바다전갈
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
바다전갈은 멸종된 절지동물로, 학명은 Eurypterida이며, 넓은 날개를 의미하는 고대 그리스어에서 유래되었다. 영어로는 sea scorpion, 중국어로는 광시후, 일본어로는 우미사소리 등으로 불린다. 몸은 두흉부와 후체구로 나뉘며, 두흉부에는 협각과 다리가, 후체구에는 생식기, 블라트퓌세, 미절 등이 존재한다. 바다전갈은 크게 광익아목과 긴다리바다전갈아목으로 구분되며, 수영 노의 유무로 나뉜다. 과거에는 말발굽게와 함께 구갑류로 분류되었으나, 현재는 거미류와 더 가까운 Sclerophorata로 분류된다. 초기 오르도비스기부터 실루리아기에 번성했으며, 데본기 후기 대멸종으로 인해 많은 종이 멸종되었다. 바다전갈에 대한 연구는 1818년 새뮤얼 L. 미칠에 의해 시작되었으며, 제임스 엘스워스 드 케이, 루이 아가시, 존 메이슨 클라크 등에 의해 다양성과 생물학이 밝혀졌다.
더 읽어볼만한 페이지
- 협각아문 - 협각류
협각류는 캄브리아기부터 존재해 온 절지동물 아문으로, 머리가슴과 배로 나뉘는 몸, 협각, 다리 형태의 부속지를 가지며, 거미, 전갈, 진드기 등을 포함한다. - 광익목 - 메가라크네
메가라크네는 아르헨티나에서 발견된 멸종된 유절지동물로, 처음에는 거대한 거미로 오인되었으나, 2005년 바다 전갈로 재분류되었다. - 광익목 - 믹소프테루스
믹소프테루스는 중생대 실루리아기에 살았던 최대 75cm 크기의 육식성 바다전갈로, 단단한 외골격과 헤엄다리를 가지며 매복 사냥을 했을 것으로 추정된다. - 1843년 기재된 화석 분류군 - 넓은이빨청상아리
넓은이빨청상아리는 마이오세에서 플라이오세 시기에 전 세계 바다에 서식했던 멸종된 악상어목 상어로, 현생 청상아리와 유사하나 톱니가 있는 이빨을 가졌으며 고래, 해양 포유류 등을 먹이로 삼았을 것으로 추정되고 넓은이빨 형태는 백상아리의 조상으로 여겨지지만 속의 분류는 불확실하다. - 1843년 기재된 화석 분류군 - 오토두스 앵거스티덴스
오토두스 앵거스티덴스는 올리고세와 마이오세에 살았던 멸종된 상어 종으로, 백상아지와 비슷하지만 더 크고 톱니 모양의 이빨을 가진 강력한 포식자였으며, 메갈로돈과 관련이 있고 전 세계에서 화석이 발견되지만 한반도에서는 현재 서식하기 어려울 것으로 예상된다.
| 바다전갈 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Eurypterida |
| 명명자 | 부르마이스터, 1843 |
| 이명 | 기간토스트라카 (Gigantostraca) 헤켈, 1866 키르토크테니다 (Cyrtoctenida) 스퇴르머 & 워터스턴, 1968 |
| 한국어 이름 | 우미사소리목 (바다전갈목), 광익목 |
| 영어 이름 | Sea scorpion (바다전갈), Eurypterid (광익류) |
| 어원 | 그리스어 εὐρύς (eurys, 넓은) 그리스어 πτερόν (pteron, 날개) |
| 크기 | 최대 2.5 m |
| 발톱 크기 | 최대 20 cm |
| 꼬리 가시 크기 | 최대 2.03 cm |
| 멸종 시기 | 페름기 말기 |
| 생존 시기 | 오르도비스기다리윌절 - 페름기창싱절 (4억 6730만 년 전 - 2억 5200만 년 전) |
| 생존 시기 참고 | 가능한 장싱기 기록 |
| 하위 분류 | |
| 아목 | †유립테리나 (Eurypterina) 부르마이스터, 1843 †스틸로누리나 (Stylonurina) 디에너, 1924 |
| 불확실한 위치 | †도르포프테루스 (Dorfopterus) †마르수피프테루스 (Marsupipterus) |
2. 명칭
학명 "'''Eurypterida'''"는 본 군의 한 속인 유리프테루스의 학명 "Eurypterus" (고대 그리스어의 "ευρυςgrc"(eurys, 넓은[5])와 "πτερονgrc"(pteron, 날개)의 합성어[10][11])에, 일반적으로 목 계급의 분류군 학명에 사용되는 어미 중 하나인 "-ida"를 붙인 것이다[12].이명](동물 분류학에서의 다른 이름)으로는 "큰 전갈목[13] Gigantostraca"와 "Cyrtoctenida"가 있다[68]。
바다전갈은 협각류처럼 몸을 크게 두흉부(머리 부분, prosoma)와 후체구(복부, opisthosoma)로 나눌 수 있다. 두흉부에는 여섯 개의 마디가 있고, 후체구에는 열두 개의 마디와 꼬리마디(telson)가 있다. 꼬리마디는 기원이 달라 마디로 치지 않는다.[78]
영어에서는 통칭인 "'''sea scorpion'''" 외에, 학명 "Eurypterida"에서 유래한 학술적 총칭 "'''eurypterid'''"가 있다[11]。중국어에서는 번역어로 "鱟중국어" (투구게류)의 글자를 더해 "廣翅鱟중국어" (간체자: )라고 부른다. 이 외에 "板足鱟중국어" (간체자: )와 "海蝎중국어"이라는 이름도 있다. 일본어에서는 영어의 "sea scorpion"과 "eurypterid"에 각각 대응하는 "'''우미사소리'''('''海蠍일본어''')"와 "'''코요쿠루이'''(広翼類)"가 있다[5][6]。
3. 형태
두흉부에는 마디마다 부속지(몸에 붙어있는 기관)가 한 쌍씩 있다. 첫째 마디의 부속지는 협각으로, 먹이를 입에 넣는 역할을 한다. 나머지 다섯 쌍은 다리로, 걷거나 먹이를 잡는 데 쓰인다. 다리 안쪽에는 밑마디(coxa)가 있고, 여기에는 악기(gnathobase)라는 톱니 모양 가시가 있어 먹이를 잘게 부순다. 두흉부 등쪽은 갑각(carapace)으로 덮여 있고, 여기에는 큰 겹눈 한 쌍과 작은 홑눈 한 쌍이 있다. 배쪽에는 메타스토마(metastoma)라는 판이 있어 배면 가운데를 감싼다.[78]
후체구는 중체부(mesosoma)와 후체부(metasoma), 또는 전복부(preabdomen)와 후복부(postabdomen)로 나눌 수 있다. 등쪽은 등판(tergite), 배쪽은 배판(sternite)으로 덮여 있다. 앞쪽 배면에는 생식기(genital appendages)가 있는데, 이는 첫째와 둘째 마디의 부속지가 합쳐진 것이다. 셋째에서 여섯째 마디까지는 부속지가 변형된 블라트퓌세(Blattfüsse)가 배면을 덮고, 이는 호흡기관을 덮는 덮개 역할을 한다.[78]
후체구 끝에는 꼬리마디가 있는데, 대부분 칼날 모양이다. 하지만 프테리고토이데아(Pterygotioidea), 히베르토프테루스과(Hibbertopteridae), 미크테로프티스과(Mycteroptidae)는 꼬리마디가 납작하여 수영할 때 방향타 역할을 했을 수 있다. 카르키노소마토이데아(Carcinosomatoidea) 상과의 일부 속, 특히 ''에우사르카나''(Eusarcana)는 전갈과 유사한 꼬리마디를 가졌고, 독을 주입하는 데 사용했을 수도 있다.[14]
후체구 밑면은 변형된 부속지가 덮고 있는데, 이는 Blattfüssede(독일어로 잎다리)라는 판 모양 구조를 이룬다.[1] 이 구조는 호흡 기관이 있는 Blattfüsse|italics=unsetde와 복부 표면 사이에 아가미실(아가미 도관)을 만든다. 둘째에서 여섯째 후체구 분절에는 타원형 또는 삼각형 기관(Kiemenplattende, 아가미 도관)이 있어 호흡을 돕는다. 이 기관은 바다 위에서 공기 호흡을 돕고, Blattfüssen|italics=unsetde은 말굽게처럼 수중 호흡 부분을 덮었을 것이다.[1]
바다전갈은 크기가 매우 다양했다. 대부분 100cm 정도였지만,[15][16] 가장 작은 종인 ''알켄롭테루스 버글라렌시스''는 2.03cm에 불과했다.[16] 반면 가장 큰 종인 ''자켈롭테루스 레나니아에''는 2.5m에 달해, 지금까지 살았던 가장 큰 절지동물이었다.[9]
3. 1. 전체 (Prosoma)
바다전갈은 다른 협각류와 마찬가지로 몸을 크게 두 부분으로 나눌 수 있는데, 앞쪽을 두흉부(prosoma, 전체), 뒤쪽을 후체구(opisthosoma)라고 한다.
두흉부에는 총 여섯 개의 마디가 있다. 마디마다 부속지가 한 쌍씩 달려 있는데, 첫째 마디의 부속지는 입으로 먹이를 집어넣는 협각이다. 나머지 다섯 쌍의 부속지는 걷거나 먹이를 잡는 데 사용되는 다리이다. 다리의 가장 안쪽 마디는 밑마디(coxa)라고 하며, 여기에는 톱니 모양의 가시인 악기(gnathobase)가 있어 먹이를 잘게 부수는 역할을 한다. 두흉부 등면은 여섯 마디를 덮는 하나의 갑각(carapace)으로 덮여있다. 갑각 위에는 한 쌍의 큰 겹눈(compound eye)과 한 쌍의 작은 홑눈(ocellus)이 중앙에 있다. 배면에는 메타스토마(metastoma)라는 판이 다섯째 다리쌍의 밑마디 사이에 있어 배면 가운데를 감싼다.[78]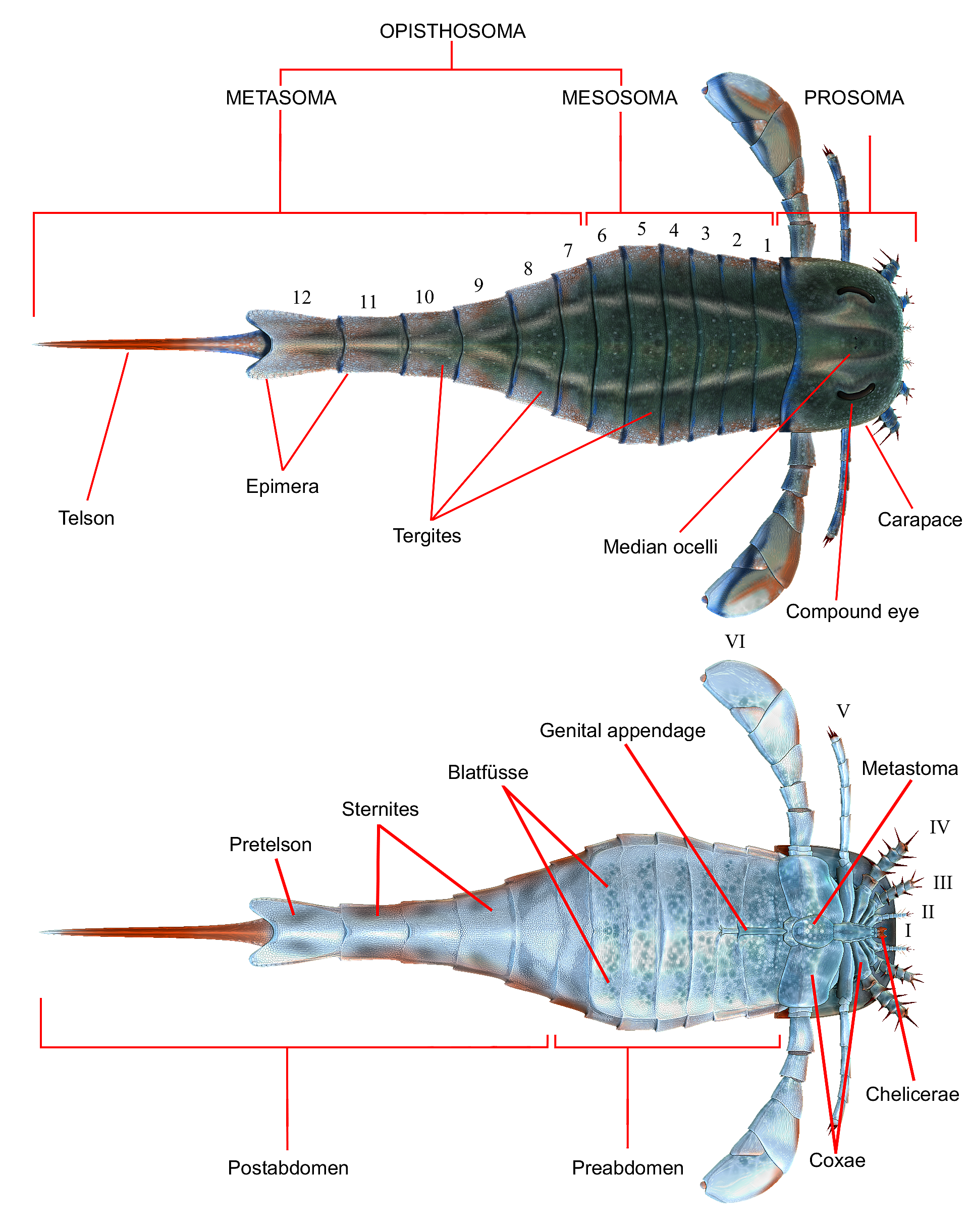
다른 모든 절지동물처럼 바다전갈은 분절된 몸체와 단백질과 키틴으로 구성된 큐티클로 덮인 관절 부속지(다리)를 가졌다. 전면은 딱지로 덮여 있었으며, 그 위에 겹눈과 홑눈이 모두 위치해 있었다.
