메가라크네
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
메가라크네는 아르헨티나에서 발견된 멸종된 유절지동물로, 한때 거대한 거미로 오인되었으나, 현재는 바다 전갈로 분류된다. 1980년 최초 발견 당시에는 거미류로 잘못 분류되어, 50cm에 달하는 다리 폭을 가진 거대한 거미의 복원 모형이 전시되기도 했다. 하지만 2005년, 더 완전한 표본이 발견되면서 유립테루스목, 즉 바다 전갈로 재분류되었으며, 현재는 미크테로프테리드인 Woodwardopterus scabrosus와 유사한 종으로 여겨진다.
더 읽어볼만한 페이지
- 석탄기의 동물 - 메가네우라
메가네우라는 석탄기 후기에 서식했으며 날개 길이가 70cm가 넘는 거대한 잠자리류 곤충 속으로, 높은 산소 농도와 포식자 부재 등으로 인해 큰 크기를 가졌고 다른 곤충이나 양서류를 잡아먹는 포식자였을 것으로 추정된다. - 석탄기의 동물 - 아르트로플레우라
아르트로플레우라는 석탄기와 페름기에 번성했던 멸종된 다지류로, 넓은 등판과 3개의 엽을 가진 외형을 특징으로 하며, 육상에서 생활하며 초식 또는 부식성 식성을 가졌을 것으로 추정된다. - 광익목 - 바다전갈
바다전갈은 고생대 오르도비스기부터 페름기까지 살았던 멸종된 절지동물로, 머리가슴, 협각, 다리, 후체로 나뉘며 서새와 "Kiemenplatten"이라는 호흡기를 가졌고, 일부는 2.5m에 달하는 큰 크기로 바다와 담수에서 서식했다. - 광익목 - 믹소프테루스
믹소프테루스는 중생대 실루리아기에 살았던 최대 75cm 크기의 육식성 바다전갈로, 단단한 외골격과 헤엄다리를 가지며 매복 사냥을 했을 것으로 추정된다. - 1980년 기재된 화석 분류군 - 제피로사우루스
제피로사우루스는 백악기 초 북아메리카 서부에 서식한 힙실로포돈과 공룡으로, 가파른 얼굴과 솟아오른 혹이 특징이며 오로드로메우스와 근연 관계를 가지며 굴을 파는 습성이 있었을 가능성이 제기된다. - 1980년 기재된 화석 분류군 - 민미
민미는 호주에서 발견된 백악기 초식성 갑옷 공룡으로, 척추를 따라 뼈 판을 가지고 있었으며, 쿤바라사우루스로 분류되기도 하였고, 유리포다 또는 안킬로사우루스류로 분류되기도 한다.
| 메가라크네 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Megarachne servinei |
| 명명자 | Hünicken, 1980 |
| 크기 | 약 54cm |
| 시대 | Gzhelian (약 3억 370만 년 전 ~ 2억 9890만 년 전) |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 절지동물문 |
| 아문 | 협각아문 |
| 강 | 퇴구강 |
| 목 | 우미사소리목(광익목) |
| 아목 | 아시나가우미사소리아목 |
| 상과 | 미크테로푸스상과 |
| 과 | 미크테롭투스과 |
| 속 | 메가라크네속 |
| 종 | 메가라크네 세르비네이 |
2. 역사
메가라크네는 협각류에 속하는 절지동물의 한 속이다. 이 생물의 정체에 대한 학계의 이해는 화석 발견과 연구를 통해 극적으로 변화해왔다.[17] 1980년 아르헨티나에서 고생물학자 마리오 휘니켄이 처음 발견하여 기술했을 때, 메가라크네는 등갑 형태 등을 근거로 거대한 원시적인 거미의 일종으로 잘못 분류되었다.[5][16] 이 잘못된 분류로 인해, 몸길이 33.9cm, 다리 폭 50cm에 달하는 역대 가장 큰 거미로 알려지면서 큰 대중적 관심을 받았다.[7][6]
