드리오피테쿠스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
''드리오피테쿠스''는 '참나무 유인원'이라는 뜻으로, 1856년 프랑스에서 최초의 화석이 발견된 멸종된 유인원 속이다. 이들은 오랑우탄과 유사한 체형을 가졌으며, 치아 구조는 현생 침팬지와 유사하다. ''드리오피테쿠스''는 족 드리오피테키니에 속하며, 오랑우탄, 아프리카 유인원, 인간의 조상 또는 별도의 계통으로 간주된다. 온난한 기후의 중신세에 유럽의 숲에서 번성했지만, 기후 변화로 인한 환경 변화로 멸종했을 것으로 추정된다.
더 읽어볼만한 페이지
- 고대 유인원 - 호모 에렉투스
호모 에렉투스는 약 200만 년 전부터 10만 년 전까지 생존하며 직립 보행을 하고 아슐리안 석기를 제작했으며 불을 사용한 최초의 인류로 추정되는 종이다. - 고대 유인원 - 네안데르탈인
네안데르탈인은 유럽과 서아시아에 살았던 고대 인류로, 큰 뇌 용량과 독특한 신체적 특징을 지녔으며, 무스테리안 문화를 사용하고 동굴에서 거주하며 매장 의례를 치르는 등 복잡한 사회생활을 영위했지만, 현생 인류와의 경쟁, 기후 변화 등으로 약 4만 년 전 멸종한 종이며, 현생 인류와 이종교배하여 현생 인류 유전체에 일부 유전자가 남아있다. - 1856년 기재된 화석 분류군 - 트로오돈
트로오돈은 후기 백악기 북아메리카에 서식한 수각류 공룡의 한 속으로, 종의 구분에 대한 논쟁과 모식표본의 불완전성으로 의문명으로 간주될 가능성이 있으며, 큰 뇌 용량으로 "중생대에서 가장 똑똑한 동물"로 여겨지기도 하고 야행성일 가능성이 제기되는 등 분류와 생태에 대한 연구가 지속적으로 진행 중이다. - 1856년 기재된 화석 분류군 - 트라코돈
트라코돈은 조지프 레이디가 1856년에 명명한 공룡 속으로 북미 공룡에게 붙여진 최초의 이름 중 하나였으나, 현재는 모식표본의 불완전성으로 인해 의문명으로 간주되는 하드로사우루스과의 육상 초식 공룡이다. - 마이오세의 포유류 - 수마트라코뿔소
수마트라코뿔소는 인도네시아 수마트라 섬에서 발견된 가장 작은 코뿔소 종으로, 두 개의 뿔과 털이 많으며 멸종 위기에 놓여 국제적인 보호를 받고 있다. - 마이오세의 포유류 - 사헬란트로푸스
사헬란트로푸스는 약 700만 년 전에서 680만 년 전에 생존했던 화석 인류의 한 속으로, 2001년 차드에서 발견된 '투마이' 화석 표본을 통해 알려져 있으며, 작은 두개골과 턱뼈, 작은 뇌 용량을 특징으로 하고, 분류 및 직립 보행 여부에 대한 논쟁에도 불구하고 인류 진화 연구에 중요한 자료로 평가받는다.
