공자새
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
공자새(Confuciusornis)는 전기 백악기(약 1억 3100만 년 ~ 1억 2000만 년 전)에 살았던 멸종된 조류의 속으로, 중국 랴오닝성 일대에서 화석이 발견되었다. 시조새 이후 부리가 있는 가장 오래된 새 중 하나로, 짧은 꼬리와 깃털을 가진 특징을 보이며, 날개 깃털의 구조와 골격 특징을 통해 비행 능력을 가졌을 것으로 추정된다. 공자새는 다양한 종으로 분류되었으나, 현재는 Confuciusornis sanctus 종만이 일반적으로 인정된다. 화석은 주로 예홀 생물군에서 발견되며, 이 지역의 지질 환경과 다양한 동식물 화석과 함께 연구된다.
더 읽어볼만한 페이지
- 백악기의 새 - 시노르니스
시노르니스는 중국에서 발견된 깃털 공룡 화석으로, 흉골과 어깨뼈 구조로 비행 능력을, 발가락 구조로 나무 위 생활에 적응했을 것으로 추정되며, 시조새와 유사하지만 짧은 꼬리와 부리 등의 차이점을 보이는 조류 진화 연구의 중요한 중간 형태이다. - 아시아의 고대 새 - 시노르니스
시노르니스는 중국에서 발견된 깃털 공룡 화석으로, 흉골과 어깨뼈 구조로 비행 능력을, 발가락 구조로 나무 위 생활에 적응했을 것으로 추정되며, 시조새와 유사하지만 짧은 꼬리와 부리 등의 차이점을 보이는 조류 진화 연구의 중요한 중간 형태이다. - 백악기 전기 아시아의 새 - 시노르니스
시노르니스는 중국에서 발견된 깃털 공룡 화석으로, 흉골과 어깨뼈 구조로 비행 능력을, 발가락 구조로 나무 위 생활에 적응했을 것으로 추정되며, 시조새와 유사하지만 짧은 꼬리와 부리 등의 차이점을 보이는 조류 진화 연구의 중요한 중간 형태이다. - 백악기 전기 아시아의 새 - 프로오르니스
프로오르니스는 신생대 에오세 중기 아르헨티나에 서식했던 날지 못하는 거대한 새이며, 초기 조류 진화의 다양성을 보여주는 중요한 증거로 여겨진다.
| 공자새 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Confuciusornis |
| 명명자 | Hou et al., 1995 |
| 모식종 | †Confuciusornis sanctus |
| 모식종 명명자 | Hou et al., 1995 |
| 종 | |
| 기타 종 | †C. dui Hou et al., 1999 †C. feducciai Zhang et al., 2009 †C. jianchangensis Li et al., 2010 †C. shifan Wang et al., 2022 |
| 속 동의어 | |
| 종 동의어 (C. sanctus) | |
| 개요 | |
| 생존 시기 | 백악기 초기, 1억 2500만 년 전 ~ 1억 2000만 년 전 |
| 과 | Confuciusornithidae |
| 목 | Confuciusornithiformes |
2. 역사

1993년 11월, 베이징의 중국 고생물학 연구소(IVPP) 고생물학자 허우 롄하이와 후 요밍은 진저우의 화석 수집가 장허의 집에서 지역 벼룩시장에서 구입한 화석 조류 표본을 보았다. 12월에 허우는 양위산이라는 농부가 발견한 두 번째 표본을 알게 되었는데, 두 표본 모두 베이퍄오 시(北票市) 샹위안에서 같은 장소에서 발견되었다.[5][6] 1995년, 허우와 동료들은 이 두 표본과 세 번째 표본을 새로운 속과 종인 ''Confuciusornis sanctus''로 공식 기재했다. 속명은 철학자 공자와 그리스어 ὄρνις(''ornis''), "새"를 결합한 것이고, 종명은 라틴어로 "거룩한 자"를 의미하며 중국어 圣贤(''shèngxián)'', "현자"를 번역한 것이다. 최초 발견된 표본(IVPP V10918)은 모식표본으로 지정되었고, 다른 두 골격(IVPP V10895, IVPP V10919–10925)은 부모식표본으로 지정되었다.[7] 부모식표본은 모식표본에서 알려지지 않은 뼈만 포함하고 있어 종에 대한 언급은 추측일 뿐이었다.[8] 그 직후 많은 수의 보존된 표본이 발견되어 단일 종을 나타낸다는 것이 확인되었다.[22]
''Confuciusornis''는 초기 포유류 ''장허수리움''과 함께 예홀 생물군의 가장 주목할만한 화석 발견으로 여겨졌다.[49][9] 1990년대 후반, ''Confuciusornis''는 시조새 이후 가장 오래된 부리가 있는 새로 여겨졌고, 이셴 층은 후기 쥐라기 (티톤절) 시대인 것으로 여겨졌다. 1992년에 ''시노르니스''와 ''카타이오르니스''가 주포탕 층에서 기재되었지만, 조각난 잔해만을 기반으로 하였다.[29][7][9] 이후 두 지층 모두 전기 백악기(바레미안에서 압트절, 1억 3100만 년에서 1억 2000만 년 전)로 연대가 측정되었다.[69]