전면에는 부속지 쌍 I에서 VI라고 하는 6쌍의 부속지가 있었다. 부속지의 첫 번째 쌍은 입 앞에 위치한 유일한 쌍으로, 협각이라고 한다. 이들은 음식 조각을 조작하고 입으로 밀어 넣는 데 사용되는 작은 집게발을 갖추고 있었다. 한 계통인 프테리고투스과(Pterygotidae)에서는 협각이 크고 길었으며, 특화된 집게 (발톱)에 강하고 잘 발달된 이빨이 있었다. 그 다음 부속지 쌍인 II에서 VI까지는 먹이를 먹는 데 사용되는 coxae (사지 분절)에 gnathobases (또는 "치판")를 가지고 있었다. 이 부속지는 일반적으로 원통형 모양의 걷는 다리였으며 일부 종에서는 가시로 덮여 있었다. 대부분의 계통에서 사지는 뒤로 갈수록 커지는 경향이 있었다. 두 개의 바다전갈 아목 중 더 큰 유리프테루스아목(Eurypterina)에서 여섯 번째 부속지 쌍은 수생 환경을 가로지르는 데 도움이 되도록 수영 패들로 변형되었다.
전체 (prosoma, 머리가슴 cephalothorax라고도 하며, 머리 자체에 해당한다[59])는 눈이나 입 등이 유래하는 전절과 6쌍의 전체 부속지 (후술)가 유래하는 직후 6절의 체절 (제1-6체절)의 융합으로 생긴 합체절로 생각된다[14]. 1장의 등갑 (carapace, prosomal dorsal shield[14])으로 덮여 있으며, 그 등쪽 좌우에 겹눈인 측안 (lateral eye)[17]과 중앙에 홑눈인 중안 (median eye)을 각각 한 쌍씩 갖는다. 등갑의 가장자리는 돌출되어 있지만, 투구게류만큼 극단적이지 않으며, 그 아래에 있는 모든 부속지 전체를 덮을 정도는 아니다. 등갑의 복면은 가장자리를 따라 접혀진 외골격 구조 (doublure[18] 또는 ventral plate[19])가 있으며, 분류군에 따라 1장부터 5장 정도로 세분된다[18][20][31][24]. 투구게류와 비슷하게, 이 외골격과 등갑의 경계, 즉 등갑의 가장자리는 탈피의 틈새이기도 하다[19]. 부속지에 둘러싸인 복면 중앙에는 "endostoma"라는 눈에 띄지 않는 두 잎 모양의 외골격이 있으며[20], 일반적으로는 투구게류와 같은 전체의 복판 (sternite, sternum) 유래로 여겨지지만[14], 바다전갈의 경우에는 문헌에 따라 제5다리 기절에서 유리된 턱기[25]·제7체절의 부속지라는 이설도 있다[21]. 이 외골격 바로 앞에는, 입과 그것을 덮어 씌운 눈에 띄지 않는 위입술 (labrum)과 구상판 (epistome)의 복합체가 있다[25][22].
다른 진협각류와 마찬가지로, 전체의 복면은 1쌍의 협각과 5쌍의 보행다리형 부속지(다리)라는 총 6쌍의 부속지(관절지)를 가진다.[14] 협각은 가위 모양으로 3절의 지절(자루부 1절과 가위 2절)로 이루어지며[23][14], 기부는 구상판과 입술의 복합체에 관절한다.[25][22] 일반적으로는 완전히 전신에 숨겨질 정도로 짧고 눈에 띄지 않지만, 밀러바다전갈과[6]는 배갑의 앞쪽 가장자리를 약간 넘을 정도로 크고[20][30], 다이오우바다전갈과(Pterygotidae)는 더욱 극단적으로 발달하여 팔처럼 길게 뻗어 있었다.[20][24][23][18][30]
협각 바로 뒤에 이어진 5쌍의 다리는 뒤쪽일수록 많은 지절로 나뉘며 (원칙적으로 제1다리 7절, 제2-3다리 8절, 제4-제5다리 9절)[25][7], 분류군이나 번호에 따라 단조로운 보행다리형에서 다소 특수한 형태로 특화된 것이 있다.[18][7][41] 각각의 다리의 기부 (기절 coxa)는 입을 둘러싼 저작기인 톱니 모양의 턱기부 (gnathobase)를 가진다.[26] 처음 1쌍의 다리는 촉지이며, 일반적으로는 형태적으로 다리와 분화되지 않지만, 분류군이나 암수[25]에 따라 어느 정도의 특화가 보이는 경우도 있다.[18] 마지막 1쌍의 다리는, 바다전갈아목에서는 제7지절 선단 안쪽에 "podomere 7A"라는 구조체가 있다.[27] 그 중 모셀롭테루스과를 제외한 종류는 현저하게 특화된 유영다리(swimming leg)이며, 제7지절, 제8지절, podomere 7A가 편평하고, 노 모양의 패들 (paddle, swimming paddle)을 구성한다.[18][27] 긴다리바다전갈아목의 경우, 이 다리는 podomere 7A를 가지지 않고, 보행다리형 그대로이다.[18][27][28]
3. 2. 후체 (Opisthosoma)
바다전갈은 다른 협각류와 마찬가지로 몸이 크게 두 부분, 즉 앞쪽의 두흉부(prosoma)와 뒤쪽의 후체구(opisthosoma)로 나뉜다. 후체구는 12개의 마디와 꼬리마디(telson)로 구성된다. 꼬리마디는 다른 마디와 기원이 달라 마디의 일부로 보지 않는다.[78]
후체구는 중체부(mesosoma)와 후체부(metasoma)로, 또는 전복부(preabdomen)와 후복부(postabdomen)로 나눌 수 있다. 등면은 등판(tergite), 배면은 배판(sternite)으로 덮여 있다. 앞쪽 배면에는 후체구의 첫째와 둘째 마디에 달린 부속지가 합쳐져 생긴 생식기(생식 부속지, genital appendages)가 있다. 셋째에서 여섯째 마디까지는 부속지가 판 모양으로 변형된 블라트퓌세(Blattfüsse)가 배면을 덮는다. 블라트퓌세는 광익류의 호흡기관을 덮는 일종의 덮개 역할을 한다.[78]
후체는 12개의 분절과 미절(telson)로 구성된다.[59] 미절은 대부분 칼날 모양이지만, 프테리고토이데아(Pterygotioidea), 히베르토프테루스과(Hibbertopteridae), 미크테로프티스과(Mycteroptidae) 등 일부 계통에서는 납작해져 수영 시 방향타 역할을 했을 수 있다. 카르키노소마토이데아(Carcinosomatoidea) 상과의 일부 속, 특히 ''에우사르카나''(Eusarcana)는 현대 전갈과 유사한 미절을 가지고 있어 독을 주입하는 데 사용했을 수도 있다.[14]
후체 밑면은 변형된 후체 부속지에서 진화한 구조로 덮여 있다. 후체 전체에 걸쳐 이 구조는 Blattfüssede(독일어로 잎다리)라는 판 모양 구조를 형성했다.[1] 이것은 호흡 기관이 포함된 앞의 Blattfüsse|italics=unsetde와 후체 자체의 복부 표면 사이에 아가미실(아가미 도관)을 만들었다. 두 번째에서 여섯 번째 후체 분절에는 호흡에 도움이 되는 기관으로 해석된 타원형 또는 삼각형 기관도 포함되어 있었다. Kiemenplattende(아가미판) 또는 "아가미 도관"이라고 하는 이 기관은 바다 위에서 바다전갈이 공기를 호흡하는 데 잠재적으로 도움이 되었을 것이며, 현대 말굽게의 기관과 유사한 Blattfüssen|italics=unsetde은 수중 호흡을 담당하는 부분을 덮었을 것이다.[1]
바다전갈의 호흡 기관은 복부 벽에 위치했다. Blattfüsse|italics=unsetde는 opisthosomal 부속지에서 진화하여 아랫부분을 덮고 "아가미 구획"이 위치한 아가미 방을 만들었다.[1] 종에 따라 바다전갈 아가미 구획은 삼각형 또는 타원형 모양이었고 쿠션과 같은 상태로 솟아 있었을 가능성이 있다. 이 아가미 구획의 표면에는 여러 개의 가시가 있어 표면적을 넓혔다. 구조 내에 많은 내함입으로 인해 스펀지 조직으로 구성되어 있었다.
일부 연구자들은 "아가미 구획"이 실제 아가미가 아니라 공기 호흡 기관, 즉 폐, 플라스트론 또는 가짜 기관지였을 것이라고 추정한다. 바다전갈 아가미 구획은 현대 등각류에서 발견되는 가짜 기관지와 가장 유사하며, 이 두 기관이 같은 방식으로 기능했을 가능성이 제기되었다.
후체는 길게 뻗어 있으며, 등면의 등판은 12절로 보이지만 실제로는 13절(제7-19체절)의 합체절이며, 그중 후체 제1절(제7체절)의 등판은 관찰할 수 없을 정도로 퇴화되었다.[14] 일반적으로 4번째 등판이 가장 넓지만,[31] 과 같이 1-2번째 등판이 가장 넓은 분류군도 있다.[28] 복면에는 체절마다 복판이 늘어서 있지만, 최종 6절만 뚜렷하게 보이며, 전반부의 복판은 퇴화되었거나 완전히 개판에 덮여 있다.[31] 최종 5절은 등판과 복판이 융합되어 링 모양의 체환이 된다.[14] 후체의 말단에는 1개의 미절이 있으며, 일반적으로 칼 모양으로 뾰족하지만, 주걱 모양도 있다(스리모니아과, 다이오우미사소리과 등).[18][29][30][14]
일부 분류군에서는 후체가 전갈처럼 명확하게 전후로 폭이 넓은 전반부와 가늘고 긴 후반부로 분화되어 있으며, 각각 중체(mesosoma)와 종체(metasoma), 혹은 전복부(preabdomen)와 후복부(postabdomen)라고 불린다.[18][14]
3. 3. 호흡 기관
바다전갈은 다른 모든 절지동물과 마찬가지로 후체 밑면에 변형된 후체 부속지에서 진화한 구조로 덮여 있었다. 후체 전체에 걸쳐 이러한 구조는 Blattfüssede(독일어로 잎 모양 다리)라고 하는 판 모양 구조를 형성했다.[1] 이것은 호흡 기관이 포함된 앞의 Blattfüssen|italics=unsetde와 후체 자체의 복부 표면 사이에 아가미실 (아가미 도관)을 만들었다. 두 번째에서 여섯 번째 후체 분절에는 호흡에 도움이 되는 기관으로 해석된 타원형 또는 삼각형 기관도 포함되어 있었다. Kiemenplattende(아가미 도관)이라고 하는 이 기관은 바다 위에서 바다전갈이 공기를 호흡하는 데 잠재적으로 도움이 되었을 것이며, 현대 말굽게의 기관과 유사한 Blattfüssen|italics=unsetde은 수중 호흡을 담당하는 부분을 덮었을 것이다.[1]
4. 생태
바다전갈은 종류에 따라 다양한 생활 방식을 가졌다. 프테리고투스과(Pterygotidae)는 집게발처럼 크게 발달한 협각을 이용해 다른 동물을 사냥하는 포식자였을 것으로 보인다. 특히 야이켈롭테루스(Jaekelopterus)나 프테리고투스(Pterygotus)와 같은 대형종은 최상위 포식자였을 가능성이 크다. 반면, 아쿠티라무스(Acutiramus)는 큰 몸집에도 불구하고 시력이 좋지 않아 매복 포식자나 시체 청소부 역할을 했을 것으로 추정된다.[89]
카르키노소마과(Carcinosomatidae)는 지느러미 다리를 제외한 모든 다리에 강력한 가시가 발달하여 먹이를 제압하는 포식자였을 것이다.
스틸로누루스과(Stylonuridae)는 첫째와 둘째 다리에 빗처럼 촘촘한 가시가 있어, 퇴적물을 긁어 가시에 걸린 먹이를 먹는 방식에 적응했을 것으로 보인다. 이들은 밑바닥 생활에 적응하여 메타스토마(metasoma)가 납작한 판 모양으로 변형되었다.[82]
힙베르톱테루스과(Hibbertopteridae)는 스틸로누루스과와 유사한 생활 방식을 가졌지만, 일부 가시가 감각모로 변형되어 퇴적물 속 먹이를 감지하는 등 더 발전된 사냥 방식을 보였다. 셋째 다리까지 먹이를 찾는 데 사용했으며, 키르토크테누스 윗테베르겐시스(Cyrtoctenus wittebergensis)는 다리 가시가 두 가지 형태로 변형되어, 머리빗 형태의 가시에 걸린 먹이를 막대 형태의 가시로 떼어내는 역할을 했을 것으로 추정된다.[90]
바다전갈은 생활 방식, 서식 환경, 분류학적 유사성 등에 따라 크기가 매우 다양했다. 대부분의 바다전갈 그룹에서 100cm 정도의 크기가 흔했다. 가장 작은 바다전갈은 알켄롭테루스(Alkenopterus)로, 몸길이가 2.03cm에 불과했다.
가장 큰 바다전갈이자 현재까지 알려진 가장 큰 절지동물은 야이켈롭테루스 레나니아에(Jaekelopterus rhenaniae)이다. 독일에서 발견된 집게 화석을 통해 추정한 몸길이는 평균 2.5m에 달하며, 집게발을 펼치면 1미터가 더해진다. 이는 다른 거대 절지동물의 최대 몸 크기를 훨씬 뛰어넘는 수치이다. 에레토프테루스(Eretopterus)와 히베르토프테루스(Hibbertopterus)도 2.5미터에 달했을 것으로 추정되지만, 전자는 단편적인 화석만이, 후자는 발자국 증거를 기반으로 추정되었다.
야이켈롭테루스(Jaekelopterus)가 속한 프테리고티드(Pterygotidae)과는 여러 대형 종으로 유명하다. 2.1m 크기의 아쿠티라무스(Acutiramus)와 1.75m 크기의 프테리고투스(Pterygotus)는 거대했다. 이러한 큰 크기는 구애 행동, 포식, 환경 자원 경쟁 등의 요인에 영향을 받았을 것으로 추정된다.
거대한 바다전갈은 프테리고티드과에만 국한되지 않았다. 카르시노소마(Carcinosoma)는 2.2m에 달해 프테리고티드과와 경쟁했으며, 펜테콥테루스(Pterygotus)는 1.7m에 달했을 것으로 추정된다.
