친타오사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
친타오사우루스는 중국 산둥성에서 발견된 하드로사우루스과 공룡의 한 속이다. 1950년 화석이 발견되었으며, 1958년 양중젠에 의해 명명되었다. 친타오사우루스는 코에 독특한 볏이 있는 것이 특징이며, 처음에는 사우롤로푸스아과로 분류되었으나 람베오사우루스아과로 변경되었다. 볏의 형태에 대한 논쟁이 있었지만, 2013년 연구를 통해 볏의 구조와 기능에 대한 새로운 가설이 제시되었다. 친타오사우루스는 람베오사우루스아과에 속하며, 유럽의 파라라브도돈, 쿠탈리사우루스와 함께 계통군을 형성할 수 있다.

더 읽어볼만한 페이지
- 1958년 기재된 화석 분류군 - 아이히스타에티사우루스
아이히스타에티사우루스는 쥐라기 후기에서 백악기 초기에 살았던 작은 도마뱀으로, 짧고 뭉툭한 주둥이와 평평한 두개골을 가지며, 독일과 이탈리아에서 화석이 발견되었고, 도마뱀붙이와 유사한 등반 생활에 적응했을 것으로 추정되지만, 불완전한 화석 자료로 인해 정확한 분류는 논쟁 중이다. - 1958년 기재된 화석 분류군 - 이크티올레스테스
이크티올레스테스는 에오세 시대에 살았던 멸종된 고래의 한 속으로, 파키스탄에서 화석이 발견되었으며, 육상 포유류와 고래류의 중간 형태를 보이며, 반수생 생활을 했을 것으로 추정된다. - 양중젠이 명명한 분류군 - 오메이사우루스
오메이사우루스는 중국 쓰촨성에서 발견된 용각류 공룡의 한 속으로, 쯔궁 지역에서 많은 화석이 발견되었으며 마멘키사우루스과에 속한다. - 양중젠이 명명한 분류군 - 마멘키사우루스
마멘키사우루스는 중국에서 발견된 용각류 공룡의 한 속으로, 매우 긴 목이 특징이며 쥐라기 후기에서 백악기 초기에 걸쳐 화석이 발견된다. - 하드로사우루스과 - 에드몬토사우루스
에드몬토사우루스는 백악기 후기에 살았던 하드로사우루스과의 대형 초식 공룡으로, 9~12m의 몸길이, 오리 부리 형태의 입, 그리고 피부 자국이 보존된 미라 화석으로 알려져 있으며, *E. regalis*와 *E. annectens* 두 종이 인정되고, 티라노사우루스와 공존했던 생태 환경과 식생, 성장 과정 등에 대한 연구가 활발히 진행되고 있다. - 하드로사우루스과 - 마이아사우라
마이아사우라는 백악기 후기에 북아메리카에서 서식한 8~9m 길이의 하드로사우루스과 초식 공룡으로, 눈 사이의 짧고 단단한 볏과 무리 생활 및 둥지 군집을 통해 새끼를 키운 흔적이 발견된 "좋은 어머니"라는 의미의 속명이 특징이다.