1995년, 지역 농부들은 베이퍄오 시 허툰 부근에서 화석 발굴을 시작했고, 1997년부터 IVPP에서 대규모 발굴이 수행되어 수백 개의 ''Confuciusornis'' 표본이 회수되었다.[5][9] 랴오닝, 허베이, 내몽골을 포함한 넓은 지역에서 예홀 생물군의 화석을 산출하는 많은 지역이 확인되었다.[49] 화석의 풍부함, 보존 상태, 상업적 가치로 인해 지역 농부들의 발굴은 많은 수의 화석을 산출했으며, 일부는 국외로 밀반출되었다.[6][29] 1999년, 중국 국립 지질 박물관은 거의 100개의 ''Confuciusornis'' 표본을 소장하고 있는 것으로 추정되었고,[22] 2010년에는 산둥 톈위 자연사 박물관이 536개의 표본을 소유하고 있다고 보고되었다.[43] 그러나 대부분의 표본은 개인 소장이라 연구에 사용할 수 없다.[57]
약 100 m2의 표면에서 40개체가 발견되었는데, 이는 응회암이 호수 퇴적물로 퇴적되도록 한 화산 폭발 후 잿더미, 열 또는 유독 가스에 의해 전체 조류 무리가 동시에 죽은 결과로 설명되었다.[10]
2. 1. 발견
1993년 11월, 베이징의 중국 고생물학 연구소(IVPP)의 고생물학자 허우 롄하이와 후 요밍은 진저우의 화석 수집가 장허의 집에서 화석 조류 표본을 보았다.[5][6] 1995년, 허우와 동료들은 이 표본들을 기반으로 새로운 속과 종인 ''Confuciusornis sanctus''를 공식적으로 기재했다. 속명은 철학자 공자와 그리스어 ὄρνις(''ornis''), "새"를 결합한 것이다.[7]거의 같은 시기에 발견된 초기 포유류 ''장허수리움''과 함께, ''Confuciusornis''는 예홀 생물군의 가장 주목할만한 화석 발견으로 여겨졌다.[49][9] 1990년대 후반, ''Confuciusornis''는 시조새 이후 가장 오래된 부리가 있는 새로 여겨졌다.[29][7][9] 이후 이셴 층과 지우포탕 층 두 지층 모두 전기 백악기(바레미안절~압트절, 131Ma영어~120Ma영어)로 연대가 측정되었다.[69]
1995년부터 지역 농부들과 IVPP에서 화석 발굴을 시작하여 수백 개의 ''Confuciusornis'' 표본을 포함한 많은 화석이 발견되었다.[5][9] 화석의 풍부함, 보존 상태, 상업적 가치로 인해 지역 농부들의 발굴은 이례적으로 많은 수의 화석을 산출했으며, 일부는 국외로 밀반출되기도 했다.[6]
2. 2. 추가 종 및 동의어
''Confuciusornis sanctus''가 처음 기술된 이후, 5개의 추가 종이 공식적으로 명명되어 기술되었다. 그러나 종 간의 차이점을 종 내에서 발생하는 변동과 구별하기 어려워, ''C. sanctus''만이 보편적으로 인정받고 있다.[49]- ''Confuciusornis chuonzhous'': 1997년 Hou에 의해 명명되었으며, 원래 ''C. sanctus''의 모식표본인 IVPP V10919를 기반으로 하였다. 종명은 베이퍄오의 옛 이름인 추안저우를 가리킨다.[11] 현재 일반적으로 ''C. sanctus''와 같은 종으로 간주된다.[12]
- ''Confuciusornis suniae'': 1997년 Hou가 발표한 논문에서 명명되었으며, 표본 IVPP V11308을 기반으로 한다. 종명은 화석을 IVPP에 기증한 Liang Shikuan의 아내인 Sun 부인을 기리기 위한 것이다.[11] 현재 일반적으로 ''C. sanctus''와 같은 종으로 간주된다.[12]
- ''Confuciusornis dui'': 1999년 Hou와 동료들에 의해 명명되었다. 종명은 기증 수집가인 Du Wengya를 기리기 위한 것이다. 모식표본(IVPP V11553)은 한 쌍의 긴 꼬리 깃털과 뿔이 있는 부리의 흔적이 있는 거의 완전한 성체 골격이다. 두 번째 표본인 모식표본 IVPP 11521은 단편적이며 일부 척추와 갈비뼈, 꼬리, 흉골 및 골반, 그리고 대퇴골을 포함한다. Hou와 동료들에 따르면, ''C. dui''는 대부분의 다른 ''Confuciusornis'' 표본보다 작고 가늘며, 모식표본은 ''C. sanctus''의 모식표본보다 약 15% 작고, 해당 종의 더 큰 개체보다 약 30% 작았다. 턱 끝은 ''C. sanctus''보다 더 뾰족했고, 하악골에는 ''C. sanctus''에서 뚜렷하게 나타나는 밑면 용골이 없었다. 첫 번째 손가락의 발톱은 ''C. sanctus''처럼 확대되지 않았다. 흉골은 더 길쭉했고 해부학적 세부 사항에서 달랐으며, 뒷다리의 하부 마디(중족골)는 꼬리의 꼬리뼈보다 짧았다.[13] 2011년 Marguán-Lobon과 동료들의 통계 분석 결과, ''C. sanctus''의 가장 작은 크기 부류에 해당하는 표본과 유의미한 차이가 나타나지 않아, 추정되는 차이점이 단일 종의 개체 변동임을 시사했다. 그러나 이들은 ''C. dui'' 모식표본의 위치를 다시 찾을 수 없었고, 이는 아마도 분실되었을 것이며, 따라서 해당 표본의 주형에 의존하여 측정을 해야 했다. 종의 유효성을 평가하기 위해 ''C. dui'' 표본에 대한 재연구가 필요할 것이다.[62]
