중생대
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
중생대는 약 2억 5100만 년 전부터 6600만 년 전까지 이어진 지질 시대로, '중간 생명'을 의미하며, 고생대와 신생대 사이에 위치한다. 중생대는 트라이아스기, 쥐라기, 백악기의 세 시기로 구분되며, 페름기-트라이아스기 멸종으로 시작해 백악기-제3기 대멸종으로 마무리된다. 중생대에는 공룡을 비롯한 다양한 파충류가 번성했으며, 초대륙 판게아가 분리되어 현재 대륙의 모습으로 변화했다. 기후는 대체로 온난했으며, 겉씨식물이 육상 식물의 주를 이루었다.
더 읽어볼만한 페이지
- 대 (지질학) - 고생대
고생대는 캄브리아기 대폭발부터 페름기 말 대멸종까지, 다양한 무척추동물과 최초의 척추동물, 육상 식물 및 동물의 출현, 그리고 초대륙 판게아 형성과 멸종을 겪은 지질 시대의 중요한 시기이다. - 대 (지질학) - 신생대
신생대는 약 6,600만 년 전부터 현재까지의 지질 시대로, 포유류의 번성, 대륙 이동과 기후 변화, 인류의 출현과 진화를 특징으로 한다. - 중생대 - 영동군
영동군은 충청북도 남부에 위치하며, 소백산맥과 노령산맥이 만나는 지점으로 높은 산지를 이루고 있으며, 포도, 감 등의 특산물과 영동난계국악축제로 유명하다. - 중생대 - 곤드와나
곤드와나는 고생대와 중생대에 존재했던 초대륙으로, 현재의 아프리카, 남아메리카, 남극, 인도, 오스트레일리아 등을 포함했으며, 판게아 분열 이후 서곤드와나와 동곤드와나로 나뉘어 각 대륙이 이동하면서 생물 지리학적 분포 패턴에도 영향을 미쳤다. - 지질학에 관한 - 석탄
석탄은 식물이 지하에서 석탄화 과정을 거쳐 생성된 고체 연료로, 산업 혁명 이후 중요한 에너지원으로 사용되었으며, 탄소 함량에 따라 등급이 나뉘고 화력 발전, 제철 등에 사용되지만, 연소 과정에서 환경 문제를 야기한다. - 지질학에 관한 - 인도-오스트레일리아판
인도-오스트레일리아판은 곤드와나 대륙 분열로 형성되어 인도판과 오스트레일리아판으로 나뉘며, 인도판이 유라시아판과 충돌하여 히말라야 산맥과 티베트 고원을 형성했고, 주변 판들과 복잡한 경계를 이루어 지진과 쓰나미가 자주 발생한다.
| 중생대 | |
|---|---|
| 지도 정보 | |
| 시대 구분 | |
| 상위 시대 | 현생누대 |
| 하위 시대 | 고생대 |
| 시작 시점 | 2억 5190만 2천 년 전 |
| 시작 시점 불확실성 | 2만 4천 년 |
| 종료 시점 | 6600만 년 전 |
| 지질학적 정보 | |
| 시대 단위 | 대 |
| 지층 단위 | 대이층 |
| 시대 형식 | 형식적 |
| 하위 경계 정의 | 코노돈트 Hindeodus parvus의 첫 등장. |
| 하위 경계 GSSP 위치 | 메이샨, 저장성, 중국 |
| 하위 경계 GSSP 승인일 | 2001년 |
| 상위 경계 정의 | 주요 운석 충격 및 후속 K-Pg 멸종 사건과 관련된 이리듐 농축층. |
| 상위 경계 GSSP 위치 | 엘 케프 단면, 엘 케프, 튀니지 |
| 상위 경계 GSSP 승인일 | 1991년 |
| 명칭 정보 | |
| 명칭 형식 | 형식적 |
| 별칭 | 파충류의 시대 공룡의 시대 침엽수의 시대 |
| 사용 정보 | |
| 천체 | 지구 |
| 사용처 | 국제 (국제층서위원회) |
| 사용된 시간 척도 | 국제층서위원회 시간 척도 |
| 정의 | |
| 시간 단위 | 대 |
| 층서 단위 | 대이층 |
| 시간 범위 형식 | 형식적 |
| 하위 경계 정의 | 코노돈트 Hindeodus parvus의 첫 출현. |
| 하위 GSSP 위치 | 메이산, 저장, 중국 |
| 하위 GSSP 승인일 | 2001 |
| 상위 경계 정의 | 대규모 운석 충돌 및 후속 K-Pg 대멸종과 관련된 이리듐 풍부층. |
| 상위 GSSP 위치 | 엘 케프 섹션, 엘 케프, 튀니지 |
| 상위 GSSP 승인일 | 1991 |
2. 명칭
현재의 명칭은 1840년 영국의 지질학자 존 필립스가 제안하였다. "중생대"는 문자 그대로 '중간 생명'을 의미하며, 그리스어 접두사 ('사이')와 ('동물, 생명체')에서 유래한다.[1][2] 이러한 방식으로 중생대는 신생대(Cenozoic|새로운 생명영어)와 고생대('오래된 생명') 그리고 원생누대('이전 생명') 에온과 비교될 수 있다.
중생대는 대략 2억 5100만 년 전부터 6600만 년 전까지의 시기이며, 트라이아스기, 쥐라기, 백악기의 세 시기로 나뉜다.[51]
중생대는 원래 "고생대" 다음에 오는 "제2기"로 묘사되었으며, 제3기에 앞선다.[3]
3. 지질 시대
고생대 말기 페름기와 트라이아스기 사이에 있었던 페름기-트라이아스기 멸종으로 시작되어 백악기와 신생대의 백악기-제3기 대멸종을 겪으며 마무리된다.[49] 페름기-트라이아스기 멸종은 지구 역사상 가장 큰 대량 멸종으로, 바다 생물종의 90%, 육상 생물종의 70%가 전멸하여 "거대한 죽음"이라고도 불린다.[52] 공룡의 멸종과 함께 중생대를 마감하는 백악기-제3기 대멸종에서도 약 50%의 생물종이 절멸되었다.[53]
중생대가 시작되었을 때 모든 육지는 판게아라는 하나의 초대륙으로 뭉쳐 있었다. 판게아는 북쪽의 로라시아와 남쪽의 곤드와나로 갈라졌다. 로라시아는 다시 북아메리카와 유라시아로 나뉘었고, 곤드와나는 남아메리카, 아프리카, 오세아니아, 남극, 인도아대륙으로 갈렸다.
