마멘키사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
마멘키사우루스(Mamenchisaurus)는 중국에서 발견된 용각류 공룡의 한 속으로, 매우 긴 목이 특징이다. 1952년 처음 발견된 M. constructus를 시작으로, M. hochuanensis, M. sinocanadorum, M. anyuensis, M. youngi, M. jingyanensis 등 여러 종이 보고되었으나, 일부 종의 분류는 불확실하다. 마멘키사우루스는 전형적인 용각류의 특징인 네 발 걷기, 큰 몸집, 작은 머리를 공유하며, 특히 목의 길이가 매우 길어 전체 길이의 절반에 가깝다. 화석은 쥐라기 후기에서 백악기 초기에 걸쳐 발견되며, 서식 환경, 먹이 섭취 방식, 분류학적 위치에 대한 연구가 진행 중이다.
더 읽어볼만한 페이지
- 1954년 기재된 화석 분류군 - 테리지노사우루스
테리지노사우루스는 백악기 후기 몽골에 서식한 거대한 수각류 공룡으로, 앞발의 매우 긴 발톱이 특징이며, 초기에 파충류로 오인되었으나 현재는 테리지노사우루스류에 속하는 마니랍토라 공룡으로 분류되고 초식 또는 잡식성 생활을 했을 것으로 추정된다. - 1954년 기재된 화석 분류군 - 레바키사우루스
레바키사우루스는 약 20m 길이의 중형 용각류 공룡으로, 목에서 꼬리까지 가시 모양 뼈 돌기가 특징이며, 아르헨티나, 모로코 등지에서 화석이 발견되었고, 레바키사우루스과에 속한다. - 양중젠이 명명한 분류군 - 오메이사우루스
오메이사우루스는 중국 쓰촨성에서 발견된 용각류 공룡의 한 속으로, 쯔궁 지역에서 많은 화석이 발견되었으며 마멘키사우루스과에 속한다. - 양중젠이 명명한 분류군 - 친타오사우루스
친타오사우루스는 백악기 후기 중국 산둥성에서 서식한 람베오사우루스아과 조각류 공룡으로, 머리에 있는 독특한 볏이 특징이며, 1958년 양중젠에 의해 명명되었고, 람베오사우루스아과 내에서 파라라브도돈과 관련이 있는 것으로 분류된다. - 쥐라기 후기 아시아의 공룡 - 알로사우루스
알로사우루스는 쥐라기 후기에 살았던 대형 수각류 공룡으로, 짧은 앞다리, 머리의 뿔, 갈고리 발톱을 특징으로 하며, 모리슨층에서 발견되었고, 다른 육식 공룡과 함께 먹이 사슬의 정점에 있었다. - 쥐라기 후기 아시아의 공룡 - 인롱
인롱은 약 1억 6천만 년 전 쥐라기 후기에 살았던 초기 각룡류 공룡으로, 쥐라기 지층에서 발견되었다는 점과 위석을 가지고 있었다는 특징을 가진다.
| 마멘키사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Mamenchisaurus |
| 어원 | zh (Mǎmíngxī, 마밍시) |
| 명명자 | 양종건 (Young), 1954 |
| 속 | 용각아목 |
| 종 | |
| 대표종 | Mamenchisaurus constructus |
| 기타 종 | M. hochuanensis Young & Zhao, 1972 M. sinocanadorum Russell & Zheng, 1993 M. youngi Pi, Ouyang & Ye, 1996 M. anyuensis He et al., 1996 "'M. guangyuanensis"' Li & Cai, 1997 (학명 미등록) M. jingyanensis Zhang, Li & Zeng, 1998 M. yunnanensis Fang et al., 2004 |
| 생물학적 분류 | |
| 속 | 용각류 |
| 시대 | |
| 시기 | 쥐라기 후기 - 백악기 전기 (옥스포디언절 - 압트절), |
| 기타 | 알비절 동안의 기록 가능성 |
| 크기 | |
| 길이 | 15~26 미터 |
| 최대 길이 추정 | 35 미터 |
| 발음 | |
| 영어 (IPA) | /məˌmʌntʃiˈsɔːrəs/ |
| 영어 (발음 구별 기호) | mə-MUN-chee-SOR-əs |
| 영어 (IPA) | /məˌmɛntʃɪˈsɔːrəs/ |
2. 역사 및 종
마멘키사우루스는 여러 종이 발견되었으며, 그 중 일부는 다른 속으로 재분류되거나 유효성에 대한 논쟁이 있다.