- ''Confuciusornis feducciai'': 2009년 Zhang Fucheng과 동료들에 의해 명명되었으며, 종명은 조류학자 앨런 페두치아를 기리고 있다. 모식표본인 D2454는 시허툰 지역에서 발견되었으며 다롄 자연사 박물관에 보관되어 있다. Zhang과 동료들에 따르면, ''C. feducciai''는 더 큰 크기, 골격 비율 및 여러 형태학적 특징에서 다른 ''Confuciusornis'' 종과 달랐다. 앞다리는 뒷다리보다 15% 더 길었고, ''C. sanctus''에서는 길이가 같았다. 상완골의 상단에는 다른 ''Confuciusornis'' 표본에서 특징적인 큰 구멍(공)이 없었다. 첫 번째 손가락의 첫 번째 마디는 더 가늘었다. 다른 차이점은 쇄골에서 나타났는데, 쇄골은 V자 모양이었고, 흉골은 길이보다 넓었으며, 좌골은 치골에 비해 길었다.[14] 2011년 Marguán-Lobon과 동료들은 이러한 진단에 문제가 있다고 주장했다. 상완골의 큰 구멍은 왼쪽 상완골에는 없는 것으로 보였지만, 모식표본의 오른쪽 상완골에는 분명히 존재했다. 또한, 그들의 통계 분석에 따르면 해당 표본은 ''C. sanctus'' 변동의 연속체 안에 잘 들어맞았다. 따라서 이들은 ''C. feducciai''가 ''C. sanctus''와 동일(하위 동의어)하다고 제안했다.[62]
- ''Confuciusornis jianchangensis'': 2010년 Li Li와 동료들에 의해 명명되었으며, 토우다잉쯔에서 발견된 표본 PMOL-AB00114를 기반으로 했다. 대부분의 다른 종들이 이셴 층에서 유래하는 것과 달리, ''C. jianchangensis''는 지우포탕 층에서 발견된다.[15]
2002년 Hou는 ''Jinzhouornis'' 속을 명명했지만, Chiappe 등(2018)과 Wang 등(2018)은 형태 계량 및 알려진 공자새류 표본 검사를 통해 이 속이 ''Confuciusornis''의 하위 동의어임을 보여주었다.[57][16]
3. 특징
孔子鳥중국어는 계통발생학적으로 기저 형질과 파생 형질이 섞여 있는 것이 특징이다. 꼬리뼈와 뼈로 된 흉골(가슴뼈)을 가지고 있어 짧은 꼬리를 가졌다는 점에서 시조새보다 더 "진보"되었지만, 앞다리에 큰 발톱이 있고, 닫힌 눈구멍을 가진 원시적인 두개골을 가지며, 비교적 작은 가슴뼈를 가지고 있다는 점에서는 현대 조류보다 더 "원시적"이었다.[39]
3. 1. 크기
孔子鳥屬|공자새속중국어는 현대의 까마귀 크기 정도로, 전체 길이는 약 50cm, 날개 길이는 최대 70cm였다. 몸무게는 0.2kg에서 0.5kg까지 추정된다.3. 2. 두개골
두개골은 옆에서 보면 거의 삼각형 모양이었으며, 이빨이 없는 부리는 튼튼하고 뾰족했다. 턱 앞쪽에는 각질성 람포테카(뿔로 덮인 부리)와 관련된 깊은 신경혈관 구멍과 홈이 있었다. 외비공(뼈로 된 콧구멍)은 거의 삼각형 모양이었고, 주둥이 끝에서 멀리 떨어져 있었다.[22] ''공자새''는 1990년대 후반에 조상 이궁류 두개골(두개골 양쪽에 두 개의 측두창이 있음)을 가진 최초의 알려진 조류로 여겨졌지만, 2018년에 엘자노프스키와 동료들은 공자조류의 측두 부위에서 보이는 구성이 이들의 그룹에 대해 자폐형질 (원시 상태에서 유지된 것이 아니라 이차적으로 진화한 고유한 특성)라고 결론 내렸다. 일부 표본에서 부리의 각질 덮개가 보존되어 종 간의 차이를 보여준다.[22]3. 3. 골격
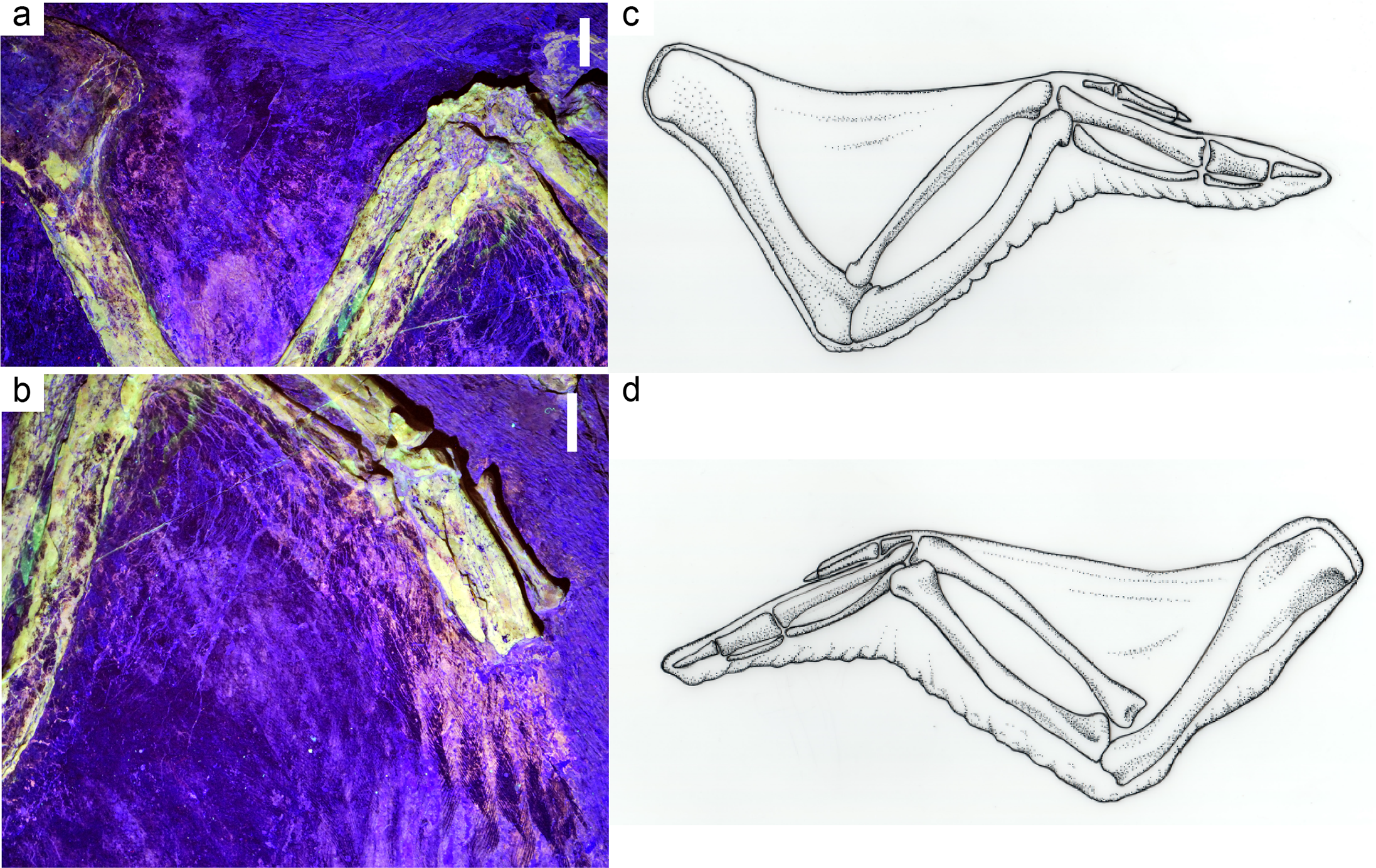
''공자새''의 날개 깃털은 길고 현대적인 모습이었다. 0.5kg 개체의 주익은 길이가 20.7cm에 달했다. 가장 긴 5개의 주익(''주우 primarii'')은 손 길이의 3.5배가 넘었고 어떤 현생 조류보다도 상대적으로 길었지만, 아래팔의 부익은 그에 비해 상당히 짧았다.[41] 가장 바깥쪽 주익은 두 번째 바깥쪽 주익보다 훨씬 짧아 비교적 둥글고 넓은 날개를 형성했다. 날개 모양은 현생 조류에서 발견되는 특정 모양과 일치하지 않는다.[25] 주익은 정도의 차이는 있지만 비대칭적이었고, 특히 가장 바깥쪽 주익에서 두드러졌다. 상완에 삼익이 있었는지는 불분명하다. 일부 표본에서는 날개 깃털 윗부분을 덮고 있는 윤곽깃털이 보존되었고, 일부 표본에서는 몸의 윤곽 깃털이 보존되었다.[22]