하지만 일부 거미학자들은 처음부터 이 분류에 의문을 제기했으며,[8] 최초 기술자인 휘니켄 본인도 일부 특징이 일반적인 거미와 다르다는 점을 인정했다.[7] 결정적인 전환점은 2005년 같은 지역에서 두 번째 표본이 발견되면서 마련되었다.[5] 영국의 고생물학자 폴 A. 셀던이 이끈 연구팀(휘니켄 포함)은 새로운 화석과 기존 모식 표본을 종합적으로 재검토하여, 메가라크네가 거미가 아니라 유라프테리드(흔히 바다 전갈로 불리는 그룹)의 일종이라는 결론을 내렸다.[2][6] 이 연구를 통해 메가라크네는 '거대 거미'라는 오해에서 벗어나 남아메리카 석탄기 지층에서 발견된 중요한 바다 전갈 화석 중 하나로 올바르게 자리매김하게 되었다.[10][17]
2. 1. 거대 거미로의 오해 (1980년대~1990년대)

''Megarachne servinei''는 1980년 아르헨티나의 고생물학자 마리오 휘니켄(Mario Hünicken)에 의해 처음 기술되었다.[16][5] 속명은 "거대한"을 의미하는 고대 그리스어 μέγας|megasgrc와 "거미"를 의미하는 라틴어 ''arachne''를 결합하여 "거대한 거미"라는 뜻이다. 모식 표본 (CORD-PZ 2110)은 아르헨티나 산루이스 주의 바호 데 벨리즈 지층(Bajo de Véliz Formation) 팔레로 멤버(Pallero Member)에서 발견되었으며, 이 지층은 석탄기 후기 그젤리안 시대에서 페름기 초기 아셀리안 시대 사이(약 3억 년 전[18])의 것으로 추정된다.[5][6][18] 현재 표본은 코르도바 국립 대학교 고생물학 박물관에 보관되어 있다.[5]
이 모식 표본은 등갑(carapace), 처음 두 개의 배판(tergite), 세 개의 부분적인 부속지, 그리고 콕사(coxa)로 추정되는 부분을 포함하고 있었다.[10][5] 휘니켄은 등갑의 형태, 특히 머리 중앙의 두 눈 사이에 위치한 15mm 폭의 둥근 눈 결절(ocular tubercle)과 첫 번째 체절 뒤의 둥근 구조(이를 "적당히 털이 많은" 배로 해석)를 근거로 이 표본을 거미 중에서도 Mygalomorphae(타란툴라 등이 속한 그룹)으로 잘못 분류했다. 그는 X선 마이크로 단층 촬영을 통해 흉판(sternum), 입술(labium), 콕사, 협각 송곳니 등 숨겨진 구조들도 거미의 특징으로 해석했다.[5][16]
이 화석이 거미의 것이라는 가정 하에 몸길이 33.9cm, 다리를 포함한 폭이 50cm로 추정되면서, ''Megarachne servinei''는 당시 알려진 가장 큰 거미인 골리앗새잡이거미(''Theraphosa blondi'', 다리 폭 약 30cm)를 훨씬 능가하는, 역대 가장 큰 거미로 여겨지게 되었다.[17][7] 이 타이틀 덕분에 메가라크네는 빠르게 대중적인 관심을 얻었고, 휘니켄의 설명을 바탕으로 거대한 거미의 모습으로 복원된 모형이 전 세계 박물관에 전시되고 다큐멘터리 등에도 등장했다.[7][6][19]
하지만 일부 거미학자들은 이 해석에 의문을 제기했다. 1989년 쉬어(Shear)와 동료들은 메가라크네가 거미강에 속하기는 하지만, 알려지지 않은 목(order)이나 ricinuleid일 가능성을 제시했다.[8] 휘니켄 자신도 특이한 큐티클 장식, 등갑이 봉합선에 의해 앞뒤로 나뉜 점, 그리고 주걱 모양의 협각(chelicerae) 등 기존 거미들과 다른 형태학적 특징들을 인정했다. 특히 주걱 모양의 협각은 다른 거미에게서는 발견되지 않는 매우 이례적인 특징이었다.[7][17] 그러나 모식 표본이 오랫동안 은행 금고에 보관되어 다른 연구자들이 직접 접근하기 어려웠고, 오직 석고 모형만 연구할 수 있었기 때문에[7][17] 초기 해석에 대한 검증이 제대로 이루어지지 못했다. 이러한 논란 속에서 메가라크네는 1980년대와 1990년대 동안 거대한 고대 거미로 잘못 알려지게 되었다.
