAM12
bar:NAM13
bar:NAM14
bar:NAM15
bar:NAM16
bar:NAM17
bar:NAM18
bar:NAM19
bar:NAM20
bar:NAM21
bar:NAM22
bar:NAM23
bar:NAM24
bar:NAM25
bar:NAM26
bar:NAM27
bar:NAM28
bar:NAM29
bar:NAM30
bar:space
bar:period
bar:space
bar:era
PlotData=
align:center textcolor:black fontsize:M mark:(line,black) width:25
shift:(2,-5)
bar:periodtop
from: -251 till: -245 color:earlytriassic text:초기
from: -245 till: -228 color:middletriassic text:중기
from: -228 till: -199.6 color:latetriassic text:후기
from: -199.6 till: -175.6 color:earlyjurassic text:초기
from: -175.6 till: -161.2 color:middlejurassic text:중기
from: -161.2 till: -145.5 color:latejurassic text:후기
from: -145.5 till: -99.6 color:earlycretaceous text:초기
from: -99.6 till: -65.5 color:latecretaceous text:후기
from: -65.5 till: -55.8 color:paleocene text:팔레오.
from: -55.8 till: -33.9 color:eocene text:에오세
from: -33.9 till: -23.03 color:oligocene text:올리고.
from: -23.03 till: -5.332 color:miocene text:마이오.
from: -5.332 till: -2.588 color:pliocene text:플.
from: -2.588 till: -0.0117 color:pleistocene text:플.
from: -0.0117 till: 0 color:holocene text:홀.
bar:eratop
from: -251 till: -199.6 color:triassic text:트라이아스기
from: -199.6 till: -145.5 color:jurassic text:쥐라기
from: -145.5 till: -65.5 color:cretaceous text:백악기
from: -65.5 till: -23.03 color:paleogene text:고제3기
from: -23.03 till: -2.588 color:neogene text:신제3기
from: -2.588 till: 0 color:quaternary text:Q.
PlotData=
align:left fontsize:M mark:(line,white) width:5 anchor:till shift:(5,-4)
color:her bar:NAM1 from:-233.23 till:-210 text:헤레라사우루스과
color:pur bar:NAM2 from:-231.4 till:-208 text:과이바사우루스과
color:pur bar:NAM3 from:-225 till:-190 text:플라테오사우루스과
color:pur bar:NAM4 from:-228 till:-213 text:리오하사우루스과
color:pur bar:NAM5 from:-227 till:-176 text:마소스폰딜루스과
color:pur bar:NAM6 from:-183 till:-175 text:불카노돈과
color:pur bar:NAM7 from:-168 till:-125 text:투리아사우리아
color:pur bar:NAM8 from:-175 till:-150 text:세티오사우루스과
color:pur bar:NAM9 from:-174 till:-93 text:디플로도쿠스상과
color:pur bar:NAM10 from:-157 till:-93 text:브라키오사우루스과
color:pur bar:NAM11 from:-140 till:-66 text:티타노사우리아
color:ther bar:NAM12 from:-221 till:-183 text:코엘로피시스상과
color:ther bar:NAM13 from:-199.3 till:-66 text:케라토사우루스류
color:ther bar:NAM14 from:-170 till:-85 text:메갈로사우루스상과
color:ther bar:NAM15 from:-175.6 till:-88 text:카르노사우루스류
color:ther bar:NAM16 from:-130 till:-66 text:메가랍토르류
color:ther bar:NAM17 from:-166 till:-66 text:티라노사우루스상과
color:ther bar:NAM18 from:-151.5 till:-108 text:콤프소그나투스과
color:ther bar:NAM19 from:-140 till:-66 text:오르니토미무스류
color:ther bar:NAM20 from:-160 till:-66 text:알바레즈사우루스류
color:ther bar:NAM21 from:-139 till:-66 text:테리지노사우루스류
color:ther bar:NAM22 from:-130 till:-66 text:오비랍토르류
color:ther bar:NAM23 from:-167 till:-66 text:데이노니쿠스류
color:ther bar:NAM24 from:-155 till:0 shift:(-45,5) text:참새류
color:orn bar:NAM25 from:-200 till:-140 text:헤테로돈토사우루스과
color:orn bar:NAM26 from:-169 till:-100 text:스테고사우루스류
color:orn bar:NAM27 from:-170.3 till:-66 text:곡룡류
color:orn bar:NAM28 from:-92 till:-66 text:후두류
color:orn bar:NAM29 from:-161 till:-66 text:각룡류
color:orn bar:NAM30 from:-164 till:-66 text:조각류
PlotData=
align:center textcolor:black fontsize:M mark:(line,black) width:25
shift:(2,-5)
bar:period
from: -251 till: -245 color:earlytriassic text:초기
from: -245 till: -228 color:middletriassic text:중기
from: -228 till: -199.6 color:latetriassic text:후기
from: -199.6 till: -175.6 color:earlyjurassic text:초기
from: -175.6 till: -161.2 color:middlejurassic text:중기
from: -161.2 till: -145.5 color:latejurassic text:후기
from: -145.5 till: -99.6 color:earlycretaceous text:초기
from: -99.6 till: -65.5 color:latecretaceous text:후기
from: -65.5 till: -55.8 color:paleocene text:팔레오.
from: -55.8 till: -33.9 color:eocene text:에오세
from: -33.9 till: -23.03 color:oligocene text:올리고.
from: -23.03 till: -5.332 color:miocene text:마이오.
from: -5.332 till: -2.588 color:pliocene text:플.
from: -2.588 till: -0.0117 color:pleistocene text:플.
from: -0.0117 till: 0 color:holocene text:홀.
bar:era
from: -251 till: -199.6 color:triassic text:트라이아스기
from: -199.6 till: -145.5 color:jurassic text:쥐라기
from: -145.5 till: -65.5 color:cretaceous text:백악기
from: -65.5 till: -23.03 color:paleogene text:고제3기
from: -23.03 till: -2.588 color:neogene text:신제3기
from: -2.588 till: 0 color:quaternary text:Q.
과학자들은 지금까지 존재했던 가장 크고 작은 공룡을 확실하게 알 수 없을 것이다. 이는 화석화된 동물이 극히 일부에 불과하고, 그 대부분이 땅에 묻혀 있기 때문이다. 발견되는 비조류 공룡 표본 중 완전한 골격은 드물고, 피부 및 기타 연조직의 흔적은 더욱 드뭅니다. 뼈의 크기와 형태를 유사하고 더 잘 알려진 종과 비교하여 완전한 골격을 재구성하는 것은 정확하지 않은 기술이며, 살아있는 동물의 근육과 기타 기관을 재구성하는 것은 기껏해야 교육받은 추측의 과정이다.[91]
잘 보존된 골격으로 알려진 가장 키가 크고 무거운 공룡은 ''기라파티탄 브란카이''(이전에는 ''브라키오사우루스''의 종으로 분류)이다. 그 유해는 1907년에서 1912년 사이에 탄자니아에서 발견되었다. 여러 비슷한 크기의 개체의 뼈가 현재 베를린 자연사 박물관에 전시된 골격에 포함되었으며; 이 마운트는 높이가 12m이고 길이가 21.8m 에서 22.5m이며,[92][93] 무게는 30000kg 에서 60000kg 사이였을 것이다. 가장 긴 완전한 공룡은 길이가 27m인 ''디플로도쿠스''로, 미국 와이오밍주에서 발견되어 1907년 피츠버그의 카네기 자연사 박물관에 전시되었다.[94] 잘 보존된 화석 재료로 알려진 가장 긴 공룡은 ''파타고티탄''이다. 뉴욕의 미국 자연사 박물관에 있는 골격 마운트는 길이가 37m이다. 아르헨티나 플라자 우인쿨의 무세오 무니시팔 카르멘 푸네스에는 길이가 39.7m인 ''아르젠티노사우루스'' 재구성 골격 마운트가 있다.[95]

더 큰 공룡이 있었지만, 그들에 대한 지식은 소수의 조각난 화석에 전적으로 기반한다. 기록된 가장 큰 초식 동물의 표본 중 대부분은 1970년대 이후에 발견되었으며, 80000kg 에서 100000kg의 무게와 30m 에서 40m의 길이에 달했을 수 있는 거대한 ''아르젠티노사우루스''가 포함된다. 가장 긴 것 중 일부는 길이가 33.5m인 ''디플로도쿠스 할로룸'' (이전에는 ''세이소사우루스''), 길이가 33m 에서 34m인 ''수퍼사우루스'',
가장 큰 육식 공룡은 ''스피노사우루스''로, 길이가 12.6m 에서 18m에 달하고 무게는 7MT 에서 20.9MT였다. 다른 큰 육식 수각류에는 ''기가노토사우루스'', ''카르카로돈토사우루스'', ''티라노사우루스''가 포함된다. ''테리지노사우루스''와 ''데이노케이루스''는 수각류 중 가장 키가 컸다. 가장 큰 조반류 공룡은 길이가 16.6m인 하드로사우루스과 ''산둥고사우루스 기가스''일 것이다.[100]

최초의 공룡 화석은 19세기 초에 발견되었으며, 1841년 리처드 오웬에 의해 이 "거대한 화석 도마뱀"을 지칭하기 위해 "공룡(dinosaur)"("무서운 도마뱀"을 의미)이라는 호칭이 만들어졌다.
공룡이라는 말은 분류학적·형태학적·생태학적인 관점에서 매우 세밀한 "분류"이다. 현대까지 생존하고 있는 유일한 생물 종인 조류는 10,700종 이상이 확인되었으며, 척추동물 중에서 가장 다양한 그룹 중 하나이다. 식성은 다양하며, 초식성·육식성의 양쪽 종이 확인되었다.