2. 발견 및 명명
1950년, 중국 동부 산둥성 라이양 칭캉커우 근처의 쓰코우에서 대형 하드로사우루스과 공룡의 다양한 유해가 발굴되었다.[3] 1951년 베이징의 양중젠에 의해 발굴이 시작되었으며, 최소 7개체의 부분적으로 연결된 골격이 발견되었다.[9] 1958년, 양중젠은 이 유해들을 모식종 ''친타오사우루스 스피노리누스''(Tsintaosaurus spinorhinus)로 명명했다. 속명은 칭다오시에서 유래되었으며, 과거에는 "친타오"로 음역되었다. 종명은 라틴어 ''spina''(가시)와 그리스어 ῥίς, ''rhis''(코)에서 유래되었으며, 코에 있는 독특한 볏을 가리킨다.[3]
모식 표본 IVPP AS V725는 캄파니아절에 속하는 왕스층군의 일부인 징강커우 지층에서 발견되었다. 이 표본은 두개골을 포함한 부분적인 골격으로 구성되어 있다. 공동 모식 표본은 두개골 지붕인 IVPP V818 표본이다.[3] 같은 지역에서 몇 개의 추가적인 부분 골격과 다수의 분리된 골격 요소들이 발견되었다. 양중젠은 이 중 일부를 ''친타오사우루스''로 분류했으며, 다른 일부는 ''타니우스 칭칸코우엔시스''(Tanius chingkankouensis) 양 1958로 명명했다. 또한 ''타니우스 라이양엔시스''(Tanius laiyangensis) 젠 1976도 존재한다. 이 두 종은 현재 주니어 동의어이거나 ''불확실명''(nomen dubium)으로 간주된다.[3]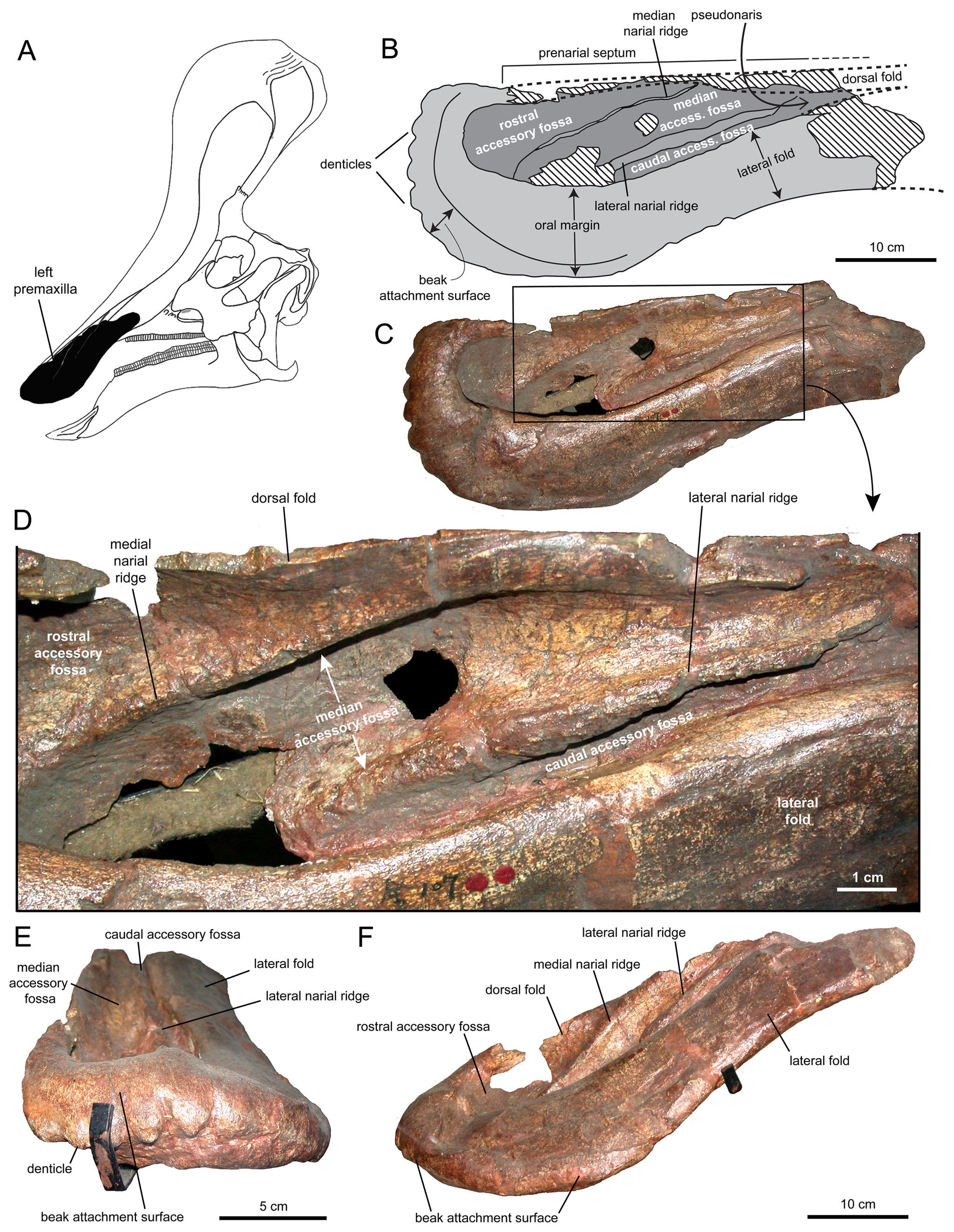
이후 연구자들은 더 많은 재료를 ''친타오사우루스''로 분류했다. 친타오사우루스는 처음에 사우롤로푸스아과로 분류되었으나, 이후 람베오사우루스아과로 변경되었다.[9] 그러나 볏 모양 돌기가 너무 특이하여 다른 람베오사우루스아과의 볏 모양 돌기와 상동 기관인지 의심받기도 했다.