거대한 바다전갈은 가벼운 몸체를 가진 것이 특징이다. 가벼운 구조는 탈피, 호흡, 에너지 비용, 외골격의 물리적 특성 등 절지동물의 크기를 제한하는 요인의 영향을 줄여준다. 프테리고티드는 특히 가벼웠으며, 대부분의 큰 몸체 화석은 얇고 비광물화된 상태로 보존되었다. 이러한 가벼운 적응은 다른 거대 고생대 절지동물에게도 나타나며, 거대 절지동물 진화에 필수적이었을 수 있다.
가벼운 거대 바다전갈 외에도, 히베르토프테리드과의 일부는 매우 큰 몸집을 가졌다. 히베르토프테루스 스코울레리(Hibbertopterus scouleri)의 갑각 폭은 65cm이며, 몸길이는 거의 2m에 달했을 것으로 추정된다. 프테리고티드보다 더 튼튼한 이 거대한 히베르토프테루스(Hibbertopterus)는 무게가 가장 큰 프테리고티드와 경쟁하거나 능가했을 가능성이 있다.
두 바다전갈아목(Eurypterina)은 주로 마지막 부속지(다리) 형태에 따라 구분된다. 스틸로누리나(Stylonurina)는 마지막 부속지가 길고 가느다란 보행 다리 형태인 반면, 바다전갈아목(Eurypterina)은 다리가 수정되어 수영 노로 넓어졌다. 많은 바다전갈아목(Eurypterina)의 다리는 해저를 기어가는 것 이상을 하기에는 너무 작았다. 반대로, 여러 스틸로누리나(Stylonurina)는 육상에서 걸을 수 있는 길고 강력한 다리를 가지고 있어 현대 게와 유사했을 것이다.
2005년 스코틀랜드에서 발견된 화석 궤적은 스틸로누리나(Stylonurina) 바다전갈 히베르토프테루스(Hibbertopterus)의 것으로 추정된다. 이는 절지동물이 만든 가장 큰 육상 궤적으로, 길이가 6m이고 너비는 평균 95cm이다. 이는 바다전갈의 육상 이동에 대한 첫 번째 기록이며, 일부 바다전갈이 적어도 짧은 기간 동안 육상 환경에서 생존할 수 있었음을 보여준다. 히베르토프테루스(Hibbertopterus)는 짧은 보폭으로 매우 느리게 기어다녔으며, 큰 텔손(telson)은 땅에 끌려 중앙 홈을 남겼다. 파라스틸로누루스(Parastylonurus)와 같은 작은 스틸로누리나(Stylonurina)는 더 빠르고 정확하게 걸었을 것이다.
바다전갈아목(Eurypterina) 수영 노의 기능은 그룹마다 달랐다. 유리프테로이데아(Eurypteroidea)에서 노는 노와 유사하게 위아래가 아닌 거의 수평면에서만 움직일 수 있었다. 프테리고티오이데아(Pterygotioidea)는 더 빨리 수영할 수 있었을 것이다. 대부분의 바다전갈아목(Eurypterina)은 게와 물벌레와 유사한 노 젓기 유형의 추진력을 사용했지만, 더 큰 개체는 바다거북과 바다사자처럼 수중 비행이 가능했을 수 있다. 이는 노의 움직임과 모양만으로 양력을 생성하는 방식으로, 더 에너지 효율적이었을 것이다.
믹소프테루스(Mixopterus)는 수영을 잘하는 것은 아니었으며, 주로 바닥에 머물면서 수영 노를 사용해 가끔 수직으로 움직였다. 걷는 동안에는 긴 복부의 무게를 두 개의 무겁고 특수한 전면 부속지가 균형을 이루었고, 중심은 꼬리를 올려 조절했다.
보존된 화석화된 바다전갈 궤적은 크고 이종각 부위를 가지는 경향이 있으며, 중앙선을 따라 텔손 끌기 자국이 있는 경우가 많다. 이러한 궤적은 남아메리카를 제외한 모든 대륙에서 발견되었다. 남아프리카 공화국과 곤드와나와 같이 바다전갈 화석 유해가 드문 곳에서는 궤적의 발견이 바다전갈 몸 화석보다 앞서고 더 많다. 바다전갈 궤적은 여러 종 식별에 언급되었으며, 가장 유명한 것은 팜미크니움(Palmichnium)이다. 팜미크니움 코신키오룸(P. kosinskiorum)은 지금까지 알려진 가장 큰 바다전갈 발자국을 보존하며, 각 궤적은 직경이 약 7.6cm이다.
Kiemenplatte|italics=unsetde는 "아가미 구획"이라고 불리지만 실제 아가미로 기능하지 않았을 수도 있다. 다른 동물에서 아가미는 물에서 산소를 흡수하는 데 사용되며 몸벽에서 밖으로 자란다. 바다전갈은 수생 동물이었지만, "아가미 구획"에는 기능적인 아가미가 포함되지 않았을 가능성이 높다. 이전의 해석은 종종 바다전갈의 "아가미"를 다른 그룹의 아가미와 상동 기관으로 식별했지만, 투구게의 아가미 구조와 바다전갈의 아가미 구조는 매우 다르다. 투구게에서 아가미는 가스 교환에 사용되는 더 큰 표면적을 제공하는 많은 라멜라(판)로 구성되어 있다. 또한, 바다전갈의 아가미 구획은 다른 그룹의 아가미와 유사하다면 그것들을 지탱하기에는 비례적으로 너무 작다. 기능적인 아가미가 되려면 효율성이 높아야 했으며 매우 효율적인 순환계가 필요했을 것이다.
대신, "아가미 구획"은 공기를 호흡하는 기관, 아마도 폐, 플라스트론 또는 가짜 기관지였을 것이라고 제안되었다. 플라스트론은 일부 절지동물이 수중에서 공기를 호흡하기 위해 이차적으로 진화한 기관이지만, 바다전갈은 처음부터 물에서 진화했으므로 설명하기 어렵다. 대신 절지동물 호흡 기관 중에서 바다전갈 아가미 구획은 현대 등각류에서 발견되는 가짜 기관지와 가장 유사하다. 이러한 기관은 공기 호흡 생물의 기관과 약간의 유사성이 있기 때문에 가짜 기관지라고 불리며 등각류의 뒷다리 내에 존재한다. 가짜 기관지의 구조는 바다전갈 아가미 구획의 스펀지 구조와 비교되었다. 두 기관이 같은 방식으로 기능했을 수 있다.
일부 연구자들은 바다전갈이 양서류 생활 방식에 적응하여 전체 아가미 구획 구조를 아가미로 사용하고 그 안의 내함입을 가짜 기관지로 사용했을 수 있다고 제안했다. 그러나 이 생활 방식은 생리학적으로 가능하지 않았을 수 있는데, 물의 압력이 내함입으로 물을 밀어 넣어 질식을 유발했을 것이기 때문이다. 또한, 대부분의 바다전갈은 평생 동안 수생 동물이었을 것이다. 육지에서 얼마나 많은 시간을 보냈든지, 수중 환경에서의 호흡 기관이 존재했어야 한다. Blattfüssen|italics=unsetde 내의 아가미 방 내에 위치했을 것으로 예상되는 실제 아가미는 바다전갈에서는 알려져 있지 않다.
''Carcinosoma'', ''Acutiramus'' 및 ''유리프테루스''(Eurypterus)를 포함한 다양한 유르프테리드의 화석에서 소화관이 보존된 화석이 보고되었다. ''버팔로프테루스''(Buffalopterus) 표본의 텔슨(telson)에서 잠재적인 항문이 보고되었지만, 현대 말굽게와 마찬가지로 항문은 텔슨 앞 마지막 마디와 텔슨 사이의 얇은 큐티클을 통해 열렸을 가능성이 더 높다.
오하이오주에서 발견된 오르도비스기 퇴적층에서 유르프테리드 배설물(coprolite)은 삼엽충 조각과 유르프테리드 ''메갈로그랍투스 오하이오엔시스''(Megalograptus ohioensis) 조각을 포함하고 있으며, 동일한 유르프테리드 종의 완전한 표본과 함께 발견되어 카니발리즘의 증거를 나타내는 것으로 추정된다. 오하이오주 오르도비스기에서 발견된 ''라나르코프테루스 돌리코스켈루스''(Lanarkopterus dolichoschelus) 종에 속하는 유사한 배설물은 무악어류 조각과 ''라나르코프테루스''(Lanarkopterus) 자체의 더 작은 표본 조각을 포함하고 있다.
최상위 포식자 역할은 매우 큰 유르프테리드에게만 국한되었겠지만, 더 작은 유르프테리드도 더 큰 친척과 마찬가지로 그 자체로 강력한 포식자였을 것이다.
하층판 이후의 후체 제2-7절은 6쌍의 덮개판(개판, operculum)이라는 평판 모양의 부속지가 있으며, 체절의 복면(복판)을 덮어 덮는 것처럼 뒤쪽으로 접혀있다.[14] 체절에 밀착되어 투구게류의 덮개판 정도의 운동 능력은 없었던 것으로 생각된다.[45] 처음 2쌍의 덮개판은 전후 융합된 생식 구개(genital operculum), 나머지 4쌍의 독립적인 덮개판은 Blatfüsse|italics=unsetde(단수: Blatfuss)라고 불린다.[14] Blatfüsse|italics=unsetde는 통상적으로 모두 좌우 융합되어 있지만, 중앙의 홈(central sulture)으로 좌우로 나뉘는 경우도 있다.[52]
처음 1쌍(후체 제2절)의 덮개판만으로 이루어진 다른 협각류의 생식 구개와는 달리, 바다 전갈의 생식 구개는 전후 융합된 처음 2쌍(후체 제2-3절)의 덮개판으로 구성되어 있으며, 이 특징은 바다 전갈을 명확히 다른 유사군(특히 카스마타스피스류)에서 구분시키는 가장 중요한 공유 파생 형질이다.[31][73][59][14] 그 중 후체 제2절의 덮개판에 해당하는 판은 "median opercular plate", 후체 제3절의 덮개판에 해당하는 판은 "posterior opercular plate"라고 불린다.[31] 긴발바다전갈아목과 기반적인 바다전갈아목 종류에서는 생식 구개는 더 나아가 "anterior opercular plate"라는 단축된 판이 전단에 있으며, 이는 후체 제1절의 복판 유래의 부분으로 생각된다.[31] 파생적인 바다전갈아목(Diploperculata) 종류에서는 anterior opercular plate를 갖지 않고,[73] 나머지 판은 더욱 융합이 진행되어 경계에 해당하는 홈(transverse suture)이 보이지 않을 정도로 일체화되었다.[31] 또한, posterior opercular plate의 중앙에는 "spatulae"라는 생식 부속지의 좌우에 늘어선 1쌍의 돌기물을 갖는 경우가 있다.[18][32]
생식 구개의 중앙에는 1개의 생식 부속지 (genital appendage)를 갖는다.[14] 이것은 후체 제2절의 덮개판에 유래하며,[31] 그 1쌍의 내지가 좌우 융합하여 생긴 생식기로 생각된다.[32] 기부는 "deltoid plate"라는 1쌍의 삼각형/오각형 외골격에 연결되어[18][31] 이는 생식 부속지의 운동에 관여하는 구조로 생각된다.[34] 생식 부속지는 동종에서 긴 타입(type A)과 짧은 타입(type B)이라는 분명한 이형이 보이며, 이는 암수를 나타내는 특징으로 생각되지만,[33][34] 각각에 대응하는 성별(어느 쪽이 수컷이고 어느 쪽이 암컷인가)은 여러 설로 나뉘어져 있다.[52] 특히 type A의 경우, 생식 부속지의 뿌리에 "horn organ"이라는 1쌍의 긴 낭상 내부 구조가 있으며, 이는 문헌 기재에 의해 (암컷의) 수란관[51] 혹은 (수컷의) 정포를 생성하는 기관[52]으로 생각된다. 이 생식 부속지, 특히 type A는 통상적으로 3절로 나뉘지만, 대왕바다전갈과의 경우에는 암수 모두 생식 부속지가 분절되지 않는다.[30][31]
바다전갈은 일반적으로 포식자로 여겨지며, 분류군에 따라 다양한 포식 방법에 특화된 양상을 보인다.[41] 유영성이며 사나운 포식에 적합한 종류도 있는가 하면, 저생성이며 퇴적물에서 작은 먹이를 걸러내는 데 적합한 종류도 있다.[42][43][17][41] 그들의 포식 행동으로 남겨진 것으로 보이는 생흔화석도 알려져 있다.[44] 바다전갈아목과 긴다리바다전갈아목의 종류는, 각각 앞서 언급한 두 생태적 지위에 특화된 경향이 있다.[16]
바다전갈아목의 유영각을 가진 종류는 그 유영각으로 물 속에서 헤엄쳤다는 것이 널리 인정되지만, 유영의 메커니즘은 여러 설로 나뉘며,[45] 문헌에 따라 유영각의 앞뒤 동작과 노의 각도 변환으로 추진력을 생성[25][46], 혹은 유영각을 날개처럼 움직여 양력으로 전진했다고 생각된다.[47][45] 중형 이하로 큰 유영각을 가진 종류(예: 바다전갈과)는 추진력, 대형으로 작은 유영각을 가진 종류(예: 대왕바다전갈과)는 양력이라는, 종류에 따라 유영 방법이 달랐다는 설도 있다.[45]
이름과는 달리, 바다전갈류는 반드시 바다에만 국한되지 않고, 담수에 서식하는 종류도 많이 알려져 있으며, 초기 바다에서 점점 얕은 바다나 담수로 진출한 것이 화석 기록에 의해 시사된다.[48][16][17] 적어도 일부 종류는 육상으로 어느 정도 진출했을 가능성이 있으며, 이를 시사하는 발자국의 생흔화석[49]도 발견되고 있다. 또한, 그 호흡기로부터 공기 호흡에 적합한 구조도 발견된다(아가미판의 구조는 육생 갑각류의 가짜 기관과 유사[36], 서새의 라멜라 틈새에는 거미류의 서폐와 공통되는 소주가 있다[50]).