2. 2. 바다전갈로의 재분류 (2005년 이후)
''Megarachne''를 거미로 식별하는 것에 대해 일부 거미학자들은 의문을 제기했다. 예를 들어, Shear와 동료들(1989)은 ''Megarachne''가 거미강에 속하지만 "미상의 목 또는 리키눌레이목(ricinulei)을 나타낼 수 있다"고 언급했다.[8] 처음 기술한 휘니켄 자신조차도 화석의 형태가 거미류와 일치하지 않는 몇 가지 특징을 인정했다. 여기에는 특이한 큐티클 장식, 봉합선에 의해 앞부분과 뒷부분으로 나뉜 두흉갑, 그리고 주걱 모양(넓고 둥근 끝을 가짐)의 협각이 포함되었다. 휘니켄은 알려진 거미 중에는 주걱 모양의 협각을 가진 종이 없다는 점을 이상하게 여겼다.[7] 그러나 모식 표본은 당시 은행 금고에 보관되어 있었기 때문에 다른 고생물학자들은 석고 모형만 연구할 수 있었다.[7]
2005년, 모식 표본이 발견된 곳과 동일한 지역 및 지질학적 지평선에서 두 번째 표본이 발견되었다. 이 표본은 부분과 상대물(압축 화석의 양면)로 구성되어 있었으며, 몸의 앞부분과 네 번째 다리 쌍에서 나왔을 가능성이 있는 콕사의 일부를 보존하고 있었다.[5] 이 새로운 화석은 모식 표본에서 "협각"과 "복부"로 해석되었던 부분은 없었지만, 대신 두흉갑의 양쪽 가장자리와 다리의 기부가 보존되어 있었다.[17]
이 새로운 발견을 바탕으로, 영국의 고생물학자이자 거미학자인 폴 A. 셀던이 이끄는 연구팀(최초 기술자인 휘니켄과 아르헨티나 거미학자 호세 A. 코론카도 참여했다)은 모식 표본을 재검토했다.[2][6][17] 2005년에 발표된 연구 결과, 그들은 ''Megarachne servinei''가 거미가 아니라 큰 유라프테리드(일반적으로 "바다 전갈"로 알려진 그룹)의 일원이라고 결론지었다.