일부 진화된 조류와 달리, ''공자새''는 알룰라, 즉 "가짜 날개"가 없었다. 현대 조류에서 알룰라는 손의 첫 번째 손가락에 고정된 깃털로 형성되지만, 이 손가락은 깃털이 없고 ''공자새''의 날개 몸체와 독립적인 것으로 보인다.[22] 디터 슈테판 페터스에 따르면, 알룰라가 없는 것을 보완하기 위해 세 번째 손가락이 주 날개 아래에 별도의 날개를 형성하여 항공기의 플랩과 같은 역할을 했을 수 있다.[24] 상대적으로 진화된 긴 날개 깃털에도 불구하고, 앞팔 뼈에는 깃털 자루 돌기(''papillae ulnares'')나 깃털 인대 부착점의 흔적이 없었다.[22]
많은 표본에서 몸 전체 길이보다 더 길어진 한 쌍의 길고 좁은 꼬리 깃털이 보존되어 있다. 대부분의 현생 조류 깃털과 달리 이 깃털은 대부분의 길이에서 중앙 깃대와 바브로 분화되지 않았다. 대신, 깃털의 대부분은 폭이 약 6mm인 리본 모양의 시트를 형성했다. 깃털의 마지막 4분의 1 지점에서, 둥근 끝을 향해 깃털이 서로 맞물린 바브가 있는 중앙 샤프트로 분화되었다. 많은 ''공자새'' 개체에게는 이러한 꼬리 깃털조차 없었는데, 이는 성적 이형성 때문일 가능성이 있다. 미골 주위의 나머지 꼬리는 현대 조류 꼬리의 익숙한 깃털 부채 대신 몸의 윤곽 깃털과 유사한 짧고 비공기역학적인 깃털 덩어리로 덮여 있었다.[22]
두 개의 ''공자새'' 표본의 레이저 형광은 연조직 해부학에 대한 추가 세부 정보를 보여주었다. ''공자새''의 날개막은 크고, 아마도 비교적 두꺼웠으며, 어깨에서 손목까지 뻗어 있었고, 이는 현대 조류와 유사했다. 후날개막의 범위 또한 현대 조류와 유사하다. 그물 모양의 비늘이 발의 아랫면을 덮고 있었고, 지골과 중족골은 크고 살이 많은 패드를 지지했지만, 지골간 패드는 작거나 전혀 없었다.[25]
3. 4. 깃털 및 연조직
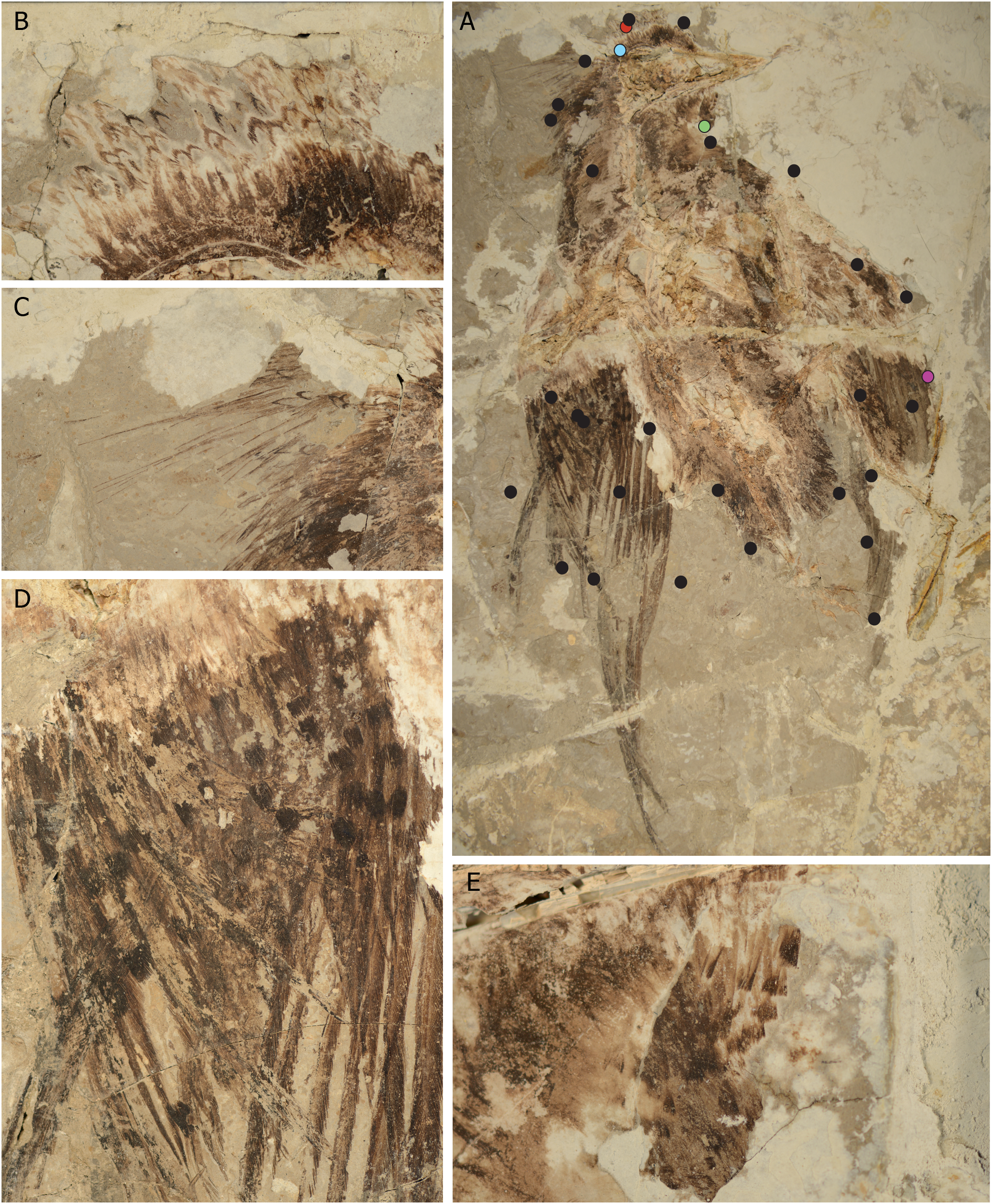
2010년 초, 장푸청(Zhang Fucheng)이 이끄는 과학자 그룹은 색상을 포함하는 멜라노솜(세포 소기관)이 보존된 화석을 조사했다. 이 화석들을 전자 현미경으로 연구하여, IVPP V13171 화석 ''공자새'' 표본에서 멜라노솜이 보존되어 있음을 발견했다. 멜라노솜은 유멜라노솜과 페오멜라노솜의 두 가지 유형으로 존재한다고 보고했는데, 이는 ''공자새''가 회색, 적갈색 및 검은색의 색상을 띠었으며, 아마도 현대의 호금조와 비슷했을 것이라는 것을 나타낸다. 또한 초기 조류 화석에서 페오멜라노솜이 보존된 것이 발견된 것은 처음이었다.[26]
그러나 두 번째 연구팀은 보고된 페오멜라노솜의 흔적을 발견하지 못했다. 2011년 연구에서는 특정 금속, 예를 들어 구리의 존재와 보존된 멜라닌 사이의 연관성을 발견했다. 멜라노솜의 화석 인상과 깃털 속 금속의 존재를 결합하여, 두 번째 과학자 팀은 ''공자새''를 어두운 색의 몸통 깃털과 날개 윗부분 깃털을 가진 모습으로 재구성했지만, 대부분의 날개 깃털에서는 멜라노솜이나 금속의 흔적을 발견하지 못했다. 그들은 ''공자새''의 날개가 흰색이거나, 카로티노이드 색소로 칠해졌을 가능성이 있다고 제안했다. 수컷 표본의 긴 꼬리 깃털 또한 전체 길이에 걸쳐 어두운 색을 띠었을 것이다.[27]
2018년 CUGB P1401 표본에 대한 연구는 ''공자새''의 날개, 목, 볏에 짙은 반점이 존재함을 나타냈다.[28]
4. 분류
1995년 허는 ''공자새''를 공자새과(Confuciusornithidae)에 분류했다. 처음에는 에난티오르니테스의 일원이며 ''고비테릭스''의 자매 분류군이라고 추정했지만, 나중에는 ''공자새''가 에난티오르니테스가 아니며 더 큰 도마뱀꼬리류(Sauriurae) 내에서 에난티오르니테스의 자매 분류군이라고 결론지었다.[29] 그러나 이는 도마뱀꼬리류가 측계통군(paraphyletic)이라고 간주한 Chiappe에 의해 비판받았다.[30] 2001년 지창은 새가슴류(Ornithothoraces)의 자매 분류군이라는 대안적인 위치를 제시했다.[31]
2002년 지의 가설은 Chiappe의 분지학(cladistic analysis)에 의해 확증되었으며, ''공자새''는 꼬리자루류(Pygostylia)의 가장 기초적인 구성원으로 정의되었다.[32] ''공자새''는 ''시조새''보다 더 "원시적인" 두개골을 가지고 있지만, ''시조새''의 긴 꼬리를 잃고 융합된 꼬리 척추인 꼬리자루를 발달시킨 최초의 새로 알려져 있다.[33]
''공자새''의 가까운 친척인 공자새과 ''창청오르니스 헝다오지엔시스'' 역시 이셴 층에서 살았다.