복잡한 생식기를 가지고 있어, 바다전갈은 말굽게류와 같은 체외수정을 하지 않고, 오히려 거미류와 같이 체내수정을 하며, 배우자 행동으로 암수가 특화된 생식지를 사용하여 정포를 주고받는 (교접)을 했을 것으로 생각된다.[51][52]
5. 분류
바다전갈은 거미강과 멸종한 카스마타스피스강(Chasmataspidida)과 함께 데카트리아타(Dekatriata)라는 분류군을 이룬다. 데카트리아타의 기본적인 형태는 후체구에 마디가 13개인 모습이다[79]. 바다전갈은 후체구의 부속지 두 쌍이 생식기로 변형되었다는 점에서 다른 데카트리아타 동물들과 구별된다[79].
데카트리아타 내에서 바다전갈과 거미강은 정포를 생성하는 기관이 있다는 공통점을 근거로 스클레로포라타(Sclerophorata)라는 분류군으로 묶인다[79]. 카스마타스피스강에서는 아직 정포 생성 기관이 발견되지 않았는데, 후대의 발견을 통해 스클레로포라타가 데카트리아타와 의미가 같아져 쓸모없는 분류군이 될 가능성도 있다[79].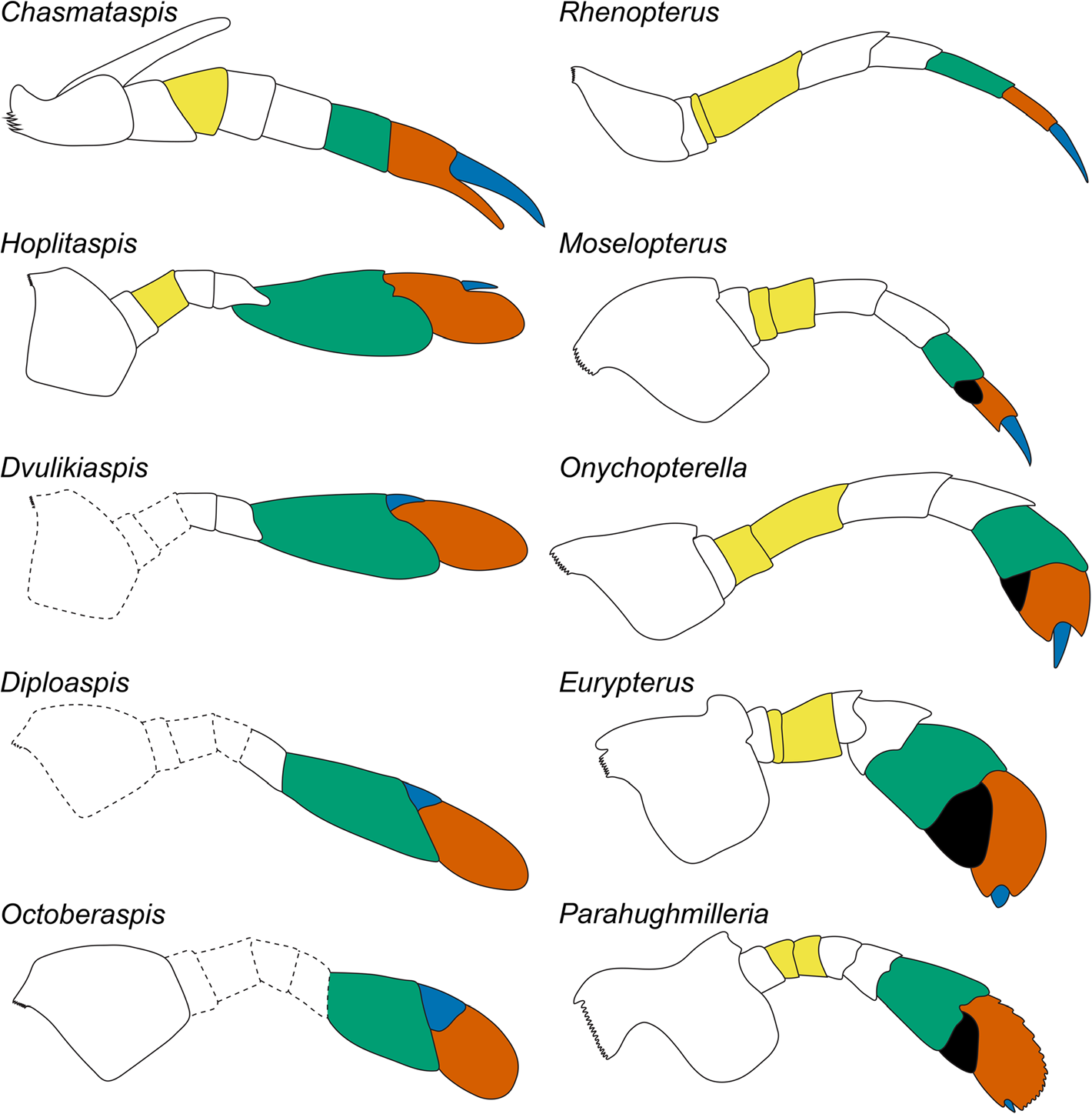
바다전갈은 광익아목(Eurypterina)과 스틸로누루스아목(Stylonurina)이라는 두 개의 아목으로 나뉜다. 광익아목은 다섯째 다리쌍의 일곱째 마디와 맨끝 마디 사이에 '사이마디'가 있는 반면, 스틸로누루스아목은 다섯째 다리쌍이 변형되지 않아 가느다란 모양이며 '사이마디'가 없다[82].
바다전갈의 생식과 성적 이형성은 화석으로만 연구가 가능하기 때문에 어려움이 있다. 바다전갈의 앞가슴은 여섯 개의 분절이 융합된 구조이며, 일곱 번째 분절은 '배판', 여덟 번째 분절은 '뚜껑'이라고 불리며 생식 개구부를 포함한다. 이 뚜껑 아랫면에는 말굽게와 유사한 생식 부속지가 있는데, "A형"과 "B형" 두 가지 형태가 존재한다.
바다전갈은 서새(book gill)와 "키에멘플라텐(Kiemenplatten)"이라는 두 종류의 호흡기를 5쌍 가지고 있으며, 모두 후체 제3-7절에 부속되어 있다[50]. 서새는 말발굽게와 비슷하지만, 미세 구조는 거미류의 서폐(book lung)에 가깝다[50]. 키에멘플라텐은 바다전갈에 특유한 호흡기로 여겨진다[36][14][50].
{{clade|style=font-size:100%;line-height:100%
|label1=절지동물|1={{clade
|label1=만다블류|1=다지류 --, 갑각류 
|label2=협각류|2={{clade
|1=해양거미류 --
|label2=진협각류|2={{clade
|1=말발굽게류 --
|label2=Dekatriata|2={{clade
|1=†카스마타스피스류 --
|label2=Sclerophorata|2={{clade
|1=†'''바다전갈류''' --
|label2=거미류|2=진드기--, 가짜전갈--, 전갈--, 거미-- 등
}} }} }} }} }}
과거에는 바다전갈류가 말발굽게류와 함께 구갑류(Merostomata)로 묶였으나[54][53][31][17], 20세기 이후 거미류 등과 함께 협각류(Chelicerata)로 묶이게 되었다[55].
최근에는 바다전갈류가 거미류에 더 가깝다는 (Sclerophorata[52] 혹은 Metastomata[56]) 설이 주류를 이루고 있다[56][57][58][52][59][14][17]. 이에 따르면 구갑류는 거미류를 제외한 측계통군이 된다[31][17].
{{cladogram
|title=
|align= right
|caption=진협각류 중에서 거미류가 다계통군이었을 경우 바다전갈류의 계통 위치[50]。
|cladogram=
{{clade|style=width:28em;font-size:100%;line-height:100%
|label1=진협각류|1={{clade
|1=다양한 거미류 (비폐포류) --
|2=다양한 거미류 (비폐포류) --
|3={{clade
|1=말발굽게류 --
|2={{clade
|1=†'''바다전갈류''' --
|2=다양한 거미류 (폐포류) ----
}} }} }} }} }}
카스마타스피스류와의 유연 관계도 논의되고 있으며, 2010년대 이후에는 독립된 단계통군이면서 바다전갈류, 거미류와 단계통군 (Dekatriata) 이 된다는 설이 주류가 되었다[59][61][62][63]。
5. 1. 광익아목 (Eurypterina)
바다전갈은 광익아목(Eurypterina)과 스틸로누루스아목(Stylonurina)이라는 두 개의 아목으로 나뉜다. 광익아목은 다섯째 다리쌍의 일곱째 마디와 맨끝 마디 사이에 일종의 '사이마디'가 세모꼴로 나 있는데[82], 이는 실제 마디가 아니고 다리에 난 돌기가 변화한 것이다. 모셀롭테루스(''Moselopterus'')와 같은 원시적인 종들은 이 다리가 가늘어서 걷는 데에만 쓰이지만, 그 외의 후대에 분화된 종들은 다섯째 쌍의 다리가 납작한 지느러미처럼 변화하여 수영을 할 수 있었다.
다른 많은 멸종된 그룹과 마찬가지로, 바다전갈의 생식과 성적 이형성은 화석화된 껍질과 등판으로만 알려져 있기 때문에 이해하고 연구하기 어렵다.
바다전갈의 앞가슴은 첫 번째 여섯 개의 외골격 분절이 융합되어 더 큰 구조를 이룬다. 일곱 번째 분절(첫 번째 뒤가슴 분절)은 '배판', 여덟 번째 분절(뚜렷한 판 모양)은 '뚜껑'이라고 하며 생식 개구부를 포함한다. 이 분절의 아랫면은 생식 뚜껑이 차지하고 있는데, 이는 원래 조상의 일곱 번째와 여덟 번째 부속지 쌍에서 진화한 구조이다. 중앙에는 현대 말굽게와 마찬가지로 생식 부속지가 있다. 이 부속지는 내부 관이 있는 길쭉한 막대로, "A형"과 "B형"의 두 가지 뚜렷한 형태가 있다.
A형 부속지는 일반적으로 B형보다 길다. 일부 속에서는 A형 부속지가 세 부분으로 나뉘는 반면, B형 부속지는 두 부분으로만 나뉘는 'Eurypterus'와 같이 다른 수의 부분으로 나뉜다. A형 부속지에는 furcala(라틴어로 '포크')라고 불리는 두 개의 곡선형 가시가 있다. B형 부속지에도 furca|italics=unsetla가 존재할 수 있으며, 이는 부속지의 융합되지 않은 끝을 나타낼 수 있다. A형 부속지와 관련된 Blattfüsse|italics=unsetde의 등쪽 및 복부 표면 사이에는 "관형 기관" 또는 "뿔 기관"으로 묘사되는 일련의 기관이 있다. 이 기관은 대부분 정자낭(정자를 저장하는 기관)으로 해석되지만, 이 기능은 아직 확실하게 입증되지 않았다. 절지동물에서 정자낭은 수컷에게서 받은 정자보를 저장하는 데 사용된다. 이는 A형 부속지가 암컷 형태이고 B형 부속지가 수컷 형태임을 의미한다.
길고 암컷으로 추정되는 A형 부속지의 주요 기능은 산란관이 절지동물 산란관보다 일반적으로 길기 때문에 산란관 역할보다는 기질에서 정자보를 생식관으로 가져가는 것이었을 것이다. 뚜껑의 측면을 회전시켜 부속지를 몸에서 내릴 수 있었을 것이다. 서로 다른 판이 해당 위치에 겹쳐져 있기 때문에 뚜껑 주변을 움직이는 근육 수축 없이는 부속지를 움직일 수 없었을 것이다. 사용하지 않을 때는 제자리에 유지되었을 것이다. A형 부속지의 furca|italics=unsetla는 정자보를 열어 내부의 자유 정자를 흡수하는 데 도움이 되었을 수 있다. "뿔 기관"(아마도 정자낭)은 관을 통해 부속지에 직접 연결된 것으로 생각되지만, 이러한 추정 관은 이용 가능한 화석 자료에서는 보존되지 않는다.
수컷으로 추정되는 B형 부속지는 부속지의 등쪽 표면에 있는 하트 모양의 구조에서 정자보를 생성, 저장 및 형성했을 것이다. 넓은 생식 개구부는 많은 양의 정자보를 한 번에 방출할 수 있게 했을 것이다. A형 부속지와 마찬가지로 내릴 수 있는 B형 부속지와 관련된 긴 furca|italics=unsetla는 기질이 정자보 침착에 적합한지 감지하는 데 사용되었을 수 있다.