[2][6] 휘니켄이 처음 ''Megarachne''를 거미로 잘못 식별했지만, 거미류와 바다 전갈 모두 협각류에 속하며 밀접한 관련이 있기 때문에[9] 그의 해석이 완전히 근거 없는 것은 아니었다.[17]
연구팀은 다른 유라프테리드와의 형태학적 비교를 통해 ''Megarachne''가 또 다른 큰 페름기-석탄기 유라프테리드인 미크테로프테리드과(Mycteroptidae)의 ''Woodwardopterus scabrosus''와 가장 유사하다고 판단했다. ''Woodwardopterus'' 역시 단일 표본만 알려져 있다.[2][17] 셀던과 동료들은 비록 알려진 표본이 단 두 개뿐이지만, ''Megarachne''가 남아메리카 석탄기 퇴적물에서 발견된 유라프테리드 중 가장 완전한 형태라고 결론지었다.[10]
재분류 과정에서 모식 표본에서 거미의 특징으로 잘못 해석되었던 부분들의 실제 정체가 밝혀졌다.[17]
| 휘니켄의 해석 (거미 특징) | 셀던 외 연구팀의 재해석 (바다 전갈 특징) |
|---|---|
| "협각" | 두흉갑의 전반부 |
| "양 협각의 경계" | 두흉갑 전반 중앙의 융기선 |
| "두흉갑" | 두흉갑의 후반부와 후체(opisthosoma) 첫 번째 등판(tergite) |
| "두흉갑 후방의 홈" | 두흉갑과 후체의 경계 |
| "집약된 수 쌍의 홑눈" | 각 1쌍의 홑눈과 겹눈 |
| "후체(복부)" | 후체 두 번째 등판 |
''메가라크네''는 비교적 희귀한 바다전갈 분류군인 스틸로누리나 아목에 속한다. 스틸로누리나 내에서 ''메가라크네''는 상과인 믹테로포이데아(Mycteropoidea)에 분류되며, 이 상과에는 밀접한 관계의 속인 ''우드워드옵테루스''(Woodwardopterus)와 ''믹테롭스(Mycterops)''가 포함된 믹테로피다에과(Mycteropidae)가 속한다.[3][21][15][23]