[9] 로제스트벤스키는 친타오사우루스 스피노리누스를 타니우스 시넨시스의 동의어로 간주했고, 호너 등은 친타오사우루스가 하드로사우루스아과와 람베오사우루스아과의 키메라일 가능성을 지적했다.[9] 그러나 고생물학자 에릭 뷔페토는 다른 여러 표본에서도 명확한 막대 모양 돌기가 남아 있다는 점, 타니우스와는 다양한 형질상의 차이로 명확히 다른 속이라는 점 등을 들어 이러한 의견을 반박했다.[9]
3. 특징
3. 1. 볏
''친타오사우루스''(Tsintaosaurus)는 원래 머리에 유니콘과 같은 볏이 있는 것으로 복원되었다.[4] 보존된 볏은 약 40cm 길이의 돌기로 구성되어 있으며, 코 뒤쪽 상단에서 거의 수직으로 돌출되어 있다.[4] 이 구조는 속이 비어 있고 윗부분이 갈라진 것처럼 보인다.[4] 1990년, 데이비드 B. 웨이샴펠과 잭 호너는 볏의 존재에 대해 의문을 제기하며, 실제로는 화석의 압착으로 인해 위로 왜곡된, 부러진 비골(nasal bone)일 것이라고 주장했다.[4] 그러나 1993년 에릭 뷔페토 등은 뼈 자체에 대한 재조사를 거쳐 볏이 왜곡된 것도 아니고 복원의 산물도 아니라고 결론지었다.[4]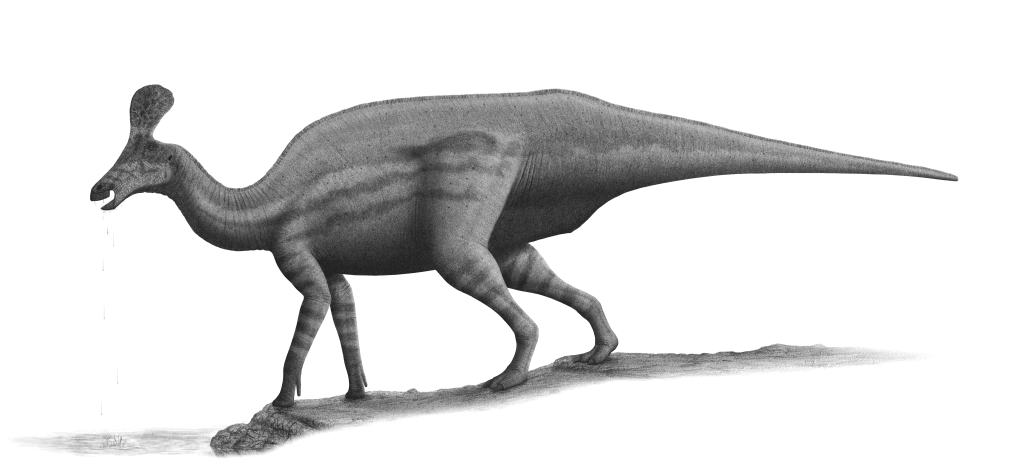

2013년, 알베르트 프리토-마르케스와 조나단 와그너는 새로운 복원을 제시했다.[5] 이들은 볏의 앞부분은 전상악골의 상향 돌기에 의해 형성되었고, 볏 뒤쪽을 형성하는 비골의 볏 돌기의 확장된 윗부분과 롬보이드 접촉 면을 이루었을 것이라고 보았다. 볏의 뒤쪽 기저부는 전두골의 외부 성장물에 의해 덮여 있었고, 융합된 비골은 속이 빈 튜브 구조를 형성했을 것이다.[5] 볏의 높이는 사각형을 따라 측정한 뒤쪽 두개골의 높이를 초과했을 것이며, 볏은 대부분 수직이지만 약간 뒤쪽을 향하고 있었을 것이다.[5]