3. 1. 트라이아스기 (Triassic)
고생대 말 페름기와 트라이아스기 사이에 있었던 페름기-트라이아스기 멸종으로 중생대가 시작되었다.[49] 트라이아스기는 약 2억 5200만 년 전부터 2억 100만 년 전까지 지속되었으며, 쥐라기 이전 시대이다. 페름기-트라이아스기 멸종 사건과 트라이아스기-쥐라기 멸종 사건이라는 두 차례의 "5대 멸종" 사건 사이에 위치하며, 초기 트라이아스세, 중기 트라이아스세, 후기 트라이아스세의 세 시대로 구분된다.[8]
약 2억 5200만 년 전부터 2억 4700만 년 전까지의 초기 트라이아스세는 초대륙 판게아 내륙의 사막이 지배적이었다. 지구는 생물의 95%가 멸종하는 대규모 멸종 사건을 겪은 직후였으며, 육상에서 가장 흔한 척추동물은 리스트로사우루스(Lystrosaurus), 유형동물(labyrinthodont), 유파르케리아(Euparkeria)와 페름기 멸종을 극복한 다른 많은 생물들이었다. 템노스폰딜류(Temnospondyl)는 초기 트라이아스기에 다양성의 정점에 달했다.[9]
2억 4700만 년 전부터 2억 3700만 년 전까지의 중기 트라이아스세에는 판게아의 분열과 테티스 해(Tethys Ocean)의 형성이 시작되었다. 생태계는 페름기 멸종에서 회복되었고, 조류, 해면, 산호, 갑각류 등이 회복되었으며, 이크티오사우루스(ichthyosaur)와 노토사우루스(nothosaur)와 같은 새로운 수생 파충류가 진화했다. 육상에서는 소나무 숲이 번성했고, 모기와 초파리와 같은 곤충류도 번성했다. 파충류는 점점 커지기 시작했고, 최초의 악어류와 공룡이 진화하여 이전에 민물 세계를 지배했던 대형 양서류와 육상의 포유류형 파충류와의 경쟁이 시작되었다.[10]
2억 3700만 년 전부터 2억 100만 년 전까지의 후기 트라이아스세에는 잦은 폭염과 중간 정도의 강수량(연간 25cm~50cm)이 특징이었다. 최근의 온난화는 대륙들이 서로 분리되기 시작하면서 육상에서 공룡의 진화 붐을 일으켰고,(2억 4300만 년 전부터 2억 1000만 년 전까지의 니아사사우루스(Nyasasaurus), 대략 2억 3500만 년 전~2억 700만 년 전, 일부는 용각류형류, 수각류, 헤레라사우루스류로 분리) 최초의 익룡(pterosaur)도 나타났다. 후기 트라이아스기에는 일부 진화된 시노돈트(cynodont)에서 최초의 포유류형(Mammaliaformes)이 출현했다. 그러나 이러한 모든 기후 변화는 트라이아스기-쥐라기 멸종 사건으로 알려진 대규모 멸종으로 이어졌는데, 이 사건으로 많은 악어형류(archosaurs)(익룡, 공룡, 악어형류(crocodylomorph) 제외), 대부분의 시냅시드(synapsid), 그리고 거의 모든 대형 양서류가 멸종했고, 해양 생물의 34%가 멸종하면서 지구의 네 번째 대멸종이 발생했다. 그 원인은 논란의 여지가 있지만,[11][12] 중앙 대서양 마그마 지역(Central Atlantic magmatic province)에서 일어난 홍수 현무암 분출(flood basalt eruption)이 그 원인 중 하나로 제시되고 있다.[13][14][15]
3. 2. 쥐라기 (Jurassic)

쥐라기는 2억 년 전부터 1억 4500만 년 전까지의 시대이며, 초기 쥐라기, 중기 쥐라기, 후기 쥐라기의 세 가지 주요 시대로 구분된다.[16]
초기 쥐라기는 2억 년 전부터 1억 7500만 년 전까지의 시대이다.[16] 기후는 열대성이었고 트라이아스기보다 훨씬 습했는데, 이는 육지 사이에 거대한 바다가 출현했기 때문이다. 바다에서는 플레시오사우루스류, 어룡, 그리고 암모나이트가 풍부했다. 육지에서는 공룡과 다른 악어형류가 지배적인 종으로 자리 잡았고, ''딜로포사우루스(Dilophosaurus)''와 같은 수각류가 먹이 사슬의 정점에 있었다. 최초의 진정한 악어가 진화하여 대형 양서류를 거의 멸종 직전까지 몰아넣었다. 전반적으로 악어형류가 세계를 지배하게 되었다. 한편, 최초의 진정한 포유류가 진화했는데, 크기는 비교적 작았지만 널리 퍼져 있었다. 예를 들어, 쥐라기의 ''카스토로카우다(Castorocauda)''는 수영, 굴착, 물고기 잡이에 적응한 특징을 가지고 있었다. 약 1억 5000만 년 전 후기 쥐라기 시대의 ''프루이타포소르(Fruitafossor)''는 다람쥐 정도의 크기였으며, 치아, 앞다리, 등의 구조는 사회성 곤충(아마도 흰개미일 것으로 추정되며, 개미는 아직 나타나지 않았다)의 둥지를 파헤쳤음을 시사한다. ''볼라티코테리움(Volaticotherium)''는 현대의 청설모처럼 짧은 거리를 활강할 수 있었다. ''루고소돈(Rugosodon)''과 같은 최초의 다구치류가 진화했다.