2020년 현재 약 250종의 바다전갈류가 기재되어 있으며, 이 중 대부분은 바다전갈아목에 속한다[48][16][68]。
바다전갈아목 중에서 모세로프테루스상과(제5다리는 보행형)와 오니코프테렐라상과(제5다리는 보행형과 유영형의 중간 형태)는 본 아목의 유영형 다리의 진화를 보여주는 것처럼 차례로 분기되며[27], 바다전갈상과는 나머지 상과에 이르는 측계통군으로 여겨진다[73]。나머지 상과(믹소프테루스상과, 상과, 아델로프탈무스상과, 프테리고투스상과)는 파생적인 바다전갈류이며[31], 생식구개는 전단의 판을 가지지 않고 나머지 2개의 판은 경계가 보이지 않을 정도로 융합되었다[31][73]。이 특징에 따라, 이러한 상과는 ("두 개"를 의미하는 고대 그리스어 "διπλόω"와 덮개판을 의미하는 "operculum"의 합성어) 하목으로 묶인다[73]。
다음은 World Spider Catalog에 게재된 화석협각류 목록표 "A summary list of fossil spiders and their relatives"(2020년 1월 15일 최종 갱신)에 근거한 바다전갈아목의 분류 체계이다[68]。
- '''바다전갈목''' †(=오오사소리목 Gigantostraca, Cyrtoctenida)(오르도비스기 - 페름기)
- * '''바다전갈아목''' [6](오르도비스기 - 페름기)
- ** '''오니코프테렐라상과'''(오르도비스기 - 실루리아기)
'''오니코프테렐라과'''
* (데본기)
* 오니코프테렐라[70] (오르도비스기)
* (실루리아기)
- ** '''모세로프테루스상과'''(실루리아기 - 데본기)
'''모세로프테루스과'''
* 모세로프테루스[70] (데본기)
* 스토엘메로프테루스[72] (실루리아기)
* (데본기)
- ** '''믹소프테루스상과'''[6](Lamsdell et al. (2013) 이후의 계통분석에서 하목에 포함됨[73][32][7][69])(오르도비스기)
'''믹소프테루스과'''[6]
*
* 메갈로그랍투스
* 펜테코프테루스 [7]
(과 소속 불확실)
* (오르도비스기)
* (오르도비스기)
'''돌리코프테루스과'''[6](실루리아기 - 데본기)
* (실루리아기)
* 돌리코프테루스[70] (실루리아기)
* (실루리아기)
'''바다전갈과'''[6] (실루리아기)
* 유리프테루스 (=)
'''에리오프테루스과'''(실루리아기 - 데본기)
* 에리오프테루스[70]
'''스트로빌로프테루스과'''(실루리아기 - 데본기)
* (실루리아기)
*스트로빌로프테루스[70] (=, )(실루리아기 - 데본기)
'''카르키노소마상과''' (= 믹소프테루스상과[6]) (오르도비스기 - 데본기)
* '''카르키노소마과'''[6] (오르도비스기 - 데본기)
**카르키노소마[70] (= ''Eurysoma'') (실루리아기)
** 에오카르키노소마[70] (오르도비스기)
** (=, 파라카르키노소마[70] ) (실루리아기 - 데본기)
** 리노카르키노소마[70] (실루리아기)
* '''믹소프테루스과'''[6] (=Lanarkopteridae) (실루리아기)
**
** 믹소프테루스
'''바에링오프테루스상과''' (정식 기재되지 않음[68]) (오르도비스기 - 데본기)
* (데본기)
* (오르도비스기)
* (실루리아기)
'''아델로프탈무스상과'''[6] (데본기 - 페름기)
* '''아델로프탈무스과'''[6]
** (=, , , ) (데본기 - 페름기)
** (실루리아기)
** (실루리아기)
** (실루리아기 - 데본기)
** (실루리아기 - 데본기)
** (실루리아기)
'''프테리고투스상과''' (실루리아기 - 데본기)
* '''휴밀레리아과'''[6] (실루리아기)
**
** 휴밀레리아
* '''슬리모니아과''' (실루리아기)
**
** 슬리모니아
* '''프테리고투스과'''[6] (= 야켈로프테루스과[6] Jaekelopteridae) (실루리아기 - 데본기)
** 아쿠티라무스 (실루리아기 - 데본기)
** 키우르콥테루스 (실루리아기)
** 에레토프테루스 (=) (실루리아기 - 데본기)
** 야켈로프테루스 (데본기)
** 네크로감마루스 (실루리아기)
** 프테리고투스 (=) (실루리아기 - 데본기)
5. 2. 스틸로누루스아목 (Stylonurina)
바다전갈은 크게 광익아목(Eurypterina)과 스틸로누루스아목(Stylonurina)의 두 아목으로 나뉜다. 스틸로누루스아목은 다섯째 다리쌍이 변형되지 않아 가느다란 모양이며, '사이마디'가 없고, 두흉부 배판에 횡방향 봉합선이 있다는 점에서 광익아목과 구별된다.[82]2020년 기준으로 약 250종의 바다전갈류가 알려져 있으며, 2개의 아목, 12개의 상과, 21개의 과, 그리고 약 70개의 속으로 분류된다.[68] 바다전갈류는 크게 스틸로누루스아목(Stylonurina)과 광익아목(Eurypterina)으로 나뉘며, 대부분의 종은 광익아목에 속한다.[48][16][68] 스틸로누루스아목은 원시적인 형태를 띠고 있어, 추가적인 계통 분석을 통해 광익아목에 대한 측계통군이 될 가능성도 있지만, 두 아목 모두 단계통군이며 자매군이라는 점이 대부분의 계통 분석에서 지지받고 있다.[16][28][73][32][7][69]
히베르토프테루스과(Hibbertopteridae)를 별도의 분류군으로 보는 의견도 있지만, 이 과는 스틸로누루스아목에 속하는 파생군으로 보는 것이 일반적이다.[28][69][68]
다음은 스틸로누루스아목의 분류 체계와 각 지질 시대이다. (각주와 특기하지 않는 한 World Spider Catalog의 화석 협각류 목록표 "A summary list of fossil spiders and their relatives"(최종 갱신일: 2020년 1월 15일)에 근거[68]) 속보다 상위 분류군은 굵게 표시한다.
- '''바다전갈목(Eurypterida)''' † (=오오사소리목 Gigantostraca, Cyrtoctenida) (오르도비스기 - 페름기)
- '''스틸로누루스아목(Stylonurina)'''[6] (=Woodwardopterina, Hibbertopterina) (오르도비스기 - 페름기)
- (상과·과 미정)
- (''Stylonuroides'') (실루리아기 - 데본기)
- '''레노프테루스상과(Rhenopteroidea)''' (=Brachyopterelloidea) (오르도비스기 - 데본기)
- '''레노프테루스과(Rhenopteridae)'''[70] (=Brachyopterellidae)
- 브라키오프테렐라(Brachyopterella)[70] (실루리아기)
- 브라키오프테루스(Brachyopterus)[70] (오르도비스기)
- 키아에로프테루스(Kiaeropterus)[70] (실루리아기)
- (Leiopterella) (데본기)
- 레노프테루스(Rhenopterus)[70] (데본기)
- '''긴꼬리바다전갈상과(Stylonuroidea)'''[6] (=Drepanopteroidea) (실루리아기 - 데본기)
- '''파라스티로누루스과(Parastylonuridae)''' (실루리아기 - 데본기)
- 파라스티로누루스(Parastylonurus)[70] (실루리아기)
- (Stylonurella) (실루리아기? - 데본기)
- '''긴꼬리바다전갈과(Stylonuridae)'''[6] (=Laurieipteridae, Pageidae) (실루리아기 - 데본기)
- (Ctenopterus) (실루리아기)
- (Laurieipterus) (실루리아기)
- 파게아(Pagea)[70] (데본기)
- 스티로누루스(Stylonurus) (데본기)
- (Soligorskopterus)[71] (데본기)
- '''코코모프테루스상과(Kokomopteroidea)''' (실루리아기)
- '''코코모프테루스과(Kokomopteridae)'''
- 코코모프테루스(Kokomopterus)
- (Lamontopterus)
- '''할디에오프테루스과(Hardieopteridae)'''
- 할립테루스(Hallipterus)[72] (실루리아기)
- 할디에오프테루스(Hardieopterus)[70] (실루리아기)
- (Tarsopterella) (실루리아기)
- '''믹테로프스상과(Mycteropoidea)''' (=히베르토프테루스상과 Hibbertopteroidea) (실루리아기 - 페름기)
- '''가마아시바다전갈과(Drepanopteridae)'''[6] (실루리아기 - 데본기)
- 드레파노프테루스(Drepanopterus)[70] (실루리아기 - 데본기)
- '''히베르토프테루스과(Hibbertopteridae)''' (=Cyrtoctenidae) (데본기 - 페름기)
- 캄필로세팔루스(Campylocephalus)[70] (석탄기 - 페름기)
- (Cyrtoctenus) (데본기 - 석탄기)
- (Dunsopterus) (데본기 - 석탄기)
- 하스티미마(Hastimima)[70] (페름기)
- 히베르토프테루스(Hibbertopterus) (석탄기 - 페름기)
- (Vernonopterus) (석탄기 - 페름기)
- '''믹테로프스과(Mycteropidae)''' (=우드워드바다전갈과[6] Woodwardopteridae) (석탄기 - 페름기)
- 메가라크네(Megarachne)[70] (석탄기 - 페름기)
- 믹테로프스(Mycterops)[70] (석탄기)
- (Woodwardopterus) (석탄기 - ?페름기[8])
6. 진화

1882년까지 실루리아기 이전의 바다전갈은 알려진 바가 없었다. 1880년대 이후의 발견으로 오르도비스기 초기 바다전갈에 대한 지식이 확장되었다.[1] 현재 알려진 가장 초기의 바다전갈인 메갈로그라프투스과의 ''펜테코프테루스''는 중기 오르도비스기의 다리윌기 (4억 6730만 년 전)로 거슬러 올라간다.[2] 모로코의 후기 트레마도키기 (초기 오르도비스기) 페조우아타 생물군에서 더 오래된 바다전갈 화석이 보고되었지만, 아직 충분히 연구되지 않았으며,[3] 페토이드 부속지일 가능성이 높다.
''펜테코프테루스''는 상대적으로 진화된 바다전갈로, 카르시노소마토이드 상과 내의 메갈로그라프투스과에 속한다. 이는 적어도 바다전갈아목 내에서 대부분의 바다전갈 분지군이 중기 오르도비스기 동안 이미 확립되었음을 시사한다.[4] 가장 초기의 스틸로누린 바다전갈인 ''브라키옵테루스''[5] 역시 중기 오르도비스기 시대이다. 두 아목 구성원의 존재는 원시적인 줄기 바다전갈이 그들보다 앞섰을 것이라는 것을 나타내지만, 화석 기록에서는 알려지지 않았다. 중기 오르도비스기 동안 여러 바다전갈 분지군이 존재한다는 것은 바다전갈이 초기 오르도비스기 동안 기원하여 최초 형태가 진화한 직후 급격하고 폭발적인 방사 및 다양화를 경험했거나, 캄브리아기 동안 기원했음을 시사한다.[4]
