코리토사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
코리토사우루스는 백악기 후기에 북아메리카에 서식했던 조각류 공룡의 한 속이다. 헬멧 모양의 볏을 가진 것이 특징이며, 코린트 투구를 뜻하는 그리스어에서 이름이 유래되었다. 1911년 캐나다 앨버타에서 화석이 발견되었으며, 현재까지 두 종이 인정된다. 볏의 기능은 소리 증폭, 시각적 과시, 종 내 의사소통 등 다양하게 추정된다. 초식 공룡으로, 숲이나 습지에서 살았을 것으로 보인다.
더 읽어볼만한 페이지
- 1914년 기재된 화석 분류군 - 카스모사우루스
카스모사우루스는 백악기 후기 북아메리카에 살았던 각룡류 공룡 속으로, 길고 넓은 프릴과 짧은 뿔이 특징이며 카스모사우루스 벨리와 카스모사우루스 루셀리 두 종이 유효하다. - 1914년 기재된 화석 분류군 - 고르고사우루스
고르고사우루스는 백악기 후기 북미에 살았던 몸길이 8~9m의 티라노사우루스과 육식 공룡으로, 알베르토사우루스와 가까운 친척이며 성장 계열이 잘 알려져 있고, 성장 단계에 따라 식단이 변화했다. - 바넘 브라운이 명명한 분류군 - 드로마에오사우루스
후기 백악기 캄파니안절에 살았던 드로마에오사우루스는 "달리는 도마뱀"이라는 뜻의 이름을 가진, 2m 정도 크기의 육식공룡으로 날카로운 이빨과 낫 모양 발톱, 강력한 턱힘을 이용해 사냥했을 것으로 추정되지만, 드로마에오사우루스과에 속하는 것 외에 정확한 분류는 아직 연구 중이다. - 바넘 브라운이 명명한 분류군 - 파키케팔로사우루스
파키케팔로사우루스는 백악기 후기에 북아메리카와 아시아에서 서식했던 조반류 공룡으로, 두꺼운 뼈로 된 돔 형태의 두개골을 가지고 이족 보행을 했을 것으로 추정되며, 종내 경쟁이나 과시용으로 두개골 돔을 사용했을 가능성이 있다. - 하드로사우루스과 - 에드몬토사우루스
에드몬토사우루스는 백악기 후기에 살았던 하드로사우루스과의 대형 초식 공룡으로, 9~12m의 몸길이, 오리 부리 형태의 입, 그리고 피부 자국이 보존된 미라 화석으로 알려져 있으며, *E. regalis*와 *E. annectens* 두 종이 인정되고, 티라노사우루스와 공존했던 생태 환경과 식생, 성장 과정 등에 대한 연구가 활발히 진행되고 있다. - 하드로사우루스과 - 마이아사우라
마이아사우라는 백악기 후기에 북아메리카에서 서식한 8~9m 길이의 하드로사우루스과 초식 공룡으로, 눈 사이의 짧고 단단한 볏과 무리 생활 및 둥지 군집을 통해 새끼를 키운 흔적이 발견된 "좋은 어머니"라는 의미의 속명이 특징이다.
| 코리토사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Corythosaurus |
| 명명자 | Brown, 1914 |
| 어원 | 투구 도마뱀 |
| 몸길이 | 7.7–9 m |
| 두개골 높이 | 70.8 cm |
| 생물학적 분류 | |
| 속 | 코리토사우루스속 |
| 종 | C. 카수아리우스 Brown, 1914 (모식종) C. 인테르메디우스 Parks, 1923 (원래 스테파노사우루스) |
| 동의어 | 코리토사우루스 엑스카바투스 Gilmore, 1923 스테파노사우루스 인테르메디우스 Parks, 1923 프로케네오사우루스 에렉토프론스 (Parks, 1931 [원래 테트라고노사우루스]) 테트라고노사우루스 에렉토프론스 Parks, 1931 코리토사우루스 비크리스타투스 Parks, 1935 코리토사우루스 브레비크리스타투스 Parks, 1935 |
| 시대 | |
| 생존 시기 | 백악기 후기 (캄파니아절), 약 7750만년 전 ~ 7480만년 전 |
| 식성 | |
| 식단 | 식물 (과일, 침엽수) |
2. 발견 및 명명

코리토사우루스의 첫 번째 표본인 AMNH 5240은 1911년 바넘 브라운이 캐나다 앨버타의 레드디어 강 유역에서 발견하여 1912년 가을에 확보했다.[3][24] 이 표본은 앨버타의 벨리 리버 그룹 지층에서 나왔으며,[24] 거의 완전한 골격과 함께 상당 부분의 피부 자국이 보존되어 있어 주목받았다.[25] 골격은 대부분 연결된 상태였으나, 꼬리 끝 약 0.61m와 앞다리 일부가 없었다.[25] 피부 자국은 골격의 넓은 부분을 덮고 있어 생존 당시의 몸 형태를 추정하는 데 도움을 주었다.[25] 1914년, 브라운과 피터 카이젠은 두 번째 표본인 AMNH 5338을 발견했다. 이 두 표본은 현재 미국 자연사 박물관에 원래 발굴된 자세 그대로 전시되어 있다.[26]