| 드리오피테쿠스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
| 학명 | Dryopithecus |
| 명명자 | Lartet, 1856 |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 목 | 영장목 |
| 아목 | 직비원아목 |
| 하목 | 원숭이하목 |
| 상과 | 유인원상과 |
| 과 | † 드리오피테쿠스과 (Dryopithecidae) |
| 속 | † 드리오피테쿠스속 (Dryopithecus) |
| 종 | |
| 하위 종 | †Dryopithecus fontani Lartet, 1856 ?†Dryopithecus carinthiacus Mottl, 1957 |
| 지질 시대 | |
| 화석 범위 | 세라발레절–토르토나절 () |
| 물리적 특징 | |
| 추정 몸무게 | 44 kg |
| 이미지 | |
 | |
2. 어원
속명 ''드리오피테쿠스''는 고대 그리스어로 '참나무'를 뜻하는 ''drus''와 '유인원'을 뜻하는 ''pithekos''의 합성어에서 유래했는데, 이는 명명자가 드리오피테쿠스가 현대 유럽과 유사한 환경의 참나무 또는 소나무 숲에 서식했다고 믿었기 때문이다.[2] 종명 ''D. fontani''는 발견자인 지역 수집가 알프레드 퐁탄(Monsieur Alfred Fontan)을 기리기 위해 명명되었다.[1]
3. 분류

드리오피테쿠스는 드리오피테키니 족에 속하며, 히스파노피테쿠스, 루다피테쿠스, 우라노피테쿠스, 아노이아피테쿠스, 피에롤라피테쿠스 등과 함께 분류된다.[4] 하지만 이들의 분류 관계는 명확하지 않아, 아노이아피테쿠스와 피에롤라피테쿠스는 드리오피테쿠스 속에 포함될 수 있고,[4] 히스파노피테쿠스와 루다피테쿠스는 서로 같은 종일 수 있으며,[15] 히스파노피테쿠스, 루다피테쿠스, 우라노피테쿠스는 별도의 족으로 분류될 수도 있다.[15]
드리오피테키니는 오랑우탄이 속한 사람아과의 한 갈래이거나,[16] 아프리카 유인원과 인간을 포함하는 사람아과의 조상이거나,[17][18][19] 혹은 독립된 드리오피테쿠스아과로 분류되기도 한다.[15][20]
드리오피테쿠스는 중신세 중기 기후가 온난했던 시기에 유럽의 넓은 숲에서 번성했던 유인원 무리의 일원으로, 중신세 중기 붕괴라는 냉각화 현상으로 다양해진 초기 또는 중기 중신세 아프리카 유인원에서 기원했을 것으로 추정된다.[21][4][22] 유인원이 유럽이나 아시아에서 먼저 진화하여 아프리카로 이동했을 가능성도 제기된다.[21][4][22]