5. 고생물학
공자새는 크고 살집이 많은 발가락 패드, 작은 발가락 사이 패드, 발바닥에 그물 모양 비늘만 있어(유연성 증가) 굽은 발톱을 가진다. 이는 현대 수목 서식 조류와 공유하는 특징으로, 공자새가 나무 위에서 생활했을 가능성을 시사한다.[25]
공자새의 눈을 지지하는 공막환은 현대 조류 및 다른 파충류와 비교했을 때, 대부분의 현대 조류와 유사하게 주행성이었을 가능성을 보여준다.[36]
공자새는 20주가 채 안 되어 성체 크기에 도달했지만, 성장률은 현대 조류보다 느렸다.[56] 성적 성숙은 최종 크기에 도달하기 훨씬 전에 발생했을 것으로 추정된다.[63]
1998년 푸청 장과 동료들은 주사 전자 현미경으로 넙다리뼈 횡단면을 분석, 뼈에 혈관이 풍부하고 단일 성장 정지선만 나타나는 것을 확인했다. 이를 통해 공자새의 성장이 빠르고 지속적이며, 온혈동물이었을 것이라고 결론지었다.[52] 뼈 구조가 현대 변온성 앨리게이터와 달리 깃털 달린 비조류 공룡 베이피아오사우루스와 유사하다는 점을 들어 이 주장을 뒷받침했다.[53]
2003년 아르망 드 리클레와 동료들은 80개 뼈 박편 연구를 통해 공자새가 비조류 수각류와 유사한 빠른 성장률을 보였다고 밝혔다.[56]
2013년 아누수야 친사미와 동료들은 수질골을 통해 꼬리가 짧은 표본을 암컷, 긴 꼬리 표본을 수컷으로 추정했다.[63]
2018년 징마이 오코너와 동료들은 수질골 식별에 의문을 제기하며, 에난티오르니테스(Enantiornithes)의 후지에서 수질골을 확인하고, 기초 조류의 공룡과 같은 성장 방식을 뒷받침했다.[64]
1999년, 중국 고생물학자 Hou Lianhai와 동료들은 부리가 위로 굽어 있고 맹금류가 아니라는 점을 들어 공자새가 초식성일 가능성이 높다고 주장했다.[13] 같은 해, 고생물학자 Dieter S. Peters와 Ji Qiang은 발가락 물갈퀴의 잔해가 보존되지는 않았지만, 다소 부드러운 부리를 사용하여 수영하면서 물속에서 먹이를 잡았다고 가설을 세웠다. 2002년, 폴란드 고생물학자 안제이 엘자노프스키는 공자새가 물이나 지면에서 먹이를 잡아 날아다니는 방식으로 먹이를 찾았다고 제안했다.[65]
2003년, 중국 고생물학자 Zhonghe Zhou와 Fucheng Zhang은 튼튼하고 이빨이 없는 턱은 씨앗을 먹었을 수 있음을 시사했다.[66] 2006년, Johan Dalsätt과 동료들은 ''C. sanctus'' 표본(IVPP V13313)에서 작은 물고기, 아마도 ''Jinanichthys''의 뼈를 발견하고 공자새가 잡식성이었을 것이라고 제안했다.[67] 안드레이 지노비예프는 공자새가 날아가면서 물고기를 잡았다고 추정했다.[68]
공자새의 두개골은 상대적으로 움직이지 않았으며, 현대 조류의 운동성이 없었다.[22]
5. 1. 비행

많은 ''공자새'' 표본은 일부 현대 극락조와 유사하게 한 쌍의 길고 깃털 모양의 꼬리 깃털을 보존하고 있다.[46] 깃털이 없는 표본도 신체 나머지 부분에는 정교하게 보존된 깃털을 가지고 있어, 깃털 유무가 단순히 보존 불량 때문이 아님을 보여준다.[22] 1998년 래리 마틴과 동료들은 당시 알려진 표본의 약 5~10%에서 긴 꼬리 깃털이 발견된다고 언급했다.[39] 2011년 예수스 마루간-로본과 동료들의 분석에 따르면, 130개의 표본 중 18%가 긴 꼬리 깃털을 가졌고, 28%는 없었으며, 나머지 54%는 보존 상태가 불량하여 깃털 유무를 판단하기 어려웠다.[62]
이러한 패턴의 생물학적 의미에 대해서는 논란이 있었다.[62] 마틴과 동료들은 이 패턴이 성적 이형성을 반영하며, 깃털 모양의 깃털은 구애 과시에 사용된 한쪽 성별(아마 수컷)에만 존재했을 것이라고 제안했다.[39] 이러한 해석은 후속 연구 대부분에서 지지되었다.[63] 1999년 치아페와 동료들은 성적 이형성이 유일한 설명은 아니지만 가장 합리적인 설명이며, 현대 조류에서 장식 깃털의 길이가 종종 성별에 따라 다르다고 언급했다.[22]
알려진 ''공자새'' 표본이 소형 및 대형 그룹으로 나뉘지만, 이러한 이봉 분포가 긴 꼬리 깃털 소유와 관련이 없다는 관찰 결과는 논란을 일으켰다. 2008년 치아페와 동료들은 이러한 크기 분포가 공룡과 유사한 성장 방식(성장 단락 참조)으로 설명될 수 있으며, 성적 이형성이 긴 꼬리 깃털 유무에 대한 가장 유력한 설명이라고 주장했다.[57] 그러나 2009년 빈프리트와 디터 페터스는 대부분의 현대 조류에서 유사한 깃털이 나타나는 경우처럼, 두 성별 모두 긴 꼬리 깃털을 가졌을 가능성이 높다고 반박했다. 다만 한쪽 성별이 다른 쪽보다 더 컸을 것이다(성적 크기 이형성). 이들은 ''공자새''의 크기와 긴 꼬리 깃털 분포가 현대 꿩꼬리물떼새(''Hydrophasianus chirurgus'')와 유사하다고 제안했는데, 이 물새는 암컷이 가장 크고 두 성별 모두 번식기에만 긴 꼬리를 가진다. ''공자새''는 가장 작은 표본에서도 긴 꼬리 깃털이 존재한다는 점에서 물떼새와 다르며, 이는 긴 꼬리 깃털이 생식 기능이 없었을 수도 있음을 시사한다.[58]
긴 꼬리 깃털이 없는 현상을 설명하기 위해 몇 가지 다른 가설이 제시되었다. 1999년 연구에서 치아페와 동료들은 개체가 털갈이 중에 사망하여 꼬리 깃털이 없을 가능성을 논의했다. 초기 조류의 털갈이에 대한 직접적인 증거는 없지만, ''공자새'' 표본의 깃털 마모가 없다는 것은 깃털이 주기적으로 갱신되었음을 시사한다. 현대 조류처럼 털갈이하는 개체와 하지 않는 개체가 함께 존재했을 수 있으며, 수컷과 암컷이 일 년 중 다른 시기에 털갈이를 했을 수 있어 긴 꼬리 깃털 유무 표본이 함께 발견되는 것을 설명할 수 있다.[22] 반면 페터스와 페터스는 ''공자새''가 여러 현존 종들이 사용하는 방어 기제인 깃털을 떨어뜨렸을 수 있다고 제안했다. 이러한 탈락은 동물을 묻은 화산 폭발로 인한 스트레스 때문에 유발되었을 수 있으며, 그 결과 깃털이 없는 많은 표본이 생겨났을 것이다.[58] 2011년 논문에서 예수스 마루간-로본과 동료들은 긴 꼬리 깃털 유무에 따라 두 개의 별개 종이 존재했을 가능성도 고려해야 한다고 언급했다. 그러나 이 가능성은 현재 다른 해부학적 차이점이 명확하지 않아 입증되지 않았다.[62]