모식종인 ''코리토사우루스 카수아리우스'' (''Corythosaurus casuarius'')는 브라운이 1912년에 확보한 첫 번째 표본(AMNH 5240)을 모식 표본으로 하여 1914년에 명명되었다.[4] 속명 ''Corythosaurus''는 머리에 있는 볏의 모양 때문에 붙여졌으며, 종명 ''casuarius''는 화식조와의 유사성 때문에 붙여졌다.[4]
한편, 1912년 찰스 H. 스턴버그가 발견했던 코리토사우루스의 가장 잘 보존된 표본 두 점은 비극적인 운명을 맞았다. 제1차 세계 대전 중이던 1916년 12월 6일, 이 표본들을 영국의 런던 자연사 박물관으로 운송하던 배 ''SS 마운트 템플''호가 대서양 한가운데에서 독일의 상선 습격선 SMS ''뫼베''에 의해 격침되면서 함께 바다에 가라앉고 말았다.[5]
과거에는 여러 종의 코리토사우루스가 보고되기도 했으나, 1975년 피터 도드슨의 연구 이후 현재는 대부분 ''C. casuarius'' 한 종만이 유효한 것으로 간주된다.[27] 일부 다른 종들이 제안되기도 했지만, 이들의 유효성에 대해서는 논란이 있다.[6][7][28][8][44]
2. 1. 종 목록
모식종인 ''코리토사우루스 카수아리우스'' (''Corythosaurus casuarius'')는 1914년 바넘 브라운에 의해 명명되었다. 이는 그가 1911년 앨버타의 레드 디어 강 근처 벨리 리버 그룹에서 발견하여 1912년에 확보한 첫 번째 표본인 AMNH 5240을 모식 표본으로 한다.[3][24][25] 1916년 브라운은 1914년에 브라운과 피터 카이젠이 발견한 또 다른 표본인 AMNH 5338을 기반으로 더 상세한 설명을 발표했으며, 이는 부모식 표본이 되었다.[26] 속명 ''Corythosaurus''는 그리스어 κόρυθος|코리토스grc("코린트 투구")에서 유래했으며, "투구를 쓴 도마뱀"이라는 뜻이다. 종명 ''casuarius''는 머리볏 모양이 유사한 화식조(''Casuarius'')를 가리킨다. 따라서 전체 이명법은 "코린트 투구 볏을 가진 화식조 같은 파충류"를 의미한다.[4]
과거에는 최대 7종의 코리토사우루스가 기술되었으나, 1975년 피터 도슨은 여러 람베오사우루스아과 공룡들의 두개골과 볏 형태 차이를 연구한 결과, 이러한 차이가 동물의 성별 및 나이에 따른 개체 변이일 수 있다고 주장했다. 이 연구 이후 ''C. casuarius'' 한 종만이 유효한 것으로 인정받고 있다.[27]
그러나 ''C. intermedius''는 일부 연구에서 여전히 유효한 종으로 간주되기도 한다. 이 종은 1920년 레비 스턴버그가 발견한 두개골(표본 ROM 776)을 기반으로 1923년 윌리엄 파크스가 명명했다. 파크스는 원래 같은 해 초에 이 표본을 ''스테파노사우루스 인테르메디우스'' (''Stephanosaurus intermedius'')로 명명했었다.[6] 종명 ''intermedius''는 파크스가 이 종이 다른 종들의 중간적인 특징을 가진다고 보았기 때문에 붙여졌다.[7][28][8] ''C. intermedius''는 ''C. casuarius''보다 지질학적으로 약간 더 후대인 캄파니아 시기 후기에 살았던 것으로 보이며, 두 종이 동일하지 않다는 주장을 뒷받침하는 2009년 연구도 있다.[44]
| 종명 | 명명자 (연도) | 기반 표본 (알려진 경우) | 비고 |
|---|---|---|---|
| C. bicristatus | Parks (1935) | 현재 C. casuarius의 동의어로 간주됨 | |
| C. brevicristatus | Parks (1935) | 현재 C. casuarius의 동의어로 간주됨 | |
| C. excavatus | Gilmore (1923) | UALVP 13 | 1920년 발견된 두개골 기반, 현재 C. casuarius의 동의어로 간주됨[9] |
| C. frontalis | Parks (1935) | 현재 C. casuarius의 동의어로 간주됨 | |
| C. intermedius | Parks (1923) | ROM 776 | 일부 연구에서 유효종으로 간주됨[6][7][28][8][44] |