따라서 정확한 바다전갈의 기원은 여전히 알려져 있지 않다. "원시 바다전갈"이라고 불리는 화석이 캄브리아기나 선캄브리아기 시대의 퇴적층에서 묘사되었지만,[6] 오늘날에는 바다전갈로, 심지어 관련 형태로도 인식되지 않는다. 이전에 원시 바다전갈로 여겨졌던 ''스트라봅스''[7]는 현재 아글라스피드류 또는 스트라보포스류로 분류된다. 아글라스피드류는 한때 원시 협각류로 여겨졌지만 현재 삼엽충과 더 밀접한 관련이 있는 그룹으로 여겨진다.[8]
오르도비스기 바다전갈의 화석 기록은 매우 부족하다. 한때 오르도비스기에서 알려진 바다전갈의 대부분은 오인 또는 가화석으로 판명되었다. 오늘날 오르도비스기 바다전갈을 나타내는 종은 11종에 불과하다. 이들은 두 가지 생태적 범주로 나뉜다. 고대 대륙 로렌시아의 크고 활동적인 포식자와 저서 (해저에서 생활) 및 대륙 아발로니아 및 곤드와나에서 온 기저 동물이다.[2] 메갈로그라프투스과(속 ''에키노그나투스'', ''메갈로그라프투스'', ''펜테코프테루스'' 포함)에 분류된 로렌시아 포식자는 후기 오르도비스기 동안 소규모 방사를 경험하면서 최초로 성공적인 바다전갈 그룹을 나타낼 가능성이 높다.[9]
유해류는 전기 데본기 동안 풍부하게 존재하며 다양성을 유지했고 (모든 절지동물 중 가장 큰 종인 프테리고티드 ''자이켈롭테루스''의 진화), 이 그룹은 후기 데본기 대멸종의 영향을 크게 받은 많은 그룹 중 하나였다. 이 멸종 사건은 해양 생물 (특히 삼엽충, 완족류, 산호초 형성 생물)에만 영향을 미치는 것으로 알려져 있으며, 유해류의 풍부함과 다양성을 약화시켰다.[10]
전기 데본기 동안 다양성이 크게 감소하기 시작했고, 유해류는 후기 데본기까지 해양 환경에서 드물게 발견되었다. 프라니안 단계 동안 4개의 과가 멸종되었고, 파메니안에서는 5개의 과가 추가로 멸종되었다.[10] 해양 그룹이 가장 큰 영향을 받았기 때문에 유해류는 주로 유해류 아목 내에서 영향을 받았다. 단 하나의 스틸로누린 그룹 (Parastylonuridae 과)만이 전기 데본기에 멸종되었다. 단 두 개의 유해류 과 (Adelophthalmidae, Waeringopteridae)만이 후기 데본기까지 살아남았다. 유해류는 전기 데본기 동안 가장 큰 감소를 경험했는데, 1천만 년 만에 다양성의 50% 이상을 잃었다. 스틸로누린은 비교적 일정한 다양성과 풍부함을 유지했지만, 후기 데본기 동안 영향을 받아 많은 오래된 그룹이 Mycteroptidae 과와 Hibbertopteridae 과의 새로운 형태로 대체되었다.[11]
유해류 아목의 파멸적인 멸종 패턴은 더 진화된 물고기의 출현과 관련이 있을 수 있다. 유해류의 감소는 무악어류가 처음으로 더 발달하게 된 시점에 시작되었으며, 북아메리카와 유럽에서 판피어류 (갑옷을 입은 물고기)의 출현과 일치한다.[12]
생존한 hibbertopterid 과와 mycteroptid과의 스틸로누린은 새로운 생태적 지위로 진화함으로써 물고기와의 경쟁을 피했다. 이들은 후기 데본기와 전기 석탄기를 통해 방산과 다양화를 경험했으며, 유해류 내에서 마지막으로 일어난 방산으로 "쓸어 먹는" (먹이를 찾기 위해 기질을 긁는) 능력을 가진 여러 새로운 형태가 등장했다.[13]
7. 연구사
유해류는 전기 데본기 동안 풍부하게 존재하며 다양성을 유지했지만, 후기 데본기 대멸종의 영향을 크게 받았다. 이 멸종은 해양 생물에만 영향을 미쳐 유해류의 풍부함과 다양성을 약화시켰다.[12] 이미 전기 데본기부터 다양성이 감소하기 시작하여 후기 데본기에는 해양 환경에서 드물게 발견되었다. 프라니안 단계 동안 4개의 과, 파메니안 단계에서 5개의 과가 멸종했다.[13] 유해류는 주로 유해류 아목 내에서 영향을 받았으며, 스틸로누린 그룹 중에서는 Parastylonuridae 과만이 전기 데본기에 멸종했다. 단 두 개의 유해류 과(Adelophthalmidae 및 Waeringopteridae)만이 후기 데본기까지 살아남았다. 유해류는 전기 데본기에 가장 큰 감소를 겪어 1천만 년 만에 다양성의 50% 이상을 잃었다. 반면 스틸로누린은 비교적 일정한 다양성과 풍부함을 유지했지만, 후기 데본기에 영향을 받아 Mycteroptidae 과와 Hibbertopteridae 과의 새로운 형태로 대체되었다.[14]
이러한 멸종 패턴은 더 진화된 물고기의 출현과 관련이 있을 수 있다. 유해류의 감소는 무악어류가 발달하기 시작한 시점과 북아메리카 및 유럽에서 판피어류의 출현과 일치한다.[15]
생존한 hibbertopterid 과와 mycteroptid과의 스틸로누린은 새로운 생태적 지위로 진화하여 물고기와의 경쟁을 피했다. 이들은 후기 데본기와 전기 석탄기를 거치며 방산과 다양화를 경험했고, "쓸어 먹는" 능력을 가진 여러 새로운 형태가 등장했다.[16]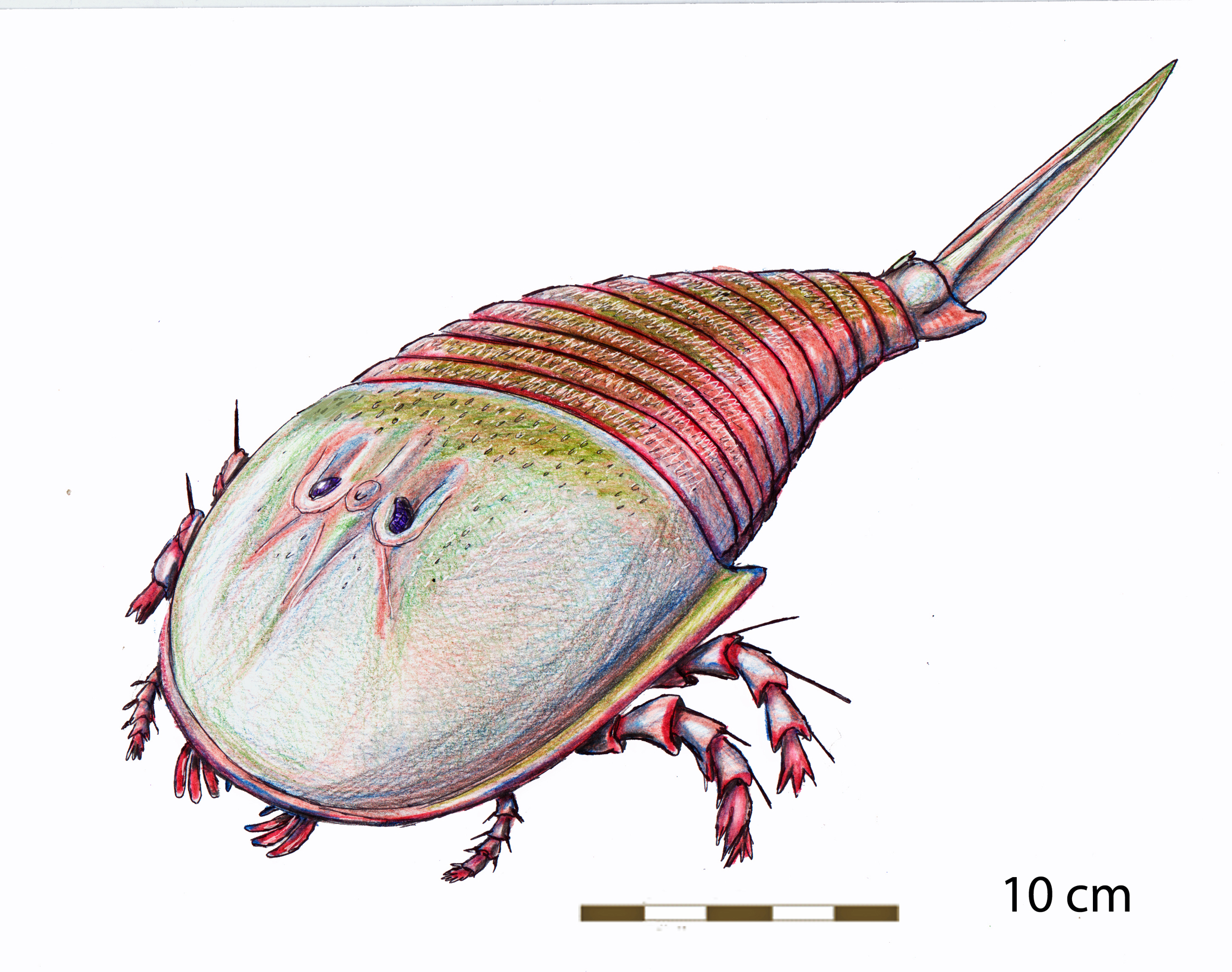
Adelophthalmidae, Hibbertopteridae 및 Mycteroptidae 3개의 유절류 과만이 멸종 사건에서 완전히 살아남았다. 이들은 모두 담수 동물로 추정되었으며, 이는 유절류가 해양 환경에서 멸종했음을 의미한다.[17] 해양 유절류 포식자가 사라지면서 육기어류인 리지도돈트 등이 새로운 최상위 포식자가 되었다.[18] 그러나 최근 발견에 따르면, 이 유절류가 강어귀, 삼각주, 석호 및 해안 연못과 같은 변두리 해양 환경에 서식했을 가능성도 제기된다.[19] 유일하게 살아남은 유절류 과인 Adelophthalmidae는 ''Adelophthalmus'' 속으로 대표된다. 히베르토프테리드, 믹테로프테리드 및 ''Adelophthalmus''는 페름기까지 살아남았다.[20]
''Adelophthalmus''는 다른 유절류가 없는 상황에서 번성하여 모든 후기 고생대 유절류 중 가장 흔해졌고, 생존하는 stylonurine보다 더 많은 수와 다양성을 보였다.[21] ''Adelophthalmus''의 33종 중 23종(69%)이 석탄기에서만 발견되었다.[22][23] 이 속은 후기 석탄기에 가장 높은 다양성을 보였다. 판게아 초대륙 형성으로 ''Adelophthalmus''는 거의 전 세계적인 분포를 얻었다.[24]
후기 석탄기와 초기 페름기 동안 ''Adelophthalmus''는 널리 분포했으며, 주로 해안 평원에 인접한 기수 및 담수 환경에서 서식했다. 그러나 판게아 형성에 따른 기후 변화로 서식지가 사라지면서 수가 줄어들었고, 초기 페름기의 Leonardian 단계에 멸종했다.[25]
믹테로프테리드와 히베르토프테리드는 일정 기간 생존했으며, 페름기 지층에서 알려진 속은 ''Hastimima''와 ''Campylocephalus''였다.[26] ''Hastimima''는 초기 페름기에 멸종했고,[27] ''Campylocephalus''는 더 오래 생존했다. 러시아 페름기 퇴적층에서 발견된 ''C. permianus'' 종의 화석은 1.4m에 달했을 수 있다. 다른 자료에서는 Kungurian-Roadian 시대(약 2억 8300만 년 전 ~ 2억 6700만 년 전)를 제시한다.[29] 오스트레일리아 페름기 말기의 ''Woodwardopterus freemanorum'' 종은 알려진 가장 어린 유절류이다.[30] 마지막 유절류는 페름기-트라이아스기 대멸종 때 또는 그 직전에 멸종했다. 이 멸종은 기록상 가장 파괴적인 대멸종으로, 삼엽충 등 다른 고생대 그룹도 멸종시켰다.[31]
7. 1. 초기 연구 (1818년 ~ 1859년)
1882년까지 실루리아기 이전의 바다전갈은 알려진 바가 없었다. 1880년대 이후의 발견으로 오르도비스기 시대의 초기 바다전갈에 대한 지식이 확장되었다.[1] 오늘날 알려진 가장 초기의 바다전갈인 메갈로그라프투스과의 ''펜테코프테루스''는 중기 오르도비스기의 다리윌기 절, 즉 4억 6730만 년 전으로 거슬러 올라간다.[2] 모로코의 후기 트레마도키기 (초기 오르도비스기) 시대의 페조우아타 생물군에서 훨씬 더 오래된 바다전갈 화석이 보고되었지만, 아직 충분히 연구되지 않았으며, 페토이드 부속지일 가능성이 높다.[3]''펜테코프테루스''는 상대적으로 진화된 바다전갈로, 카르시노소마토이드 상과 내의 메갈로그라프투스과에 속한다. 이러한 진화된 위치는 적어도 바다전갈아목 내에서 대부분의 바다전갈 분지군이 중기 오르도비스기 동안 이미 확립되었음을 시사한다.[4] 가장 초기의 알려진 스틸로누린 바다전갈인 ''브라키옵테루스''[5] 역시 중기 오르도비스기 시대이다. 두 아목 모두의 구성원이 존재한다는 것은 원시적인 줄기 바다전갈이 그들보다 앞섰을 것이라는 것을 나타내지만, 이것들은 지금까지 화석 기록에서 알려지지 않았다. 중기 오르도비스기 동안 여러 바다전갈 분지군이 존재한다는 것은 바다전갈이 초기 오르도비스기 동안 기원하여 최초 형태가 진화한 직후 급격하고 폭발적인 방사 및 다양화를 경험했거나, 그룹이 훨씬 더 일찍, 아마도 캄브리아기 동안 기원했음을 시사한다.[4]
따라서 정확한 바다전갈의 기원은 여전히 알려져 있지 않다. "원시 바다전갈"이라고 불리는 화석이 때때로 캄브리아기나 심지어 선캄브리아기 시대의 퇴적층에서 묘사되었지만,[6] 오늘날에는 바다전갈은 물론 관련 형태로도 인식되지 않는다. 이전에 원시 바다전갈로 여겨졌던 일부 동물, 예를 들어 미주리주의 캄브리아기에서 발견된 ''스트라봅스''[7]는 현재 아글라스피드류 또는 스트라보포스류로 분류된다. 한때 원시 협각류로 여겨졌던 아글라스피드류는 현재 삼엽충과 더 밀접한 관련이 있는 그룹으로 여겨진다.[8]
7. 2. 연구의 발전 (1859년 ~ 1912년)
1882년까지 실루리아기 이전의 바다전갈은 알려진 바가 없었다. 1880년대 이후의 동시대 발견은 오르도비스기 시대의 초기 바다전갈에 대한 지식을 확장시켰다.[1] 오늘날 알려진 가장 초기의 바다전갈인 메갈로그라프투스과의 ''펜테코프테루스''는 중기 오르도비스기의 다리윌기 절, 즉 4억 6730만 년 전으로 거슬러 올라간다.[2] 모로코의 후기 트레마도키기 (초기 오르도비스기) 시대의 페조우아타 생물군에서 훨씬 더 오래된 바다전갈 화석이 보고되었지만, 아직 충분히 연구되지 않았으며,[3] 페토이드 부속지일 가능성이 높다.