공룡은 현존하는 종(조류)과 화석을 통해 모든 대륙에 분포했음이 알려져 있다. 20세기 전반까지, 조류가 공룡으로 인식되기 전에는 공룡은 둔중한 변온 동물이라는 인식이 일반적이었다. 그러나 1970년대에 일어난 공룡 르네상스에 의해 일부 공룡은 신진대사가
12. 고생물학
공룡 화석은 수천 년 동안 알려져 왔지만, 그 실체는 제대로 알려지지 않았다. 중국에서는 용의 뼈로 여겨 기록했고, 중의학에 사용하기도 했다. 유럽에서는 성경 속 거인의 유해로 생각했다.[20]
17세기 후반 영국에서 공룡 뼈에 대한 학술적 설명이 처음 등장했다. 1676년 옥스퍼드셔주 콘웰 근처에서 발견된 뼈(현재 ''메갈로사우루스''의 대퇴골)를 로버트 플롯이 연구하여 거대한 인간의 뼈라고 결론지었다.[21][22][23] 에드워드 류이드는 ''루텔룸 임피카툼''이라는 용각류 이빨 화석을 연구했다.
1815년에서 1824년 사이, 윌리엄 버클랜드는 ''메갈로사우루스''의 화석을 더 수집하여 과학 저널에 발표했다. ''이구아노돈''은 1822년 메리 앤 맨텔이 발견했다고 알려져 있지만, 기데온 맨텔이 먼저 발견했다는 주장도 있다. 기데온 맨텔은 1825년에 연구 결과를 발표했다.
1842년, 리처드 오웬은 "공룡"이라는 용어를 만들었다.[7][8][9][24] 이 용어는 δεινόςgrc (deinos, 무시무시한)와 σαῦροςgrc (sauros, 도마뱀)에서 유래했다. 오웬은 ''이구아노돈'', ''메갈로사우루스'', ''힐라에오사우루스''가 뚜렷한 특징을 공유한다고 보고 별개의 분류군으로 제시했다.[7][8] 앨버트 공의 지원으로 런던 자연사 박물관이 설립되었다.[25]
1858년, 윌리엄 파커 포울크는 뉴저지주 해던필드에서 미국 최초의 공룡 화석(''하드로사우루스'')을 발견했다.[26] 이는 1834년 메이드스톤에서 발견된 이구아노돈에 이어 거의 완전한 공룡 골격 중 두 번째로 발견된 것이었으며, 두 발로 걷는 생물이었다. 이 발견으로 미국에서 공룡 열풍이 불었다.
에드워드 드링커 코프와 오스니엘 찰스 마시는 뼈 전쟁이라 불리는 치열한 경쟁을 벌였다. 이 싸움은 30년 이상 지속되었으며, 코프가 사망한 1897년에 끝났다. 이들의 경쟁으로 많은 표본이 손상되거나 파괴되었지만, 고생물학에 큰 기여를 했다. 마시는 86개, 코프는 56개의 새로운 공룡 종을 발견했다. 코프의 수집품은 미국 자연사 박물관에, 마시의 수집품은 피바디 자연사 박물관에 있다.
1861년, 독일 졸른호펜에서 발견된 시조새 화석은 새와 파충류의 특징을 모두 가지고 있었다. 1868년, 토마스 헉슬리는 새의 조상이 공룡이라고 주장했다.[277] 1969년 존 오스트롬 등이 데이노니쿠스를 기재하면서 새와 공룡의 관계가 다시 주목받았다.[282] 자크 고티에의 분지학적 방법 발달[283][284]과 새로운 화석 발견으로 이 관계는 더욱 확실해졌다.[265]
1969년에 기술된 ''데이노니쿠스'' 연구는 "크고 느린 변온 동물"이라는 공룡의 이미지를 "항온성이고 활동적인 동물"로 바꾸었다. 1970년대 로버트 배커 등이 주도한 패러다임 시프트는 '''공룡 르네상스'''라고 불렸다.
12. 1. 크기
공룡은 트라이아스기 후기에 출현하여 백악기 후기까지 지배파충류로 번성하였다.[317] 중생대에 번성한 공룡은 특히 쥐라기와 백악기에 융성하였으며, 다른 동물들은 공룡의 크기에 비할 수 없었다. 당시 포유류는 대부분 설치류였고, 크기는 지금의 고양이만 했다.[319] 예외적으로 레페노마무스와 같은 삼돌기치목 포유류는 12kg-14kg에 달해 작은 프시타코사우루스를 사냥하기도 했다.[320]2006년까지 500여 속의 공룡이 밝혀졌으며, 이 중 75% 정도는 골격 화석이 발견되었다.[321] 초기 연구에서는 3400여 속으로 추정하기도 했으나, 오늘날 화석 기록과는 차이가 있다.[322] 2008년 10월 17일 현재 확인된 공룡의 종은 1047개이다.[323]
공룡은 생김새, 크기, 식성, 행동 양식 등에서 매우 다양했다. 초식과 육식 공룡이 모두 있었고, 2족 보행이나 4족 보행을 했다. 플라테오사우루스나 이구아노돈처럼 2족 보행과 4족 보행을 모두 할 수 있는 공룡도 있었다. 몸을 보호하기 위해 뿔, 등딱지, 골갑, 볏 등을 가진 종류도 있었다. 공룡은 남극을 포함한 지구 어디에서나 서식했다.[324]
12. 1. 1. 최대 및 최소
공룡은 그 크기가 매우 다양했다. 닭 정도의 작은 크기부터, 육상 동물 중에서는 최대 크기까지 존재했다. 가장 큰 공룡은 용각류였으며, 그 중에서도 수퍼사우루스가 가장 큰 것으로 알려져 있다. 용각류는 수생 고래류를 제외하면 지구 역사상 가장 큰 동물이며, 육상 동물 중에서는 지구상 최대이다.[244] 마라아푸니사우루스나 불하토카요사우루스처럼 더 거대한 크기에 달했을 것으로 추정되는 용각류도 있었지만, 화석이 현존하지 않아 불확실하다.[245][246] 부분적인 표본으로는 바로사우루스 또는 수퍼사우루스에 속한다고 여겨지는 대형 표본이 있으며, 일부 추정에서는 전체 길이 40~50미터 범위에 달했을 것으로 추정된다.[247][248]체중은 브라키오사우루스과의 기라파티탄 등이 80톤 이상이었다는 설도 있지만, 이 수치는 다소 과장되었다는 의견도 있다.[249]
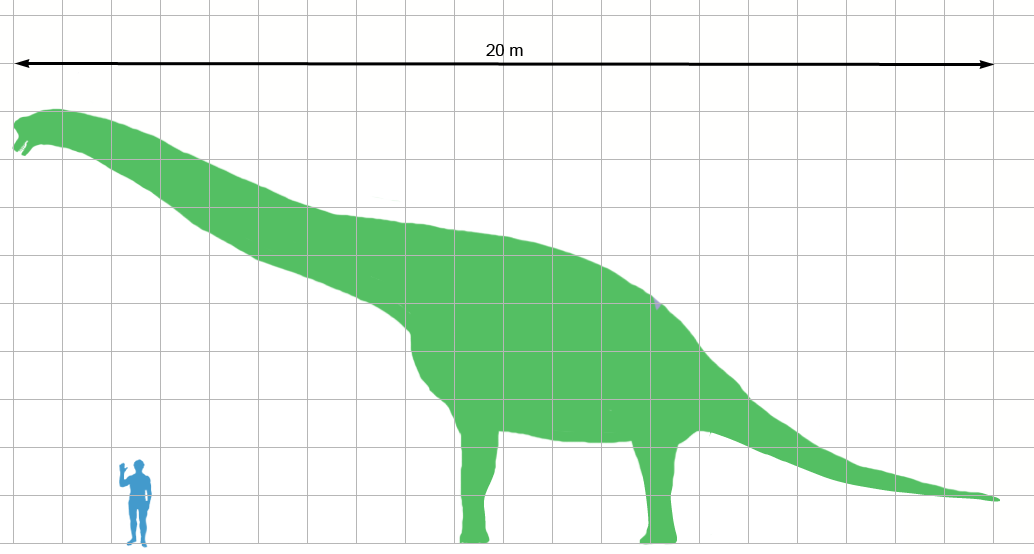
공룡의 지리적 분포, 모양, 행동, 크기는 매우 다양했기 때문에 "평균 크기"를 정확하게 정의하기는 어렵다.[328] 빌 에릭슨에 따르면, 무게가 500kg과 5톤 사이로 추정되는 공룡 중 80%는 미국 서부의 모리슨층에서 발견된다. 스테고사우루스와 용각류 등 평균 20톤 이상인 공룡은 오늘날의 일반적인 대형 포유류와 비교되지 않을 정도로 거대하다. 63세대에 따른 공룡의 평균 무게는 395.9kg(회색곰의 무게)를 초과하며, 일반 대중 중간값은 2톤(기린과 비교)이다. 이는 평균 무게 631g인 멸종 포유류(788세대)와 크게 대조되며, 현재까지 발견된 가장 작은 공룡의 3분의 2 이상의 크기였다. 모든 현대 포유 동물, 공룡의 거의 대부분은 개인의 2%를 제외한 모든 살아있는 포유 동물보다 더 컸다.[329]
평균적으로 크기가 컸던 공룡으로는 아르헨티노사우루스, 세이스모사우루스가 있고, 육식공룡 중에서는 스피노사우루스, 티라노사우루스, 기가노토사우루스, 수코미무스가 있다.
가장 잘 알려진 공룡들은 큰 크기로 유명하지만, 많은 중생대 공룡은 인간 크기이거나 그보다 작았으며, 현대 조류는 일반적으로 작은 크기이다.