몸 전체 길이는 약 10m에서 13m, 전체 높이는 약 5m, 무게는 약 4ton에서 5ton으로 추정된다. 파라사우롤로푸스(''Parasaurolophus'')나 람베오사우루스(''Lambeosaurus'')처럼 머리 부분에 속이 비어 있는 돌기를 가지고 있다. 코리토사우루스의 경우, 이름의 유래가 된 반원형의 돌기가 머리 꼭대기에 솟아 있다. 이 돌기는 코뼈가 확장된 것으로 복잡한 내부 구조를 가지며, 성별에 따라 크기가 달라 수컷의 것이 더 컸을 것으로 추정된다. 이 돌기는 후각을 발달시키거나 울음소리를 증폭하는 데 사용되었을 것으로 생각되며, 모형 실험에서는 안개 경적과 비슷한 소리를 냈다고 한다. 달리는 속도는 시속 48km 정도로 추정된다.
3. 형태
머리를 지탱하는 목은 S자 모양이었으며, 척추의 극돌기가 길게 뻗어 등 쪽에 능선을 형성했다. 꼬리뼈는 등뼈에서 이어지는 골질 결합 조직으로 단단히 연결되어 있어 움직임이 제한적이었을 것으로 보인다.
3. 1. 골격

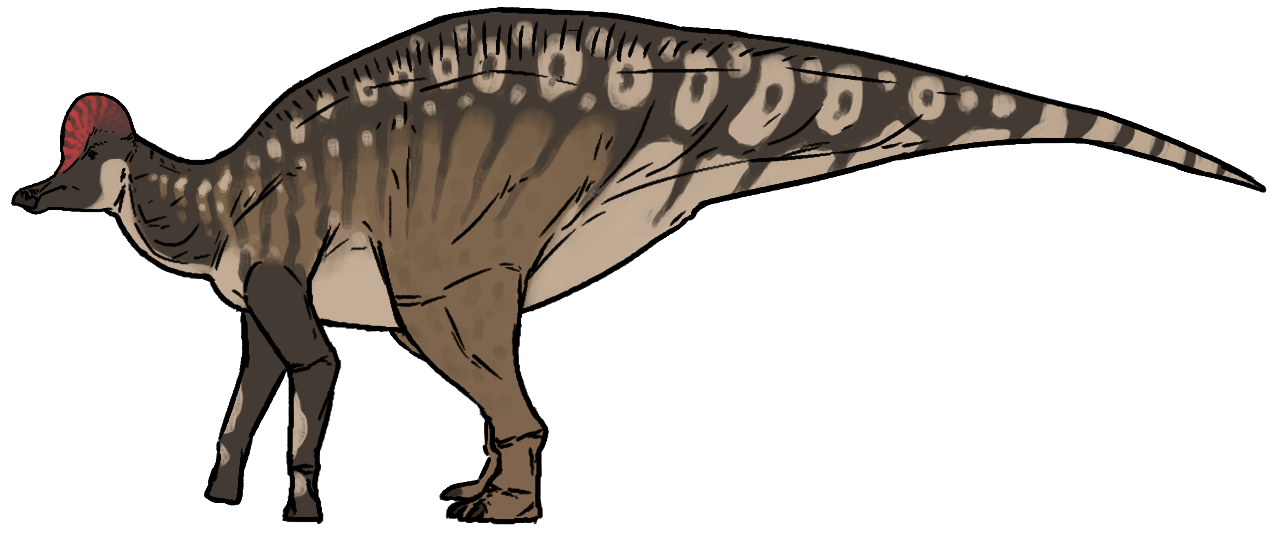
코리토사우루스의 몸 전체 크기는 약 10m에서 13m 정도로 추정된다. 2012년 벤슨(Benson) 등은 평균 길이를 9m로 추정했다.[32] 1962년 에드윈 H. 콜버트는 모형을 이용해 무게를 추정했는데, 평균 무게는 약 3.82ton로 결론 내렸다.[10] 미국 자연사 박물관(AMNH)의 표본 AMNH 5240은 총 길이가 8.1m이고 무게는 3.1ton에 가까웠다.[11] 2016년 그레고리 S. 폴은 ''C. casuarius'' 종의 길이를 8m, 무게를 2.8MT로, ''C. intermedius'' 종의 길이를 7.7m, 무게를 2.5MT로 추정했다.[12] 성체 크기로 보이는 ''C. casuarius'' 표본은 길이가 약 9m로 측정되었다.[13]
파라사우롤로푸스(''Parasaurolophus'')나 람베오사우루스(''Lambeosaurus'')처럼 머리 부분에 속이 비어 있는 볏 모양의 돌기를 가지고 있다. 이 볏은 화식조의 것이나 고대 그리스의 고린도 투구를 닮았으며,[32] 성별에 따라 크기 차이가 있어 수컷의 것이 더 컸을 것으로 추정된다.[32][40] 두개골은 에드몬토사우루스(''Edmontosaurus'')나 크리토사우루스(''Kritosaurus''), 사우롤로푸스(''Saurolophus'')보다 비례적으로 짧고 작지만, 볏을 포함하면 표면적은 거의 비슷하다.[29]
이 공룡의 두개골은 20개 이상 발견되었다.[40] 볏은 앞위턱뼈, 비골, 앞이마뼈, 이마뼈가 합쳐져 형성되었으며,[29] 내부에는 길쭉한 콧구멍 통로가 복잡하게 얽혀 있다. 콧구멍은 볏 안으로 들어가 측면의 주머니를 거쳐 중앙의 방으로 이어지고 호흡기로 연결된다.[32][40] 이 구조는 후각을 발달시키거나 울음소리를 증폭하는 데 쓰였을 것으로 생각된다. 모형 실험에서는 안개 경적과 비슷한 소리를 냈다고 한다. 볏의 두 반쪽은 정중 봉합선으로 나뉘며, 눈구멍 앞쪽의 볏은 두꺼운 뼈로 이루어져 있다.[29]
초기 연구에서 바넘 브라운은 볏의 구조를 일부 잘못 해석했다. 그는 비골이 볏의 대부분을 차지하고 앞위턱뼈에 의해 분리되지 않는다고 생각했으나, 실제로는 주둥이 대부분과 볏 앞부분은 앞위턱뼈로 이루어져 있으며 이것이 비골을 분리한다. 또한 브라운은 볏 위쪽과 뒤쪽이 이마뼈로 덮여 있다고 생각했지만, 이는 실제로는 비골이었다. 실제 이마뼈는 볏 기저부의 내부 구조를 이루며 측면에서는 보이지 않는다.[14][29]
코리토사우루스의 입은 좁으며, 앞위턱뼈는 콧구멍을 주둥이 앞쪽까지 연장하는 공기 통로를 감싼다. 이 통로는 '가짜 코'라고 불리는 뼈로 된 콧구멍에서 끝나는데, 실제 콧구멍은 눈구멍 위, 볏 안쪽에 위치한다.[29] 모식 표본의 아래턱 길이는 66.9cm, 깊이는 10cm이다. 부리부터 볏 꼭대기까지의 전체 길이는 83.7cm, 두개골 길이는 81.2cm, 높이는 70.8cm이다.[33]
머리를 지탱하는 목은 S자 모양이다. 척추는 극돌기가 길게 뻗어 등쪽에 능선을 형성했다. 바넘 브라운에 따르면 척추는 15개의 목뼈, 19개의 등뼈, 8개의 천추, 61개 이상의 꼬리뼈로 구성된다.[30] 꼬리뼈는 등뼈에서 이어지는 골질 결합 조직으로 단단히 연결되어 있어 움직임이 제한적이었을 것으로 보인다.