- 지공고사우루스 푸시엔시스 (''Zigongosaurus fuxiensis''): 1976년에 명명되었으나, 이후 연구자들에 의해 ''오메이사우루스''로 재분류되거나 ''마멘키사우루스 푸시엔시스''(M. fuxiensis)로 할당되기도 했다. 그러나 2019년에는 진단 불가능한 것으로 간주되었다.[3][16]
- 마멘키사우루스 광위안엔시스 ("Mamenchisaurus guangyuanensis"): 공식적으로 기술되지 않은 ''노멘 누둠'' 상태이며, 상샤시먀오 지층에서 유년기부터 성체까지 여러 개체의 유해가 발견되었다.
- 마멘키사우루스 윈난엔시스 (''M. yunnanensis''): 2004년에 기술되었으나, ''마멘키사우루스'' 속으로 분류하는 것에 대한 의문이 제기되었다.[3]
- 오메이사우루스 창쇼우엔시스 (''Omeisaurus changshouensis''): 꼬리 척추 특징으로 인해 ''마멘키사우루스''에 더 가깝다고 간주되었으나, 2004년에 진단 불가능하다는 의견이 제시되었다.[26]
- 모시류 ("Moshisaurus"): 일본에서 발견된 불완전한 상완골을 바탕으로 ''마멘키사우루스''의 일종으로 추정되었으나, 특정 용각류 그룹으로 분류하기 어렵다는 재평가를 받았다.[24][25]

NSM PV17656
2. 1. 발견 및 명명

''마멘키사우루스''(Mamenchisaurus)는 1952년 중국 쓰촨성 이탕 고속도로 건설 현장에서 처음 발견되었다. 이 화석지는 늦은 쥐라기에 해당하는 상샤시먀오 지층에 속한다.[3] 이 부분적인 골격 화석은 이후 연구되어 1954년 중국의 저명한 고생물학자인 양중젠 교수에 의해 ''마멘키사우루스 콩스트룩투스''(Mamenchisaurus constructus)로 명명되었다.
모식 표본(IVPP V. 790)은 단편적이고, 정돈되지 않았으며, 기술적인 방식으로 발굴되지 않았다. 재료는 5개의 등쪽 척추, 30개의 꼬리 척추, 갈비뼈 조각, 등쪽 신경 가시 및 혈관궁을 포함했다. 14개의 목 척추가 보존되었지만 완전한 것은 없었다. 양중젠은 일부 목 척추가 누락되었을 수 있다고 언급했다. 사지 재료는 두 조각의 대퇴골, 완전한 경골, 비골, 거골, 중족골, 발가락 뼈, 발톱을 포함했다. 두개골, 앞다리, 골반대는 없었다.[7]
''마멘키사우루스''는 '마멘치 도마뱀'을 의미하며, 중국어 병음 'mǎ'(马 '말')와 'mén'(门 '문'), 'chi'는 'xī'(溪 '개울' 또는 '시내')의 대체 음역으로, 접미사 '-saurus'(고대 그리스어 'sauros'는 '도마뱀'을 의미)와 결합한 것이다. 이는 화석이 처음 발견된 장소의 이름을 따서 속명을 지으려는 의도였다. 그러나 양중젠의 억양 혼동으로 인해, 지명 ''Mǎmíngxī''(马鸣溪 '말 울음 시내')가 ''Mǎménxī''(马门溪 '말 문 시내')로 오인되었다.[8] 첫 번째 ''마멘키사우루스'' 화석이 건설 작업으로 인해 발견되었다는 사실은 양중젠이 모식종을 ''마멘키사우루스 콩스트룩투스''(Mamenchisaurus constructus)로 명명하게 했다.[7]
1958년 양중젠은 간쑤성에서 수집된 추가적인 용각류 유해를 기술했다. 유해는 다양한 부분 표본(IVPP V. 945, V. 946, V. 947, V. 948)으로 구성되었으며, 대부분이 ''M. constructus''에 할당되었다.[9] 1972년, 이 표본 중 하나가 ''M. hochuanensis''로 재할당되었다.[10]
2. 2. 주요 종

마멘키사우루스는 현재까지 여러 종이 발견되었으며, 각 종은 발견된 지역과 화석의 특징에 따라 구분된다.