3. 1. 분류사
최초의 ''드리오피테쿠스'' 화석은 1856년 프랑스 고생물학자 에두아르 라르테에 의해 프랑스 피레네 산맥에서 기술되었는데,[1] 이는 찰스 다윈이 그의 저서 ''종의 기원''을 출판하기 3년 전이었다. 이후 연구자들은 현존하는 아프리카 유인원과의 유사성을 언급했다. 다윈은 그의 저서 ''인간의 유래''에서 ''드리오피테쿠스''가 유인원의 아프리카 기원에 의문을 제기한다고 간략하게 언급했다.[3]
''드리오피테쿠스''의 분류는 많은 혼란을 겪어 왔으며, 미세한 차이를 근거로 새로운 표본이 새로운 종 또는 속의 기초가 되어 현재는 폐지된 여러 종이 생겨났다.[4] 1960년대까지 모든 비인간 유인원은 현재 무효가 된 과인 사람과에 분류되었고, 멸종된 유인원은 드리오피테쿠스과에 분류되었다.[4] 1965년, 영국의 고인류학자 데이비드 필빔과 미국의 고생물학자 엘윈 L. 시몬스는 당시 구세계 전역에서 발견된 표본을 포함하는 이 속을 유럽의 ''드리오피테쿠스'', 아시아의 ''시바피테쿠스'', 아프리카의 ''프로콘술''의 세 아속으로 나누었다. 그 후, 각 아속을 속으로 승격시킬지에 대한 논의가 있었다. 1979년, ''시바피테쿠스''는 속으로 승격되었고, ''드리오피테쿠스''는 다시 유럽의 ''드리오피테쿠스'', 아프리카의 ''프로콘술'', ''림노피테쿠스'', ''랑가피테쿠스''로 아속이 세분되었다.[5] 그 이후로 몇몇 종이 더 지정되고 이동되었으며, 21세기까지 이 속에는 ''D. fontani'', ''D. brancoi'',[6][7] ''D. laietanus'',[8] 그리고 ''D. crusafonti''가 포함되었다.[9] 그러나 2009년 ''D. fontani''의 부분 두개골 발견으로 인해 많은 종들이 새로운 속, 예를 들어 새롭게 세워진 ''히스파노피테쿠스'' 등으로 분리되었는데, 그 이유는 ''드리오피테쿠스'' 모식표본의 단편적인 특성, 불분명하고 불완전한 진단적 특징이 혼란을 야기했기 때문이다.[10][4]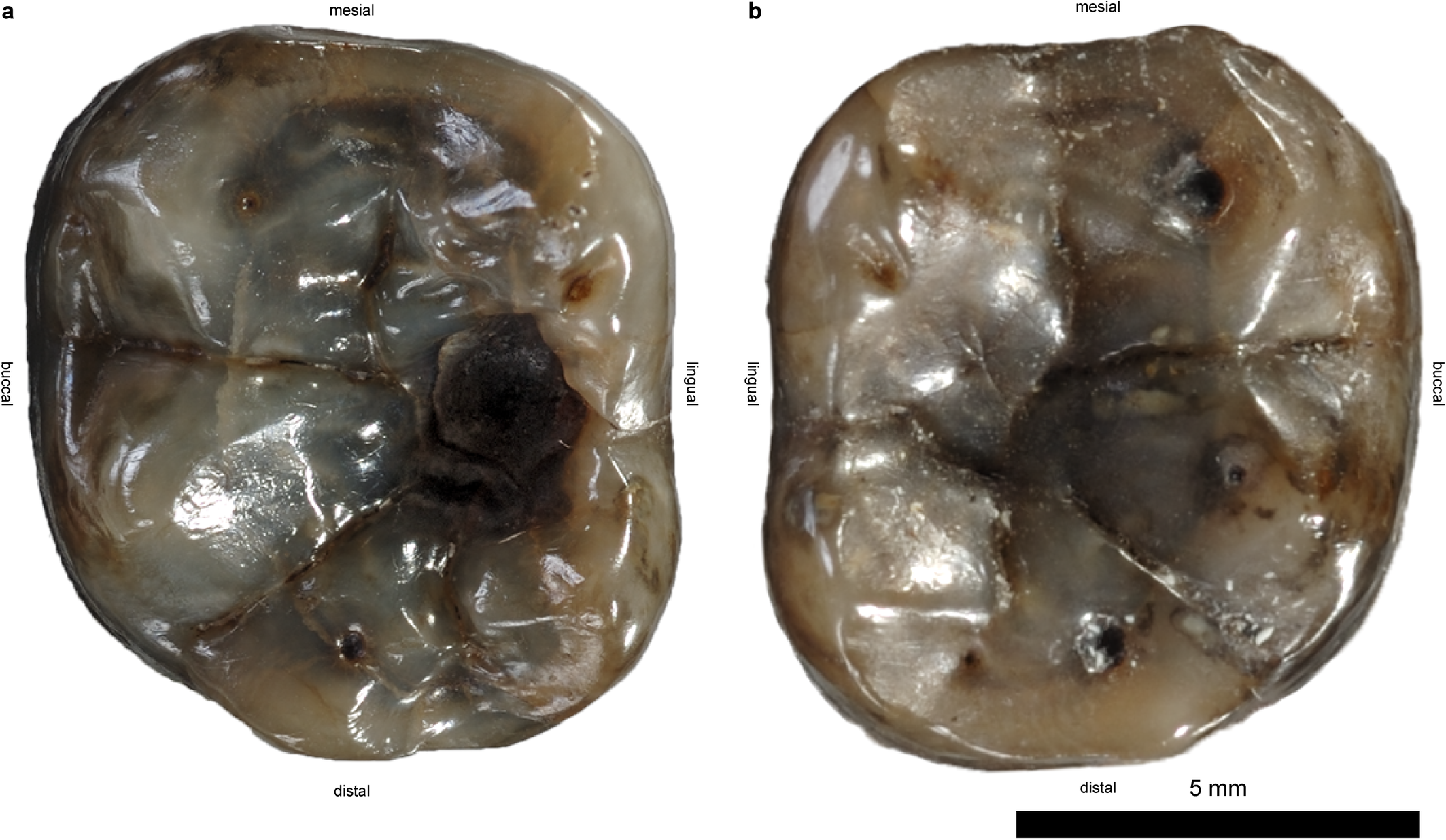
현재, 논란의 여지가 없는 종은 ''D. fontani'' 하나뿐이다. 표본은 다음과 같다.