프리토-마르케스와 와그너는 볏의 내부 공기 통로에 대한 새로운 가설도 제시했다.[5] ''친타오사우루스''가 주둥이에 표준 람베오사우루스류 배열을 가지고 있었을 것이라고 추정했으며, 공기는 흡입할 때 윗부리 뒤쪽의 전상악골의 "가짜 콧구멍"인 쌍을 이룬 ''가상 비공''(pseudonares)을 통해 두개골로 들어갔을 것이다.[5] 거기에서 공기는 전상악골의 중앙 돌기 아래에 있는 쌍을 이룬 통로를 통해 볏 상단으로 운반된 다음, 엽 내의 공통 중앙 챔버로 들어갔을 것이다.[5] 챔버의 뒤쪽은 비골에 의해 형성되었으며 아마도 비강(nasal cavity)과 상동(相同)일 것이다. 챔버는 전상악골의 굽은 중앙 돌기에 의해 앞쪽과 뒤쪽에 각각 하나씩 두 개의 작은 구멍으로 나뉘어, 구멍 사이의 통로를 둘러싼 갈고리를 형성했다.[5] 뒤쪽 구멍에서 공기는 아래로 운반되어 내부 두개골 구멍으로 향했다.[5]
후방 비골의 튜브형 구조가 공기 통로가 아니라는 결론으로 인해 프리토-마르케스와 와그너는 그 기능에 대한 대안적인 설명을 찾아야 했다. 그들은 튜브가 상대적인 강도와 낮은 뼈 질량을 결합하여 볏의 무게를 줄이는 역할을 했을 것이라고 제안했다.[5]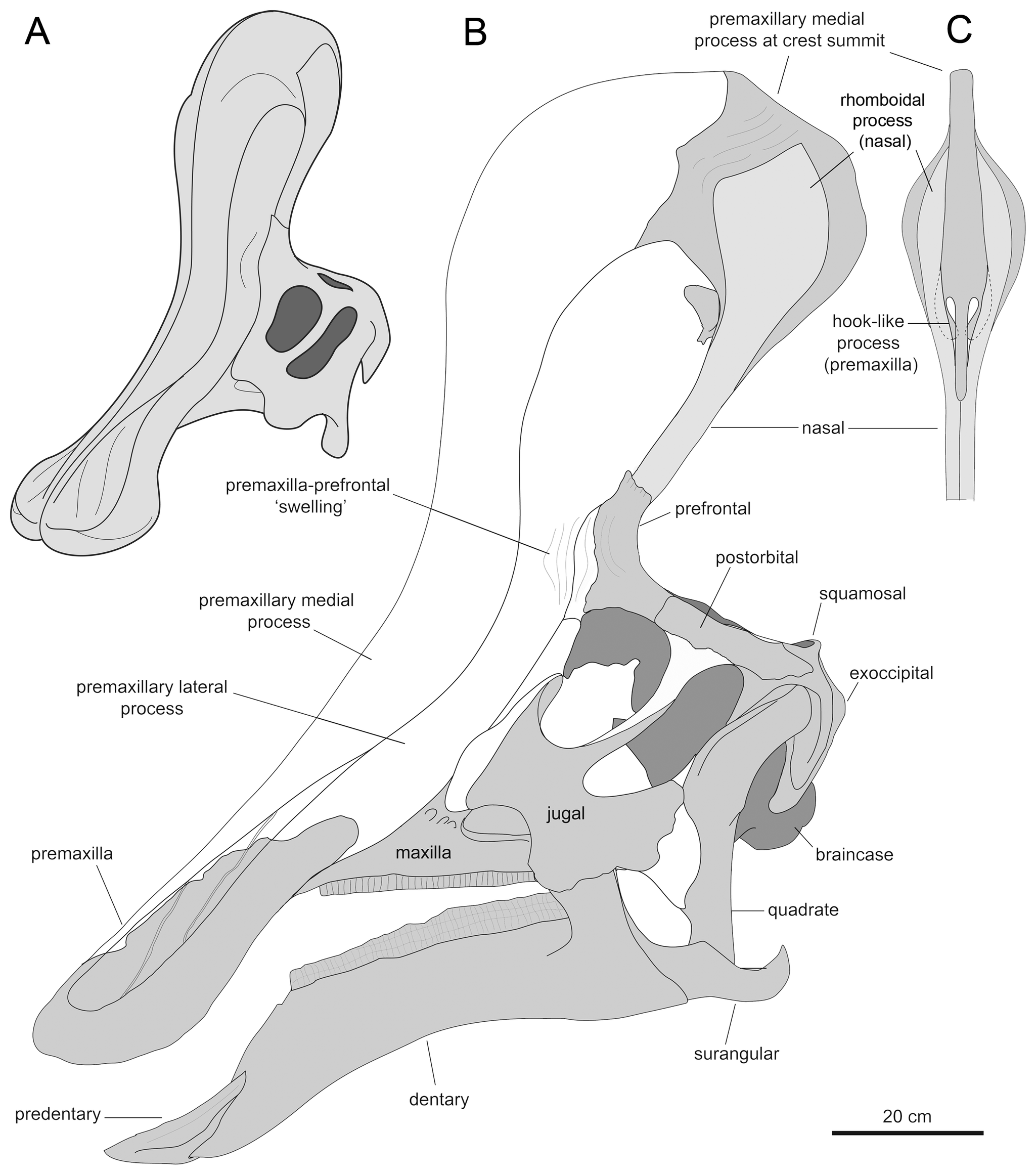


3. 2. 기타 특징
사람과 비교한 ''친타오사우루스''의 크기]]볏을 제외하고, 프리토-마르케스와 바그너는 ''친타오사우루스''의 몇 가지 다른 독특한 특징(자포모르피)을 확인했다.[5] 위쪽 부리의 가장자리는 둥글고 두꺼우며, 콧구멍 주변 앞쪽 함몰부의 가로 너비보다 넓다. 이 함몰부가 앞위턱뼈에 위치해 있는 한, 각 측면은 아래쪽과 옆쪽으로 비스듬히 이어지는 두 개의 능선에 의해 세로로 나뉜다. 내부적으로 융합된 코뼈는 뇌두 앞쪽에 뼈 블록을 형성한다. 코뼈의 뒤쪽은 이마뼈의 앞쪽 연장에 의해 잘려 있으며, 이 중 가장 위쪽은 두개골 지붕에 비해 솟아 있다. 앞위턱뼈의 위쪽 가지는 뒤쪽, 아래쪽 및 약간 안쪽을 향하는 내부 돌기를 가지고 있으며, 정중선에서 공유된 공간을 나눈다. 