3. 2. 연조직

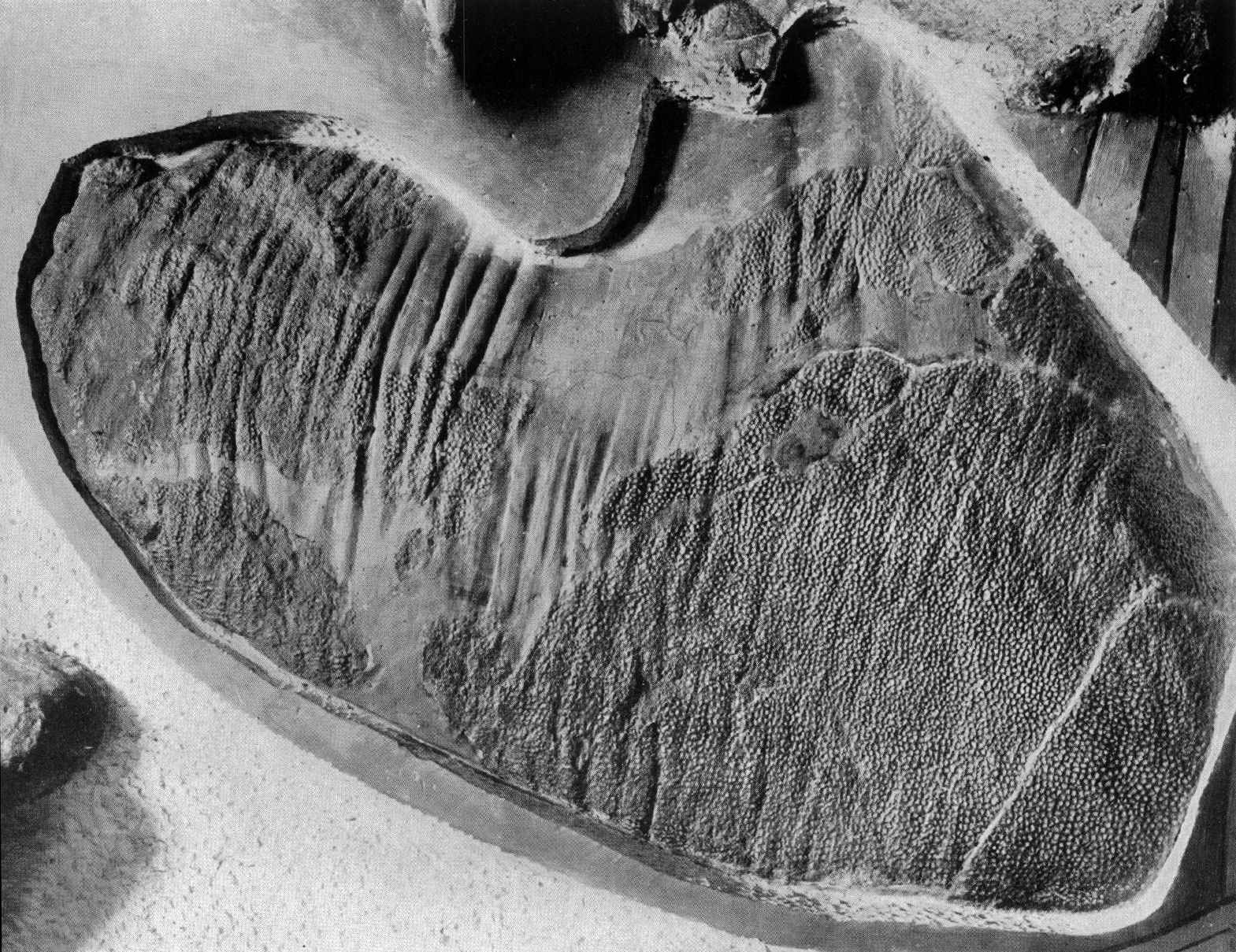
''코리토사우루스 카수아리우스''(''C. casuarius'')의 모식 표본에서는 옆구리와 꼬리가 여러 종류의 비늘로 덮여 있었던 흔적이 발견되었다. 몸 전체에는 작은 돌기로 덮인 다각형의 결절 비늘이 있었으며, 그 크기는 부위마다 다양했다. 원뿔형이면서 삿갓조개 모양을 한 비늘은 경골(정강이뼈) 뒤쪽 피부 주름에서만 보존되었는데, 이는 다리 피부가 아니라 배 아랫부분의 피부였을 가능성이 있다.[33] ''C. casuarius''의 다각형 비늘 사이에는 방패 모양의 비늘이 줄지어 촘촘하게 배열되어 있었다.[15] 목뼈 부분을 제외한 모든 척추뼈에는 골화된 힘줄이 존재했다. 이 힘줄은 척추뼈의 횡돌기 아래로는 뻗어 있지 않았다. 각 힘줄은 시작 부분(기부)에서 납작하고, 중앙 부분은 가로로 타원형이며, 끝부분은 둥근 형태를 띠었다.[31]
''코리토사우루스 카수아리우스'' 외에도 여러 하드로사우루스과(Hadrosauridae) 공룡들에게서 광범위한 피부 자국이 발견되었다. 특히 에드몬토사우루스 아넥텐스(''Edmontosaurus annectens'')에서 주목할 만한 외피가 발견되었으며, 브라킬로포사우루스 카나덴시스(''Brachylophosaurus canadensis''), 그리포사우루스 노타빌리스(''Gryposaurus notabilis''), 파라사우롤로푸스 워커리(''Parasaurolophus walkeri''), 람베오사우루스 마그니크리스타투스(''Lambeosaurus magnicristatus''), ''L. lambei'', 사우롤로푸스 안구스티로스트리스(''Saurolophus angustirostris'') 및 아직 종이 확인되지 않은 조각류 공룡에서도 피부 흔적이 발견되었다. 이 중 ''L. lambei'', ''C. casuarius'', ''G. notabilis'', ''P. walkeri'', ''S. angustirostris''는 다각형 비늘을 가지고 있었다. ''L. lambei'', ''S. angustirostris'', ''C. casuarius''의 비늘은 서로 유사한 형태를 보였다. 코리토사우루스는 뒷다리와 발의 피부 자국이 보존된 몇 안 되는 하드로사우루스과 공룡 중 하나이다. 2013년 연구에 따르면, 하드로사우루스과 중에서는 사우롤로푸스 안구스티로스트리스(''Saurolophus angustirostris'')가 가장 상태가 좋고 완전한 발과 다리 외피를 보존하고 있었지만, 사우롤로푸스 오스본니(''S. osborni''), 에드몬토사우루스 아넥텐스(''Edmontosaurus annectens''), 그리고 ''람베오사우루스 람베이''(''L. lambei'', ''L. clavinitialis''와 동일 종)와 같은 다른 종들도 해당 부위에 상당량의 연조직이 보존되어 있다.[15]
과거에는 코리토사우루스의 손과 발에 물갈퀴가 있는 것처럼 보여 주로 물속에서 생활했을 것이라고 생각되었다.[36] 그러나 이후 연구를 통해 이것이 실제 물갈퀴가 아니라, 오늘날 많은 포유류에서 볼 수 있는 것과 매우 유사한 얇은 발바닥 패드(pad)임이 밝혀졌다.[32][42]
4. 분류

처음에 바넘 브라운(Barnum Brown)은 코리토사우루스(''Corythosaurus'')를 당시 트라코돈과(Trachodontidae)로 불리던 분류군[24](현재는 하드로사우루스과(Hadrosauridae)로 알려짐[32])의 일원으로 분류했다. 트라코돈과 내에는 트라코돈아과(Trachodontinae)와 사우롤로푸스아과(Saurolophinae)가 있었는데, 브라운은 트라코돈아과에 ''하드로사우루스(Hadrosaurus)'', ''트라코돈(Trachodon)'', ''클라오사우루스(Claosaurus)'', ''크리토사우루스(Kritosaurus)''를 포함시켰고,[38] 사우롤로푸스아과에는 ''코리토사우루스(Corythosaurus)'', ''스테파노사우루스(Stephanosaurus)'', ''사우롤로푸스(Saurolophus)''를 포함시켰다.[39]