| 표본 | 발견 지역 | 특징 | 기타 |
|---|---|---|---|
| MNHNP AC 36 (모식표본) | 프랑스 피레네 산맥 생고댕 | 치아가 있는 수컷 하악골 조각 3개[4][11] | 침팬지의 치아 발달을 기준으로 6~8세[12] |
| NMB G.a.9., FSL 213981 | 프랑스 생탈방드호슈 | 상부 절치, 암컷 상부 어금니[13] | |
| IPS35026, IPS41724 | 스페인 카탈루냐 바예스 페네데스 | 수컷 부분 안면, 대퇴골[10][4] | |
| LMK-Pal 5508 | 오스트리아 케른텐주 생 슈테판 | 치아가 있는 암컷 하악골[4][24][14] | 1250만 년 전, 별도의 종 "D. carinthiacus"로 간주될 수 있음[4] |
''드리오피테쿠스''는 족인 드리오피테키니에 분류되며, ''히스파노피테쿠스'', ''루다피테쿠스'', ''우라노피테쿠스'', ''아노이아피테쿠스'', 그리고 ''피에롤라피테쿠스''와 함께 분류되지만, 후자의 두 종은 ''드리오피테쿠스''에 속할 수 있고,[4] 전자의 두 종은 동의어가 될 수 있으며, 전자의 세 종은 자체적인 족에 배치될 수도 있다.[15] 드리오피테키니는 오랑우탄 (사람아과)의 분파,[16] 아프리카 유인원과 인간 (사람아과)의 조상,[17][18][19] 또는 자체적인 별도의 계통 (드리오피테쿠스아과)으로 간주된다.[15][20]
''드리오피테쿠스''는 적응 방산의 일부였으며, 온난한 기후의 중신세 기후 최적기에 유럽의 확장된 숲에서 유인원이 번성했고, 아마도 진행되는 중신세 중기 붕괴 (냉각 현상)에서 다양화된 초기 중신세 또는 중기 중신세 아프리카 유인원으로부터 기원했을 것이다. 유인원이 먼저 유럽이나 아시아에서 진화한 다음 아프리카로 이주했을 가능성이 있다.[21][4][22]
3. 2. 현생 분류군과의 관계
최초의 ''드리오피테쿠스'' 화석은 1856년 프랑스 고생물학자 에두아르 라르테에 의해 프랑스 피레네 산맥에서 기술되었는데,[1] 이는 찰스 다윈이 그의 저서 ''종의 기원''을 출판하기 3년 전이었다. 이후의 연구자들은 현존하는 아프리카 유인원과의 유사성을 언급했다. 다윈은 그의 저서 ''인간의 유래''에서 ''드리오피테쿠스''가 유인원의 아프리카 기원에 의문을 제기한다고 간략하게 언급했다.[3]''드리오피테쿠스''의 분류는 많은 혼란을 겪어 왔으며, 미세한 차이를 근거로 새로운 표본이 새로운 종 또는 속의 기초가 되어 현재는 폐지된 여러 종이 생겨났다.[4] 1960년대까지 모든 비인간 유인원은 현재 무효가 된 과인 사람과에 분류되었고, 멸종된 유인원은 드리오피테쿠스과에 분류되었다.[4] 1965년, 영국의 고인류학자 데이비드 필빔과 미국의 고생물학자 엘윈 L. 시몬스는 당시 구세계 전역에서 발견된 표본을 포함하는 이 속을 유럽의 ''드리오피테쿠스'', 아시아의 ''시바피테쿠스'', 아프리카의 ''프로콘술''의 세 아속으로 나누었다. 그 후, 각 아속을 속으로 승격시킬지에 대한 논의가 있었다. 1979년, ''시바피테쿠스''는 속으로 승격되었고, ''드리오피테쿠스''는 다시 유럽의 ''드리오피테쿠스'', 아프리카의 ''프로콘술'', ''림노피테쿠스'', ''랑가피테쿠스''로 아속이 세분되었다.[5] 그 이후로 몇몇 종이 더 지정되고 이동되었으며, 21세기까지 이 속에는 ''D. fontani'', ''D. brancoi'',[6][7] ''D. laietanus'',[8] 그리고 ''D. crusafonti''가 포함되었다.[9] 그러나 2009년 ''D. fontani''의 부분 두개골 발견으로 인해 많은 종들이 새로운 속, 예를 들어 새롭게 세워진 ''히스파노피테쿠스'' 등으로 분리되었는데, 그 이유는 ''드리오피테쿠스'' 모식표본의 단편적인 특성, 불분명하고 불완전한 진단적 특징이 혼란을 야기했기 때문이다.[10][4]
현재, 논란의 여지가 없는 종은 ''D. fontani'' 하나뿐이다. 표본은 다음과 같다.