앞이마뼈는 눈물뼈의 아래쪽 부분에서 앞이마뼈의 위쪽 돌기의 아래쪽 부분까지 이어지고 앞위턱뼈의 측면의 돌기에 연결되어 볏 기저부 측면에 융기를 형성하는 플랜지를 가지고 있다. 앞이마뼈의 측면과 밑면은 깊은 수직 홈을 보인다. 상측두창은 가로 방향으로 길이가 너비보다 넓다.[5]
좌측 전두골]]
마르케스 등은 볏 모양 돌기 외에도 친타오사우루스의 몇 가지 다른 고유 특징을 확인했다. 상악골의 부리 가장자리는 콧구멍 주변의 전방 함몰부 폭보다 넓고 둥글고 두껍다. 이 함몰부는 비스듬히 아래와 옆으로 이어지는 두 개의 융기에 의해 세로로 나뉘어져 있다. 내부적으로는 융합된 비골이 뇌상자 앞에 뼈 블록을 형성한다. 비골 후부는 전두골 연장에 의해 고정되며, 그 최상부는 두개골 정점을 향해 솟아 있다. 전상악골의 위쪽 돌기는 정중선의 공유 공간을 분할하고, 측두골 후방은 약간 내측 후방으로 향한다. 전두골은 누관 하부에서 전두골 하부까지 이어지는 플랜지를 가지며, 볏 모양 돌기 정점의 측면에 융기를 형성하기 위해 전상악골 측면의 측두골에 연결된다. 전두골의 측면과 하면에는 깊은 수직 홈이 보인다. 척추 전정의 개구부는 가로로 길다[11]。
4. 분류
''친타오사우루스''는 람베오사우루스아과에 속하며, 유럽의 파라라브도돈과 쿠탈리사우루스(파라라브도돈의 동의어일 가능성이 있음)와 함께 분기도를 형성할 수 있다.[6] 특히 이들과 친타오사우루스족이라는 클레이드를 공유한다.[12]
2013년 Prieto-Márquez 외 연구에 따르면, ''친타오사우루스''는 람베오사우루스아과 내에서 다음과 같은 위치에 있다:[7][13]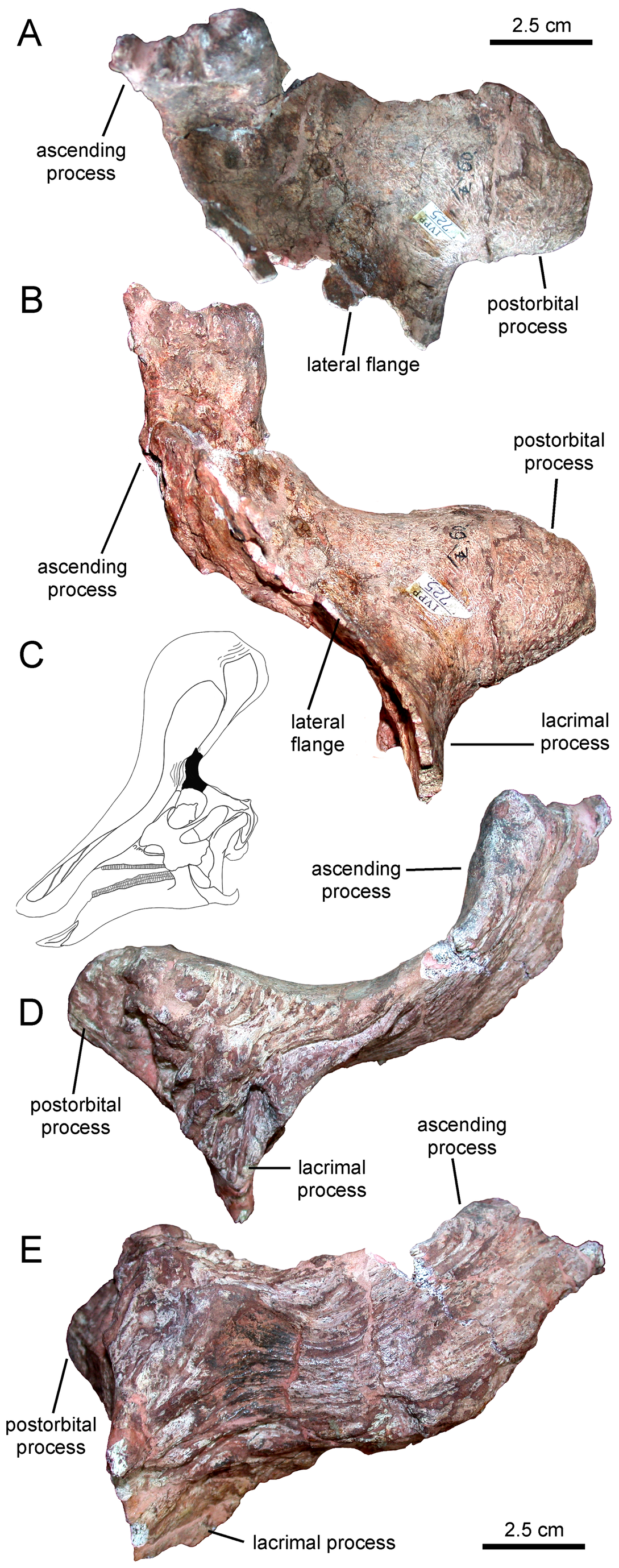
5. 고생태학
산둥성 왕시 계열의 여러 지층, 그중에서도 가장 최근 지층인 징강커우 층의 공룡 알 연구에 따르면, 이 지역은 공룡 다양성이 높았으며, 기후는 이전의 장장딩 층보다 건조해졌음을 알 수 있다.[8]