중기 쥐라기는 1억 7500만 년 전부터 1억 6300만 년 전까지의 시대이다.[16] 이 시대에는 공룡이 번성하여 ''브라키오사우루스(Brachiosaurus)''와 ''디플로도쿠스(Diplodocus)''와 같은 거대한 용각류의 무리가 양치식물 초원을 가득 채웠고, ''알로사우루스(Allosaurus)''와 같은 많은 새로운 포식자들이 이들을 사냥했다. 침엽수림이 숲의 큰 부분을 차지했다. 바다에서는 플레시오사우루스류가 매우 흔했고, 어룡도 번성했다. 이 시대는 파충류의 전성기였다.[17]
후기 쥐라기는 1억 6300만 년 전부터 1억 4500만 년 전까지의 시대이다.[16] 이 시대에는 ''시조새(Archaeopteryx)''와 같은 최초의 아비알라에가 작은 수각류 공룡으로부터 진화했다. 해수면 상승으로 대서양 해로가 열렸고, 오늘날까지 계속해서 확장되었다. 대륙의 추가적인 분리는 새로운 공룡의 다양화를 위한 기회를 제공했다.
3. 3. 백악기 (Cretaceous)
백악기는 중생대에서 가장 긴 기간이지만, 초기 백악기와 후기 백악기의 두 시기로만 나뉜다.[18]
초기 백악기는 1억 4,500만 년 전부터 1억 년 전까지이다.[18] 초기 백악기에는 해로가 확장되었고, 용각류, 스테고사우루스류와 같이 고지대에서 먹이를 찾는 공룡 무리의 다양성이 감소했다. 특히 북아메리카에서는 용각류가 드물었다.[19] 섬을 건너다니는 일부 공룡(예: 유스트렙토스폰딜루스)은 고대 유럽의 해안 천해와 작은 섬에 적응하여 진화했다. 쥐라기-백악기 멸종으로 남은 빈 공간을 채우기 위해 카르카로돈토사우루스와 스피노사우루스와 같은 다른 공룡들이 나타났다. 계절이 다시 나타나고 극지방은 계절적으로 더 추워졌지만, 레엘리나사우라와 무타부라사우루스와 같이 일부 공룡은 극지방 숲에 연중 서식했다. 극지방은 악어에게는 너무 추웠고, 쿨라수쿠스와 같은 대형 양서류의 마지막 보루가 되었다. 타페자라와 오르니토케이루스와 같은 속이 진화하면서 익룡은 더 커졌다. 포유류는 서식 범위를 계속 확장했다. 유트리코노돈트류는 레페노마무스와 고비코돈과 같은 상당히 크고 족제비와 같은 포식자를 낳았고, 초기 수궁류는 후대류와 진수류로 확장하기 시작했으며, 치몰로돈트 다람쥐류는 화석 기록에서 흔해졌다.
후기 백악기는 1억 년 전부터 6,600만 년 전까지이다. 후기 백악기에는 신생대까지 계속될 냉각 경향이 나타났다. 결국 열대 지방은 적도로 제한되었고 열대 지방을 벗어난 지역에서는 극심한 계절 변화를 겪었다. 티라노사우루스, 안킬로사우루스, 트리케라톱스 및 하드로사우루스류와 같은 새로운 분류군이 먹이 사슬을 지배하면서 공룡은 여전히 번성했다. 바다에서는 모사사우루스류가 지배하여 쇠퇴한 후 세노마니안-투로니안 경계 사건에서 사라진 어룡의 역할을 대신했다. 같은 사건에서 플리오사우루스류가 멸종했지만, 엘라스모사우루스와 같은 긴 목의 플레시오사우루스류는 계속 번성했다. 트라이아스기까지 거슬러 올라갈 수도 있는 속씨식물이 처음으로 실제로 우점종이 되었다. 후기 백악기의 익룡은 이유는 잘 알려져 있지 않지만 쇠퇴했지만, 이는 화석 기록의 경향 때문일 수도 있다. 왜냐하면 그 다양성은 이전에 생각했던 것보다 훨씬 높은 것으로 보이기 때문이다. 조류는 점점 더 흔해졌고 다양한 역조류와 조류 형태로 분화되었다. 대부분 작았지만, 해양 헤스페로르니테스류는 비교적 크고 날지 못하는 형태로 진화하여 공해에서의 생활에 적응했다. 후대류와 원시 진수류도 흔해졌고, 디델포돈과 쇼왈테리아와 같은 크고 특수화된 속을 낳았다. 그럼에도 불구하고 우점 포유류는 다람쥐류였으며, 북쪽에는 치몰로돈트류가, 남쪽에는 곤드와나테레류가 있었다. 백악기 말기에 데칸 트랩과 기타 화산 폭발로 대기가 오염되었다. 이것이 계속됨에 따라, 6,600만 년 전에 거대한 운석이 지구에 충돌하여 칙술루브 크레이터를 생성한 것으로 여겨진다. 이 사건은 K-Pg 멸종(이전의 K-T 멸종)으로 알려져 있으며, 5번째이자 가장 최근의 대량 멸종 사건으로, 비조류 공룡을 포함한 생명체의 75%가 멸종되었다.[20][21][22]
4. 초대륙의 분열
고생대 말 지구의 대륙들은 서로 연결되어 하나의 거대한 대륙인 판게아를 이루고 있었다. 그러나 중생대에 들어 판게아가 분리되기 시작하였고 쥐라기 초중기인 1억 7500만 년 전쯤 남북으로 분열되어 로라시아와 곤드와나로 나뉘게 되었고 분열된 두 대륙 사이에 바다가 들어서게 되었다.[54]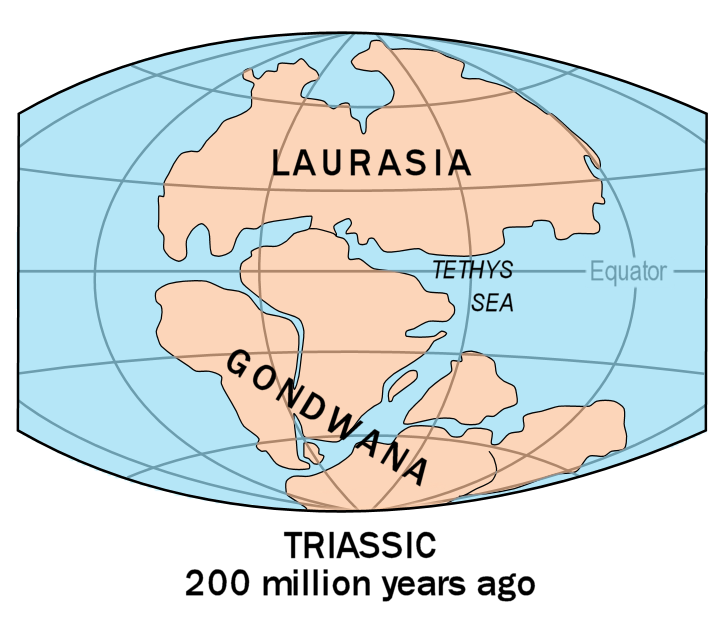
후기 고생대의 활발한 수렴판 조산 작용과 비교하여 중생대의 지각 변형은 상대적으로 온화했다. 중생대의 유일한 주요 조산 운동은 현재 북극해에 해당하는 지역에서 발생하여 이누이티아 조산대, 브룩스 산맥, 시베리아의 베르호얀스크 산맥과 체르스키 산맥, 그리고 만주 지역의 흥안령을 형성했다.