| 표본명 | 발견 지역 | 특징 |
|---|---|---|
| MNHNP AC 36 | 프랑스 피레네 산맥의 생고댕 | 치아가 있는 수컷 하악골 조각 3개,[4][11] 침팬지의 치아 발달을 기준으로 6~8세[12] |
| NMB G.a.9., FSL 213981 | 프랑스 생탈방드호슈 | 상부 절치, 암컷 상부 어금니[13] |
| IPS35026, IPS41724 | 스페인 카탈루냐 바예스 페네데스 | 수컷 부분 안면, 대퇴골[10][4] |
| LMK-Pal 5508 | 오스트리아 케른텐주 생 슈테판 | 치아가 있는 암컷 하악골, 별도의 종, "D. carinthiacus"로 간주될 수 있음[4][24][14] |
''드리오피테쿠스''는 족인 드리오피테키니에 분류되며, ''히스파노피테쿠스'', ''루다피테쿠스'', ''우라노피테쿠스'', ''아노이아피테쿠스'', 그리고 ''피에롤라피테쿠스''와 함께 분류되지만, 후자의 두 종은 ''드리오피테쿠스''에 속할 수 있고,[4] 전자의 두 종은 동의어가 될 수 있으며, 전자의 세 종은 자체적인 족에 배치될 수도 있다.[15] 드리오피테키니는 오랑우탄 (사람아과)의 분파,[16] 아프리카 유인원과 인간 (사람아과)의 조상,[17][18][19] 또는 자체적인 별도의 계통 (드리오피테쿠스아과)으로 간주된다.[15][20]
''드리오피테쿠스''는 적응 방산의 일부였으며, 온난한 기후의 중신세 기후 최적기에 유럽의 확장된 숲에서 유인원이 번성했고, 아마도 진행되는 중신세 중기 붕괴 (냉각 현상)에서 다양화된 초기 중신세 또는 중기 중신세 아프리카 유인원으로부터 기원했을 것이다. 유인원이 먼저 유럽이나 아시아에서 진화한 다음 아프리카로 이주했을 가능성이 있다.[21][4][22]
4. 특징
드리오피테쿠스는 두 발로 서서 걸을 수도 있었지만 주로 네 발로 걸었다. 넙다리뼈의 구조는 고관절의 이동성에 중요하며, 매달리기보다는 사지보행 방식에 더 적합했음을 보여준다.[10]
4. 1. 해부학적 특징
넙다리뼈 머리 측정을 바탕으로 수컷 ''드리오피테쿠스''의 생존 시 체중은 44kg으로 추정된다.[10]''드리오피테쿠스''의 치아는 현생 침팬지와 가장 유사하다. 치아는 작고 에나멜 층이 얇다. ''드리오피테쿠스''는 턱이 가늘어 거칠거나 단단한 음식을 먹는 데 적합하지 않았음을 시사한다. 현생 유인원과 마찬가지로 수컷은 송곳니가 뚜렷하다.[26][16] 어금니는 넓고 작은 어금니는 더 넓다. 입천장, 긴 주둥이 (전돌증), 얼굴에 거의 수직으로 향하는 큰 코를 가지고 있다. 전체적으로 얼굴은 고릴라와 많은 유사점을 보인다. 초기에서 중기 미오세 아프리카 유인원은 이러한 유사점을 공유하지 않으므로 고릴라와 같은 특징은 가까운 친연관계의 결과라기보다는 ''드리오피테쿠스''에서 독립적으로 진화했을 가능성이 높다.[10]
약 265mm를 측정하는 위팔뼈는 보노보와 크기와 형태가 유사하다. 보노보와 마찬가지로 몸통은 바깥쪽으로 구부러져 있으며, 삼두근과 삼각근의 삽입이 제대로 발달하지 않아 ''드리오피테쿠스''는 오랑우탄만큼 매달리기 행동에 능숙하지 않았음을 시사한다.[23] 넙다리뼈 목은 넙다리뼈 머리를 넙다리뼈 몸통에 연결하며, 매우 길거나 가파르지 않다. 넙다리뼈 머리는 큰 돌기에 낮게 위치하며, 작은 돌기는 뒷면을 향해 더 많이 위치한다. 이 모든 특징은 고관절의 이동성에 중요하며, 매달리기보다는 사지보행 방식을 나타낸다.[10] 그러나 오스트리아 ''드리오피테쿠스'' 시대와 지역의 과일 나무는 일반적으로 5m에서 12m 높이였고 얇은 말단 가지에서 열매를 맺어 이를 잡기 위한 매달리기 행동을 시사한다.[24]