12. 2. 행동
현대 조류는 매우 사회적이며 무리를 지어 생활하는 경우가 많다. 조류와 악어류(조류의 가장 가까운 현존 친척)에서 흔히 나타나는 일부 행동이 멸종된 공룡 그룹에서도 흔히 나타났다는 데에는 대체로 의견이 일치한다. 화석 종의 행동에 대한 해석은 일반적으로 골격의 자세와 그들의 서식지, 생체역학에 대한 컴퓨터 시뮬레이션 및 유사한 생태적 틈새의 현대 동물과의 비교를 기반으로 한다.조류 외에 많은 공룡 그룹에서 흔히 나타나는 광범위한 행동으로서의 무리지어 다니는 또는 무리에 대한 첫 번째 잠재적 증거는 1878년에 발견된 31마리의 이구아노돈(''Iguanodon'')이다. 당시 이들은 깊고 물이 찬 싱크홀에 빠져 익사한 후 벨기에 베르니사르에서 함께 죽은 것으로 여겨졌다. 그 이후 다른 대량 사망 현장이 발견되었다. 이러한 현장은 여러 개의 발자국과 함께 많은 초기 공룡 종에서 군집 행동이 흔했음을 시사한다. 수백, 수천 마리의 초식동물의 발자국은 오리 부리 공룡(하드로사우루스과)이 아메리카 들소나 아프리카 스프링복과 같이 큰 무리로 이동했을 수 있음을 나타낸다. 용각류 발자국은 이러한 동물들이 적어도 잉글랜드 옥스퍼드셔에서는 여러 종으로 구성된 그룹으로 이동했음을 보여주지만, 구체적인 무리 구조에 대한 증거는 없다. 무리를 이루는 것은 방어, 철새 목적, 또는 어린 개체를 보호하기 위해 진화했을 수 있다. 다양한 수각류, 용각류, 곡룡류, 조각류, 각룡류를 포함한 많은 유형의 느리게 성장하는 공룡이 미성숙한 개체의 집단을 형성했다는 증거가 있다. 한 예로 중국 내몽골 지역의 한 유적지에서 1~7세의 20마리 이상의 시노르니토미무스(''Sinornithomimus'') 유해가 발견되었다. 이 집단은 진흙에 갇힌 사회적 집단으로 해석된다. 공룡을 군집 동물로 해석하는 것은 육식 수각류가 큰 먹이를 잡기 위해 함께 일하는 팩 헌터로 묘사하는 것으로 확장되었다. 그러나 이러한 생활 방식은 현대 조류, 악어 및 기타 파충류에서는 흔하지 않으며, ''데이노니쿠스''(Deinonychus)와 ''알로사우루스''(Allosaurus)와 같은 수각류에서 포유류와 같은 팩 사냥을 시사하는 매장학적 증거는 많은 현대 이궁류 포식자에서 볼 수 있듯이 먹이를 먹는 동물 간의 치명적인 분쟁의 결과로 해석될 수도 있다.

마기노케팔리아류, 수각류, 람베오사우루스아과와 같은 일부 공룡의 볏과 프릴은 능동적인 방어에 사용하기에는 너무 약했을 수 있으며, 성적 또는 공격적인 과시에 사용되었을 가능성이 높지만, 공룡의 짝짓기와 영토성에 대해서는 알려진 바가 거의 없다. 물린 자국에서 머리에 상처가 난 것은 적어도 수각류가 적극적인 공격적 대결에 참여했음을 시사한다.
행동적 관점에서 볼 때 가장 가치 있는 공룡 화석 중 하나는 1971년 고비 사막에서 발견되었다. 여기에는 벨로키랍토르(''Velociraptor'')가 원시각룡(''Protoceratops'')을 공격하는 모습이 포함되어 있어, 공룡이 실제로 서로 공격했다는 증거를 제공한다. 살아있는 먹이를 공격했다는 추가 증거는 하드로사우루스과 공룡인 ''에드몬토사우루스(Edmontosaurus)''의 부분적으로 치유된 꼬리이다. 꼬리는 티라노사우루스에게 물려 생존했음을 보여주는 방식으로 손상되었다. 일부 공룡 종 사이의 동족 포식은 2003년 마다가스카르에서 수각류인 ''마준가사우루스(Majungasaurus)''가 관련된 이빨 자국에서 확인되었다.
공룡과 현대 조류 및 파충류의 공막 고리를 비교하여 공룡의 일상 활동 패턴을 추론하는 데 사용되었다. 대부분의 공룡이 낮에 활동했다는 주장이 있었지만, 이러한 비교를 통해 드로마에오사우루스과, ''유라베나토르(Juravenator)'', ''메가프노사우루스(Megapnosaurus)''와 같은 소형 포식 공룡이 야행성일 가능성이 높다는 것을 보여주었다. 각룡류, 용각류, 하드로사우루스과, 조류공룡류와 같은 대형 및 중형 초식 및 잡식 공룡은 하루 종일 짧은 간격으로 활동하는 주야혼합성일 수 있지만, 소형 조반류인 ''아길리사우루스(Agilisaurus)''는 주행성으로 추론되었다.
''오릭토드로메우스(Oryctodromeus)''와 같은 공룡의 화석 증거를 바탕으로 일부 조반류 종은 부분적으로 굴성 (굴을 파는) 생활 방식을 이끌었던 것으로 보인다. 많은 현대 조류는 수목 생활을 하며, 특히 엔안티오르니테스류를 포함한 많은 중생대 조류도 마찬가지였다.[104] 일부 초기 조류와 같은 종(드로마에오사우루스과 포함)도 이미 수목 생활을 했을 수 있지만(예: ''미크로랍토르''), 대부분의 비조류 공룡은 육상 이동에 의존했던 것으로 보인다. 공룡이 어떻게 땅 위에서 움직였는지에 대한 정확한 이해는 공룡 행동 모델의 핵심이다. 로버트 맥닐 알렉산더가 개척한 생체역학은 이 분야에서 중요한 통찰력을 제공했다. 예를 들어, 근육과 중력이 공룡의 골격 구조에 가하는 힘에 대한 연구를 통해 공룡이 얼마나 빨리 달릴 수 있는지, 디플로도쿠스과가 채찍과 같은 꼬리 채찍질을 통해 음속 폭음을 만들 수 있는지, 그리고 용각류가 물에 뜰 수 있는지 등을 조사했다.
12. 3. 생식
공룡이 어떤 방법으로 번식했는지는 확실하지 않다. 그러나 공룡알 화석이 발견되어, 적어도 몇 종류는 다른 파충류와 마찬가지로 알을 낳았을 것이라 추측하고 있다. 암컷은 땅을 파서 둥지를 만든 후 12~24개의 알을 낳았다. 그 중에는 알에서 깬 새끼를 일일이 돌보는 것도 있었고, 새끼가 제 나름으로 살아가도록 내버려두는 것도 있었다.[105]모든 공룡은 양막란을 낳았다. 공룡 알은 보통 둥지에 낳았다. 대부분의 종은 컵, 돔, 판, 침대 긁기, 둔덕 또는 굴과 같은 다소 정교한 둥지를 만든다. 일부 현대 조류 종은 둥지가 없다. 절벽에 둥지를 트는 바다오리는 맨바위에 알을 낳고, 수컷 황제펭귄은 몸과 발 사이에 알을 보관한다. 원시 조류와 많은 비조류 공룡은 수컷이 주로 알을 품는 공동 둥지에 알을 낳는 경우가 많다. 현대 조류는 하나의 기능적인 난관만 가지고 있고 한 번에 하나의 알을 낳지만, 더 원시적인 조류와 공룡은 악어처럼 두 개의 난관을 가지고 있었다. ''트로오돈''과 같은 일부 비조류 공룡은 성체가 1~2일마다 한 쌍의 알을 낳고 모든 알이 낳일 때까지 포육을 지연시켜 동시 부화를 보장하는 반복적 산란을 보였다.[106]
암컷은 알을 낳을 때 딱딱한 바깥 뼈와 골수 사이의 사지에서 특별한 유형의 뼈를 성장시킨다. 칼슘이 풍부한 이 해면질 뼈는 알 껍질을 만드는 데 사용된다. ''티라노사우루스'' 골격의 특징 발견은 멸종된 공룡의 해면질 뼈에 대한 증거를 제공했고, 고생물학자들이 화석 공룡 표본의 성별을 처음으로 결정할 수 있게 했다. 추가 연구를 통해 육식 공룡 ''알로사우루스''와 조반류 ''테논토사우루스''에서 해면질 뼈가 발견되었다. ''알로사우루스''와 ''티라노사우루스''를 포함하는 공룡 계통은 공룡 진화 초기에 ''테논토사우루스''로 이어진 계통에서 분기되었기 때문에, 해면질 조직의 생산은 모든 공룡의 일반적인 특징임을 시사한다.

현대 조류의 또 다른 널리 퍼진 특징은 부화 후 새끼에 대한 부모의 보살핌이다. 잭 호너가 1978년 몬태나에서 발견한 ''마이아사우라''("착한 어머니 도마뱀") 둥지 터는 조반류에서 부모의 보살핌이 출생 후 오랫동안 지속되었음을 보여주었다. 오비랍토르류 ''Citipati osmolskae''의 표본이 1993년에 닭과 같은 포육 자세로 발견되었는데, 이는 알을 따뜻하게 유지하기 위해 깃털의 단열층을 사용하기 시작했음을 나타낼 수 있다.[107] 기초 용각류 ''마소스폰딜루스''의 배아가 이가 없는 채로 발견되었는데, 이는 어린 공룡에게 먹이를 주기 위해 어느 정도 부모의 보살핌이 필요했음을 나타낸다. 트랙웨이는 또한 스코틀랜드 북서부 스카이 섬에서 조반류의 부모 행동을 확인했다.
그러나 많은 공룡 종, 특히 수각류에서 조숙성 또는 초조숙성에 대한 충분한 증거가 있다. 예를 들어, 비조류형류는 느린 성장률, 둥우리와 같은 알 묻기 행동 및 출생 후 곧바로 날 수 있는 능력이 있음이 풍부하게 입증되었다.[108][109][110][111] ''티라노사우루스''와 ''트로오돈'' 모두 명확한 초조숙성을 가진 유체를 가지고 있었고 성체와 다른 생태적 지위를 차지했을 가능성이 있다.[106] 용각류에 대해 초조숙성이 추론되었다.[112]
생식기 구조는 색소 침착 또는 잔류 인산칼슘염을 통해 보존될 수 있는 비늘이 없으므로 화석화될 가능성이 낮다. 2021년, ''프시타코사우루스''의 공룡 총배설강 외부의 가장 잘 보존된 표본이 기술되었으며, 양성 모두의 사회적 과시에 사용되는 악어류 사향선과 유사한 측면 부종과 신호 기능을 반영할 수 있는 색소 침착된 부위를 보여주었다. 그러나 이 표본만으로는 이 공룡이 성적 신호 기능을 가지고 있는지 여부를 결정하기에 충분한 정보를 제공하지 못하며, 가능성만 뒷받침한다. 총배설강 시각적 신호는 살아있는 조류의 수컷 또는 암컷에서 발생할 수 있으므로 멸종된 공룡의 성별을 결정하는 데 유용하지 않을 가능성이 높다.[113]
공룡도 다른 동물과 마찬가지로 교미를 하고 산란한다. 구애 행동을 했을 가능성이 있다. 또한, 한 번에 많은 알을 낳을 수 있기 때문에 번식 효율도 높아진다.