| 종 | 발견 연도 | 특징 |
|---|---|---|
| 마멘키사우루스 호추안엔시스 (Mamenchisaurus hochuanensis) | 1972년 | 중국에서 가장 큰 용각류 중 하나 |
| 마멘키사우루스 시노카나도룸 (Mamenchisaurus sinocanadorum) | 1993년 | 신장티탄과 밀접한 관련이 있는 것으로 추정 |
| 마멘키사우루스 아니우엔시스 (Mamenchisaurus anyuensis) | 1996년 | 다른 종들보다 약 3천만 년 더 젊어 속의 구성원일 가능성이 낮음 |
| 마멘키사우루스 영기 (Mamenchisaurus youngi) | 1996년 | 매우 완전한 골격과 두개골이 보존된 표본 발견 |
| 마멘키사우루스 징옌엔시스 (Mamenchisaurus jingyanensis) | 1998년 | 여러 표본이 발견되어 다양한 골격 정보를 제공 |
2. 2. 1. 마멘키사우루스 호추안엔시스 (Mamenchisaurus hochuanensis)
1972년, 양중젠(Young)과 자오시진(Xijin Zhao)은 두 번째 종인 ''Mamenchisaurus hochuanensis''를 기술했다. 화석은 중국 충칭 북쪽 허촨의 한 마을 근처, 쓰촨 성(원래 쓰촨성의 일부)에서 발견되었는데, 푸강 위의 200m 높이 산비탈에 위치해 있었다.[10] ''M. hochuanensis'' 화석 부지는 늦은 쥐라기까지 거슬러 올라가는 ''M. constructus'' 모식 표본 위치와 매우 가까운 상부 샤시먀오 지층에 속했다.[3] 현지인들은 중국 혁명 이전에 유해를 처음 발견했으나, 유해는 결국 방치되어 현장에서 풍화되었다. 발굴은 1957년에야 시작되었다.모식 표본(CCG V 20401)은 거의 완전하고 연결된 척추 열로 구성되어 있으며, 거의 온전하게 보존된 19개의 길쭉한 경추, 12개의 등쪽 척추, 4개의 천골, 마지막 몇 개를 제외한 35개의 미추를 포함했다. 또한 골격에는 앞다리와 두개골의 대부분이 없었다. ''M. hochuanensis''가 처음 기술되었을 때, 그것은 중국에서 알려진 가장 큰 용각류였다.[11]
1958년, 양중젠은 간쑤성 융덩의 하이시완 지역에서 마멘키사우루스 표본(IVPP V. 946)을 기술했다. 이 표본은 처음에는 ''M. constructus''로 분류되었으나, 1972년에 부모식 표본으로 ''M. hochuanensis''로 재분류되었다. 이 표본은 모식 표본보다 약간 작았고 전체적으로 더 적은 재료로 구성되었지만, 모식 표본에서 누락된 일부 해부학적 세부 사항이 포함되어 있었다.[10]