이후 브라운은 코리토사우루스의 계통발생학적 위치를 수정하여, 이 공룡이 ''하이파크로사우루스(Hypacrosaurus)''와 매우 밀접한 관련이 있으며, 어쩌면 그 조상일 수도 있다고 보았다. 그가 두 속 사이에서 발견한 유일한 차이점은 척추의 발달 정도와 다리 비율이었다.[30] 1920년대에 알프레드 셔우드 로머(Alfred Sherwood Romer)는 공룡 골반뼈 연구를 통해 두 주요 공룡 목(目)이 별도로 진화했을 가능성과, 새가 골반뼈 모양과 비율을 근거로 조반류에서 진화했을 가능성을 제기했다. 그는 ''티라노사우루스(Tyrannosaurus)''와 ''코리토사우루스(Corythosaurus)''를 기본 모델로 사용하여 두 가설을 검토했다. 로머는 비록 새가 현재 수각류로 분류되지만, 만약 ''코리토사우루스''와 같은 조반류에서 진화했다면 새 특유의 골반 근육 구조와 해부학적 특징이 나타나는 것이 충분히 가능하다고 보았다.[16] 그러나 현재는 ''코리토사우루스''를 포함한 조반류의 골반 구조가 새와 표면적으로 유사해 보일지라도, 새는 고도로 분화된 마니랍토라 수각류 공룡의 후손으로 여겨진다.[32]
현재 ''코리토사우루스''는 하드로사우루스과의 하위 분류군인 람베오사우루스아과(Lambeosaurinae)로 분류된다. 이 아과에는 ''하이파크로사우루스(Hypacrosaurus)'', ''람베오사우루스(Lambeosaurus)'', ''올로로티탄(Olorotitan)'' 등 다른 하드로사우루스류 공룡들이 포함된다. 이들은 ''올로로티탄''을 제외하고는 대체로 비슷한 모양의 두개골과 볏을 가지고 있다. 하지만 2003년에 발표된 연구에서는 독특한 볏 모양에도 불구하고 ''올로로티탄''이 ''코리토사우루스''의 가장 가까운 친척일 수 있다고 제안했다.[37] 벤슨(Benson) 등(2012)의 연구에서는 ''코리토사우루스''가 부채 모양의 볏을 가진 람베오사우루스류 그룹에 속하며, ''벨라프론스(Velafrons)'', ''니포노사우루스(Nipponosaurus)'', ''하이파크로사우루스(Hypacrosaurus)''와 밀접한 관련이 있다고 분석했다.[32]
2014년, ''장헝롱(Zhanghenglong)''이라는 새로운 공룡을 기술한 연구가 학술지 ''PLOS ONE''에 게재되었다. 이 연구에는 하드로사우루스상과(Hadrosauroidea) 공룡들의 관계를 보여주는 거의 완전한 분기도(cladogram)가 포함되었는데, 여기서 ''코리토사우루스''는 가장 진화된 람베오사우루스류 중 하나이자 ''하이파크로사우루스''의 자매 분류군으로 나타났다.[17]
5. 고생물학
''코리토사우루스''와 현생 파충류의 공막륜을 비교한 결과, 이 공룡은 짧은 간격으로 하루 종일 활동하는 경일주성 생활을 했을 가능성이 제기된다.[42] 또한 ''로포로톤''과 같은 하드로사우루스과 공룡들처럼, 코리토사우루스 역시 길어진 라게나 덕분에 청각이 매우 발달했을 것으로 보인다.[18] 파충류의 막대 모양 귀뼈인 등골이 얇고 고막이 크다는 점은 예민한 중이를 가졌음을 시사한다.[42] 이러한 해부학적 특징을 바탕으로, 하드로사우루스류의 귀는 현생 악어만큼 소리를 잘 감지할 수 있을 정도로 민감했을 것으로 추정된다.[18]
5. 1. 볏의 기능
파라사우롤로푸스(''Parasaurolophus'')나 람베오사우루스(''Lambeosaurus'')처럼 코리토사우루스도 머리 부분에 속이 빈 골질 돌기, 즉 볏을 가지고 있다. 이 볏은 코뼈가 확장된 것으로, 겉보기와 달리 내부 구조는 상당히 복잡하다. 볏의 크기는 성별에 따라 차이가 있어 수컷의 것이 더 컸을 것으로 추정된다.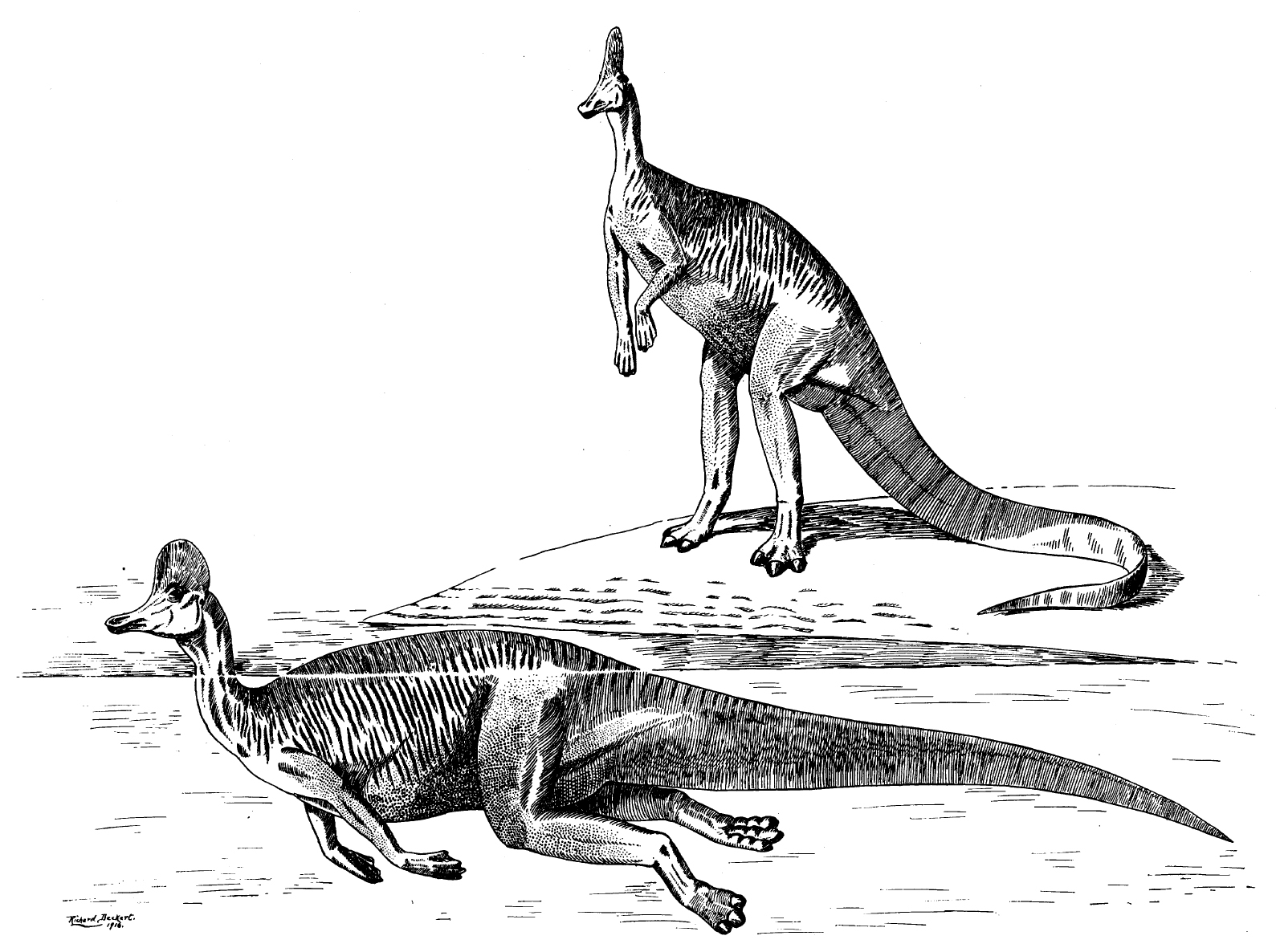
코리토사우루스 볏의 복잡한 내부 구조는 소리를 내는 데 사용되었을 것으로 보인다. 코리토사우루스를 포함한 람베오사우루스아과(Lambeosaurinae) 공룡들의 콧구멍 통로는 복잡한 형태를 띠는데, 코리토사우루스, 히파크로사우루스, 람베오사우루스는 S자 형태이고 파라사우롤로푸스는 U자 형태의 관을 가진다.[18] 어떤 소리든 이 정교한 공간을 통과하면서 증폭되었을 것이다.[32][40] 과학자들은 코리토사우루스가 트롬본과 같은 금관 악기와 유사하게 크고 낮은 음의 울음소리를 낼 수 있었을 것으로 추측한다.[40][19] 모형 실험에서는 안개 경적과 비슷한 소리가 재현되기도 했다.