이 조산 운동은 북극해의 열림과 중국 북부와 시베리아 지괴가 아시아에 봉합되는 것과 관련이 있다.[23] 반대로, 이 시대는 초대륙 판게아의 극적인 분리를 특징으로 하며, 점차 북쪽 대륙인 로라시아와 남쪽 대륙인 곤드와나로 분열되었다. 이는 오늘날 대서양 연안(예: 미국 동부 해안)의 대부분을 특징짓는 대륙주변부를 생성했다.[24]
이 시대 말까지 대륙들은 거의 현재의 형태로 분리되었지만, 현재의 위치는 아니었다. 로라시아는 북아메리카와 유라시아가 되었고, 곤드와나는 남아메리카, 아프리카, 오스트레일리아, 남극 대륙 및 인도 아대륙으로 분열되었으며, 인도 아대륙은 신생대에 아시아판과 충돌하여 히말라야 산맥을 형성했다.
5. 기후
고생대와 신생대에 빙하기가 주기적으로 나타났던 것과 달리, 판게아가 분열된 뒤로 해안선이 길어지면서 대륙에 고온 다습한 지역이 늘어 겉씨식물이 출현하는 등 중생대 동안에는 빙하기가 발생하지 않아 일반적으로 온도가 높았다.[55][56]
트라이아스기는 일반적으로 건조했으며, 특히 판게아 내부에서는 계절 변화가 컸다. 낮은 해수면 또한 기온 변화를 심화시켰을 것이고, 내륙에는 광대한 사막이 있었을 가능성이 높다. 풍부한 적색층과 할라이트와 같은 증발암이 이러한 결론을 뒷받침하지만, 일부 증거는 트라이아스기의 일반적으로 건조한 기후가 강우량 증가의 시기로 중단되었음을 시사한다.[25] 가장 중요한 습윤 시기는 카르니아절 홍수 사건과 트라이아스기-쥐라기 멸종 사건 수백만 년 전의 레티아절의 시기였다.
쥐라기 동안 해저 확장의 증가로 해수면이 상승하기 시작했다. 지표면 아래 새로운 지각의 형성은 해양의 물을 현재 해수면보다 최대 200m까지 끌어올려 해안 지역을 침수시켰다. 게다가, 판게아는 더 작은 대륙으로 분열하기 시작하여 테티스 해 주변에 새로운 해안선을 만들었다. 기온은 계속 상승하다가 안정되기 시작했다. 물의 근접성으로 습도도 증가했고, 사막은 후퇴했다.[26]
백악기의 기후는 확실하지 않고 논쟁의 여지가 많다. 아마도 대기 중 이산화탄소의 높은 수준이 남북 기온 경도를 거의 없애버렸을 것으로 생각된다. 전 세계의 기온은 거의 같았고, 현재보다 약 10℃ 높았다. 심해로의 산소 순환 또한 방해받았을 가능성이 있으며, 많은 양의 유기물의 분해를 방지하여 결국 "흑색 셰일"로 퇴적되었다.[27][28]
중생대의 여러 시기에 대기 중 산소량에 대한 연구 결과는 다양하다.
6. 대한민국
한국에는 평안 누층군의 상부 지층이 중생대 초 트라이아스기에 퇴적되었다.