2005년 영국 다큐멘터리 ''괴물과의 산책'' 제작 당시, ''메가라크네''는 석탄기 부분을 다루는 에피소드에서 고양이 크기의 파충류인 ''페트로라코사우루스''를 사냥하는 거대한 타란툴라 같은 거미로 등장할 예정이었다. 이 재구성은 시리즈 제작 당시 메가라크네에 대해 알려진 내용을 면밀히 따른 것이었다. 그러나 제작이 한창 진행된 후에 이 속이 실제로는 유르프테리드라는 사실이 밝혀졌고, 이때는 재구성을 수정하기에는 너무 늦은 상황이었다. 해당 장면들은 그대로 유지되었지만, 거대한 거미는 실제로 존재하는 원시 거미 아목 중심거미아목에 속하는, 특정되지 않은 종으로 이름이 바뀌었다. 그러나 이 아목에 속하는 속들은 프로그램에 등장하는 거미보다 훨씬 작고 외형도 상당히 달랐다.[6]
[1]
논문
Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
ScienceBlogs
2010-04-23
또한, 1980년 휘니켄이 X선 마이크로 단층 촬영으로 발견했다고 보고한 모식 표본 뒷면의 흔적들은 화석 자체의 구조가 아니라, 단순히 화석 아래 암석의 갈라진 틈이었음이 지적되었다.[17]
3. 형태
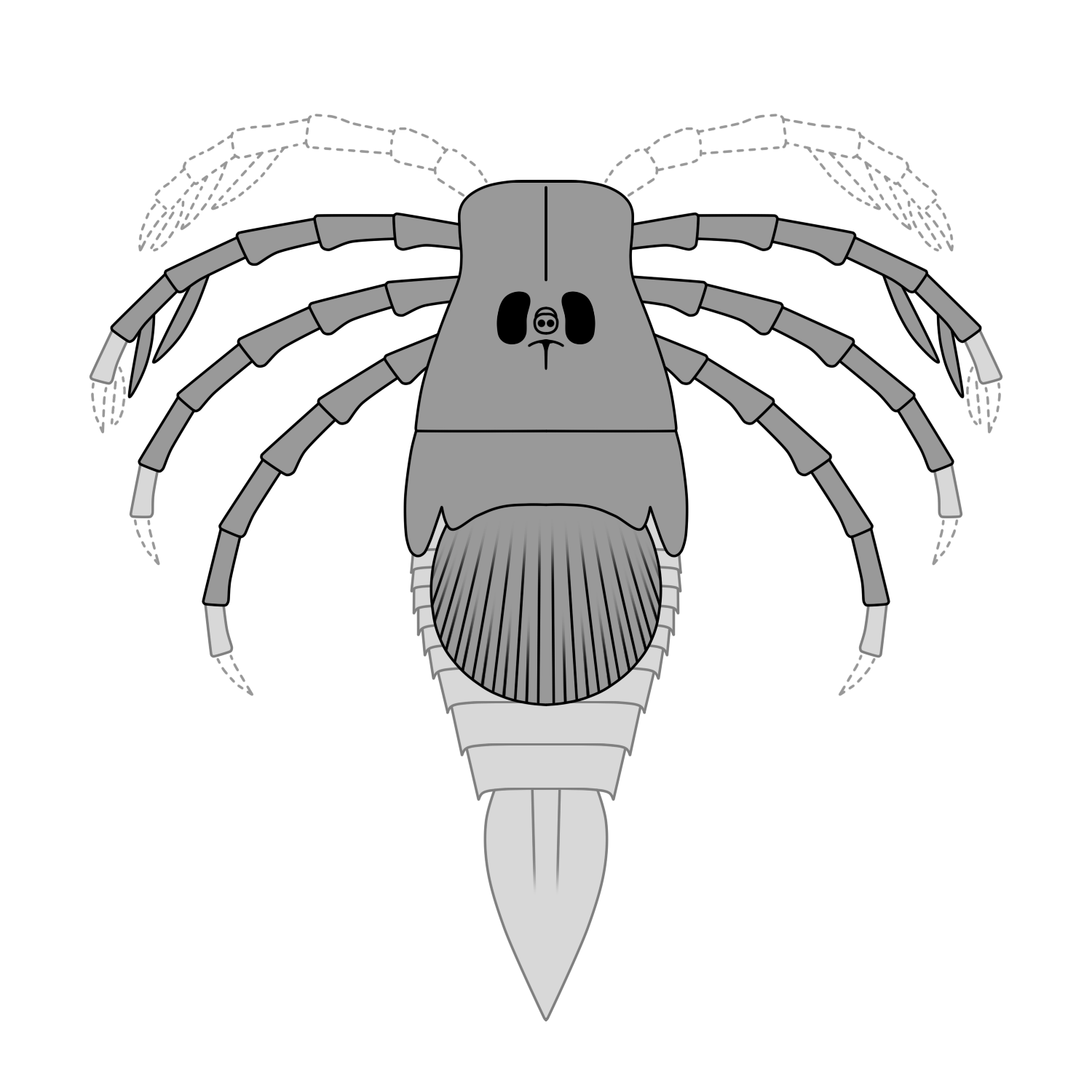
알려진 화석에 따르면 ''메가라크네''의 몸길이는 54cm에 달하는 것으로 추정된다.[20] 이는 절지동물로서는 큰 편에 속하지만, 1.5m를 넘기도 하는 가까운 친척 ''히베르토프테루스''[1] 등 다른 바다전갈류에 비하면 작은 크기였다.