- '''수각류'''
주로 수각류는 얕게 판 구멍에 알을 낳고, 흙이나 식물을 덮는 단순한 둥지를 만들었다. 알이 둥지 중앙에 공간이 생기도록 도넛 모양으로 2~3층으로 겹쳐 낳고, 모든 알을 낳은 뒤 흙이나 식물로 덮어 알 없는 둥지 중앙에 어미가 웅크리고 앉아 알을 보호했다고 생각되는 경우, 둥지가 지면을 얕게 판 U자형으로, 가장자리가 낮게 솟아 있으며, 알은 두 개씩 짝을 이루어 부드럽게 한 둥지 안의 땅에 알의 뾰족한 부분이 꽂히도록 낳은 경우가 있다. 또한 산란 후에는 현대의 조류처럼 알을 품어 따뜻하게 했다고 생각된다.[270]
- '''용각류''' (주로 티타노사우루스형류)
용각류는 주로 뒷다리를 사용하여 한 방향으로 구멍을 파고 얕고 길쭉한 둥지를 만들어 알을 낳았다고 추측된다. 한 번의 산란으로 낳는 알의 수가 100개를 넘는 경우도 있었다고 생각된다.[270]
- '''조각류'''
조각류 중에서도 마이아사우라는, 절구 모양의 둥지에 최대 40개의 알을 낳고, 식물을 알 위에 덮었다고 생각된다. 둥지 안의 알의 배치에는 규칙성이 없이 제각각이었다.[270]
지중에 둥지를 파고, 땅 속에 알을 낳는 방법은, 알을 보호하고 부화까지 온도와 습도를 조절할 수 있는 이점이 있었다. 또한, 한 번에 많은 알을 낳을 수 있기 때문에, 번식 효율도 높아졌다.[271]
12. 4. 생리학
공룡은 현생 악어와 조류처럼 4개의 심방을 가진 심장을 가졌을 것으로 추정된다. 이는 모든 주룡류가 공유하는 특성일 가능성이 높다. 현생 조류는 높은 신진대사를 보이며 내온성("온혈")이지만, 공룡 계통에서 이 특성이 얼마나 오래되었는지에 대한 논쟁은 1960년대부터 활발하게 진행되어 왔다.[114] 다양한 연구 결과, 공룡은 내온성, 외온성("냉혈"), 또는 그 중간 어디쯤에 해당한다고 주장되어 왔다.[114]최근 연구자들 사이에서는 공룡의 서로 다른 계통이 서로 다른 신진대사를 가졌지만, 대부분은 다른 파충류보다 높은 신진대사율을 가졌고 현생 조류와 포유류보다는 낮았다는 합의가 이루어지고 있다.[115] 이를 중온성이라고 부르기도 한다.[116] 악어와 그들의 멸종된 친척들에 대한 증거는 이러한 높은 신진대사가 공룡과 악어의 공통 조상인 초기 주룡류에서 발달했을 수 있음을 시사한다.[117][118]

비조류 공룡이 발견된 후, 고생물학자들은 처음에는 이들이 외온성이라고 가정했다. 이는 고대 공룡이 비교적 느리고 둔한 생물임을 암시하는 데 사용되었지만, 많은 현대 파충류는 체온을 조절하기 위해 외부 열원에 의존함에도 불구하고 빠르고 가볍다. 공룡을 외온성으로 보는 생각은 로버트 T. 배커가 1968년에 이 주제에 대한 영향력 있는 논문을 발표할 때까지 널리 퍼져 있었다. 배커는 해부학적, 생태학적 증거를 사용하여 땅에 꼬리를 끌고 다니는 수생 동물로 묘사되었던 용각류가 활발한 육상 생활을 했던 내온성 동물이라고 주장했다. 1972년, 배커는 에너지 요구량과 포식자-피식자 비율을 기반으로 자신의 주장을 확장했다. 이것은 공룡 르네상스를 이끈 중요한 결과 중 하나였다.[119][120][27]
공룡 생리학에 대한 현대적 이해에 가장 크게 기여한 것 중 하나는 고생물 조직학이다.[121][122] 1960년대부터 아르망 드 리클레스는 섬유판 골 - 불규칙하고 섬유질의 질감을 가지고 혈관으로 채워진 뼈 조직 - 이 일관된 빠른 성장과 따라서 내온성을 나타낸다고 제안했다. 섬유판 골은 공룡과 익룡에서 흔히 발견되었지만,[123][124] 보편적으로 존재하지는 않았다.[125][126] 이로 인해 다양한 공룡 계통에서 성장 곡선을 재구성하고 성장 속도의 진화를 모델링하는 상당한 연구가 이루어졌으며,[127] 이는 공룡이 현생 파충류보다 더 빨리 성장했음을 시사했다.[122]
내온성을 시사하는 다른 증거로는 많은 계통에서 깃털 및 기타 유형의 신체 덮개가 존재한다는 점, 외온성 동물과 비교하여 골 조직 내 산소-18 동위원소의 일관된 비율이 특히 위도와 기온이 변함에 따라 안정적인 내부 온도를 시사한다는 점,[128][129] 그리고 극지 공룡의 발견 등이 있다.[131][132][133][134]
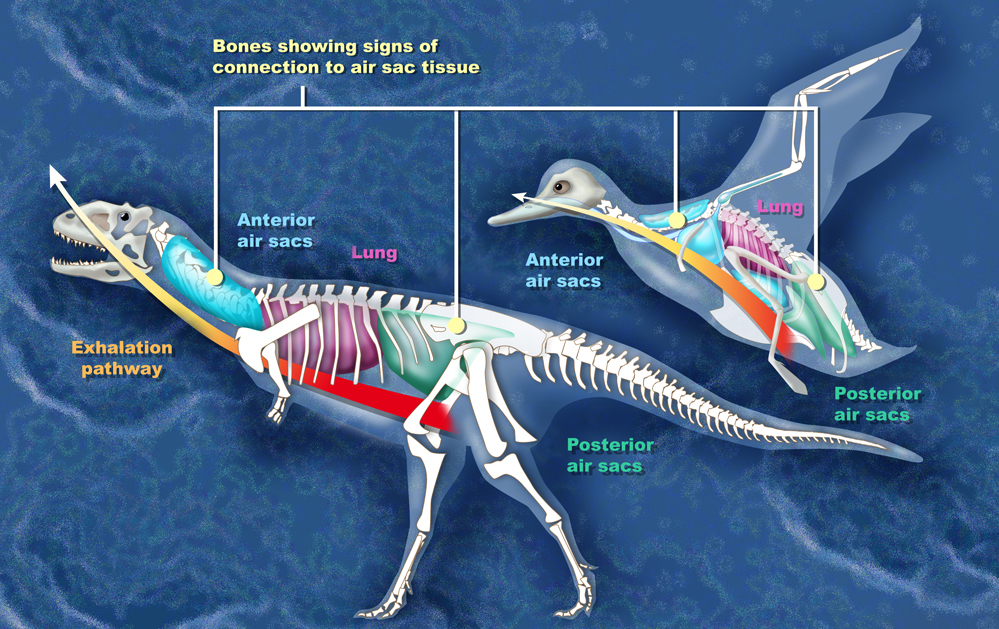
수각류 공룡에서 더 높은 신진대사는 기낭 시스템을 특징으로 하는 조류 호흡 시스템의 진화에 의해 뒷받침되었다. 가장 초기의 수각류에서 나타났을 수 있는 이러한 호흡 시스템은[135] 비슷한 크기의 포유류에 비해 더 많은 산소를 공급하는 동시에 더 큰 안정시 일회 호흡량을 가지고 호흡 빈도를 낮추어 더 높은 활동 수준을 유지할 수 있게 해준다.[90] 빠른 공기 흐름은 또한 효과적인 냉각 메커니즘이었으며, 이는 낮은 신진대사율과 함께[136] 거대한 용각류가 과열되는 것을 막아주었을 것이다. 이러한 특성은 용각류가 거대한 크기로 빠르게 성장할 수 있게 해주었을 것이다.[137][138] 용각류는 또한 크기의 이점을 얻었을 수 있는데, 표면적 대 부피 비율이 작다는 것은 거대 온열이라고 불리는 현상으로, 더 쉽게 체온을 조절할 수 있다는 것을 의미했다.[90][139]
다른 파충류와 마찬가지로, 공룡은 주로 요산뇨성인데, 즉, 그들의 신장은 혈류에서 질소성 폐기물을 추출하여 요소나 암모니아 대신 요산으로 요관을 통해 창자로 배설한다. 이것은 물을 절약하는 데 도움이 되었을 것이다.[115] 대부분의 현생 종에서 요산은 반고체 폐기물로 대변과 함께 배설된다.[140][141] 그러나 적어도 일부 현대 조류 (예: 벌새)는 선택적으로 암모니아뇨성이 될 수 있으며, 대부분의 질소성 폐기물을 암모니아로 배설한다.[142] 이 물질은 창자의 배출물과 함께 총배설강에서 나온다.[143][144] 또한 많은 종이 펠릿을 토해내며,[145] 화석 펠릿은 쥐라기 시대부터 ''안키오르니스''에서 알려져 있다.[146]
13. 조류의 기원
공룡이 새의 조상일 가능성은 1868년 토머스 헨리 헉슬리에 의해 처음 제기되었다. 20세기 초 게르하르트 하일만의 연구로 새가 공룡의 후손이라는 이론 대신 테코돈트의 후손이라는 주장이 나왔는데, 이는 공룡에게 쇄골이 없다는 것이 주요 증거였다. 그러나 이후 쇄골(또는 분리된 쇄골이 융합된 갈비뼈)이 실제로 존재한다는 사실이 밝혀졌다. 1924년 이전에 ''오비랍토르''에서 발견되었지만, 쇄골간으로 잘못 알려졌다. 1970년대에 오스트롬이 공룡-새 이론을 부활시켰고, 계통분류학의 발전과 함께 소형 수각류와 초기 새의 발견이 늘면서 이 이론은 지지를 얻었다.