6. 1. 경상 분지
한국에는 평안 누층군의 상부 지층이 중생대 초 트라이아스기에 퇴적되었으며, 경상 분지는 백악기에 형성된 남한 최대의 퇴적 분지이다.6. 2. 한반도의 공룡
경상 분지는 백악기에 형성된 남한 최대의 퇴적 분지이다.7. 생물상

중생대는 이전 시기인 고생대와 이후 시기인 신생대에 빙하기가 주기적으로 나타났던 것과 달리, 전반적으로 온도가 높았다.[55] 초대륙 판게아가 분열되면서 해안선이 길어져 고온 다습한 지역이 늘어났고, 이는 육상 식물이 널리 퍼지는 데 기여했다.[56]
트라이아스기에는 판게아 내륙의 사막이 지배적이었고, 페름기-트라이아스기 멸종 이후 리스트로사우루스, 유형동물, 유파르케리아 등이 육상에서 가장 흔한 척추동물이었다. 템노스폰딜류는 이 시기에 다양성이 정점에 달했다.[9] 중기 트라이아스기에는 판게아의 분열과 테티스 해의 형성이 시작되면서 생태계가 회복되었고, 이크티오사우루스와 노토사우루스와 같은 새로운 수생 파충류가 진화했다. 육상에서는 소나무 숲과 곤충류가 번성했으며, 파충류는 점점 커지기 시작했고, 최초의 악어류와 공룡이 진화하여 이전의 지배적인 동물들과 경쟁했다.[10]
후기 트라이아스기에는 잦은 폭염과 온난화로 인해 육상에서 공룡의 진화 붐이 일어났고, 최초의 익룡도 나타났다. 일부 진화된 키노돈트에서 최초의 포유류형이 출현했다. 그러나 트라이아스기-쥐라기 멸종 사건으로 많은 파충류와 대형 양서류가 멸종했다.[11][12]
쥐라기에는 기후가 열대성이었고 트라이아스기보다 훨씬 습했다. 바다에서는 플레시오사우루스류, 어룡, 암모나이트가 풍부했다. 육지에서는 딜로포사우루스와 같은 수각류 공룡과 다른 파충류가 지배적인 종으로 자리 잡았고, 최초의 진정한 악어가 진화하여 대형 양서류를 거의 멸종 직전까지 몰아넣었다. 한편, 최초의 진정한 포유류가 진화하여 널리 퍼져 있었다. 카스토로카우다는 수영, 굴착, 물고기 잡이에 적응했고, 프루이타포소르는 흰개미 둥지를 파헤쳤으며, 볼라티코테리움는 짧은 거리를 활강할 수 있었다. 루고소돈과 같은 최초의 다구치류도 진화했다. 중기 쥐라기에는 브라키오사우루스와 디플로도쿠스와 같은 거대한 용각류 공룡과 알로사우루스와 같은 포식자들이 번성했다. 침엽수림이 숲의 큰 부분을 차지했고, 바다에서는 플레시오사우루스류와 어룡이 번성했다.[17] 후기 쥐라기에는 시조새와 같은 최초의 조류가 작은 수각류 공룡으로부터 진화했고, 해수면 상승으로 대서양 해로가 열렸다.
백악기 초기에는 해로가 확장되고 용각류와 스테고사우루스류의 다양성이 감소했다. 카르카로돈토사우루스와 스피노사우루스와 같은 새로운 공룡들이 나타났고, 레엘리나사우라와 무타부라사우루스와 같이 일부 공룡은 극지방 숲에 서식했다. 타페자라와 오르니토케이루스와 같은 속이 진화하면서 익룡은 더 커졌고, 포유류는 서식 범위를 계속 확장했다. 유트리코노돈트류는 레페노마무스와 고비코돈과 같은 포식자를 낳았고, 초기 수궁류는 후대류와 진수류로 확장했으며, 치몰로돈트 다구치류는 화석 기록에서 흔해졌다.
후기 백악기에는 냉각 경향이 나타났고, 티라노사우루스, 안킬로사우루스, 트리케라톱스 및 하드로사우루스류와 같은 새로운 분류군이 먹이 사슬을 지배하면서 공룡은 여전히 번성했다. 바다에서는 모사사우루스류가 지배하여 어룡의 역할을 대신했고, 플리오사우루스류는 멸종했지만, 엘라스모사우루스와 같은 긴 목의 플레시오사우루스류는 계속 번성했다. 속씨식물이 처음으로 우점종이 되었고, 조류는 다양하게 분화되었다. 대부분 작았지만, 해양 헤스페로르니테스류는 비교적 크고 날지 못하는 형태로 진화했다. 후대류와 원시 진수류도 흔해졌고, 디델포돈과 쇼왈테리아와 같은 크고 특수화된 속을 낳았다. 그럼에도 불구하고 우점 포유류는 다구치류였다. 백악기 말기에 데칸 트랩과 기타 화산 폭발로 대기가 오염되었고, 6,600만 년 전에 거대한 운석이 지구에 충돌하여 칙술루브 크레이터를 생성한 것으로 여겨지는 백악기-팔레오기 멸종 사건으로 비조류 공룡을 포함한 생명체의 75%가 멸종했다.[20][21][22]
페름기 말 대멸종 이후, 파레이아사우루스류와 고르고놉시아류의 멸종으로 생긴 빈 생태적 지위를 채우며 다양한 생명체가 방산하게 되었다. 살아남은 키노돈트류와 디키노돈트류도 있었지만, 후자는 멸종했다.
복잡한 생태계가 재정립되는 데는 오랜 시간이 걸렸다. 멸종 후 400만 년에서 600만 년 후인 트라이아스기 중기에 시작되어,[40] 3천만 년이 지나서야 완전히 번성했다.[41] 이 시기 동물 생태계는 공룡, 익룡, 그리고 어룡, 플레시오사우루스, 모사사우루스류와 같은 수생 파충류 등 다양한 원시 파충류가 지배했다.
쥐라기 후기와 백악기의 기후 변화는 추가적인 적응 방산을 촉진했다. 쥐라기는 원시 파충류 다양성이 절정에 달했던 시기였으며, 최초의 조류와 진수류 포유류도 나타났다.
트라이아스기에 파충류에서 공룡이 출현하였으며, 쥐라기에는 공룡이 번성하고, 조류가 출현하였다. 백악기에도 공룡의 번영은 계속되었으나, 백악기 말(K-T 경계, K-Pg 경계)에 지구 규모의 대량 멸종이 일어나 조류를 제외한 공룡류는 멸종했다.(단, 알라모사우루스 등의 일부 속은 이를 극복하고 살아남았을 가능성이 있음[47]).