처음에는 거대한 거미로 잘못 알려지기도 했으나,[2] 여러 특징들이 ''메가라크네''가 바다전갈류임을 명확히 보여준다. 특히 갑각(머리를 덮는 외골격) 표면에 융기된 초승달 모양의 무늬와 전체에 걸쳐 중단 없이 이어지는 분할 능선(무크론) 형태의 큐티클 조각은 바다전갈류의 전형적인 특징이다.[2] 몸 표면에는 작은 융기가 빽빽하게 나 있다.[17] 현재까지 알려진 화석 부위는 배갑, 후방 다리, 그리고 후체 앞부분의 등판 두 개뿐이다.[17]
머리가슴 부분의 배갑은 세로로 길며 좌우가 약간 오목한 형태이다. 한 쌍의 겹눈과 홑눈은 배갑 중앙에 모여 있으며,[17] 겹눈은 대략 사다리꼴 모양이다.[4] 눈 앞쪽에는 융기선이 하나 있고, 뒤쪽으로는 T자 모양의 융기선이 배갑의 정중선을 따라 이어진다.[17]
머리가슴의 6쌍의 부속지 중에서는 뒤쪽 3쌍의 다리만 화석으로 발견되었다. 앞의 3쌍 부속지인 협각, 촉지(제1 다리), 그리고 제2 다리는 알려져 있지 않다.[17][21] 알려진 다리들은 턱기부가 있는 기절 이후의 마디들이 길고 홈이 파여 있으며, 각 마디 끝은 약간 부풀어 있다.[17][21] 다리 끝부분(마지막 마디)은 발견되지 않았다.[17] 이 중 첫 번째 쌍(전체 중 네 번째 다리)의 중간 마디에는 칼날 모양의 돌기가 나 있다.[17] 이러한 칼날 모양 구조는 먹이 섭취 방식과 관련이 있었을 것으로 추정되는데, 늪이나 강바닥의 부드러운 퇴적물을 앞다리 칼날로 긁어 작은 무척추동물을 포획하는 방식(스윕 피딩, sweep-feeding)으로 먹이를 섭취했을 가능성이 있다.[2]
후체(복부)는 앞부분의 두 개 등판만 알려져 있다. 첫 번째 등판은 배갑에 붙어 있으며 가로로 넓고, 양옆은 뒤쪽을 향해 잎 모양으로 돌출되어 있다.[17] 두 번째 등판은 크고 둥근 원반 모양으로 튀어나와 있는 것이 특징이며, 표면에는 근육 부착 흔적이 있다.[17] 이 구조의 기능은 아직 밝혀지지 않았다.[2]
전체적인 외형은 다른 마이케르옵테류 바다전갈과 매우 유사했을 것으로 보인다. 이들은 전두부(머리 앞부분)의 포물선 모양 가장자리, 고대 로마의 검(글라디우스)을 닮은 칼 모양의 미부(꼬리)와 그 아래쪽의 쌍을 이루는 용골 모양 돌출부[3] 등의 특징을 공유한다.[4] (다만, ''메가라크네'' 화석에서는 꼬리 부분이 발견되지 않아 복원도에서는 근연종인 ''우드워드옵테루스''를 참고하여 그려졌다.)
4. 생태

알려진 ''메가라크네'' 화석은 몸길이가 54cm 정도로 나타난다. 이는 절지동물로서는 큰 편이지만, 다른 대형 전갈류에 비하면 작은 크기이다. 예를 들어 비교적 가까운 친척인 ''히베르토프테루스''는 몸길이가 1.5m를 넘을 수 있었다.[1]
''메가라크네''는 부속지(다리)에 칼날 모양의 구조를 가지고 있었다. 이를 이용해 늪이나 강과 같은 수생 생태계의 부드러운 퇴적물을 긁어 작은 무척추동물을 포획하여 먹는 '스윕피딩'(sweep-feeding) 방식으로 먹이를 섭취했을 것으로 추정된다.[2] 이러한 먹이 전략은 다른 마이케르옵테루스류에서도 흔하게 나타난다.[3]
''메가라크네''의 두 표본은 모두 아르헨티나의 바호 데 벨리즈 지층에서 발견되었으며, 후기 석탄기인 젤리안 시대로 연대가 추정된다.[5][6] 바호 데 벨리즈 지층의 환경은 유르프테리드, 특히 헤엄치는 유르프테리나 아목의 일반적인 서식 환경과는 달리, 홍수터의 담수 환경이었던 것으로 보인다.[5][12] 퇴적학적 증거 역시 이들의 서식 환경이 해양이 아니었음을 시사한다.[17] 화석과 함께 발견된 식물 잔해는 당시 식물상이 다양한 종류의 씨고사리와 부리고사리류가 주를 이루었음을 보여준다.[13]