5. 2. 꼬리 깃털
많은 ''공자새'' 표본에서 발견되는 한 쌍의 길고 깃털 모양의 꼬리 깃털은, 일부 현대의 극락조에서 나타나는 것과 유사하다.[46] 이러한 깃털이 없는 표본도 몸의 나머지 부분에는 깃털이 잘 보존되어 있는 경우가 있어, 깃털이 없는 것이 단순히 보존 상태가 나빠서가 아니라는 것을 알 수 있다.[22]이러한 꼬리 깃털의 존재 유무에 따른 패턴의 생물학적 의미는 논란이 있는데,[62] 1998년 래리 마틴과 동료들은 꼬리 깃털이 성적 이형성을 반영하여 수컷에게만 존재하며 구애 과시에 사용되었을 것이라고 제안했다.[39] 이후 연구에서도 이러한 해석이 지지되었다.[63] 1999년, 치아페와 동료들은 성적 이형성이 유일한 설명은 아니지만 가장 합리적인 설명이라고 언급했다.[22]
2009년 빈프리트와 디터 페터스는, 대부분의 현대 조류에서 유사한 깃털이 나타나는 경우처럼, 암수 모두 긴 꼬리 깃털을 가지고 있었을 가능성이 높다고 반박했다. 다만 한쪽 성별이 다른 쪽보다 더 컸을 것이라고 추정했다(성적 크기 이형성). 이들은 ''공자새''의 크기와 긴 꼬리 깃털의 분포가 현대의 꿩꼬리물떼새와 유사하다고 제안했는데, 꿩꼬리물떼새는 암컷이 가장 크고, 암수 모두 번식기에만 긴 꼬리를 가진다. 그러나 ''공자새''는 가장 작은 표본에서도 긴 꼬리 깃털이 존재한다는 점에서 꿩꼬리물떼새와는 다르며, 이는 긴 꼬리 깃털이 생식 기능이 없었을 수도 있음을 시사한다.[58]
긴 꼬리 깃털이 없는 현상을 설명하기 위해 다른 가설들도 제시되었다. 1999년 치아페와 동료들은 털갈이 중에 꼬리 깃털이 빠졌을 가능성을 제시했다. ''공자새'' 표본에 깃털 마모가 없다는 것은 깃털이 주기적으로 갱신되었음을 의미하며, 현대 조류처럼 털갈이를 하는 개체와 하지 않는 개체가 함께 있었을 수 있고, 암수가 다른 시기에 털갈이를 했을 수 있다는 것이다.[22] 페터스와 페터스는 ''공자새''가 스트레스를 받으면 깃털을 떨어뜨리는 방어 기제를 사용했을 수 있다고 제안했다.[58] 2011년 예수스 마루간-로본과 동료들은 긴 꼬리 깃털이 있는 종과 없는 종, 즉 두 개의 별개 종이 존재했을 가능성도 고려해야 한다고 언급했지만, 다른 해부학적 차이점이 명확하지 않아 입증하기는 어렵다고 보았다.[62]
5. 3. 번식
2007년, 오른쪽 발 근처에 알이 보존된 ''공자새'' 골격이 보고되었는데, 이는 암컷일 가능성을 시사한다.[47] 이 알은 둥근 모양이며 지름이 17mm인데, 동물의 머리보다 약간 작다. 2010년 연구에서는 ''공자새'' 알의 폭이 비슷한 크기의 현대 조류에서 예상되는 것보다 작다는 것을 보여주었다.[48] 2016년에는 큰 표본의 구멍이 최대 23mm의 알 지름을 나타낸다고 언급했다.[49]2018년 연구에서는 중생대 조류의 알이 몸집에 비해 작았을 것이며, 조류 방식의 접촉 부화(번식을 위해 알 위에 앉는 것)가 불가능했을 것이라고 주장했다.[50] 2007년 연구에서는 ''공자새''가 개방된 둥지에서 알을 품지 않았을 가능성이 높으며, 나무 틈새를 사용했을 수 있다고 주장했다.[47] 2016년 연구에서는 중생대 조류가 알을 땅에 묻었을 것이라고 주장했다.[51]
5. 4. 성장
1998년 푸청 장과 동료들은 주사 전자 현미경을 사용하여 넙다리뼈 횡단면을 분석, 뼈에 혈관이 풍부하고 단일 성장 정지선만 나타나는 것을 확인했다. 이를 통해 ''공자새''의 성장이 빠르고 지속적이며, 온혈동물이었을 것이라고 결론지었다.[52] 뼈 구조가 현대 변온성 앨리게이터와 달리 깃털 달린 비조류 공룡 ''베이피아오사우루스''와 유사하다는 점을 들어 이 주장을 뒷받침했다.[53]2003년 아르망 드 리클레와 동료들은 80개 뼈 박편 연구를 통해 ''공자새''가 20주가 채 안 되어 성체 크기에 도달하며, 성장률은 현대 조류보다 낮지만 비조류 수각류와 유사한 빠른 성장률을 보였다고 밝혔다.[56]
2008년 치아페와 동료들은 106개 표본을 분석하여 신체 크기와 긴 꼬리 깃털 소유 간 관계를 연구했다. 그 결과 성장 급증 직전의 성장 단계에서 많은 개체가 화석화되는 것을 막았을 것이라는 가설을 제시했다.[57]
2009년 빈프리트와 디터 페터스는 더 작은 크기 등급의 신체 크기가 가장 어린 성장 단계를 나타낼 가능성이 높으며, 두 개의 더 큰 크기 등급이 성장 단계가 아닌 두 성별을 나타낸다고 주장했다.[58]
2013년 아누수야 친사미와 동료들은 수질골을 통해 꼬리가 짧은 표본을 암컷, 긴 꼬리 표본을 수컷으로 추정했다. 또한 성적 성숙이 최종 크기에 도달하기 훨씬 전에 발생했을 것이라고 결론 내렸다.[63]
2018년 징마이 오코너와 동료들은 수질골 식별에 의문을 제기하며, 에난티오르니테스(Enantiornithes)의 후지에서 수질골을 확인하고, 기초 조류의 공룡과 같은 성장 방식을 뒷받침했다.[64]
5. 5. 식단
1999년, 중국 고생물학자 Hou Lianhai와 동료들은 부리가 위로 굽어 있고 맹금류가 아니라는 점을 들어 ''공자새''가 초식성일 가능성이 높다고 주장했다.[13] 1999년, 고생물학자 Dieter S. Peters와 Ji Qiang은 발가락 물갈퀴의 잔해가 보존되지는 않았지만, 다소 부드러운 부리를 사용하여 수영하면서 물속에서 먹이를 잡았다고 가설을 세웠다. 2002년, 폴란드 고생물학자 안제이 엘자노프스키는 ''공자새''가 물이나 지면에서 먹이를 잡아 날아다니는 방식으로 먹이를 찾았다고 제안했다.[65]