7. 1. 식물상
중생대는 1억 8000만 년에 걸친 긴 시간으로 기후 역시 다양한 변화를 겪었다. 그러나 일반적으로 보아 온도가 높았다고 말할 수 있는데, 이전 시기인 고생대와 이후 시기인 신생대에 빙하기가 주기적으로 나타났던 것과 달리 중생대 동안에는 발생하지 않았기 때문이다.[55] 또한, 초대륙 판게아가 분열된 뒤로 해안선이 길어지면서 대륙에 고온 다습한 지역이 늘었다. 이로 인해 육상 식물이 널리 퍼질 수 있는 조건이 마련되었고, 겉씨식물이 출현하였다.[56]당시 육상 식물의 우점종은 겉씨식물(gymnosperm영어)이었다. 겉씨식물은 관다발을 가지고 있으며, 구과를 맺고, 꽃이 피지 않는 식물로, 솔잎나무와 같이 씨앗에 껍질이 없는 식물이다. 이는 현재 지구의 식물상과 대조되는데, 현재는 종의 수를 기준으로 볼 때 속씨식물(angiosperm영어)이 육상 식물의 우점종이다. 은행나무속(*Ginkgo*)의 가장 오래된 종은 중기 쥐라기에 처음 등장했다. 이 속은 오늘날 단일 종인 *Ginkgo biloba* 은행나무로 대표된다.[36] 현대 침엽수 그룹은 쥐라기 동안 다양화되기 시작했다.[37] 벤네티테스목(Bennettitales)은 멸종된 겉씨식물 그룹으로, 외관상으로는 소철과 비슷한 잎을 가지고 있으며, 후기 트라이아스기 동안 전 세계적으로 분포하여 중생대 종자식물에서 가장 흔한 그룹 중 하나였다.[38]
속씨식물은 초기 백악기에, 처음에는 열대 지방에서 다양화되었지만, 고른 온도 기울기 덕분에 백악기 전체에 걸쳐 극지방으로 퍼져나갈 수 있었다. 백악기 말까지 속씨식물은 많은 지역에서 나무 식물상을 지배하게 되었지만, 일부 증거에 따르면 생물량은 백악기-팔레오기 멸종 이후까지도 소철과 양치류가 여전히 지배하고 있었던 것으로 보인다. 일부 식물 종들은 이후 시대와는 현저히 다른 분포를 보였는데, 예를 들어 양치류 목인 쉬자목(Schizeales)은 중생대에는 북반구에 치우쳐 있었지만, 현재는 남반구에 더 많이 분포한다.[39]
7. 2. 동물상
트라이아스기에는 초대륙 판게아 내륙의 사막이 지배적이었고, 페름기-트라이아스기 멸종 이후 리스트로사우루스, 유형동물, 유파르케리아 등이 육상에서 가장 흔한 척추동물이었다. 템노스폰딜류는 이 시기에 다양성이 정점에 달했다.[9] 중기 트라이아스기에는 판게아의 분열과 테티스 해의 형성이 시작되었고, 생태계가 회복되면서 다양한 생물들이 번성했다. 이크티오사우루스와 노토사우루스와 같은 새로운 수생 파충류가 진화했고, 육상에서는 소나무 숲과 곤충류가 번성했다. 파충류는 점점 커지기 시작했고, 최초의 악어류와 공룡이 진화하여 이전의 지배적인 동물들과 경쟁했다.[10]후기 트라이아스기에는 잦은 폭염과 온난화로 인해 육상에서 공룡의 진화 붐이 일어났고, 최초의 익룡도 나타났다. 일부 진화된 키노돈트에서 최초의 포유류형이 출현했다. 그러나 트라이아스기-쥐라기 멸종 사건으로 많은 파충류와 대형 양서류가 멸종했다.[11][12]
쥐라기에는 기후가 열대성이었고 트라이아스기보다 훨씬 습했다. 바다에서는 플레시오사우루스류, 어룡, 암모나이트가 풍부했다. 육지에서는 딜로포사우루스와 같은 수각류 공룡과 다른 파충류가 지배적인 종으로 자리 잡았고, 최초의 진정한 악어가 진화하여 대형 양서류를 거의 멸종 직전까지 몰아넣었다. 한편, 최초의 진정한 포유류가 진화하여 널리 퍼져 있었다. 카스토로카우다는 수영, 굴착, 물고기 잡이에 적응했고, 프루이타포소르는 흰개미 둥지를 파헤쳤으며, 볼라티코테리움는 짧은 거리를 활강할 수 있었다. 루고소돈과 같은 최초의 다구치류도 진화했다. 중기 쥐라기에는 브라키오사우루스와 디플로도쿠스와 같은 거대한 용각류 공룡과 알로사우루스와 같은 포식자들이 번성했다. 침엽수림이 숲의 큰 부분을 차지했고, 바다에서는 플레시오사우루스류와 어룡이 번성했다.[17] 후기 쥐라기에는 시조새와 같은 최초의 조류가 작은 수각류 공룡으로부터 진화했고, 해수면 상승으로 대서양 해로가 열렸다.
백악기 초기에는 해로가 확장되고 용각류와 스테고사우루스류의 다양성이 감소했다. 카르카로돈토사우루스와 스피노사우루스와 같은 새로운 공룡들이 나타났고, 레엘리나사우라와 무타부라사우루스와 같이 일부 공룡은 극지방 숲에 서식했다. 타페자라와 오르니토케이루스와 같은 속이 진화하면서 익룡은 더 커졌고, 포유류는 서식 범위를 계속 확장했다. 유트리코노돈트류는 레페노마무스와 고비코돈과 같은 포식자를 낳았고, 초기 수궁류는 후대류와 진수류로 확장했으며, 치몰로돈트 다구치류는 화석 기록에서 흔해졌다.