이러한 소리는 동족에게 먹이가 있는 곳을 알리거나 포식자의 접근과 같은 잠재적 위협을 경고하는 등 의사소통 수단으로 사용되었을 수 있다.[40] 또한 짝을 유인하는 데에도 사용되었을 가능성이 있다. 볏의 형태는 종마다 다르기 때문에, 각기 다른 소리를 내어 종을 구별하는 데 쓰였을 수도 있다.[19] 코리토사우루스를 포함한 람베오사우루스아과 공룡들이 내는 소리는 주로 저주파(400 Hz 미만)였을 것으로 추정된다. 저주파 소리는 주변 환경의 영향을 덜 받고 더 먼 거리까지 퍼져나가는 장점이 있다.[18] 이 외에도 볏이 후각 발달에 기여했을 가능성도 제기된다.
과거에는 볏이 있는 하드로사우루스류가 물속에서 생활하는 반수생 동물로 여겨지기도 했다.[19] 이는 발에서 발견된 피부 흔적을 물갈퀴로 잘못 해석했기 때문이다.[32][36] 당시 이론은 이들이 물속 깊이 잠수할 때 볏을 스노클처럼 사용하여 호흡하거나 공기를 저장했을 것이라는 내용이었다. 그러나 볏 끝에는 외부로 통하는 구멍이 없으며, 불과 3m 깊이의 물속에서도 수압 때문에 폐로 공기를 보낼 수 없다는 사실이 밝혀지면서 이 가설은 현재 받아들여지지 않는다.[19]
5. 2. 성장

코리토사우루스 카수아리우스(''Corythosaurus casuarius'')는 람베오사우루스류 중에서 어린 개체의 화석이 발견된 몇 안 되는 종 중 하나이다. 람베오사우루스 람베이(''Lambeosaurus lambei''), 하이파크로사우루스 스테빈게리(''Hypacrosaurus stebingeri''), 하이파크로사우루스 알티스피누스(''H. altispinus'')와 함께 어린 개체 화석이 발견되었다. 어린 개체는 성체의 특징인 큰 볏이 아직 발달하지 않았기 때문에 종을 특정하기가 더 어렵다. 람베오사우루스류의 볏은 나이가 들면서 점차 성장하여 성체가 되면 뚜렷해지는 경향이 있다.
공룡 공원 지층에서는 다양한 속에서 유래한 50개 이상의 연결된 표본이 발견되었는데, 이 중 어린 개체는 종 수준에서 식별하기 어려웠다. 과거 고생물학자들은 크기와 볏 모양의 차이를 기준으로 분류군을 구별하여, 이 지층 지역에서 4개의 속과 13개의 종을 식별하기도 했다. 가장 작은 표본들은 테트라고노사우루스(현재는 프로케네오사우루스의 동의어로 간주됨)로 분류되었고, 더 큰 골격은 코리토사우루스나 람베오사우루스로, 심지어 일부 성체는 파라사우롤로푸스로 식별되기도 했다.[41] 호스슈 캐니언 지층에서 발견된 작은 람베오사우루스류는 체네오사우루스로 분류되었다.[41]
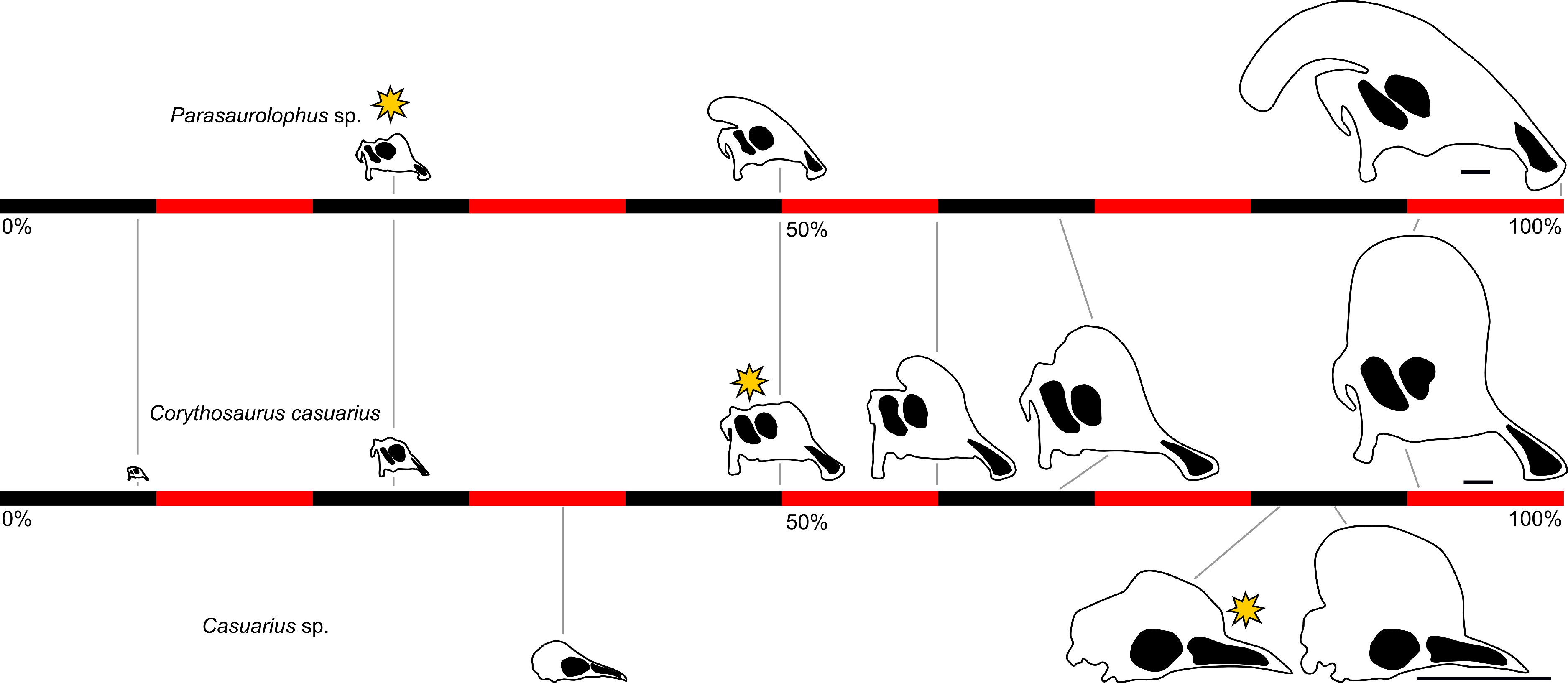
코리토사우루스는 성체 크기의 절반 정도가 되었을 때 볏이 발달하기 시작했다. 이는 성체 크기의 25% 정도일 때 볏이 자라기 시작하는 파라사우롤로푸스의 어린 개체와 비교된다. 어린 코리토사우루스는 성체와 마찬가지로 전상악골-비골 사이에 샘(gland) 구조를 가지고 있었다. 또한, 어린 코리토사우루스와 성체의 두개골 봉합선은 매끄럽고 직선이 아닌 물결 모양인데, 이는 람베오사우루스나 하이파크로사우루스와 유사하며 파라사우롤로푸스와는 다른 특징이다. 이 특징은 파라사우롤로푸스족(Parasaurolophini)과 람베오사우루스족(Lambeosaurini)을 구별하는 데 도움이 된다. 일반적으로 코리토사우루스, 람베오사우루스, 하이파크로사우루스 스테빈게리와 같은 람베오사우루스류의 어린 개체, 파라사우롤로푸스와 같은 파라사우롤로푸스족, 그리고 카자크람비아(''Kazaklambia'')와 같은 원시적인 람베오사우루스류의 볏은 서로 다른 특징으로 구별될 수 있지만, 상당히 유사한 점도 많다.[20]
1975년 피터 도드슨(Peter Dodson)의 연구는 앨버타 지역에 존재한다고 여겨졌던 람베오사우루스류의 종 수가 실제로는 훨씬 적다는 것을 밝혀냈다.[27][41] 이 연구를 통해 테트라고노사우루스는 코리토사우루스 또는 람베오사우루스의 어린 개체임이 드러났다. 특히 '테트라고노사우루스 에렉토프론스'(''T. erectofrons'')는 주로 생물 측정학적 정보를 바탕으로 코리토사우루스의 어린 개체로 분류되었다. 다만, 'T. 에렉토프론스'로 분류되었던 표본 중 유일하게 비전형적인 표본 하나는 나중에 하이파크로사우루스로 재분류될 가능성이 제기되었지만, 이 종의 모식 표본(기준 표본)은 여전히 코리토사우루스에 속하는 것으로 간주된다.[41]
5. 3. 식성
''코리토사우루스''는 조각류 공룡으로, 초식동물이었다. 벤슨(Benson) 등은 2012년 연구에서 ''코리토사우루스''의 부리가 얕고 섬세하여 부드러운 식물을 먹는 데 사용되었을 것이라고 보았다. 후기 백악기의 기후를 바탕으로, 이들은 ''코리토사우루스''가 가장 즙이 많은 과일과 어린 잎만을 먹는 선택적인 섭식자였을 것으로 추측했다.[32] 한 ''코리토사우루스'' 표본의 흉강(가슴 안쪽 공간)에서는 마지막으로 먹은 것으로 보이는 내용물이 보존된 채 발견되기도 했다. 이 내용물에는 침엽수 잎, 씨앗, 잔가지, 그리고 과일의 잔해가 포함되어 있었는데, 이는 ''코리토사우루스''가 이러한 다양한 식물을 먹었으며, 브라우저(browser)였음을 시사한다.[35]6. 고생태학