새는 수각류 공룡과 100개 이상의 뚜렷한 해부학적 특징을 공유하며, 가장 가까운 고대 친척으로 여겨진다. 새는 마니랍토란 코일루로사우루스와 가장 밀접하게 관련되어 있다. 앨런 페듀치아와 래리 마틴 등 일부 과학자들은 하일만의 원시 아르코사우루스 제안을 수정하거나, 마니랍토란 수각류가 새의 조상이지만 공룡이 아니라 수렴하는 다른 진화 경로를 제안하기도 했다.
목, 치골, 손목 (반달 모양 손목뼈), 팔, 어깨뼈대, 쇄골 (Y자 뼈), 가슴뼈 등은 조류와 공룡이 골격에서 유사성을 보이는 중요한 부위이다. 분지학적 분석을 통한 골격 비교는 이러한 연관성을 강화한다.[155]
공룡과 조류의 관계는 거의 확실하지만, 앞다리 손가락에는 불일치가 있었다. 공룡은 조류에 가까워질수록 다섯 손가락 중 5, 4번째 손가락이 퇴화하는(1, 2, 3번째 손가락이 남는) 경향이 있지만, 현생 조류의 손가락은 발생학적으로 2, 3, 4번째 손가락으로 관찰되기 때문이다.[285] 귄터 바그너 등은 비조류 공룡에서 조류로 진화하면서 앞다리 손가락 위치 관계가 변했다고 설명했고,[286][287] 타무라 등은 닭 배아 실험을 통해 발생 과정에서 손가락 원기가 위치 변화를 일으킨다는 것을 보였다.[288] 2009년에는 1, 5번째 손가락이 퇴화한(2, 3, 4번째 손가락이 발달한) 림사우루스가 보고되기도 했지만,[289] 손가락에 대한 의문은 여전히 남아있다.
13. 1. 깃털
공룡이 새의 조상일 가능성은 1868년 토머스 헨리 헉슬리에 의해 처음 제기되었다. 1970년대에 오스트롬은 공룡-새 이론을 부활시켰고, 계통분석의 출현으로 수십 년 동안 이 이론에 대한 지지가 커졌으며, 소형 수각류와 초기 새의 발견이 크게 증가했다. 특히 제홀 생물군 화석이 주목할 만한데, 다양한 수각류와 초기 새가 발견되었으며, 종종 어떤 종류의 깃털을 가지고 있었다.[35] 새는 수각류 공룡과 100개 이상의 뚜렷한 해부학적 특징을 공유하며, 현재 일반적으로 가장 가까운 고대 친척으로 받아들여지고 있다. 새는 마니랍토란 코일루로사우루스와 가장 밀접한 관련이 있다.
깃털은 현대 조류의 가장 특징적인 특징 중 하나이며, 여러 비조류 공룡도 공유하는 특성이다. 현재 화석 증거의 분포를 기반으로 볼 때, 깃털은 조상 공룡의 특징이었지만 일부 종에서는 선택적으로 사라졌을 수 있다.[147] 깃털 또는 깃털과 유사한 구조의 직접적인 화석 증거는 사우루스류와 조반목 공룡을 포함한 다양한 비조류 공룡 그룹에서 발견되었다.[35] 단순하고 분지된 깃털과 유사한 구조는 헤테로돈토사우루스과, 원시 신조반목류[148] 및 수각류, 그리고 원시 각룡류에서 알려져 있다. 현대 조류의 날개깃과 유사한 진정한 날개깃에 대한 증거는 수각류 하위 그룹인 마니랍토라에서만 발견되었으며, 여기에는 오비랍토르류, 트로오돈과, 드로마이오사우루스과, 그리고 조류가 포함된다.
그러나 연구자들은 이러한 구조가 계통 간에 공통적인 기원을 공유하는지(즉, 상동적)[150][153] 또는 조류형류 사이의 피부 덮개에 대한 광범위한 실험의 결과인지에 대해 동의하지 않는다.[151] 전자의 경우, 필라멘트는 조류형류 계통에서 흔했을 수 있으며 공룡 자체가 나타나기 전에 진화했을 수 있다.[147]
''시조새''는 공룡과 조류 사이의 잠재적인 연결을 보여주는 최초의 화석이었다. 찰스 다윈의 ''종의 기원''(1859) 출판 2년 만에 발견된 이 화석은 진화 생물학과 창조론 지지자들 사이의 논쟁을 촉발시켰다. 이 초기 조류는 공룡과 매우 유사하여, 주변 암석에서 깃털의 명확한 흔적이 없으면 적어도 하나의 표본이 작은 수각류인 ''콤프소그나투스''로 오인되었다. 1990년대 이후, 깃털을 가진 공룡의 수가 더 많이 발견되었으며, 공룡과 현대 조류 사이의 밀접한 관계에 대한 더욱 강력한 증거를 제공했다.
14. 문화적 묘사
인간의 기준에서 공룡은 환상적인 모습과 종종 거대한 크기를 가진 생물이었다. 이처럼 공룡은 대중의 상상력을 사로잡았고 인간 문화의 지속적인 일부가 되었다. "공룡"이라는 단어가 일반적인 방언에 들어간 것은 동물의 문화적 중요성을 반영한다. 영어에서 "dinosaur"는 실용적이지 않게 크거나, 구식이거나, 멸종될 운명에 처한 모든 것을 묘사하는 데 일반적으로 사용된다.
공룡에 대한 대중의 열광은 처음 빅토리아 시대 잉글랜드에서 시작되었는데, 1854년, 공룡 유해에 대한 최초의 과학적 묘사 후 30년이 지나자 런던 크리스탈 팰리스 공원에 실물과 같은 크리스탈 팰리스 공룡 조각상이 공개되었다. 크리스탈 팰리스 공룡은 매우 인기를 끌었고 곧 더 작은 복제품 시장이 활발하게 형성되었다. 그 후 수십 년 동안 공룡 전시회가 전 세계의 공원과 자연사 박물관에서 열리면서 다음 세대들이 몰입적이고 흥미로운 방식으로 이 동물들을 접할 수 있게 되었다. 공룡의 지속적인 인기는 결국 공룡 과학에 대한 상당한 공적 자금 지원으로 이어졌으며, 새로운 발견을 자주 자극했다. 예를 들어 미국에서는 대중의 관심을 끌기 위한 박물관 간의 경쟁이 1880년대와 1890년대의 골 전쟁으로 직접 이어졌으며, 이 기간 동안 두 명의 다투는 고생물학자가 엄청난 과학적 기여를 했다.
공룡에 대한 대중의 관심은 문학, 영화, 기타 미디어에 공룡의 등장을 보장했다. 1852년 찰스 디킨스의 황폐한 집에서 잠깐 언급된 것을 시작으로, 공룡은 수많은 소설 작품에 등장했다. 쥘 베른의 1864년 소설 ''지구 속 여행'', 아서 코난 도일 경의 1912년 저서 ''잃어버린 세계'', 1914년 애니메이션 영화 ''공룡 거티''(최초의 애니메이션 공룡 등장), 상징적인 1933년 영화 ''킹콩'', 1954년 ''고질라''와 그 수많은 속편, 마이클 크라이튼의 1990년 베스트셀러 소설 ''쥬라기 공원''과 1993년 영화 각색은 소설에서 공룡이 등장하는 몇 가지 주목할 만한 예시이다. 저명한 고생물학자를 포함한 공룡에 대한 일반적인 관심의 논픽션 저술가들은 종종 이 동물들을 독자들에게 과학 전반에 대한 교육을 제공하는 수단으로 활용하려고 했다. 공룡은 광고에서 흔하게 등장하며, 수많은 회사들은 자사의 제품을 판매하거나 경쟁사를 느리고, 멍청하거나, 구식으로 묘사하기 위해 인쇄물 또는 텔레비전 광고에서 공룡을 언급했다.[211]
최초의 공룡 화석은 19세기 초에 발견되었으며, 1841년 리처드 오웬에 의해 이 "거대한 화석 도마뱀"을 지칭하기 위해 "공룡(dinosaur)"("무서운 도마뱀"을 의미)이라는 호칭이 만들어졌다. 그 이후, 화석 공룡의 골격 표본은 전 세계 박물관의 주요 볼거리가 되었고, 공룡은 대중 문화의 영속적인 일부가 되었다. 현존하는 생물에 비해 매우 큰 체격과 괴물적인 외관을 가진 종이 많기 때문에, 쥬라기 공원에 대표되는 SF 작품에 등장하는 경우가 많으며, 고질라를 비롯한 다수의 창작 몬스터의 모티프로도 사용되었다.
참조
[1]
논문
A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution
2018
[2]
논문
The assembly of the avian body plan: a 160-million-year long process
https://www.bolletti[...]
2018
[3]
학술지
A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone
2007
[4]
학술지
The Late Triassic dinosauromorph ''Sacisaurus agudoensis'' (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities
https://sp.lyellcoll[...]
2013
[5]
학술지
A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet
2016
[6]
학술지
A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs
2020-08-26
[7]
웹사이트
The 'birth' of dinosaurs
https://morethanadod[...]
2023-03-15
[8]
뉴스
The Birth of Dinosaurs: Richard Owen and Dinosauria
https://blog.biodive[...]
2023-03-15
[9]
서적
The Complete Dinosaur
https://books.google[...]