후기 백악기에는 냉각 경향이 나타났고, 티라노사우루스, 안킬로사우루스, 트리케라톱스 및 하드로사우루스류와 같은 새로운 분류군이 먹이 사슬을 지배하면서 공룡은 여전히 번성했다. 바다에서는 모사사우루스류가 지배하여 어룡의 역할을 대신했고, 플리오사우루스류는 멸종했지만, 엘라스모사우루스와 같은 긴 목의 플레시오사우루스류는 계속 번성했다. 속씨식물이 처음으로 우점종이 되었고, 조류는 다양하게 분화되었다. 대부분 작았지만, 해양 헤스페로르니테스류는 비교적 크고 날지 못하는 형태로 진화했다. 후대류와 원시 진수류도 흔해졌고, 디델포돈과 쇼왈테리아와 같은 크고 특수화된 속을 낳았다. 그럼에도 불구하고 우점 포유류는 다구치류였다. 백악기 말기에 데칸 트랩과 기타 화산 폭발로 대기가 오염되었고, 6,600만 년 전에 거대한 운석이 지구에 충돌하여 칙술루브 크레이터를 생성한 것으로 여겨지는 백악기-팔레오기 멸종 사건으로 비조류 공룡을 포함한 생명체의 75%가 멸종했다.[20][21][22]
7. 2. 1. 공룡
페름기 말 대멸종 이후, 파레이아사우루스류와 고르고놉시아류의 멸종으로 생긴 빈 생태적 지위를 채우며 다양한 생명체가 방산하게 되었다. 살아남은 키노돈트류와 디키노돈트류도 있었지만, 후자는 멸종했다.
복잡한 생태계가 재정립되는 데는 오랜 시간이 걸렸다. 멸종 후 400만 년에서 600만 년 후인 트라이아스기 중기에 시작되어,[40] 3천만 년이 지나서야 완전히 번성했다.[41] 이 시기 동물 생태계는 공룡, 익룡, 그리고 어룡, 플레시오사우루스, 모사사우루스류와 같은 수생 파충류 등 다양한 원시 파충류가 지배했다.
쥐라기 후기와 백악기의 기후 변화는 추가적인 적응 방산을 촉진했다. 쥐라기는 원시 파충류 다양성이 절정에 달했던 시기였으며, 최초의 조류와 진수류 포유류도 나타났다.
트라이아스기에 파충류에서 공룡이 출현하였으며, 쥐라기에는 공룡이 번성하고, 조류가 출현하였다. 백악기에도 공룡의 번영은 계속되었으나, 백악기 말(K-T 경계, K-Pg 경계)에 지구 규모의 대량 멸종이 일어나 조류를 제외한 공룡류는 멸종했다.(단, 알라모사우루스 등의 일부 속은 이를 극복하고 살아남았을 가능성이 있음[47]).
7. 2. 2. 기타 파충류
페름기 말 대멸종 이후, 동물 생태계는 다양한 원시 파충류에 의해 지배되었습니다. 여기에는 공룡, 익룡과 어룡, 플레시오사우루스류, 모사사우루스류와 같은 수생 파충류가 포함됩니다.[40][41]7. 2. 3. 포유류
페름기 말 대량 멸종 이후, 여러 생물들이 방산을 통해 다양하게 진화했다. 특히, 대형 초식성 파레이아사우루스류와 육식성 고르고놉시아류가 사라진 자리를 키노돈트류와 디키노돈트류 등이 채웠으나, 디키노돈트류는 이후 멸종했다.복잡한 생태계가 다시 형성되기까지는 오랜 시간이 걸렸다. 멸종 후 400만~600만 년 뒤인 트라이아스기 중기에 시작되어,[40] 3천만 년이 지나서야 완전히 번성했다.[41] 이 시기에는 공룡, 익룡, 어룡, 플레시오사우루스, 모사사우루스류 등 다양한 파충류가 번성했다.
쥐라기 후기와 백악기의 기후 변화는 생물들의 추가적인 적응 방산을 이끌었다. 쥐라기는 파충류 다양성이 정점에 달했던 시기였으며, 최초의 조류와 진수류 포유류가 나타났다.
7. 2. 4. 조류
쥐라기는 원시 파충류의 다양성이 절정에 달했던 시기이며, 최초의 조류가 나타난 시기이다.[41]8. 멸종
페름기-트라이아스기 대멸종은 고생대 말 페름기와 트라이아스기 사이에 일어난 지구 역사상 가장 큰 대멸종으로, "거대한 죽음"이라고도 불린다. 이 멸종으로 바다 생물종의 90%, 육상 생물종의 70%가 사라졌다.[52]
백악기-팔레오기 멸종(K-Pg 멸종)은 공룡의 멸종과 함께 중생대를 마감하는 사건으로, 약 50%의 생물종이 절멸되었다.[53]
8. 1. 트라이아스기-쥐라기 멸종
트라이아스기 말, 약 2억 100만 년 전에 발생한 트라이아스기-쥐라기 멸종은 지구 역사상 네 번째 대멸종 사건이다.[11] 이 멸종으로 익룡, 공룡, 악어형류(crocodylomorph)를 제외한 많은 악어형류(archosaurs), 대부분의 시냅시드(synapsid), 그리고 거의 모든 대형 양서류가 멸종했다.[11] 또한, 해양 생물의 34%가 멸종했다.[11]이 멸종의 원인에 대해서는 논란이 있지만,[11][12] 중앙 대서양 마그마 지역(Central Atlantic magmatic province)에서 일어난 홍수 현무암 분출(flood basalt eruption)이 원인 중 하나로 제시되고 있다.[13][14][15]
8. 2. 백악기-팔레오기 멸종 (K-Pg 멸종)
백악기 말, K-T 경계 또는 K-Pg 경계라고 불리는 시기에 지구 규모의 대량 멸종이 일어났다. 이 멸종으로 조류를 제외한 모든 공룡이 멸종했다. 다만, 알라모사우루스와 같은 일부 속은 멸종을 이겨내고 살아남았을 가능성도 있다.[47] 백악기-팔레오기 멸종 사건은 유카탄 반도에 칙술루브 크레이터를 만든 소행성 충돌이 원인일 수 있다.[6] 후기 백악기에는 대규모 화산 폭발도 멸종에 영향을 주었을 것으로 여겨진다.[7] 이 멸종으로 모든 속의 약 50%가 사라졌다.[53]참조
[1]
서적
Penny Cyclopaedia of the Society for the Diffusion of Useful Knowledge
Charles Knight and Co.