코리토사우루스의 화석은 캐나다 앨버타의 상부 올드만 층과 하부 공룡 공원 층에서 발견되었다.[21] 올드만 층은 약 7,750만 년 전에서 7,650만 년 전 사이의 캄파니아절 후기에 해당하며,[44] 공룡 공원 층은 약 7,660만 년 전에서 7,480만 년 전 사이의 시기에 해당한다.[44][43] 코리토사우루스는 약 7,700만 년 전에서 7,570만 년 전 사이에 서식했다. 공룡 공원 층에서는 ''C. casuarius''가 7,660만 년 전에서 7,590만 년 전까지, ''C. intermedius''는 7,580만 년 전에서 7,570만 년 전까지 살았다. 올드만 층에서는 유일하게 발견되는 코리토사우루스 종인 ''C. casuarius''가 약 7,700만 년 전에서 7,650만 년 전 사이에 살았던 것으로 보인다.[44] 모식 표본은 해변으로 떠밀려 온 사체로 추정되며, 주변에서 ''Unio'' 조개껍데기, 물에 마모된 뼈 조각, 배니드 거북 화석 등이 함께 발견되었다.[34] 코리토사우루스는 주로 숲 지대에 서식했으며, 때때로 습지 지역으로 이동했을 가능성이 있다.[32]
올드만 층 상부에서는 코리토사우루스(''Corythosaurus casuarius''와 ''C. intermedius'') 외에도 다양한 공룡들이 함께 서식했다. 여기에는 수각류인 다스플레토사우루스와 사우르니톨레스테스, 하드로사우루스과인 브라킬로포사우루스, 그리포사우루스, 파라사우롤로푸스, 안킬로사우루스과인 스코롤로사우루스, 그리고 각룡류인 코로노사우루스와 케스모사우루스 등이 포함된다. 다른 속들도 발견되지만, 이들은 코리토사우루스와 같은 시기에 살았던 것은 아니다.[44]
''코리토사우루스 카수아리우스''는 공룡 공원 층의 하부에서 널리 발견된다.[22] 이 지층에서 코리토사우루스는 특히 각룡류인 켄트로사우루스 아페르투스와 밀접한 연관성을 보인다. 이러한 연관성은 공룡 공원, 주디스 강 층, 메사베르데 층, 윈드 리버 분지, 위트랜드 카운티 등 여러 지역에서 확인되었다.[45] 코리토사우루스는 이 시기 동안 다양한 대형 초식 공룡들과 공존했다. 지층의 초기 단계에는 하드로사우루스과인 그리포사우루스와 파라사우롤로푸스, 각룡류인 켄트로사우루스와 케스모사우루스, 안킬로사우루스과인 스코롤로사우루스, 에드몬토니아, 디플로사우루스 등이 함께 살았고,[44][44] 중기에는 디플로사우루스, 파노플로사우루스, 유오플로케팔루스가,[44][44] 후기 단계에는 유오플로케팔루스가 공존했다.[44] 이들 공룡의 턱 구조와 기능을 연구한 결과, 매우 혼잡한 생태 환경 속에서 먹이 경쟁을 피하기 위해 서로 약간 다른 생태적 지위를 차지했을 가능성이 제기되었다.[22] 코리토사우루스가 살았던 시기의 유일한 대형 포식자로는 티라노사우루스과인 고르고사우루스 리브라투스와 아직 명명되지 않은 다스플레토사우루스 종이 알려져 있다.[44]
토마스 M. 레만은 코리토사우루스가 해당 지역에서 가장 풍부한 주디스절 공룡 중 하나임에도 불구하고, 캐나다 남부 앨버타 외의 지역에서는 발견되지 않았다는 점을 지적했다.[45] 후기 백악기 북아메리카에 살았던 하드로사우루스와 같은 대형 초식 동물들은 큰 몸집과 높은 이동성을 가졌음에도 불구하고 "상당히 좁은 지리적 범위"를 보였다.[45] 이는 오늘날 대륙의 넓은 지역에 걸쳐 분포하는 대형 포유류 초식 동물의 분포 양상과는 매우 대조적이다.[45]
참조
[1]
웹사이트
Corythosaurus
2022-09-27
[2]
서적
Plants in Mesozoic Time: Morphological Innovations, Phylogeny, Ecosystems
Indiana University Press
2020-10-13
[3]
서적
Barnum Brown: The Man Who Discovered'' Tyrannosaurus rex
University of California Press
2011
[4]
웹사이트
DinoGuide: Corythosaurus casuarius
http://www.carnegiem[...]
Carnegie Museum of Natural History
2014-07-27
[5]
간행물
2001
[6]
논문
New species of crested trachodont dinosaur
[7]
논문
Corythosaurus intermedius, a new species of trachodont dinosaur
[8]
논문
Glishades ericksoni, an indeterminate juvenile hadrosaurid from the Two Medicine Formation of Montana: implications for hadrosauroid diversity in the latest Cretaceous (Campanian-Maastrichtian) of western North America
[9]
논문
Reuniting the "head hunted" Corythosaurus excavatus (Dinosauria: Hadrosauridae) holotype skull with its dentary and postcranium
http://dx.doi.org/10[...]
2017-08
[10]
논문
The Weight of Dinosaurs
http://digitallibrar[...]
[11]
논문
A New Method to Calculate Allometric Length-Mass Relationships of Dinosaurs