4. 2. 이동 방식
드리 오피테쿠스는 두 발로 서서 걸을 수도 있었지만 주로 네 발로 걸었다. 넙다리뼈 목은 넙다리뼈 머리를 넙다리뼈 몸통에 연결하며, 매우 길거나 가파르지 않다. 넙다리뼈 머리는 큰 돌기에 낮게 위치하며, 작은 돌기는 뒷면을 향해 더 많이 위치한다. 이 모든 특징은 고관절의 이동성에 중요하며, 매달리기보다는 사지보행 방식을 나타낸다.[10] 약 265mm를 측정하는 위팔뼈는 보노보와 크기와 형태가 유사하다. 보노보와 마찬가지로 몸통은 바깥쪽으로 구부러져 있으며, 삼두근과 삼각근의 삽입이 제대로 발달하지 않아 ''드리오피테쿠스''는 오랑우탄만큼 매달리기 행동에 능숙하지 않았음을 시사한다.[23] 그러나 오스트리아 ''드리오피테쿠스'' 시대와 지역의 과일 나무는 일반적으로 5m에서 12m 높이였고 얇은 말단 가지에서 열매를 맺어 이를 잡기 위한 매달리기 행동을 시사한다.[24]5. 고생물학
드리오피테쿠스는 주로 과일을 먹는 과식성 동물이었을 것으로 추정되며, 높은 당 섭취는 충치 발생과 관련이 있다. 치아 마모 패턴은 부드럽고 딱딱한 음식을 모두 섭취했음을 시사하며, 이는 다양한 먹이 섭취 또는 대체 먹이 섭취 전략을 나타낸다. 특화되지 않은 치아와 큰 체구는 소화하기 어려운 음식도 처리할 수 있는 유연한 식성을 가졌음을 보여주며, 현대 유인원처럼 기근 시기에는 잎과 같은 엽식성 음식도 섭취했을 수 있다. 현대 유인원과 달리, 드리오피테쿠스는 높은 탄수화물과 낮은 섬유질 식단을 가졌을 가능성이 높다. 높은 과당 섭취는 요산 수치 상승과 관련이 있는데, 이는 대형 유인원을 제외한 대부분의 동물에서 유리카제에 의해 중화된다. 대형 유인원은 1500만 년 전 유리카제 생산을 중단하여 혈압 상승, 활동성 증가, 지방 축적 능력 향상을 겪었을 것으로 추정된다. 후기 마이오세 오스트리아의 고환경은 1년 중 9~10개월 동안 과일 나무와 꿀이 풍부했음을 보여주며, 드리오피테쿠스는 늦겨울 동안 이러한 지방 비축량에 의존했을 수 있다. 혈액 내 높은 요산 수치는 지능 증가와도 관련이 있다. 수컷 드리오피테쿠스는 암컷보다 큰 송곳니를 가졌는데, 이는 현대 영장류에서 높은 수준의 공격성과 관련된다.[26]
드리오피테쿠스의 유해는 곰포테리움, 라르테토테리움, 리스트리오돈, 미오트라게로스, 안키테리움, 프로틱티테리움, 슈도아엘루루스 등 다양한 대형 포유류와 함께 발견된다. 또한, 히스파노피테쿠스, 아노이아피테쿠스, 피에롤라피테쿠스, 플리오피테쿠스 등 다른 유인원과 공존했다. 이러한 동물군은 따뜻하고 숲이 우거진 준열대 습지 환경과 일치하며, 계절성 기후에서 살았을 가능성이 있다. 오스트리아의 드리오피테쿠스는 벚나무, 포도나무, 검은 뽕나무, 스트로베리 나무, 히코리, 밤나무 등이 주요 과일 공급원이었고, 참나무, 너도밤나무, 느릅나무, 소나무 등이 꿀 공급원이었을 것으로 추정된다.[24]
후기 마이오세는 유럽의 건조화 추세가 시작된 시기였다. 지중해 지역의 계절성, 가뭄 증가, 지중해성 기후 출현은 숲과 삼림을 관목지로 대체했고, 알프스 산맥 융기는 중부 유럽의 식생 변화를 야기하여 유럽 유인원 멸종의 원인이 되었을 가능성이 있다.[30]
5. 1. 식성