1840
[1]
서적
Bulletin 769: The Geologic Time Classification of the United States Geological Survey Compared With Other Classifications, accompanied by the original definitions of era, period and epoch terms
https://books.google[...]
U.S. Government Printing Office
1925
[2]
온톨로지
Mesozoic
[3]
웹사이트
Mesozoic Era
https://www.britanni[...]
Encyclopædia Britannica
2019-09-05
[4]
서적
When life nearly died: the greatest mass extinction of all time
Thames & Hudson
2005
[5]
학술지
Estimates of the magnitudes of major marine mass extinctions in earth history
National Academy of Sciences
2016-10-03
[6]
서적
A Geologic Time Scale 2004
https://www.cambridg[...]
Cambridge University Press
2005
[7]
학술지
End-Cretaceous extinction in Antarctica linked to both Deccan volcanism and meteorite impact via climate change
https://www.research[...]
2016
[8]
백과사전
Triassic Period
https://www.britanni[...]
2024-01-31
[9]
학술지
Calibrated Diversity, Tree Topology and the Mother of Mass Extinctions: The Lesson of Temnospondyls
2008-11
[10]
웹사이트
Middle Triassic
http://palaeos.com/m[...]
[11]
백과사전
Impact Crises and Mass Extinctions: A Working Hypothesis
https://books.google[...]
Geological Society of America
1996
[12]
웹사이트
Late Triassic life
http://www.enchanted[...]
[13]
학술지
Anthropogenic-scale CO2 degassing from the Central Atlantic Magmatic Province as a driver of the end-Triassic mass extinction
https://www.scienced[...]
2022-02
[14]
학술지
Deep CO2 in the end-Triassic Central Atlantic Magmatic Province
2020-04-07
[15]
학술지
Platinum-group elements link the end-Triassic mass extinction and the Central Atlantic Magmatic Province
https://www.nature.c[...]
2020-02-26
[16]
백과사전
Jurassic Era
https://www.britanni[...]
2024-02-07
[17]
웹사이트
Middle Jurassic
http://www.enchanted[...]
[18]
백과사전
Cretaceous
https://www.britanni[...]
[19]
학술지
Diversity patterns amongst herbivorous dinosaurs and plants during the Cretaceous: implications for hypotheses of dinosaur/angiosperm co-evolution
2009-03
[20]
학술지
Repeated Blows
2002
[21]
웹사이트
Cretaceous
http://www.ucmp.berk[...]
[22]
웹사이트
K-T Extinction event
http://www.universet[...]
2015-02-03
[23]
학술지
The case for creation of the North Pacific Ocean during the Mesozoic Era
https://www.scienced[...]
1975-08
[24]
서적
Earth System History
W.H. Freeman and Company
1999
[25]
학술지
Triassic climates – State of the art and perspectives
https://www.scienced[...]
2010-04-15
[26]
학술지
Geographic range of plants drives long-term climate change
https://www.nature.c[...]
2024-02-28
[27]
학술지
Oceanic anoxic events and plankton evolution: Biotic response to tectonic forcing during the mid-Cretaceous: OCEANIC ANOXIC EVENTS AND PLANKTON EVOLUTION
2002-09
[28]
학술지
Cretaceous oceanic anoxic event 2 triggered by a massive magmatic episode
https://www.research[...]
2008-07-17
[29]
웹사이트
Oxygen and Evolution
https://www.science.[...]
2007-04-27
[30]
학술지
GEOCARBSULF: a combined model for Phanerozoic atmospheric O2 and CO2
http://rspb.royalsoc[...]
2006
[31]
학술지
Phanerozoic atmospheric oxygen: New results using the GEOCARBSULF model
http://www.ajsonline[...]
2009
[32]
학술지
A new model for atmospheric oxygen over phanerozoic time
http://rspb.royalsoc[...]
1989
[33]
웹사이트
Phanerozoic Eon
https://web.archive.[...]
2013-04-27
[34]
학술지
Phanerozoic concentrations of atmospheric oxygen reconstructed from sedimentary charcoal
2010
[35]
학술지
COPSE: a new model of biogeochemical cycling over Phanaerozoic time
http://rspb.royalsoc[...]
2004
[36]
웹사이트
Mesozoic Plants
http://www.fossilnew[...]
fossilnews.com
2000-07-28
[37]
논문
An overview of extant conifer evolution from the perspective of the fossil record
http://doi.wiley.com[...]
2018-09-01
[38]
논문
Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group
2021-03-26
[39]
웹사이트
Fern
http://www.eoearth.o[...]
Encyclopedia of Earth. National council for Science and the Environment
2011-11-09
[40]
논문
Timing of recovery from the end-Permian extinction: Geochronologic and biostratigraphic constraints from south China
2006-12-01
[41]
논문
Recovery from the most profound mass extinction of all time
2008-01-01
[42]
논문
Declining metal availability in the Mesozoic seawater reflected in phytoplankton succession
https://www.nature.c[...]
2022-10-31
[43]
기타
[44]
기타
[45]
뉴스
[46]
웹사이트
INTERNATIONAL CHRONOSTRATIGRAPHIC CHART (国際年代層序表)
http://www.geosociet[...]
日本地質学会
2014-03-19
[47]
뉴스
読売新聞
2011-02-05
[48]
웹사이트
INTERNATIONAL CHRONOSTRATIGRAPHIC CHART (国際年代層序表)
http://www.geosociet[...]
日本地質学会
2014-03-19
[49]
서적
화석 지구 46억년의 비밀
시그마프레스
2003
[50]
웹인용
스콧 R. 쇼. <곤충 연대기>(원제
[51]
논문
Time scales of critical events around the Cretaceous-Paleogene boundary
[52]
웹사이트
The Great Dying
http://science.nasa.[...]
NASA
[53]
서적
A Geologic Time Scale 2004
http://www.cambridge[...]
[54]
서적
Visualizing Earth Science
Wiley
[55]
서적
지구가 정말 이상하다
살림
2005
[56]
서적
연꽃의 세계
김영사
2007
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com