http://dinoweb.ucoz.[...]
[12]
서적
The Princeton Field Guide to Dinosaurs
http://worldcat.org/[...]
Princeton University Press
[13]
논문
Testing Size–frequency Distributions As a Method of Ontogenetic Aging: A Life-history Assessment of Hadrosaurid Dinosaurs from the Dinosaur Park Formation of Alberta, Canada, with Implications for Hadrosaurid Paleoecology
[14]
서적
The Dinosauria
University of California Press
2004
[15]
논문
Standardized terminology and potential taxonomic utility for hadrosaurid skin impressions: A case study for Saurolophus from Canada and Mongolia
[16]
논문
The Ilium in Dinosaurs and Birds
http://digitallibrar[...]
[17]
논문
A New Basal Hadrosauroid Dinosaur (Dinosauria: Ornithopoda) with Transitional Features from the Late Cretaceous of Henan Province, China
[18]
논문
Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)
http://www.hopkinsme[...]
[19]
간행물
2000
[20]
논문
Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids
[21]
논문
Reuniting the "head hunted" Corythosaurus excavatus (Dinosauria: Hadrosauridae) holotype skull with its dentary and postcranium
http://dx.doi.org/10[...]
2017-08
[22]
논문
Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada
[23]
간행물
2000
[24]
간행물
1914
[25]
간행물
1914
[26]
간행물
2000
[27]
논문
Taxonomic implications of relative growth in lambeosaurine dinosaurs
[28]
논문
Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis
[29]
간행물
1914
[30]
간행물
1916
[31]
간행물
1916
[32]
간행물
2012
[33]
간행물
1914
[34]
간행물
1916
[35]
간행물
2000
[36]
서적
1916
[37]
논문
A remarkable hollow-crested hadrosaur from Russia: an Asian origin for lambeosaurines
http://doc.rero.ch/r[...]
[38]
서적
1914
[39]
서적
1914
[40]
서적
The Age of Dinosaurs
Publications International, LTD
[41]
서적
Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed
Indiana University Press
[42]
논문
Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology
[43]
서적
2005
[44]
서적
2009
[45]
서적
2001
[46]
웹사이트
https://www.ous.ac.j[...]
[47]
웹사이트
コリトサウルス、米国にも 特定の決めては「とさか」、岡山理科大
https://www.sankei.c[...]
2022-12-26
[48]
웹사이트
貴重な発見「約7600年前の草食恐竜」が岡山理科大学に!? 当時の生態を知る手掛かり【岡山】 {{!}} OHK 岡山放送
https://www.ohk.co.j[...]
2023-09-01
[49]
웹사이트
米で発見の化石はコリトサウルス カナダ以外で初 岡山理科大に展示:山陽新聞デジタル|さんデジ
https://www.sanyonew[...]
2023-09-01
[50]
웹사이트
恐竜「コリトサウルス」の全身骨格を公開 岡山理科大学|NHK 岡山県のニュース
https://web.archive.[...]
2023-09-01
[51]
웹사이트
コリトサウルス、米国の化石にも 広範囲に生息した可能性 岡山理科大に保管:朝日新聞デジタル
https://www.asahi.co[...]
2023-09-01
[52]
웹사이트
恐竜の化石から新発見 約7600万年前の草食恐竜 “コリトサウルス” はアメリカにも生息していた! 国際的な学術誌に掲載&標本展示の見学も可能【岡山】
https://newsdig.tbs.[...]
2023-09-01
[53]
문서
이 머리 장식 스타일의 서양 투구를 "코뤼스" 또는 "코린트식(헬멧)"이라고 부른다.
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com