2003년, 중국 고생물학자 Zhonghe Zhou와 Fucheng Zhang은 튼튼하고 이빨이 없는 턱은 씨앗을 먹었을 수 있음을 시사했다.[66] 2006년, Johan Dalsätt과 동료들은 ''C. sanctus'' 표본(IVPP V13313)에서 작은 물고기, 아마도 ''Jinanichthys''의 뼈를 발견하고 ''공자새''가 잡식성이었을 것이라고 제안했다.[67] 안드레이 지노비예프는 ''공자새''가 날아가면서 물고기를 잡았다고 추정했다.[68]
두개골은 상대적으로 움직이지 않았으며, 현대 조류의 운동성이 없었다.[22]
6. 고환경 및 고생태학
''공자새''(Confuciusornis)는 이셴 층과 지우포탕 층에서 발견되었으며, 제홀 생물군에 속한다.[67] 두 지층은 잦은 화산 폭발로 인해 응회암이 상당량 포함되어 있으며, 이셴 층에서 화산 폭발이 약간 더 빈번했다. 셰일과 이암 역시 이 지층들의 주요 구성 요소이다. 응회암은 40Ar-39Ar 동위원소를 사용하여 지층의 정확한 연대 측정을 가능하게 했으며, 그 결과 이셴 층은 약 1억 2500만 년에서 1억 2000만 년 전, 지우포탕 층은 약 1억 2030만 년 전으로 추정된다.[69] 화석은 홍수와 화산 쇄설물의 결과로 매장되었으며,[70] 화산 지층은 상세한 깃털 자국과 같은 다양한 연조직의 보존을 가능하게 했다. 2010년 연구에서는 지층에서 발견된 파충류 뼈의 산소 동위원소를 사용하여 이셴을 포함한 동아시아의 많은 지층이 서늘한 온대 기후를 가지고 있음을 밝혀냈다. 이셴 층의 평균 기온은 10°C ± 4°C로 추정되었으며, 당시 온대 지역에서 알려진 나무의 일종인 ''크세녹실론''(Xenoxylon)의 화석이 이 지역 전체에서 발견되었다. 또한 악어와 같이 열을 필요로 하는 파충류는 발견되지 않는다.[71]
제홀 식물의 대부분은 이셴 층 하부에서 발견되었다. 이 식물상에는 이끼, 석송류, 속새, 양치류, 씨앗고사리류, 체칸노프스키목, 은행나무목, 베네티테스목, 네토목, 구과식물, 그리고 소수의 속씨식물을 포함한 중생대 식물의 대부분의 그룹이 포함된다. 제홀 생물군에 존재했던 동물군에는 조개류, 복족류, 이매패류, 곤충, 어류, 도롱뇽[70], 포유류, 도마뱀, 코리스토데라류, 익룡, 그리고 공룡(새 포함)이 포함된다.[69] 이 화석들은 예외적으로 잘 보존되어 있으며, 공룡 화석은 종종 섬유질과 깃털 자국을 보존하고 때로는 ''마이크로랩터''(지우포탕 층의 공중 포식자),[72] ''프시타코사우루스''(두 지층 모두에 광범위하게 분포하는 작은 각룡류),[73] 그리고 ''시노사우로프테릭스''(이셴에서 발견된 최초의 공룡 중 하나인 콤프소그나투스류)[74]와 같은 색소까지 보존한다. 제홀 생물군의 다른 깃털 공룡으로는 복부 내용물에서 ''공자새''(Confuciusornis)의 뼈와 함께 발견된 대형 콤프소그나투스류인 ''시노칼리오프테릭스 기가스''(Sinocalliopteryx gigas),[75] 작은 초식성 오비랍토르류인 ''카우디프테릭스''(Caudipteryx),[69] 그리고 대형 티라노사우루스류인 ''유티라누스''(Yutyrannus)가 있는데, 모두 이셴 층에서 발견되었다.[76] 제홀 조류는 기저 조류(''공자새''(Confucisornis), ''제홀오르니스''(Jeholornis), ''사페오르니스''(Sapeornis) 등), 더 파생된 이반새류(''에오에난티오르니스''(Eoenantiornis), ''롱기로스트라비스''(Longirostravis), ''시노르니스''(Sinornis), ''볼루오키아''(Boluochia), ''롱기프테릭스''(Longipteryx) 등), 그리고 더욱 파생된 참새류(''랴오닝오르니스''(Liaoningornis), ''이셴오르니스''(Yixianornis), ''얀오르니스''(Yanornis) 등)를 포함하여 20개 이상의 속으로 대표된다.[69]
참조
[1]
논문
A new confuciusornithid bird with a secondary epiphyseal ossification reveals phylogenetic changes in confuciusornithid flight mode
[2]
서적
The Complete Encyclopedia of Fossils
Rebo Publishers
[3]
논문
Exceptional preservation of soft tissue in a new specimen of ''Eoconfuciusornis'' and its biological implications
[4]
논문
Non-Avian dinosaur fossils from the Lower Cretaceous Jehol Group of western Liaoning, China
[5]
서적
Mesozoic birds: above the heads of dinosaurs
University of California Press
2002
[6]
논문
The discovery of Early Cretaceous birds in China
[7]
논문
''Confuciusornis sanctus'', a new Late Jurassic sauriurine bird from China
[8]
논문
The first 85 million years of avian evolution
1995
[9]
서적
The Jehol Fossils: The Emergence of Feathered Dinosaurs, Beaked Birds and Flowering Plants
https://books.google[...]
Academic Press
2008
[10]
논문
Taphonomy and mass mortality of ''Confuciusornis'' and feathered dinosaurs at the Sihetun and Zhangjiagou sites in western Liaoning, China
[11]
서적
Mesozoic Birds of China
https://paleoglot.or[...]