''드리오피테쿠스''는 과일(과식성)을 주로 먹었을 가능성이 높으며, 오스트리아 ''드리오피테쿠스''의 치아에 나타난 충치 증거는 높은 설탕 섭취, 즉 익은 과일과 꿀에서 유래했을 가능성이 높다는 것을 나타낸다.[24] 치아 마모는 ''드리오피테쿠스''가 부드럽고 딱딱한 음식을 모두 먹었다는 것을 나타내는데, 이는 다양한 음식을 섭취했거나 더 어려운 음식을 대체식으로 먹었을 수 있음을 의미한다.[25] 그럼에도 불구하고, 특화되지 않은 치아는 유연한 식단을 가지고 있음을 나타내며, 큰 체구는 소화하기 어려운 음식을 처리하는 데 도움이 되는 큰 창자를 허용했을 것이고, 현대 유인원처럼 기근 시기에는 잎(엽식성)과 같은 음식을 포함했을 수 있다. 현대 유인원과 달리, ''드리오피테쿠스''는 높은 탄수화물, 낮은 섬유질 식단을 가졌을 가능성이 높다.[24]
높은 과당 식단은 요산 수치 상승과 관련이 있으며, 이는 대형 유인원을 제외한 대부분의 동물에서 유리카제에 의해 중화된다. 이들은 1500만 년 전에 생산을 중단하여 혈압이 상승하고, 이로 인해 활동성이 증가하고, 지방 축적 능력이 향상된 것으로 생각된다. 후기 미오세 오스트리아의 고환경은 1년 중 9~10개월 동안 과일 나무와 꿀이 풍부했음을 나타내며, ''드리오피테쿠스''는 늦겨울 동안 이러한 지방 비축량에 의존했을 수 있다. 혈액 내 높은 요산 수치는 또한 지능 증가와 관련이 있다.[24]
''드리오피테쿠스'' 수컷은 암컷보다 더 큰 송곳니를 가지고 있었으며, 이는 현대 영장류에서 높은 수준의 공격성과 관련이 있다.[26]
5. 2. 고환경과 멸종
''드리오피테쿠스''(Dryopithecus)의 유해는 코끼리과(예: ''곰포테리움(Gomphotherium)''), 코뿔소(예: ''라르테토테리움(Lartetotherium)''), 돼지과(예: ''리스트리오돈(Listriodon)''), 소과(예: ''미오트라게로스(Miotragocerus)''), 말과(예: ''안키테리움(Anchitherium)''), 하이에나(예: ''프로틱티테리움(Protictitherium)''), 고양이과(예: ''슈도아엘루루스(Pseudaelurus)'')와 같은 몇몇 대형 포유류와 자주 관련되어 있다. 다른 관련 영장류는 유인원인 ''히스파노피테쿠스''(Hispanopithecus), ''아노이아피테쿠스''(Anoiapithecus), ''피에롤라피테쿠스''(Pierolapithecus)이며, 플리오피테쿠스과 유인원인 ''플리오피테쿠스(Pliopithecus)''가 있다.[27][28] 이러한 동물군은 따뜻하고 숲이 우거진, 준열대 습지 환경과 일치하며,[27][29] 계절성 기후에서 살았을 가능성이 있다. 오스트리아의 ''드리오피테쿠스''의 경우, ''벚나무(Prunus)'', 포도나무, 검은 뽕나무, 스트로베리 나무, 히코리, 밤나무와 같은 식물이 중요한 과일 공급원이었을 수 있으며, 후자의 두 식물과 참나무, 너도밤나무, 느릅나무, 소나무가 꿀 공급원이었을 수 있다.[24]
후기 마이오세는 유럽에서 건조해지는 추세가 시작된 시기였다. 지중해 지역의 계절성과 가뭄 증가, 지중해성 기후의 출현은 숲과 삼림이 열린 관목지로 대체되는 결과를 낳았을 가능성이 높았고, 알프스 산맥의 융기는 중부 유럽의 열대 및 온난 기후 식생이 중위도 및 고산 식물로 후퇴하게 만들었다. 이는 유럽의 유인원 멸종을 초래했을 가능성이 높다.[30]
참조
[1]
논문
Note sur un grand Singe fossile qui se rattache au groupe des Singes Supérieurs
[2]
서적
Hominid Adaptations and Extinctions
UNSW Press
[3]
서적
The Descent of Man
D. Appleton and Company
[4]
서적
"Dryopithecus"
Wiley Online Library
[5]
서적
Evolutionary History of the Primates
https://books.google[...]