Indiana University Press
2012-06-27
[10]
웹사이트
Using the tree for classification
https://evolution.be[...]
University of California
2019-10-14
[11]
뉴스
Shaking Up the Dinosaur Family Tree
https://www.nytimes.[...]
2019-10-30
[12]
학술지
A new hypothesis of dinosaur relationships and early dinosaur evolution
Nature Research
[13]
학술지
How has our knowledge of dinosaur diversity through geologic time changed through research history?
2018-02-19
[14]
학술지
How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model
2016
[15]
학술지
How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model
Royal Society
[16]
웹사이트
Most Dinosaur Species Are Still Undiscovered
https://www.national[...]
2021-06-06
[17]
학술지
Welcome
https://www.worldbir[...]
[18]
학술지
The Cretaceous–Tertiary biotic transition
Geological Society of London
[19]
웹사이트
Dinosauria: Morphology
https://ucmp.berkele[...]
University of California Museum of Paleontology
2019-10-16
[20]
서적
2000
[21]
서적
1677
[22]
서적
1677
[23]
웹사이트
Robert Plot
http://www.oum.ox.ac[...]
Oxford University Museum of Natural History
2019-11-14
[24]
웹사이트
Dinosauria
2019-11-10
[25]
서적
1994
[26]
학술지
The dinosaur ''Hadrosaurus foulkii'', from the Campanian of the East Coast of North America, with a reevaluation of the genus
https://app.pan.pl/a[...]
Institute of Paleobiology, Polish Academy of Sciences
2019-11-05
[27]
학술지
Sauropod dinosaur research: a historical review
[28]
서적
The Great Dinosaur Discoveries
A & C Black Publishers Ltd.
[29]
학술지
Results roll in from the dinosaur renaissance
[30]
서적
1986
[31]
학술지
Ten more years of discovery: revisiting the quality of the sauropodomorph dinosaur fossil record
[32]
학술지
Fossil quality and naming dinosaurs
[33]
학술지
Skeletal completeness of the non-avian theropod dinosaur fossil record
[34]
서적
The Complete Dinosaur
Indiana University Press
[35]
뉴스
That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail
https://www.nytimes.[...]
2016-12-08
[36]
논문
Reading About Dinosaurs – An Annotated Bibliography of Books
[37]
논문
Dinosaurs and the Cretaceous Terrestrial Revolution
[38]
논문
Soft Tissue Preservation in Terrestrial Mesozoic Vertebrates
[39]
논문
Integument of the iguanodont dinosaur ''Trachodon''
[40]
논문
II—On the Integument of ''Iguanodon bernissartensis'', Boulenger, and of ''Morosaurus becklesii'', Mantell
https://zenodo.org/r[...]
[41]
서적
The Hadrosaurs: Proceedings of the International Hadrosaur Symposium
Princeton University Press
[42]
논문
Exceptional preservation and the fossil record of tetrapod integument
[43]
논문
Dinosaur fossils with soft parts
http://doc.rero.ch/r[...]
[44]
논문
Vertebrate assemblages of the Jurassic Yanliao Biota and the Early Cretaceous Jehol Biota: Comparisons and implications
[45]
논문
Feathered Dinosaurs
[46]
논문
Reconstructing Vertebrate Paleocolor
[47]
논문
Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds
http://oro.open.ac.u[...]
[48]
논문
Countershading and Stripes in the Theropod Dinosaur ''Sinosauropteryx'' Reveal Heterogeneous Habitats in the Early Cretaceous Jehol Biota
[49]
논문
3D Camouflage in an Ornithischian Dinosaur
[50]
논문
Recent advances in amniote palaeocolour reconstruction and a framework for future research
[51]
논문
Interpreting melanin-based coloration through deep time: a critical review
[52]
논문
Melanosomes and ancient coloration re-examined: a response to Vinther 2015 (DOI 10.1002/bies.201500018)
[53]
논문
The Jehol Biota, an Early Cretaceous terrestrial Lagerstätte: new discoveries and implications
[54]
논문
The evolution of the modern avian digestive system: insights from paravian fossils from the Yanliao and Jehol biotas
[55]
논문
Dino DNA: the Hunt and the Hype
[56]
논문
Cells, Collagen Fibrils and Vessels in Dinosaur Bone
[57]
논문
Blood vessels and red blood cells preserved in dinosaur bones
[58]
논문
A chemical framework for the preservation of fossil vertebrate cells and soft tissues
[59]
논문
Biomolecular characterization and protein sequences of the Campanian hadrosaur ''B. canadensis''
[60]
논문
Molecular Phylogenetics of Mastodon and ''Tyrannosaurus rex''
[61]
논문
Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules
Elsevier
[62]
논문
Evidence of proteins, chromosomes and chemical markers of DNA in exceptionally preserved dinosaur cartilage
[63]
논문
Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens
[64]
논문
Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms
[65]
논문
Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs
[66]
논문
A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination
[67]
논문
Africa's oldest dinosaurs reveal early suppression of dinosaur distribution
[68]
논문
The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high-resolution chronostratigraphy, and faunal correlations
[69]
논문
A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina
Pensoft Publishers
[70]
논문
Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications
[71]
논문
The osteology of the early-diverging dinosaur ''Daemonosaurus chauliodus'' (Archosauria: Dinosauria) from the Coelophysis Quarry (Triassic: Rhaetian) of New Mexico and its relationships to other early dinosaurs
2021
[72]
논문
U-Pb age constraints on dinosaur rise from south Brazil
Elsevier
2018-05
[73]
논문
New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India
[74]
논문
The precise temporal calibration of dinosaur origins
[75]
논문
The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania
Royal Society
[76]
논문
The precise temporal calibration of dinosaur origins
[77]
논문
Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs
https://www.pure.ed.[...]
American Association for the Advancement of Science
2019-10-22
[78]
간행물
2013
[79]
논문
An elephant-sized Late Triassic synapsid with erect limbs
American Association for the Advancement of Science
[80]
뉴스
Fossil tracks in the Alps help explain dinosaur evolution
https://www.economis[...]
2018-04-19
[81]
간행물
2011
[82]
논문
A Review of the Bathornithidae (Aves, Gruiformes), with Remarks on the Relationships of the Suborder Cariamae
https://digitallibra[...]
American Museum of Natural History
2019-10-22
[83]
논문
The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay
https://www.research[...]
E. Schweizerbart
2019-10-22
[84]
간행물
2009
[85]
논문
Why sauropods had long necks; and why giraffes have short necks
[86]
웹사이트
Classification diagrams
https://equatorialmi[...]
2022-09-06
[87]
논문
Exceptional soft-tissue preservation in a theropod dinosaur from Italy
http://doc.rero.ch/r[...]
Nature Research
1998-03-26
[88]
논문
Dinosaur Biology
Annual Reviews
1995-11
[89]
간행물
2004
[90]
논문
Biology of the sauropod dinosaurs: the evolution of gigantism
Cambridge Philosophical Society
2011-02
[91]
간행물
2010
[92]
논문
Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs
http://www.miketaylo[...]
Taylor & Francis
[93]
논문
Die Skelettrekonstruktion von ''Brachiosaurus brancai''
https://paleoglot.or[...]
E. Schweizerbart
2019-10-24
[94]
conference
Reappraisal of ''Seismosaurus'', a Late Jurassic Sauropod Dinosaur From New Mexico
https://gsa.confex.c[...]
Geological Society of America
2019-10-25
[95]
논문
March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs
PLOS
[96]
논문
Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado
[97]
논문
Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals
http://www.gspauldin[...]
2019
[98]
논문
The lost titan of Cauvery
2022-06-27
[99]
논문
Body mass estimate of ''Bruhathkayosaurus'' and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales
2023-04-11
[100]
논문
Zhuchengosaurus maximus from Shandong Province
Chinese Academy of Geological Sciences
[101]
간행물
2004
[102]
간행물
2000
[103]
웹사이트
Bee Hummingbird (''Mellisuga helenae'')
https://www.birds.co[...]
Paley Media
2019-10-27
[104]
간행물
2002
[105]
간행물
2000
[106]
논문
Embryos and eggs for the Cretaceous theropod dinosaur ''Troodon formosus''
Taylor & Francis for the Society of Vertebrate Paleontology
[107]
간행물
[108]
논문
A Precocial Avian Embryo from the Lower Cretaceous of China
American Association for the Advancement of Science
[109]
논문
A drowned nesting colony of Late Cretaceous birds
https://blogs.scient[...]
Scientific American
2012-05-15
[110]
논문
A Large Accumulation of Avian Eggs from the Late Cretaceous of Patagonia (Argentina) Reveals a Novel Nesting Strategy in Mesozoic Birds
PLOS
[111]
논문
Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs
http://eprints.linco[...]
Wiley-Blackwell on behalf of the European Society for Evolutionary Biology
2018-05
[112]
논문
Evidence for gregarious behavior and age segregation in sauropod dinosaurs
http://doc.rero.ch/r[...]
Elsevier
[113]
논문
A cloacal opening in a non-avian dinosaur
Elsevier
2021-02-22
[114]
논문
Biomechanics of running indicates endothermy in bipedal dinosaurs
[115]
논문
Dinosaur Macroevolution and Macroecology
[116]
논문
Evidence for mesothermy in dinosaurs
[117]
논문
Palaeohistological Evidence for Ancestral High Metabolic Rate in Archosaurs
[118]
논문
Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution
[119]
논문
The Superiority of Dinosaurs
[120]
논문
Anatomical and Ecological Evidence of Endothermy in Dinosaurs
[121]
논문
On dinosaur growth
[122]
논문
Dinosaur paleohistology: review, trends and new avenues of investigation
[123]
논문
Evolution of endothermy: histological evidence
http://www.stuartsum[...]
[124]
서적
A Cold Look at the Warm-Blooded Dinosaurs
American Association for the Advancement of Science
[125]
논문
Growth in small dinosaurs and pterosaurs: the evolution of archosaurian growth strategies
http://doc.rero.ch/r[...]