''메가라크네''가 살았던 시기, 아르헨티나를 포함한 남아메리카는 고대 초대륙 곤드와나의 일부였다. 이 시기 곤드와나는 북쪽의 다른 대륙들과 융합하여 판게아를 형성하기 시작했다.[13] 바호 데 벨리즈 지층에서는 ''메가라크네'' 외에도 ''리가토프테라''와 같은 다양한 화석 곤충들이 발견되지만, 담수 포식자였던 ''메가라크네''가 이들을 직접 먹이로 삼았을 가능성은 낮다. 대신, 앞서 언급된 스윕피딩 방식으로 강의 부드러운 퇴적물 속 작은 무척추동물을 주된 먹이로 삼았을 것이다.[2]
후기 석탄기는 초기 석탄기의 비교적 따뜻했던 기후와 달리 전반적으로 추운 시기였다. 이러한 기후 변화는 대기 중 이산화 탄소(CO2) 농도 감소와 높은 산소 농도와 관련이 있을 수 있다. 아르헨티나가 위치했던 남반구는 현대의 북극 및 남극과 유사한 거대한 대륙 빙상이나 여러 개의 작은 빙하 중심지로 빙하 작용을 겪었을 가능성이 있다. 빙상의 확산은 해수면 변동에도 영향을 미쳤다. 당시 식물상은 다양성은 낮았지만 곤드와나 전역에 걸쳐 비교적 균일하게 분포했다. 주요 식물로는 ''노토라코프테리스'', ''트리필롭테리스'', ''보트리키옵시스''와 같은 씨고사리 나무와 ''말란자니아'', ''레피도덴드롭시스'', ''붐부덴드론''과 같은 부리고사리류 나무가 있었다. 발견된 식물 화석들은 특정 시기에 몬순의 영향을 받았을 가능성도 시사한다.[13]
그러나 ''메가라크네''의 구체적인 생태, 예를 들어 크고 둥근 두 번째 후체부(복부의 두 번째 등쪽 분절)의 기능 등에 대해서는 여전히 불명확한 점이 많다.[2][17]
5. 분류
2005년, 영국의 고생물학자이자 거미학자인 폴 A. 셀던이 이끄는 연구팀은 새로운 화석 발견을 바탕으로 기존 모식 표본을 재검토하여, ''Megarachne servinei''가 거미가 아니라 큰 바다전갈(유립테루스류)이라고 결론지었다.[2][6] 다른 유립테루스와의 형태학적 비교를 통해 ''메가라크네''는 같은 과의 ''우드워드옵테루스''와 가장 유사하다는 것이 밝혀졌다.[2] 셀던과 동료들은 ''우드워드옵테루스''의 두 번째 등판 화석과 ''메가라크네''의 화석을 비교하여 거의 동일하다는 것을 발견했지만, 두 가지 주요 차이점을 확인했다. ''메가라크네''는 등판과 등딱지의 미세 돌기가 ''우드워드옵테루스''보다 덜 조밀하게 배열되어 있으며, 등딱지 앞쪽 중앙 부분에 ''우드워드옵테루스''에는 없는 돌출부가 있다.[2]
믹테로피다에 과에 속하는 네 개의 속 중 ''믹테롭스'', ''우드워드옵테루스'', ''메가라크네'' 세 속은 형태와 표본 크기에 따라 서로 다른 개체 발생 단계(동물의 생애 동안의 다른 발달 단계)를 나타낼 수 있다는 주장이 제기되었다.[3][17] 이 해석에 따르면, ''메가라크네''의 덜 조밀한 미세 돌기는 ''우드워드옵테루스''보다 상당히 큰 크기 때문일 수 있으며, 가장 작은 속인 ''믹테롭스''는 등딱지와 등판에 훨씬 더 조밀한 장식을 가지고 있어 가장 어린 개체 발생 단계일 수 있다.[2] 만약 이 세 속이 실제로 동일한 동물의 다른 성장 단계를 나타낸다면, 명명 우선권 원칙에 따라 1886년에 가장 먼저 명명된 ''믹테롭스''가 유효한 속명이 될 것이다.[11]
람스델과 동료들의 2010년 연구에 따른 분기도 분석에 따르면,[3] ''메가라크네''는 넓은 후체 제1-2 등판을 공유파생형질로 가지며[17][21], 믹테로피다에 과 내에서 ''믹테롭스'' 속과 가장 가까운 관계에 있는 것으로 여겨진다.[21][23] 이들은 함께 ''우드워드옵테루스'' 속과 자매군을 형성한다. 스틸로누리나 아목 내에서 믹테로피다에 과는 히베르토프테리데 과와 함께 믹테로포이데아 상과를 이루며, 이는 드레파노프테루스 속과 자매군 관계이다. 믹테로포이데아 상과는 코코모프테로이데아 상과(코코모프테리데 과와 하디이옵테리데 과 포함)와 함께 더 큰 분기군을 형성한다. 이 분기군은 다시 스틸로누로이데아 상과 및 렌옵테리데 과와 단계적으로 더 큰 그룹을 형성한다.