Phoenix Valley Provincial Aviary of Taiwan
[12]
논문
Basal birds from China: a brief review
[13]
논문
A diapsid skull in a new species of the primitive bird ''Confuciusornis''
[14]
논문
Diversification in an Early Cretaceous avian genus: evidence from a new species of ''Confuciusornis'' from China
[15]
논문
'[Chinese] [A new species of ''Confuciusornis'' from Lower Cretaceous of Jianchung, Liaoning, China]'
[16]
논문
A taxonomical revision of the Confuciusornithiformes (Aves: Pygostylia)
http://www.vertpala.[...]
2019-01-20
[17]
서적
Dinosaurs: the most complete, up-to-date encyclopedia for dinosaur lovers of all ages
https://archive.org/[...]
Random House
[18]
논문
Cranial morphology of the Early Cretaceous bird
2018
[19]
서적
Mesozoic birds: above the heads of dinosaurs
University of California Press
2002-12-05
[20]
논문
On the Preservation of the Beak in Confuciusornis (Aves: Pygostylia)
2019
[21]
논문
New Information on the Keratinous Beak of ''Confuciusornis'' (Aves: Pygostylia) From Two New Specimens
[22]
문서
"Anatomy and systematics of the Confuciusornithidae (Theropoda:Aves) from the Late Mesozoic of northeastern China"
[23]
논문
Scapular orientation in theropods and basal birds, and the origin of flapping flight
http://www.app.pan.p[...]
2006-08-22
[24]
논문
Muβte ''Confuciusornis'' klettern?
[25]
논문
Laser Fluorescence Illuminates the Soft Tissue and Life Habits of the Early Cretaceous Bird Confuciusornis
2016
[26]
논문
Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds
http://oro.open.ac.u[...]
[27]
논문
Trace metals as biomarkers for eumelanin pigment in the fossil record
[28]
논문
Elaborate plumage patterning in a Cretaceous bird
2018-11-02
[29]
논문
''Confuciusornis'' and the early evolution of birds
[30]
논문
The Chinese early bird ''Confuciusornis'' and the paraphyletic status of Sauriurae
[31]
논문
'[Chinese] "New advances in the study of the primitive bird ''Confuciusornis'''
[32]
문서
"Basal bird phylogeny: problems and solutions"
University of California Press
[33]
문서
"The Morphology and Phylogenetic Position of ''Apsaravis ukhaana'' from the Late Cretaceous of Mongolia"
http://hdl.handle.ne[...]
American Museum of Natural History
[34]
논문
Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of ''Yixianornis grabaui''
[35]
논문
Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species
[36]
논문
Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology
[37]
서적
"Flight capability and habits of ''Confuciusornis''"
Peabody Museum of Natural History. Yale University, New Haven
[38]
웹사이트
Senter, 2006 by Felipe Elias - Issuu
https://issuu.com/fe[...]
2022-03-25
[39]
논문
"''Confuciusornis sanctus'' compared to ''Archaeopteryx lithographica''"
[40]
논문
Re-evaluation of the posture and claws of ''Confuciusornis''
[41]
논문
Narrow primary feather rachises in ''Confuciusornis'' and ''Archaeopteryx'' suggest poor flight ability
[42]
논문
Comment on 'Narrow Primary Feather Rachises in ''Confuciusornis'' and ''Archaeopteryx'' Suggest Poor Flight Ability.'
[43]
논문
Comment on "Narrow Primary Feather Rachises in ''Confuciusornis'' and ''Archaeopteryx'' Suggest Poor Flight Ability
[44]
논문
Response to comments on "Narrow Primary Feather Rachises in ''Confuciusornis'' and ''Archaeopteryx'' Suggest Poor Flight Ability
[45]
논문
An exceptionally preserved Lower Cretaceous ecosystem
2003
[46]
논문
Sexual selection in prehistoric animals: detection and implications
2013
[47]
서적
The inner bird: anatomy and evolution
UBC Press
2010
[48]
논문
Cracking a developmental constraint: egg size and bird evolution
2010
[49]
서적
Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs
JHU Press
2016
[50]
논문
Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs
http://eprints.linco[...]
2018
[51]
논문
Reproduction in Mesozoic birds and evolution of the modern avian reproductive mode
2016
[52]
논문
Osteological microstructure of ''Confuciusornis'': preliminary report
[53]
논문
Some microstructure difference among ''Confuciusornis'', ''Alligator'' and a small theropod dinosaur, and its implications
[54]
논문
Growth rings in Mesozoic birds
1994
[55]
논문
Dinosaurian growth rates and bird origins
[56]
논문
Osteohistology of ''Confuciusornis sanctus'' (Theropoda: Aves)
[57]
논문
Life history of a basal bird: morphometrics of the Early Cretaceous ''Confuciusornis''
[58]
논문
Life history, sexual dimorphism and 'ornamental' feathers in the mesozoic bird ''Confuciusornis sanctus''
[59]
논문
Palaeobiology of the Cretaceous bird ''Confuciusornis'': a comment on Peters & Peters (2009)
2010
[60]
논문
Bone growth marks reveal protracted growth in New Zealand kiwi (Aves, Apterygidae)
2009
[61]
논문
Sexual size dimorphism is the most consistent explanation for the body size spectrum of ''Confuciusornis sanctus''
2010
[62]
논문
Quantitative patterns of morphological variation in the appendicular skeleton of the Early Cretaceous bird Confuciusornis
2011
[63]
논문
Gender identification of the Mesozoic bird ''Confuciusornis sanctus''
[64]
논문
Medullary bone in an Early Cretaceous enantiornithine bird and discussion regarding its identification in fossils
2018
[65]
서적
"Biology of basal birds and the origin of avian flight"
Science, Beijing
[66]
논문
Anatomy of the primitive bird ''Sapeornis chaoyangensis'' from the Early Cretaceous of Liaoning, China
2003
[67]
논문
Food remains in ''Confuciusornis sanctus'' suggest a fish diet
[68]
논문
An attempt to reconstruct the lifestyle of confuciusornithids (Aves, Confuciusornithiformes)
[69]
논문
Evolutionary radiation of the Jehol Biota: chronological and ecological perspectives
[70]
논문
The Chinese Pompeii? Death and destruction of dinosaurs in the Early Cretaceous of Lujiatun, NE China
[71]
논문
Oxygen isotopes of East Asian dinosaurs reveal exceptionally cold Early Cretaceous climates
[72]
논문
Reconstruction of ''Microraptor'' and the Evolution of Iridescent Plumage
2012
[73]
논문
3D Camouflage in an Ornithischian Dinosaur
2016
[74]
논문
Countershading and Stripes in the Theropod Dinosaur ''Sinosauropteryx'' Reveal Heterogeneous Habitats in the Early Cretaceous Jehol Biota
2017
[75]
논문
IV, Ji S, Miyashita T, Burns ME, ''et al.'' (2012) Abdominal Contents from Two Large Early Cretaceous Compsognathids (Dinosauria: Theropoda) Demonstrate Feeding on Confuciusornithids and Dromaeosaurids
[76]
논문
A gigantic feathered dinosaur from the Lower Cretaceous of China
http://www.xinglida.[...]
[77]
서적
한 권으로 끝내는 공룡
Gbrain
2013-06-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com