[126]
논문
Osteohistology and growth dynamics of the Brazilian noasaurid ''Vespersaurus paranaensis'' Langer et al., 2019 (Theropoda: Abelisauroidea)
[127]
논문
Modeling growth rates for sauropod dinosaurs
http://doc.rero.ch/r[...]
[128]
논문
Oxygen isotopes from biogenic apatites suggest widespread endothermy in Cretaceous dinosaurs
http://doc.rero.ch/r[...]
[129]
논문
Oxygen and carbon isotope compositions of middle Cretaceous vertebrates from North Africa and Brazil: ecological and environmental significance
[130]
논문
Dinosaur bones: fossils or pseudomorphs? The pitfalls of physiology reconstruction from apatitic fossils
http://doc.rero.ch/r[...]
[131]
논문
Physiological, migratorial, climatological, geophysical, survival, and evolutionary implications of Cretaceous polar dinosaurs
[132]
논문
Paleoecological implications of Alaskan terrestrial vertebrate fauna in latest Cretaceous time at high paleolatitudes
[133]
논문
Polar dinosaurs
[134]
논문
Polar dinosaurs and the question of dinosaur extinction: a brief review
http://doc.rero.ch/r[...]
[135]
논문
Evolution of archosaurian body plans: skeletal adaptations of an air-sac-based breathing apparatus in birds and other archosaurs
[136]
논문
Dinosaur body temperatures determined from isotopic (13C-18O) ordering in fossil biominerals
[137]
논문
Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs
http://doc.rero.ch/r[...]
[138]
논문
Implications of an avian-style respiratory system for gigantism in sauropod dinosaurs
[139]
논문
All-time giants: the largest animals and their problems
https://www.palass.o[...]
[140]
논문
Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores
https://jeb.biologis[...]
[141]
논문
Does the ostrich (''Struthio camelus'') coprodeum have the electrophysiological properties and microstructure of other birds?
[142]
논문
Ammonia excretion by hummingbirds
[143]
논문
The Regulation of Urea-Biosynthesis Enzymes in Vertebrates
[144]
논문
The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates
[145]
논문
Pellet Regurgitation by Captive Sparrow Hawks (''Falco sparverius'')
https://sora.unm.edu[...]
2019-10-30
[146]
논문
Exceptional dinosaur fossils reveal early origin of avian-style digestion
[147]
논문
Rise of the fuzzy dinosaurs
https://www.nature.c[...]
Nature Research
2012-07-02
[148]
논문
A Jurassic ornithischian dinosaur from Siberia with both feathers and scales
http://palaeo.gly.br[...]
2016-07-27
[149]
논문
The soft tissue of ''Jeholopterus'' (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane
Royal Society
[150]
논문
Structure and homology of ''Psittacosaurus'' tail bristles
https://research-inf[...]
[151]
논문
Evolution of dinosaur epidermal structures
[152]
논문
Beta-keratin localization in developing alligator scales and feathers in relation to the development and evolution of feathers
Nuova Immagine Editrice
[153]
논문
The Early Origin of Feathers
[154]
논문
Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid, ''Shuvuuia deserti''
Wiley-Blackwell
[155]
웹사이트
"''Archaeopteryx'': An Early Bird"
https://ucmp.berkele[...]
University of California Museum of Paleontology
2019-10-30
[156]
뉴스
Study: Predatory Dinosaurs had Bird-Like Pulmonary System
https://www.ohio.edu[...]
Ohio University
2005-07-13
[157]
논문
A nesting dinosaur
Nature Research
[158]
논문
Avian Paternal Care Had Dinosaur Origin
American Association for the Advancement of Science
[159]
논문
Mass extinction of birds at the Cretaceous–Paleogene (K–Pg) boundary
[160]
논문
Time scales of critical events around the Cretaceous-Paleogene boundary
[161]
논문
The extinction of the dinosaurs
https://www.research[...]
[162]
논문
Extinctions: a paleontological perspective
[163]
논문
Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary
[164]
논문
Early evolution of modern birds structured by global forest collapse at the end-Cretaceous mass extinction
[165]
논문
Dental disparity and ecological stability in bird-like dinosaurs prior to the end-Cretaceous mass extinction
[166]
논문
Paleobiogeography and biodiversity of Late Maastrichtian dinosaurs: how many dinosaur species went extinct at the Cretaceous-Tertiary boundary?
[167]
논문
Late Cretaceous Extinctions
[168]
논문
Evidence suggesting gradual extinction of latest Cretaceous dinosaurs
[169]
논문
The gradual decline of the dinosaurs—fact or fallacy?
[170]
논문
Shape of Mesozoic dinosaur richness
http://doc.rero.ch/r[...]
[171]
서적
Late Cretaceous vertebrates from the Western Interior
[172]
논문
Ecological niche modelling does not support climatically-driven dinosaur diversity decline before the Cretaceous/Paleogene mass extinction
[173]
논문
A refined modelling approach to assess the influence of sampling on palaeobiodiversity curves: new support for declining Cretaceous dinosaur richness
[174]
논문
Dinosaurs in decline tens of millions of years before their final extinction
[175]
논문
Dinosaur diversity and the rock record
[176]
간행물
'2015'
[177]
논문
Shocked quartz in the Cretaceous-Tertiary boundary clays: Evidence for a global distribution
https://zenodo.org/r[...]
[178]
논문
Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico
[179]
논문
The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary
https://lirias.kuleu[...]
[180]
논문
The Chicxulub impact event and its environmental consequences at the Cretaceous–Tertiary boundary
[181]
논문
Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction
[182]
논문
Numerical Modeling of the Largest Terrestrial Meteorite Craters
[183]
논문
Generation and propagation of a tsunami from the Cretaceous-Tertiary impact event
[184]
논문
Survival in the first hours of the Cenozoic
http://webh01.ua.ac.[...]
2011-06-15
[185]
논문
K-Pg extinction: Reevaluation of the heat-fire hypothesis
[186]
논문
Energy, volatile production, and climatic effects of the Chicxulub Cretaceous/Tertiary impact
[187]
논문
Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction
[188]
논문
Organic matter from the Chicxulub crater exacerbated the K–Pg impact winter
[189]
논문
Production of sulphate-rich vapour during the Chicxulub impact and implications for ocean acidification
[190]
논문
Determination of rapid Deccan eruptions across the Cretaceous-Tertiary boundary using paleomagnetic secular variation: 2. Constraints from analysis of eight new sections and synthesis for a 3500-m-thick composite section
https://hal.archives[...]
[191]
논문
U-Pb constraints on pulsed eruption of the Deccan Traps across the end-Cretaceous mass extinction
[192]
논문
Deccan Traps mantle degassing in the terminal Cretaceous marine extinctions
[193]
논문
Volatile fluxes during flood basalt eruptions and potential effects on the global environment: A Deccan perspective
[194]
논문
Modeling climatic effects of carbon dioxide emissions from Deccan Traps volcanic eruptions around the Cretaceous–Paleogene boundary
[195]
논문
Selective environmental stress from sulphur emitted by continental flood basalt eruptions
http://eprints.white[...]
[196]
논문
Cretaceous-Tertiary boundary events: Mass extinctions, iridium enrichment and Deccan volcanism
[197]
논문
Multiple factors in the origin of the Cretaceous/Tertiary boundary: the role of environmental stress and Deccan Trap volcanism
[198]
간행물
Mass Extinctions Caused by Large Bolide Impacts
https://www.osti.gov[...]
Lawrence Berkeley Laboratory
2021-01-27
[199]
논문
State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact
https://escholarship[...]
[200]
논문
Triggering of the largest Deccan eruptions by the Chicxulub impact
https://escholarship[...]
[201]
서적
Trigger Effects in Geosystems
Springer
[202]
논문
Cretaceous extinctions: multiple causes
[203]
논문
Cretaceous extinctions: the volcanic hypothesis
[204]
논문
Deccan volcanism, the Chicxulub impact, and the end-Cretaceous mass extinction: Coincidence? Cause and effect
[205]
논문
Response—Cretaceous extinctions
https://lirias.kuleu[...]
[211]
서적
Disney Stories
2012
[212]
간행물
恐竜類の分岐分類におけるクレード名の和訳について
https://doi.org/10.1[...]
日本古生物学会
[213]
서적
生物分類表
岩波書店
[214]
서적
恐竜の学校
"[[ニュートン (雑誌)|ニュートンプレス]]"
[215]
간행물
Dinosaurs that did not die: evidence for Paleocene dinosaurs in the Ojo Alamo sandstone, San Juan Basin, New Mexico.
https://www.research[...]
[216]
문서
生物学的な分類群としてのトカゲ類ではなく、爬虫類やトカゲのような形状の動物全般を指す。
[217]
문서
Report on British Fossil Reptiles
John Murray
1842
[218]
서적
The Complete Dinosaur
[219]
웹사이트
恐竜は「恐ろしい竜」ではない
http://www.dino-para[...]
恐竜の楽園
2011-01-19
[220]
웹사이트
悩ましい翻訳語―科学用語の由来と誤訳
https://zokkon.haten[...]
"[[はてな (企業)|はてな]]"
2011-06-14
[221]
서적
恐竜の大常識
"[[ポプラ社]]"
[222]
문서
[223]
문서
[224]
문서
Origin and relationships of Dinosauria
University of California Press
[225]
문서
もっとも、両者ともそれぞれの系統の代表格であるとともに、保存や研究の状態が良好(現生鳥類は当然だが)であるため、互いの系統関係が揺らぐ可能性が小さい。この点で定義づけを行うには有効な選択と言える。既知の化石試料が断片的だったり、系統関係に不明な点が多い場合、定義自体が破綻してしまう。
[226]
문서
An annotated checklist of dinosaur species by continent
[227]
간행물
The logical basis of phylogenetic taxonomy
[228]
서적
恐竜の教科書 最新研究で読み解く進化の謎
"[[創元社]]"
2019-02-20
[229]
뉴스
"[[読売新聞]]"
2011-02-05
[230]
웹사이트
鳥は恐竜から進化した ー 論争についに終止符 {{!}} Nature ダイジェスト {{!}} Nature Portfolio
https://www.natureas[...]