2001년, 또 다른 표본(ZDM0126)이 기술되어 ''M. hochuanensis''로 언급되었다. 이 표본은 1995년 쓰촨성 쯔궁시 후이둥 신구의 건설 현장에서 발견되었다. 이 표본은 거의 완전하고 대부분 연결되어 있으며, 두개골, 가슴 거들 및 앞다리 재료와 같이 모식 표본에서 누락된 특징을 보존했다. 그러나 2020년 Moore와 동료들의 계통 발생 분석은 이 표본을 ''M. hochuanensis''로 언급하는 것에 의문을 제기했다.[46]
2. 2. 2. 마멘키사우루스 시노카나도룸 (Mamenchisaurus sinocanadorum)
1987년 8월, 중국-캐나다 공룡 프로젝트 탐사대의 Z-M. 동이 절벽에서 튀어나온 목갈비 화석을 발견했다. 화석은 신장 위구르 자치구 정가얼 분지의 시슈고우 지층 상부에서 발견되었는데, 이는 쓰촨 분지 밖에서 발견된 몇 안 되는 마멘키사우루스 중 하나이다.[3][4][12] 이 지역은 약 1억 6220만 년 전으로 추정된다. 발견 장소는 원래 버려진 마을인 장쥔먀오에서 북쪽으로 22km 떨어진 곳으로 보고되었으나, 이후 연구에서 좌표가 수정되었다.[12] 표본이 보존된 거칠고 약한 사암과 뼈의 크고 부서지기 쉬운 특성 때문에 발굴이 어려웠고, 가장 앞쪽의 척추뼈만 회수되었다.[12][4] 1993년 데일 러셀과 종 정은 이 표본을 ''Mamenchisaurus''의 새로운 종으로 명명하여 ''M. sinocanadorum''이라 불렀다. 종명은 중국-캐나다 공룡 프로젝트를 의미한다.[4]표본 IVPP V10603은 완전한 왼쪽 아래턱, 오른쪽 치골, 서골, 오른쪽 익상골, 가능한 외익상골, 오른쪽 사골, 두 번째부터 네 번째 경추, 온전한 왼쪽 목갈비로 구성되어 있다. 척추의 신경 아치는 중심과 잘 융합되어 성숙한 동물임을 나타냈다. 이빨이 완전히 솟아났지만 마모되지 않은 점으로 보아, 이 동물이 굶어 죽었을 가능성이 제기되었다. 절벽 기저부에 뼈가 제한적으로 존재했기 때문에 경추가 부패 전에 부러져 머리와 목의 일부가 하류로 흘러가 얕은 물에 묻혔을 것으로 추정된다.[4] 2023년 앤드류 무어와 동료들은 ''M. sinocanadorum''을 재연구하면서 서골, 오른쪽 익상골, 가능한 외익상골, 오른쪽 사골 및 오른쪽 치골을 찾을 수 없었다. 이들의 계통 발생 분석 결과, ''M. sinocanadorum''은 다른 ''Mamenchisaurus'' 종들과는 다른 분류군에 속하며, 신장티탄과 밀접한 관련이 있는 것으로 나타났다. 그러나 기준종인 ''M. constructus''가 재평가될 때까지는 분류학적 조치가 취해지지 않았다.[12]