참조
[1]
웹사이트
Dinosauria Translation and Pronunciation Guide T
http://www.dinosauri[...]
2002
[2]
서적
The Princeton Field Guide to Dinosaurs
https://archive.org/[...]
Princeton University Press
[3]
논문
The dinosaurian remains of Laiyang, Shantung
Whole Number
1958
[4]
논문
"''Tsintaosaurus spinorhinus'' Young and ''Tanius sinensis'' Wiman: a preliminary comparative study of two hadrosaurs (Dinosauria) from the Upper Cretaceous of China"
https://www.research[...]
1993
[5]
논문
The 'Unicorn' Dinosaur That Wasn't: A New Reconstruction of the Crest of ''Tsintaosaurus'' and the Early Evolution of the Lambeosaurine Crest and Rostrum
2013
[6]
논문
"''Pararhabdodon isonensis'' and ''Tsintaosaurus spinorhinus'': a new clade of lambeosaurine hadrosaurids from Eurasia"
2009
[7]
논문
Diversity, relationships, and biogeography of the Lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin ''Canardia garonnensis''
2013
[8]
논문
Dinosaur diversity during the transition between the middle and late parts of the Late Cretaceous in eastern Shandong Province, China: Evidence from dinosaur eggshells
2013
[9]
논문
The dinosaurian remains of Laiyang, Shantung
Whole Number
1958
[10]
간행물
"''Tsintaosaurus spinorhinus'' Young and ''Tanius sinensis'' Wiman: a preliminary comparative study of two hadrosaurs (Dinosauria) from the Upper Cretaceous of China"
C.R. Academy of Science Paris
1993
[11]
논문
The ‘Unicorn’ Dinosaur That Wasn’t: A New Reconstruction of the Crest of ''Tsintaosaurus'' and the Early Evolution of the Lambeosaurine Crest and Rostrum
2013
[12]
논문
"''Pararhabdodon isonensis'' and ''Tsintaosaurus spinorhinus'': a new clade of lambeosaurine hadrosaurids from Eurasia"
2009
[13]
논문
Diversity, relationships, and biogeography of the Lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin ''Canardia garonnensis''
2013
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com