2020년 기준으로, 메가라크네 속에는 ''Megarachne servinei''라는 한 종만이 포함된다.[15]
6. 대중 문화
참조
[2]
논문
The true identity of the supposed giant fossil spider Megarachne – 4. Discussion
ScienceBlogs
2005-03-22
[3]
논문
The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida)
[4]
서적
Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata
[5]
논문
The true identity of the supposed giant fossil spider Megarachne – 3. Results
ScienceBlogs
2005-03-22
[6]
논문
"Megarachne', the Giant Spider That Wasn't"
http://scienceblogs.[...]
ScienceBlogs
2012-11-16
[7]
논문
The true identity of the supposed giant fossil spider Megarachne – 1. Introduction
ScienceBlogs
2005-03-22
[8]
논문
A Devonian spinneret: early evidence of spiders and silk use
1989
[9]
논문
Revised systematics of Palaeozoic 'horseshoe crabs' and the myth of monophyletic Xiphosura
2013-01-01
[10]
논문
The true identity of the supposed giant fossil spider Megarachne – Abstract
ScienceBlogs
2005-03-22
[11]
간행물
A summary list of fossil spiders and their relatives
http://www.wsc.nmbe.[...]
Natural History Museum Bern
2018
[12]
웹사이트
Bajo de Veliz (CORD collection), Carboniferous of Argentina
https://paleobiodb.o[...]
2021-12-17
[13]
논문
Response of Late Carboniferous And Early Permian Plant Communities To Climate Change
https://repository.u[...]
[14]
논문
'Rigattoptera ornellasae' n. g. n. sp., a new fossil insect from the Carboniferous of Argentina
https://www.research[...]
1996
[15]
간행물
A summary list of fossil spiders and their relatives
https://wsc.nmbe.ch/[...]
Natural History Museum Bern
2020
[16]
논문
A giant fossil spider (''Megarachne servinei'') from Bajo de Véliz, Upper Carboniferous, Argentina
[17]
논문
The true identity of the supposed giant fossil spider ''Megarachne''
https://www.ncbi.nlm[...]
[18]
웹사이트
Megarachne
http://fossilworks.o[...]
Paleobiology Database
2012-11-16
[19]
웹사이트
"Megarachne', the Giant Spider That Wasn’t"
http://scienceblogs.[...]
ScienceBlogs
2012-11-16
[20]
논문
Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
https://www.research[...]
2010-04-23
[21]
논문
The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida)
https://doi.org/10.1[...]
2010-03-15
[22]
서적
節足動物の多様性と系統
https://www.worldcat[...]
裳華房
2008
[23]
논문
From success to persistence: Identifying an evolutionary regime shift in the diverse Paleozoic aquatic arthropod group Eurypterida, driven by the Devonian biotic crisis: CHANGING EVOLUTIONARY REGIMES DURING THE DEVONIAN
https://www.research[...]
2017-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com