2019년 그레고리 S. 폴은 ''M. sinocanadorum''과 동일한 지층에서 발견된 두 개의 큰 경추를 크기와 기원을 근거로 이 종에 비공식적으로 언급하며, 이들이 알려진 가장 큰 공룡 중 하나를 나타낼 수 있다고 제안했다.[5][13] 그러나 무어와 동료들은 이 척추뼈들이 기준 표본과 해부학적으로 중첩되지 않아 진단을 위해 비교할 수 없다는 이유로 ''M. sinocanadorum''으로 분류하는 것을 지지하지 않았다. 또한, 베이징의 중국지질대학교에 전시된 미기재 표본에 대해서도 직접 평가되거나 과학 문헌에 언급된 적이 없어 이 종으로 분류하기에는 시기상조라고 판단했다.[12]
2. 2. 3. 마멘키사우루스 아니우엔시스 (Mamenchisaurus anyuensis)
1996년 신루 허와 동료들에 의해 ''M. anyuensis''가 처음으로 기술되었다. 1987년 쓰촨 분지 롱차오샹 마을 근처 두 곳에서 유해가 발견되었는데, 한 채석장에서는 최소 5~6개체, 다른 채석장에서는 4개체가 발견되었다. 이 중 하나가 홀로타입 AL001이 되었으며, 이는 골격의 2/3를 차지한다. AL002, AL003, AL101-106 표본도 보고되었으며, 이들은 더 많은 골격 정보를 제공한다. 종명은 발견된 안웨 현을 참조한다.[14] ''M. anyuensis''는 수이닝 지층 상부와 펑라이전 지층 하부에서 모두 발견되었다. 우라늄-납 연대 측정에 따르면 ''M. anyuensis''는 수이닝 지층에서 1억 1440만 년 전으로 측정되었다. 이는 다른 ''마멘키사우루스'' 종보다 약 3천만 년 더 젊은 것이므로, ''M. anyuensis''가 실제로 이 속의 구성원일 가능성은 낮다.[3]홀로타입 표본에는 8개의 후방 경추, 12개의 등뼈, 5개의 천추, 여러 미추, 그리고 완전한 골반이 보존되어 있다. 또 다른 표본(AL102)은 목 중간 부분에서 5개의 연결된 경추를 보존하고 있다. 손가락 뼈를 제외하면 앞다리는 여러 개체에 의해 완전히 알려져 있다.[14]
2. 2. 4. 마멘키사우루스 영기 (Mamenchisaurus youngi)
1996년에는 상 샤시먀오 층에서 발견된 ''마멘키사우루스 영기''가 기술되었다.[3][27] 이 화석은 1987년 쓰촨성 쯔궁 시의 한 마을 근처 채석장에서 발견되었다. 이 종은 C. C. 영을 기리기 위해 명명되었다.[27]기준 표본(ZDM 0083)은 매우 완전했고 대부분 연결된 상태로, 머리에서 꼬리뼈 8번째까지 모든 척추뼈가 보존되어 있었다. 또한 가슴뼈대, 골반뼈대, 그리고 네 다리에서 나온 재료도 보존되었다. 이 표본은 거의 완전한 두개골도 보존하고 있었다.[27]
2. 2. 5. 마멘키사우루스 징옌엔시스 (Mamenchisaurus jingyanensis)