''펜테코프테루스''는 상대적으로 진화된 바다전갈로, 카르시노소마토이드 상과 내의 메갈로그라프투스과에 속한다. 이러한 진화된 위치는 적어도 바다전갈아목 내에서 대부분의 바다전갈 분지군이 중기 오르도비스기 동안 이미 확립되었음을 시사한다.[4] 가장 초기의 알려진 스틸로누린 바다전갈인 ''브라키옵테루스''[5] 역시 중기 오르도비스기 시대이다. 두 아목 모두의 구성원이 존재한다는 것은 원시적인 줄기 바다전갈이 그들보다 앞섰을 것이라는 것을 나타내지만, 이것들은 지금까지 화석 기록에서 알려지지 않았다. 중기 오르도비스기 동안 여러 바다전갈 분지군이 존재한다는 것은 바다전갈이 초기 오르도비스기 동안 기원하여 최초 형태가 진화한 직후 급격하고 폭발적인 방사 및 다양화를 경험했거나, 그룹이 훨씬 더 일찍, 아마도 캄브리아기 동안 기원했음을 시사한다.[6]
따라서 정확한 바다전갈의 기원은 여전히 알려져 있지 않다. "원시 바다전갈"이라고 불리는 화석이 때때로 캄브리아기나 심지어 선캄브리아기 시대의 퇴적층에서 묘사되었지만,[7] 오늘날에는 바다전갈로, 심지어 관련 형태로도 인식되지 않는다. 이전에 원시 바다전갈로 여겨졌던 일부 동물, 예를 들어 미주리주의 캄브리아기에서 발견된 ''스트라봅스'' 속[8]은 현재 아글라스피드류 또는 스트라보포스류로 분류된다. 한때 원시 협각류로 여겨졌던 아글라스피드류는 현재 삼엽충과 더 밀접한 관련이 있는 그룹으로 여겨진다.[9]
오르도비스기 바다전갈의 화석 기록은 매우 부족하다. 한때 오르도비스기에서 알려진 바다전갈의 대부분은 오인 또는 가화석으로 판명되었다. 오늘날 오르도비스기 바다전갈을 나타내는 것으로 확실하게 식별할 수 있는 종은 11종에 불과하다. 이러한 분류군은 두 가지 뚜렷한 생태적 범주로 나뉜다. 고대 대륙 로렌시아의 크고 활동적인 포식자와 저서 (해저에서 생활) 및 대륙 아발로니아 및 곤드와나에서 온 기저 동물이다.[10] 메갈로그라프투스과(''에키노그나투스'', ''메갈로그라프투스'', ''펜테코프테루스'' 포함)에 분류된 로렌시아 포식자는 후기 오르도비스기 동안 소규모의 방사를 경험하면서 최초로 성공적인 바다전갈 그룹을 나타낼 가능성이 높다.[11]
유해류는 전기 데본기 동안에도 풍부하게 존재하며 다양성을 유지했고 (예를 들어, 모든 절지동물 중 가장 큰 종인 프테리고티드 ''Jaekelopterus''의 진화를 이끌었다), 이 그룹은 후기 데본기 대멸종의 영향을 크게 받은 많은 그룹 중 하나였다. 이 멸종 사건은 해양 생물 (특히 삼엽충, 완족류 및 산호초를 형성하는 생물)에만 영향을 미치는 것으로 알려져 있으며, 이전에 유해류 내에서 관찰되었던 풍부함과 다양성을 효과적으로 약화시켰다.[12]
이미 전기 데본기 동안 다양성이 크게 감소하기 시작했고, 유해류는 후기 데본기까지 해양 환경에서 드물게 발견되었다. 프라니안 단계 동안 4개의 과가 멸종되었고, 이후 파메니안에서는 5개의 과가 추가로 멸종되었다.[13] 해양 그룹이 가장 큰 영향을 받았기 때문에 유해류는 주로 유해류 아목 내에서 영향을 받았다. 단 하나의 스틸로누린 그룹 (Parastylonuridae 과)만이 전기 데본기에 멸종되었다. 단 두 개의 유해류 과 (Adelophthalmidae 및 Waeringopteridae)만이 후기 데본기까지 살아남았다. 유해류는 전기 데본기 동안 가장 큰 감소를 경험했는데, 이 기간 동안 단 1천만 년 만에 다양성의 50% 이상을 잃었다. 반면에 스틸로누린은 비교적 일정한 다양성과 풍부함을 유지하며 이 기간을 거쳤지만, 후기 데본기 동안 영향을 받아 많은 오래된 그룹이 Mycteroptidae 과와 Hibbertopteridae 과의 새로운 형태로 대체되었다.[14]
유해류 아목에서 관찰된 파멸적인 멸종 패턴은 더 진화된 물고기의 출현과 관련이 있을 수 있다. 유해류의 감소는 무악어류가 처음으로 더 발달하게 된 시점에 시작되었으며, 북아메리카와 유럽에서 판피어류 (갑옷을 입은 물고기)의 출현과 일치한다.[15]
생존한 hibbertopterid 과와 mycteroptid 과의 스틸로누린은 새로운 독특한 생태적 지위로 진화함으로써 물고기와의 경쟁을 완전히 피했다. 이 과들은 후기 데본기와 전기 석탄기를 통해 방산과 다양화를 경험했으며, 유해류 내에서 마지막으로 일어난 방산으로 "쓸어 먹는" (먹이를 찾기 위해 기질을 긁는) 능력을 가진 여러 새로운 형태가 등장했다.[16]
단 3개의 유절류 과 - Adelophthalmidae, Hibbertopteridae 및 Mycteroptidae - 만이 멸종 사건에서 완전히 살아남았다. 이들은 모두 담수 동물로 추정되었고, 이는 유절류가 해양 환경에서 멸종했을 것이라는 것을 의미한다.[17] 해양 유절류 포식자가 사라지면서, 육기어류, 예를 들어 리지도돈트와 같은 종이 해양 환경의 새로운 최상위 포식자가 되었다.[18] 그러나, 다양한 최근의 발견은 이에 대한 의문을 제기하며, 이 유절류가 강어귀, 삼각주, 석호 및 해안 연못과 같은 변두리 해양 환경에 서식했던 유염성 형태였을 가능성을 시사한다. 한 가지 주장은 고생물지리학적이다. 프테리고토이드의 분포는 해양 분산을 필요로 하는 것으로 보인다.[19] 최근 Adelophthalmoidea에 대한 검토에서는 "이전에 인정되었던 것보다 많은 'Adelophthalmus'가 산출되는 많은 구역에서 해양의 영향이 훨씬 더 크다"고 인정했다.[2] 마찬가지로, 뉴멕시코의 석탄기 유절류 ''Hibbertopterus''에 대한 연구는 일부 유절류의 서식지가 "재평가될 필요가 있을 수 있다"고 결론지었다.[3] 유일하게 살아남은 유절류 과인 Adelophthalmidae는 단일 속인 ''Adelophthalmus''로 대표되었다. 히베르토프테리드, 믹테로프테리드 및 ''Adelophthalmus''는 페름기까지 살아남았다.[20]
''Adelophthalmus''는 다른 유절류가 없는 상황에서 번성하며, 모든 후기 고생대 유절류 중에서 가장 흔해졌고, 생존하는 stylonurine보다 더 많은 수와 다양성을 보였다.[21] ''Adelophthalmus''에 속하는 33종 중 23종(69%)이 석탄기에서만 발견되었다.[22][23] 이 속은 후기 석탄기에 가장 높은 다양성을 보였다. 비록 ''Adelophthalmus''가 이미 후기 데본기에 비교적 널리 분포하고 있었고 모든 주요 육지를 대표했지만, 고생대 마지막 두 기간 동안 판게아가 전 세계적인 초대륙으로 합쳐지면서 ''Adelophthalmus''는 거의 전 세계적인 분포를 얻을 수 있었다.[24]
후기 석탄기와 초기 페름기 동안 ''Adelophthalmus''는 널리 분포했으며, 주로 해안 평원에 인접한 기수 및 담수 환경에서 서식했다. 이러한 환경은 유리한 기후 조건에 의해 유지되었다. 판게아의 형성에 따른 기후 변화가 전 세계의 퇴적 및 식생 패턴을 변화시키면서, 이들은 지속되지 못했다. 서식지가 사라지면서 ''Adelophthalmus''는 수가 줄어들었고, 이미 초기 페름기의 Leonardian 단계까지 멸종했다.[25]
믹테로프테리드와 히베르토프테리드는 일정 기간 생존을 지속했으며, 각각의 그룹에서 페름기 지층에서 알려진 속은 ''Hastimima''와 ''Campylocephalus''였다.[26] ''Hastimima''는 ''Adelophthalmus''와 마찬가지로 초기 페름기에 멸종했고,[27] ''Campylocephalus''는 더 오래 생존했다. 러시아의 페름기 퇴적층에서 발견된 거대한 불완전한 등딱지는 ''C. permianus'' 종의 유일한 화석 유물이다. 원래 후기 페름기 시대라고 여겨졌지만[28] 다른 자료에서는 이 표본에 대해 Kungurian-Roadian 시대(약 2억 8300만 년 전 ~ 2억 6700만 년 전)를 제시한다.[29] 유절류 큐티클의 조각인 ''Woodwardopterus freemanorum'' 종은 오스트레일리아의 페름기 말기에서 알려져 있으며, 이는 알려진 가장 어린 유절류를 나타낸다.[30] 페름기보다 높은 화석층에서는 유절류가 알려져 있지 않다. 이는 마지막 유절류가 종말적인 멸종 사건이 일어났을 때, 또는 그 직전에 죽었음을 나타낸다. 이 멸종 사건인 페름기-트라이아스기 대멸종은 기록된 가장 파괴적인 대멸종이며, 삼엽충과 같은 다른 성공적인 고생대 그룹을 멸종시켰다.[31]
7. 3. 현대 연구 (1912년 이후)
1882년까지 실루리아기 이전의 바다전갈은 알려진 바가 없었다. 1880년대 이후의 동시대 발견은 오르도비스기 시대의 초기 바다전갈에 대한 지식을 확장시켰다.[1] 오늘날 알려진 가장 초기의 바다전갈인 메갈로그라프투스과의 ''펜테코프테루스''는 중기 오르도비스기의 다리윌기 절, 즉 4억 6730만 년 전으로 거슬러 올라간다.[2] 모로코의 후기 트레마도키기 (초기 오르도비스기) 시대의 페조우아타 생물군에서 훨씬 더 오래된 바다전갈 화석이 보고되었지만, 아직 충분히 연구되지 않았으며,[3] 페토이드 부속지일 가능성이 높다.
''펜테코프테루스''는 상대적으로 진화된 바다전갈로, 카르시노소마토이드 상과 내의 메갈로그라프투스과에 속한다. 이러한 진화된 위치는 적어도 바다전갈아목 내에서 대부분의 바다전갈 분지군이 중기 오르도비스기 동안 이미 확립되었음을 시사한다.[4] 가장 초기의 알려진 스틸로누린 바다전갈인 ''브라키옵테루스''[5] 역시 중기 오르도비스기 시대이다. 두 아목 모두의 구성원이 존재한다는 것은 원시적인 줄기 바다전갈이 그들보다 앞섰을 것이라는 것을 나타내지만, 이것들은 지금까지 화석 기록에서 알려지지 않았다. 중기 오르도비스기 동안 여러 바다전갈 분지군이 존재한다는 것은 바다전갈이 초기 오르도비스기 동안 기원하여 최초 형태가 진화한 직후 급격하고 폭발적인 방사 및 다양화를 경험했거나, 그룹이 훨씬 더 일찍, 아마도 캄브리아기 동안 기원했음을 시사한다.[6]
정확한 바다전갈의 기원은 여전히 알려져 있지 않다. "원시 바다전갈"이라고 불리는 화석이 때때로 캄브리아기나 심지어 선캄브리아기 시대의 퇴적층에서 묘사되었지만,[7] 오늘날에는 바다전갈로, 심지어 관련 형태로도 인식되지 않는다. 이전에 원시 바다전갈로 여겨졌던 일부 동물, 예를 들어 미주리주의 캄브리아기에서 발견된 ''스트라봅스'' 속[8]은 현재 아글라스피드류 또는 스트라보포스류로 분류된다. 한때 원시 협각류로 여겨졌던 아글라스피드류는 현재 삼엽충과 더 밀접한 관련이 있는 그룹으로 여겨진다.[9]
오르도비스기 바다전갈의 화석 기록은 매우 부족하다. 한때 오르도비스기에서 알려진 바다전갈의 대부분은 오인 또는 가화석으로 판명되었다. 오늘날 오르도비스기 바다전갈을 나타내는 것으로 확실하게 식별할 수 있는 종은 11종에 불과하다. 이러한 분류군은 두 가지 뚜렷한 생태적 범주로 나뉜다. 고대 대륙 로렌시아의 크고 활동적인 포식자와 저서 (해저에서 생활) 및 대륙 아발로니아 및 곤드와나에서 온 기저 동물이다.[10] 메갈로그라프투스과(''에키노그나투스'', ''메갈로그라프투스'', ''펜테코프테루스'' 포함)에 분류된 로렌시아 포식자는 후기 오르도비스기 동안 소규모의 방사를 경험하면서 최초로 성공적인 바다전갈 그룹을 나타낼 가능성이 높다.[11]
유해류는 전기 데본기 동안에도 풍부하게 존재하며 다양성을 유지했고 (예를 들어, 모든 절지동물 중 가장 큰 종인 프테리고티드 ''Jaekelopterus''의 진화를 이끌었다), 이 그룹은 후기 데본기 대멸종의 영향을 크게 받은 많은 그룹 중 하나였다. 이 멸종 사건은 해양 생물 (특히 삼엽충, 완족류 및 산호초를 형성하는 생물)에만 영향을 미치는 것으로 알려져 있으며, 이전에 유해류 내에서 관찰되었던 풍부함과 다양성을 효과적으로 약화시켰다.[12]
이미 전기 데본기 동안 다양성이 크게 감소하기 시작했고, 유해류는 후기 데본기까지 해양 환경에서 드물게 발견되었다. 프라니안 단계 동안 4개의 과가 멸종되었고, 이후 파메니안에서는 5개의 과가 추가로 멸종되었다.[13] 해양 그룹이 가장 큰 영향을 받았기 때문에 유해류는 주로 유해류 아목 내에서 영향을 받았다. 단 하나의 스틸로누린 그룹 (Parastylonuridae 과)만이 전기 데본기에 멸종되었다. 단 두 개의 유해류 과 (Adelophthalmidae 및 Waeringopteridae)만이 후기 데본기까지 살아남았다. 유해류는 전기 데본기 동안 가장 큰 감소를 경험했는데, 이 기간 동안 단 1천만 년 만에 다양성의 50% 이상을 잃었다. 반면에 스틸로누린은 비교적 일정한 다양성과 풍부함을 유지하며 이 기간을 거쳤지만, 후기 데본기 동안 영향을 받아 많은 오래된 그룹이 Mycteroptidae 과와 Hibbertopteridae 과의 새로운 형태로 대체되었다.[14]
유해류 아목에서 관찰된 파멸적인 멸종 패턴은 더 진화된 물고기의 출현과 관련이 있을 수 있다. 유해류의 감소는 무악어류가 처음으로 더 발달하게 된 시점에 시작되었으며, 북아메리카와 유럽에서 판피어류 (갑옷을 입은 물고기)의 출현과 일치한다.[15]
생존한 hibbertopterid 과와 mycteroptid 과의 스틸로누린은 새로운 독특한 생태적 지위로 진화함으로써 물고기와의 경쟁을 완전히 피했다. 이 과들은 후기 데본기와 전기 석탄기를 통해 방산과 다양화를 경험했으며, 유해류 내에서 마지막으로 일어난 방산으로 "쓸어 먹는" (먹이를 찾기 위해 기질을 긁는) 능력을 가진 여러 새로운 형태가 등장했다.[16]
단 3개의 유절류 과 - Adelophthalmidae, Hibbertopteridae 및 Mycteroptidae - 만이 멸종 사건에서 완전히 살아남았다. 이들은 모두 담수 동물로 추정되었고, 이는 유절류가 해양 환경에서 멸종했을 것이라는 것을 의미한다.[17] 해양 유절류 포식자가 사라지면서, 육기어류, 예를 들어 리지도돈트와 같은 종이 해양 환경의 새로운 최상위 포식자가 되었다.[18] 그러나, 다양한 최근의 발견은 이에 대한 의문을 제기하며, 이 유절류가 강어귀, 삼각주, 석호 및 해안 연못과 같은 변두리 해양 환경에 서식했던 유염성 형태였을 가능성을 시사한다. 한 가지 주장은 고생물지리학적이다. 프테리고토이드의 분포는 해양 분산을 필요로 하는 것으로 보인다.[19] 최근 Adelophthalmoidea에 대한 검토에서는 "이전에 인정되었던 것보다 많은 'Adelophthalmus'가 산출되는 많은 구역에서 해양의 영향이 훨씬 더 크다"고 인정했다.[2] 마찬가지로, 뉴멕시코의 석탄기 유절류 ''Hibbertopterus''에 대한 연구는 일부 유절류의 서식지가 "재평가될 필요가 있을 수 있다"고 결론지었다.[3] 유일하게 살아남은 유절류 과인 Adelophthalmidae는 단일 속인 ''Adelophthalmus''로 대표되었다. 히베르토프테리드, 믹테로프테리드 및 ''Adelophthalmus''는 페름기까지 살아남았다.[20]
''Adelophthalmus''는 다른 유절류가 없는 상황에서 번성하며, 모든 후기 고생대 유절류 중에서 가장 흔해졌고, 생존하는 stylonurine보다 더 많은 수와 다양성을 보였다.[21] ''Adelophthalmus''에 속하는 33종 중 23종(69%)이 석탄기에서만 발견되었다.[22][23] 이 속은 후기 석탄기에 가장 높은 다양성을 보였다. 비록 ''Adelophthalmus''가 이미 후기 데본기에 비교적 널리 분포하고 있었고 모든 주요 육지를 대표했지만, 고생대 마지막 두 기간 동안 판게아가 전 세계적인 초대륙으로 합쳐지면서 ''Adelophthalmus''는 거의 전 세계적인 분포를 얻을 수 있었다.[24]
후기 석탄기와 초기 페름기 동안 ''Adelophthalmus''는 널리 분포했으며, 주로 해안 평원에 인접한 기수 및 담수 환경에서 서식했다. 이러한 환경은 유리한 기후 조건에 의해 유지되었다. 판게아의 형성에 따른 기후 변화가 전 세계의 퇴적 및 식생 패턴을 변화시키면서, 이들은 지속되지 못했다. 서식지가 사라지면서 ''Adelophthalmus''는 수가 줄어들었고, 이미 초기 페름기의 Leonardian 단계까지 멸종했다.[25]
믹테로프테리드와 히베르토프테리드는 일정 기간 생존을 지속했으며, 각각의 그룹에서 페름기 지층에서 알려진 속은 ''Hastimima''와 ''Campylocephalus''였다.[26] ''Hastimima''는 ''Adelophthalmus''와 마찬가지로 초기 페름기에 멸종했고,[27] ''Campylocephalus''는 더 오래 생존했다. 러시아의 페름기 퇴적층에서 발견된 거대한 불완전한 등딱지는 ''C. permianus'' 종의 유일한 화석 유물이며, 길이가 1.4m에 달했을 수 있다. 원래 후기 페름기 시대라고 여겨졌지만[28] 다른 자료에서는 이 표본에 대해 Kungurian-Roadian 시대(약 2억 8300만 년 전 ~ 2억 6700만 년 전)를 제시한다.[29] 유절류 큐티클의 조각인 ''Woodwardopterus freemanorum'' 종은 오스트레일리아의 페름기 말기에서 알려져 있으며, 이는 알려진 가장 어린 유절류를 나타낸다.[30] 페름기보다 높은 화석층에서는 유절류가 알려져 있지 않다. 이는 마지막 유절류가 종말적인 멸종 사건이 일어났을 때, 또는 그 직전에 죽었음을 나타낸다. 이 멸종 사건인 페름기-트라이아스기 대멸종은 기록된 가장 파괴적인 대멸종이며, 삼엽충과 같은 다른 성공적인 고생대 그룹을 멸종시켰다.[31]
참조
[1]
웹사이트
Selectivity in the evolution of Palaeozoic arthropod groups, with focus on mass extinctions and radiations: A phylogenetic approach
https://core.ac.uk/r[...]