Academic Press
[6]
논문
Paleoenvironment of ''Dryopithecus brancoi'' at Rudabánya, Hungary: evidence from dental meso- and micro-wear analyses of large vegetarian mammals
[7]
논문
A new cranium of Dryopithecus from Rudabánya, Hungary
[8]
논문
A ''Dryopithecus'' skeleton and the origins of great-ape locomotion
[9]
논문
"Dryopithecus crusafonti sp. nov., a new Miocene Hominoid species from Can Ponsic (northeastern Spain)"
[10]
논문
First partial face and upper dentition of the Middle Miocene hominoid ''Dryopithecus fontani'' from Abocador de Can Mata (Vallès-Penedès Basin, Catalonia, NE Spain): taxonomic and phylogenetic implications
[11]
논문
Updated chronology for the Miocene hominoid radiation in Western Eurasia
[12]
논문
Mandibular ontogeny in the Miocene great ape ''Dryopithecus''
[13]
논문
Taxonomic attribution of the La Grive hominoid teeth
[14]
논문
Bericht über die neuen Menschenaffenfunde aus Österreich, von St. Stefan im Lavanttal, Kärnten. Carinthia II.
https://www.zobodat.[...]
[15]
PhD
The craniodental anatomy of Miocene apes from the Vallès-Penedès Basin (Primates: Hominidae): Implications for the origin of extant great apes
Universitat Autònoma de Barcelona
[16]
논문
Fossil apes from the Vallès-Penedès basin
[17]
논문
"Sivapithecus is east and Dryopithecus is west, and never the twain shall meet"
https://www.jstage.j[...]
[18]
논문
Dryopithecins, Darwin, de Bonis, and the European origin of the African apes and human clade
http://sciencepress.[...]
[19]
논문
European Miocene hominids and the origin of the African ape and human clade
[20]
서적
Hominoid evolution and climatic change in Europe
Cambridge University Press
[21]
논문
"Dryopithecus crusafonti sp. nov., a new Miocene Hominoid species from Can Ponsic (northeastern Spain)"
[22]
논문
Primate evolution in the Pannionian Basin: In situ evolution, dispersals, or both?
[23]
논문
Biological sciences: humerus of ''Dryopithecus'' from Saint Gaudens, France
[24]
논문
Earliest evidence of caries lesion in hominids reveal sugar-rich diet for a Middle Miocene dryopithecine from Europe
[25]
논문
Dietary Specialization during the Evolution of Western Eurasian Hominoids and the Extinction of European Great Apes
[26]
논문
Canine reduction in the miocene hominoid ''Oreopithecus bambolii'': behavioural and evolutionary implications
[27]
논문
The Miocene mammal record of the Vallès-Penedès Basin (Catalonia)
[28]
논문
Sur l'âge relatif des différents dépôts karstiques miocènes de La Grive-Saint-Alban (Isère)
https://www.persee.f[...]
[29]
논문
A limnic ostracod fauna from the surroundings of the Central Paratethys (Late Middle Miocene/Early Late Miocene; Styrian Basin; Austria)
[30]
논문
Ruminant diets and the Miocene extinction of European great apes
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com