1998년 장이홍, 리쿠이, 쩡칭화가 ''M. jingyanensis''를 명명했다. 모식 표본 (CV00734)은 징옌 현의 메이왕 진 행정 본부에서 200m 떨어진 곳에서 발견되었다. 다른 표본 (JV002)은 메이왕에서 약 10km 떨어진 싼장 마을에서 발견되었으며, 부모식 표본이 되었다. 세 번째 표본 (CV00219)은 싼장에서 약 10km 떨어진 뚜자 마을 근처에서 발견되었다.[15] 화석은 쓰촨 분지의 상사시먀오 지층에서 발견되었다.[3][15] 종명은 대부분의 표본이 발굴된 징옌 현을 가리킨다.[15]모식 표본은 부분적인 어깨뼈와 완전한 부리뼈, 앞다리 뼈, 완전한 궁둥뼈, 비교적 완전한 두개골, 그리고 설골을 포함한다. 부모식 표본은 세 개의 앞쪽 목뼈, 여러 개의 풍화된 등뼈, 거의 연결된 열을 포함하는 다양한 꼬리뼈, 비교적 완전한 뒷다리와 앞다리, 그리고 고립된 이빨을 포함한다. 세 번째 표본은 연결된 갈비뼈가 있는 목뼈 열, 4개의 융합된 천추, 여러 개의 꼬리뼈, 완전한 어깨뼈와 부리뼈, 다양한 사지 뼈, 그리고 고립된 이빨을 보존하고 있다.[15]

2. 3. 기타 마멘키사우루스 종 및 관련 자료
수년에 걸쳐 다른 종의 ''마멘키사우루스''가 명명되었다. 어떤 경우에는 다른 속의 종이 ''마멘키사우루스''로 이전되었지만, 이러한 이전의 유효성에 대한 이견이 있다. 이러한 종 중 일부는 단편적인 유해를 바탕으로 하며 진단할 수 없는 것으로 간주되었다. 다른 종들은 무효이거나 ''노멘 누둠''으로 간주된다.[3][16]1976년, 허우, 차오, 추는 새로운 속인 ''지공고사우루스 푸시엔시스''를 명명했다.[11][17] 상샤시마오 지층에서 최소 4개의 표본이 발견되었다. ''지공고사우루스''가 기술된 이후, 다른 연구자들은 이 속의 유효성에 대해 이견을 보였다. 1983년, 둥, 저우, 장은 일부 유해를 유사하게 명명된 ''오메이사우루스 푸시엔시스''에 할당했고, 나머지는 ''오메이사우루스 중시엔시스''에 할당했다. 장과 천은 1996년에 유해를 ''마멘키사우루스''에 ''M. 푸시엔시스''로 할당했다.[19] 리와 차이는 1997년에 이를 ''노멘 누둠''으로 간주했다.[11] 1999년, 발레리 마틴-롤란드(Valérie Martin-Rolland)는 ''지공고사우루스''를 유효한 속으로 간주했다. 왕과 동료들은 2019년에 이를 진단할 수 없는 것으로 간주했다.[3][26][16]