[2]
논문
Phylogeny and palaeoecology of the Adelophthalmoidea (Arthropoda; Chelicerata; Eurypterida)
https://doi.org/10.1[...]
2008-01-01
[3]
논문
A new species of the eurypterid Hibbertopterus from the Carboniferous of New Mexico, and a review of the Hibbertopteridae
https://doi.org/10.1[...]
2023-02-01
[4]
웹사이트
Gigantostraca Haeckel 1866
http://www.insecta.b[...]
Nomina circumscribentia insectorum (World Wide Web electronic database)
2010-04-15
[5]
서적
海の中に棲む巨大なサソリ
東海大学出版会
2002
[6]
서적
ウミサソリ綱 Class Eurypteria
裳華房
2008
[7]
논문
The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa
https://bmcevolbiol.[...]
2015-09-01
[8]
논문
The last eurypterid – a southern high-latitude record of sweep-feeding sea scorpion from Australia constrains the timing of their extinction
https://doi.org/10.1[...]
2021-11-30
[9]
논문
Giant claw reveals the largest ever arthropod
https://www.ncbi.nlm[...]
2008-02-23
[10]
웹사이트
Eurypterus - definition, etymology and usage, examples and related words
http://www.finedicti[...]
2020-11-26
[11]
웹사이트
eurypterid {{!}} Origin and meaning of eurypterid by Online Etymology Dictionary
https://www.etymonli[...]
2020-11-26
[12]
웹사이트
Definition of EURYPTERIDA
https://www.merriam-[...]
2020-11-26
[13]
서적
動物分類名
丸善
1988
[14]
논문
Segmentation and tagmosis in Chelicerata
https://www.academia[...]
[15]
논문
Distribution and dispersal history of Eurypterida (Chelicerata)
https://www.scienced[...]
2007-09-03
[16]
논문
Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
http://rsbl.royalsoc[...]
2010-04-23
[17]
논문
Insights into the 400 million-year-old eyes of giant sea scorpions (Eurypterida) suggest the structure of Palaeozoic compound eyes
https://www.nature.c[...]
2019-11-28
[18]
논문
Morphology, taxonomy, and classification of the order Eurypterida, Burmeister, 1843
https://www.semantic[...]
1989
[19]
논문
Ecdysis in sea scorpions (Chelicerata: Eurypterida)
http://www.sciencedi[...]
2008-08-11
[20]
서적
The Eurypterida of New York
https://www.biodiver[...]
New York State Education Department,
1912
[21]
논문
The Eurypterid Endostoma and Its Homology with Other Chelicerate Structures
https://bioone.org/j[...]
2022-09
[22]
논문
Ex vivo three-dimensional reconstruction of Acutiramus : a giant pterygotid sea scorpion (American Museum novitates, no. 4004)
https://hdl.handle.n[...]
2023-11-20
[23]
논문
Autecology of Silurian eurypterids
https://www.research[...]
1984
[24]
논문
A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)
https://www.jstor.or[...]
1964
[25]
논문
Functional morphology of the prosoma of ''Baltoeurypterus tetragonophthalmus'' (Fischer) (Chelicerata: Eurypterida)
https://www.research[...]
1981
[26]
논문
The evolution of feeding within Euchelicerata: data from the fossil groups Eurypterida and Trigonotarbida illustrate possible evolutionary pathways
https://peerj.com/ar[...]
2020-08-13
[27]
논문
Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina)
https://doi.org/10.1[...]
2007-01-01
[28]
논문
The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida)
http://www.tandfonli[...]
2010-03-15
[29]
논문
A collection of eurypterids from the Silurian of Lesmahagow collected pre 1900
https://www.research[...]
[30]
논문
The origin of pterygotid eurypterids (Chelicerata: Eurypterida)
https://onlinelibrar[...]
2009
[31]
논문
The eurypterid Stoermeropterus conicus from the lower Silurian of the Pentland Hills, Scotland
https://www.research[...]
[32]
논문
Babes in the wood – a unique window into sea scorpion ontogeny
https://doi.org/10.1[...]
2013-05-10
[33]
논문
Selectivity in the evolution of Palaeozoic arthropod groups, with focus on mass extinctions and radiations: a phylogenetic approach
https://kuscholarwor[...]
2014-05-31
[34]
논문
The functional morphology of mating in the Silurian eurypterid,Baltoeurypterus tetragonophthalmus(Fischer, 1839)
http://www.sciencedi[...]
1997-08-01
[35]
간행물
Gill structure in the Lower Devonian eurypterid ''Tarsopterella scotica''
https://foreninger.u[...]
1975
[36]
논문
The respiratory organs of eurypterids
https://www.palass.o[...]
Palaeontology
1995
[37]
간행물
Lamellate book-gills in a late Ordovician eurypterid from the Soom Shale, South Africa: support for a eurypterid-scorpion clade
https://www.academia[...]
1999
[38]
간행물
XXIV.—The Anatomy and Relations of the Eurypteridæ
https://www.cambridg[...]
1895
[39]
간행물
On Gill-like Structures in the Eurypterida
https://www.cambridg[...]
1941-02
[40]
간행물
Eurypterid respiration
https://www.research[...]
1985-04-02
[41]
간행물
Discerning the diets of sweep-feeding eurypterids: assessing the importance of prey size to survivorship across the Late Devonian mass extinction in a phylogenetic context
https://www.cambridg[...]
[42]
간행물
What big eyes you have: the ecological role of giant pterygotid eurypterids
https://royalsociety[...]
2014-07-31
[43]
간행물
All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids
https://www.ncbi.nlm[...]
2015-08
[44]
간행물
Ichnology of the Devonian (Emsian) Campbellton Formation, New Brunswick, Canada
https://journals.lib[...]
2017-02-08
[45]
간행물
Palaeontology: How did eurypterids swim?
https://www.nature.c[...]
1986-04
[46]
간행물
New trace fossil evidence for eurypterid swimming behaviour
https://onlinelibrar[...]
[47]
간행물
Lift based mechanisms for swimming in eurypterids and portunid crabs
https://www.cambridg[...]
1985
[48]
간행물
Distribution and dispersal history of Eurypterida (Chelicerata)
https://www.scienced[...]
2007-09-03
[49]
간행물
A gigantic fossil arthropod trackway
http://www.nature.co[...]
2005-12
[50]
간행물
Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion
https://www.cell.com[...]
2020-09-10
[51]
간행물
The functional morphology of mating in the Silurian eurypterid, Baltoeurypterus tetragonophthalmus (Fischer, 1839)
http://www.academia.[...]
[52]
간행물
Sperm carriers in Silurian sea scorpions
https://www.academia[...]
2011-10
[53]
서적
The Eurypterida of New York
https://en.wikisourc[...]
[54]
서적
A monograph of the British fossil Crustacea, belonging to the order Merostomata.
http://www.biodivers[...]
Printed for the Palæontographical Society,
1866
[55]
웹사이트
Die Entwicklungsgeschichte der Scolopender
https://www.schweize[...]
2020-11-26
[56]
간행물
Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata
https://onlinelibrar[...]
1979
[57]
간행물
Evolutionary Morphology and Phylogeny of Araghnida
https://onlinelibrar[...]
1990
[58]
간행물
A phylogenetic analysis of the arachnid orders based on morphological characters
https://academic.oup[...]
2007-06-01
[59]
간행물
Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura
https://academic.oup[...]
2013-01-01
[60]
간행물
A diverse chasmataspidid (Arthropoda: Chelicerata) fauna from the Early Devonian (Lochkovian) of Siberia
https://onlinelibrar[...]
2014
[61]
간행물
An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China
https://onlinelibrar[...]
2015
[62]
간행물
A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids
https://www.research[...]
2015-10
[63]
간행물
Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation
https://www.research[...]
2016-03
[64]
간행물
The origins of tetrapulmonate book lungs and their significance for chelicerate phylogeny
https://www.academia[...]
[65]
웹사이트
Current views on chelicerate phylogeny—A tribute to Peter Weygoldt
https://www.semantic[...]
2018-11-09
[66]
간행물
Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks
http://link.springer[...]
2019-03
[67]
간행물
Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders
https://peerj.com/ar[...]
2014-11-13
[68]
논문
A summary list of fossil spiders and their relatives
https://wsc.nmbe.ch/[...]
World Spider Catalog. Natural History Museum Bern
2020
[69]
간행물
From success to persistence: Identifying an evolutionary regime shift in the diverse Paleozoic aquatic arthropod group Eurypterida, driven by the Devonian biotic crisis: CHANGING EVOLUTIONARY REGIMES DURING THE DEVONIAN
https://www.research[...]
2017-01
[70]
서적
ぞわぞわした生きものたち: 古生代の巨大節足動物
https://www.worldcat[...]
ソフトバンククリエイティブ
2012-03
[71]
논문
A new genus of eurypterid (Chelicerata, Eurypterida) from the Upper Devonian salt deposits of Belarus
https://www.cambridg[...]
2018-09
[72]
서적
大昔の生きもの
https://www.worldcat[...]
ポプラ社
2014
[73]
논문
A new Ordovician eurypterid (Arthropoda: Chelicerata) from southeast Turkey: Evidence for a cryptic Ordovician record of Eurypterida
https://www.research[...]
2013-01
[74]
서적
とても巨大な絶滅せいぶつ図鑑
https://www.worldcat[...]
世界文化社
2019
[75]
뉴스
Giant claw points to monster sea scorpion - life - 21 November 2007 - New Scientist
http://www.newscient[...]
[76]
논문
The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa
https://bmcevolbiol.[...]
2015-12
[77]
논문
The last eurypterid – a southern high-latitude record of sweep-feeding sea scorpion from Australia constrains the timing of their extinction
https://www.tandfonl[...]
2021
[78]
논문
The functional morphology of mating in the Silurian eurypterid, Baltoeurypterus tetragonophthalmus (Fischer, 1839)
https://academic.oup[...]
1997-08
[79]
논문
Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura: Re-evaluating the Monophyly of Xiphosura
https://academic.oup[...]
2013-01
[80]
논문
Segmentation and tagmosis in Chelicerata
https://linkinghub.e[...]
2017-05
[81]
논문
Sperm carriers in Silurian sea scorpions
http://link.springer[...]
2011-10
[82]
논문
The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida)
https://doi.org/10.1[...]
2010-03-15
[83]
논문
Autecology of Silurian eurypterids
2018
[84]
논문
The trail of a large Silurian eurypterid
1975
[85]
논문
Problems of functional morphology and classification in stylonuroid eurypterids (Chelicerata, Merostomata), with observations on the Scottish Silurian Stylonuroidea*
https://www.cambridg[...]
1979
[86]
논문
A gigantic fossil arthropod trackway
https://www.nature.c[...]
2005-12
[87]
논문
The functional morphology of mating in the Silurian eurypterid, Baltoeurypterus tetragonophthalmus (Fischer, 1839)
https://academic.oup[...]
1997-08
[88]
논문
Babes in the wood – a unique window into sea scorpion ontogeny
http://bmcevolbiol.b[...]
2013
[89]
논문
All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids
https://royalsociety[...]
2015-08
[90]
논문
Cyrtoctenus wittebergensis sp. nov. (Chelicerata: Eurypterida), a large sweep-feeder from the Carboniferous of South Africa
https://www.cambridg[...]
1985
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com