2022-01-24
[231]
웹사이트
驚きの恐竜展を開催、もはや鳥展、米NYで
https://natgeo.nikke[...]
2022-01-24
[232]
문서
The functional origin of dinosaur bipedalism: cumulative evidence from bipedally inclined reptiles and disinclined mammals
2017
[233]
문서
[234]
문서
A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs
2007
[235]
서적
恐竜学がわかる。
"[[朝日新聞社]]"
[236]
간행물
Extraterrestrial Cause for the Cretaceous-Tertiary Extinction
[237]
서적
白亜紀に夜がくる-恐竜の絶滅と現代地質学
"[[青土社]]"
[238]
잡지
ムー (雑誌)
"[[Gakken|学研]]"
2008-01
[239]
간행물
The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary
[240]
웹사이트
恐竜やアンモナイト等の絶滅は「小惑星衝突により発生したすすによる気候変動」が原因だった
http://www.tohoku.ac[...]
東北大学公式ホームページ
[241]
뉴스
小惑星が少しずれていたら…恐竜生存? 東北大など発表
http://www.asahi.com[...]
朝日新聞
[242]
웹사이트
Asteroid that killed dinosaurs also intensified volcanic eruptions - study
http://www.theguardi[...]
2015-10-02
[243]
웹사이트
鳥は「恐竜の子孫」ではありません、恐竜なんです | ギズモード・ジャパン
https://www.gizmodo.[...]
[244]
문서
[245]
간행물
Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado
https://giw.utahgeol[...]
2018-10-19
[246]
논문
Body mass estimate of Bruhathkayosaurus and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales
https://www.idunn.no[...]
2023-04-11
[247]
웹사이트
The size of the BYU 9024 animal
https://svpow.com/20[...]
2019-06-16
[248]
서적
Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs
https://books.google[...]
Princeton University Press
[249]
뉴스
2億年前の大型恐竜、本当は重くなかった 英米研究
https://www.afpbb.co[...]
2012-06-07
[250]
기타
[251]
기타
[252]
문서
Theoretical aspects of cursorial adaptations in dinosaurs
[253]
뉴스
Dinosauromorph research sheds light on dinosaur evolution
CBC NEWS
2015
[254]
문서
What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs
[255]
기타
[256]
기타
[257]
기타
[258]
기타
[259]
기타
[260]
기타
[261]
서적
トリビアの泉〜へぇの本〜 3
講談社
[262]
웹사이트
Vinther, J., Briggs, D. E. G., Prum, R. O. and Saranathan, V. 2008 The colour of fossil feathers.
http://www.jakobvint[...]
[263]
웹사이트
恐竜の色はどのようにして復元するの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館
https://www.dinosaur[...]
2022-01-22
[264]
웹사이트
Yale Scientists First to Reveal Flamboyant Colors of a Dinosaur’s Feathers
https://news.yale.ed[...]
2010-02-04
[265]
문서
鳥類の起源参照
[266]
서적
系統樹をさかのぼって見えてくる進化の歴史 僕たちの祖先を探す15億年の旅
ベレ出版
[267]
웹사이트
骨「年輪」、草食哺乳類にも=恐竜冷血説を否定-欧州チーム
http://www.jiji.com/[...]
[268]
웹사이트
Seasonal bone growth and physiology in endotherms shed light on dinosaur physiology
https://www.nature.c[...]
Nature Publishing Group
2012-07-19
[269]
기타
[270]
웹사이트
恐竜はどうやって巣を作るの?卵はどんなふうに並べるの?
https://www.dinosaur[...]
2023-11-16
[271]
웹사이트
恐竜の繁殖戦略と孵化方法の多様性
https://dinosaur.blu[...]
2023-11-16
[272]
웹사이트
Plot R. (1677) The natural history of Oxford-shire : Being an essay toward the natural history of England. Chap. V. Of formed stones.
http://www.biodivers[...]
[273]
문서
真鍋真 (2005)「人間と恐竜との出会い」. 恐竜博2005-恐竜から鳥への進化. 図録
[274]
웹사이트
Buckland W. (1824) Notice on the Megalosaurus or great fossil lizard. Stonesfield. Transactions of the Geological Society of London 1(2):390-396.
http://www.biodivers[...]
[275]
논문
Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex
[276]
문서
"Archaeopteryx litographica (Vogel-Feder) und Pterodactylus von Solenhofen." Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: 678-679.
[277]
문서
"On the animals which are most nearly intermediate between birds and reptiles". Annals of the Magazine of Natural History 4 (2): 66–75.
[278]
서적
The Origin of Birds
Witherby
[279]
서적
Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds
https://books.google[...]
JHU Press
[280]
서적
The Dinosauria
University of California Press
[281]
논문
A new type of small theropod dinosaur from the Navajo Sandstone of Arizona
[282]
논문
The ancestry of birds
[283]
간행물
Saurischian monophyly and the origin of birds
[284]
서적
Classification and evolution of the dinosaur groups
St. Martin's Press
[285]
논문
Developmental patterns and the identification of homologies in the avian hand.
[286]
논문
1,2,3 = 2,3,4: a solution to the problem of the homology of the digits in the avian hand.
[287]
논문
"The evolution of HoxD-11 expression in the bird wing: insights from Alligator mississippiensis."
http://www.plosone.o[...]
[288]
논문
Embryological evidence identifies wing digits in birds as digits 1, 2, and 3.
[289]
논문
A Jurassic ceratosaur from China helps clarify avian digital homologies.
[290]
논문
Ontogeny of cranial epi-ossifications in Triceratops.
[291]
논문
Torosaurus is not Triceratops: ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy.
[292]
서적
[293]
문서
MIT’s New Evidence on Dinosaur Evolution
2020
[294]
서적
[295]
서적
哺乳類型爬虫類 ヒトの知られざる祖先
朝日新聞社
[296]
논문
Nipponosaurus Sachalinensis : A New Genus and Species of Trachodont Dinosaur from Japanese Saghalien
https://hdl.handle.n[...]
The Hokkaido Imperial University
1936-07
[297]
간행물
北海道中川町の恐竜化石を新属新種「パラリテリジノサウルス・ジャポニクス」と命名~恐竜類テリジノサウルス科の爪の進化~
https://www.hokudai.[...]
北海道大学、岡山理科大学、中川町エコミュージアムセンター
2022-05-09
[298]
논문
日本産の中生代恐竜化石目録 2022年版
https://www.gmnh.pre[...]
群馬県立自然史博物館
2023-03
[299]
웹사이트
通称・むかわ竜、学名が「カムイサウルス・ジャポニクス」に決定 新属新種の恐竜だと正式に認定される
https://nlab.itmedia[...]
2021-01-07
[300]
웹사이트
Neotheropoda
http://archosaur.us/[...]
2009-11-17
[301]
웹사이트
新種化石06「桑島化石壁産出の恐竜化石に学名がつきました」|白山市公式ホームページ
https://www.city.hak[...]
2023-03-22
[302]
뉴스
福井の恐竜化石発掘調査30年 きっかけの発見者、松田亜規さん
https://mainichi.jp/[...]
2021-02-14
[303]
논문
A new iguanodontian (Dinosauria; Ornithopoda), form the lower Cretaceous Kitadani Formation of Fukui Prefecture, Japan
[304]
웹사이트
フクイラプトルという学名になりました
https://www.dinosaur[...]
福井県立恐竜博物館
2020-06-27
[305]
논문
'Fukuititan nipponensis, a new titanosauriform sauropod from the Early Cretaceous Tetori Group of Fukui Prefecture, Japan'
[306]
웹사이트
勝山市の発掘現場から発見された恐竜に新しい学名が付けられました
http://www.dinosaur.[...]
福井県立恐竜博物館
2017-07-02
[307]
웹사이트
新恐竜はティラノサウルスもどき 98年から発掘の化石、福井 {{!}} 全国のニュース
https://www.fukuishi[...]
2023-09-08
[308]
논문
A new genus and species of carnivorous dinosaur from the Lower Cretaceous Kwanmon Group, northern Kyushu.
[309]
웹사이트
恐竜博物館の調査研究情報、長崎初の大型獣脚類(肉食恐竜)の化石について
http://www.dinosaur.[...]
福井県立恐竜博物館
2017-07-02
[310]
웹사이트
国内最古級の恐竜化石発見、熊本 獣脚類の肋骨か - 共同通信 | This Kiji
https://web.archive.[...]
共同通信
2019-01-29
[311]
간행물
"Report on British Fossil Reptiles." Part II
[312]
웹사이트
Liddell-Scott-Jones Lexicon of Classical Greek
http://www.perseus.t[...]
2008-08-05
[313]
서적
Preface
Indiana University Press
[314]
서적
Origin and relationships of Dinosauria
University of California Press
[315]
논문
An annotated checklist of dinosaur species by continent.
[316]
서적
Basal Avialae
University of California Press:Berkeley
2004
[317]
서적
Dinosaurs: The Encyclopedia
McFarland & Co.
1997
[318]
서적
The Dinosaur Data Book
Avon Books
1990
[319]
서적
Nondinosaurian vertebrates of the Mesozoic
Indiana University Press
1997
[320]
논문
Large Mesozoic mammals fed on dinosaurs
2005
[321]
논문
Estimating the Diversity of Dinosaurs
2006
[322]
논문
China and the lost worlds of the dinosaurian era
1995
[323]
뉴스
Will the real dinosaurs stand up?
http://news.bbc.co.u[...]
BBC
2008-09-17
[324]
논문
The Cretaceous–Tertiary biotic transition
http://findarticles.[...]
1997
[325]
문서
포유류와 같은 단궁강
[326]
서적
Classification and evolution of the dinosaur groups
St. Martin's Press
2000
[327]
논문
A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil
1999
[328]
웹인용
공룡 사이즈
http://dml.cmnh.org/[...]
2021-08-15
[329]
웹사이트
공룡과 포유류
http://microlnx.com/[...]
[330]
문서
예전의 분류는 조각류·검룡류·곡룡류·각룡류
[331]
서적
공룡의 세계
계몽사
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com