또 다른 종인 "마멘키사우루스 광위안엔시스"는 공식적으로 기술되지 않았으며, 이는 ''노멘 누둠''을 만든다. 상샤시마오 지층에서 유년기부터 성체에 이르기까지 여러 개체의 유해로 알려져 있다. 가장 큰 개체는 16m의 길이로 추정되었다. 이 종은 원래 1981년 장수핑의 논문에서 "오메이사우루스 광위안엔시스"로 기술되었지만,[34] 1997년 리와 차이는 이를 ''마멘키사우루스''의 종으로 등재했다. ''마멘키사우루스''와 마찬가지로, ''오메이사우루스''와 달리 이 종은 갈라진 신경 가시와 전강 꼬리 척추를 보여준다.[34] 왕과 동료들은 2019년에 이를 진단할 수 없는 것으로 간주했다.[3]
''M. 윈난엔시스''는 2004년 팡과 동료들에 의해 쓰촨 분지 윈난의 안닝 지층에서 기술되었다. 모식 표본은 분리된 앞다리, 뒷다리 및 골반 재료로 구성되었다.[20] 왕과 동료들은 2019년에 ''마멘키사우루스''에의 할당에 의문을 제기했다.[3]
1988년, 허와 동료들은 꼬리 척추의 특징을 바탕으로 ''오메이사우루스 창쇼우엔시스''가 ''마멘키사우루스''에 더 가깝다고 간주했다. 2019년, 탄과 동료들은 이러한 평가에 동의했다.[21] 1996년, 장과 천은 ''오메이사우루스 창쇼우엔시스''와 ''오메이사우루스 쿵지엔엔시스''를 ''마멘키사우루스''에 할당할 수 있다고 간주했다.[19] 그러나 2004년, 업처치는 ''O. 창쇼우엔시스''를 진단할 수 없다고 간주했다.[26]
1978년, 일본의 초기 백악기 시대의 미야코 군의 층에서 발견된 불완전한 용각류 상완골(NSM PV17656)은 아마도 ''마멘키사우루스''에 속하는 것으로 간주되었다. 1991년, 하세가와와 동료들은 이를 ''?마멘키사우루스 sp''에 할당했다.[22][23] 그러나 상완골은 1998년 아즈마 & 토미다와 2002년 바렛과 동료들에 의해 재평가되었다. 이들 저자는 상완골을 특정 용각류 그룹에 배치할 수 있는 구별되는 특징을 찾을 수 없었다. 그들은 이를 불확정 용각류로 간주했다.[24][25] 이 유해는 비공식적인 이름인 "모시류"가 주어졌다. "모시"는 그것이 발견된 장소의 지역 이름이고, "류"는 일본어로 용을 의미한다.[22]—또한 "모시사우루스"라고도 불린다.[23]
3. 형태
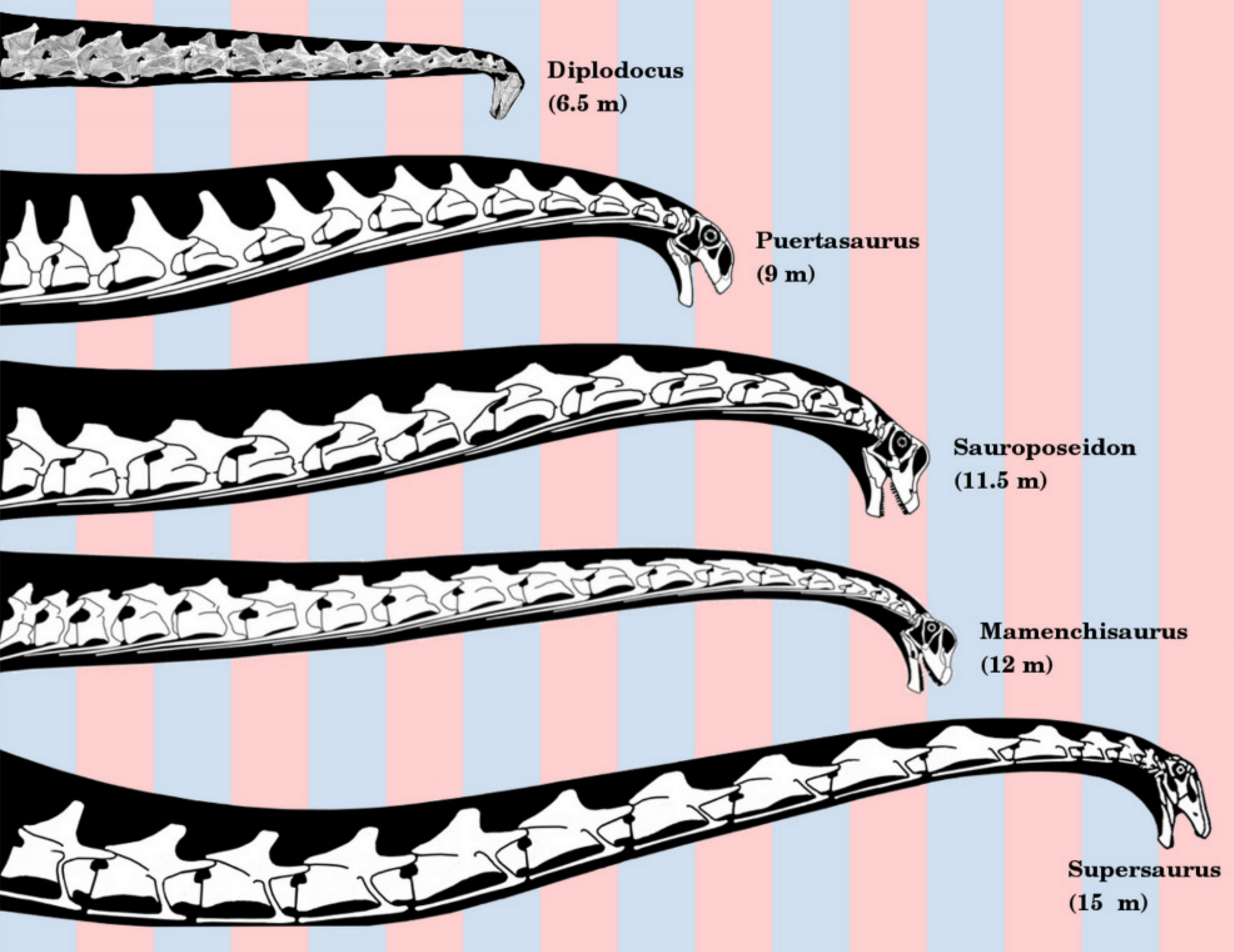
극도로 긴 목을 가진 용각류이다. 마멘키사우루스 속은 총 5종이 기재되어 있지만, 그 중 최대 종인 ''M. sinocanadorum''은 재연구 결과에 따르면 다른 종과는 다른 클레이드에 배치되어 오히려 최대급의 마멘키사우루스과 용각류인 신장티탄에 더 가깝다는 결과가 나왔다.[51] 종에 따라서는 전체 길이의 절반 이상을 목이 차지한다. 아시아 최대의 공룡 중 하나로 꼽힌다. 용각류의 경추는 15개 이하가 보통이지만 마멘키사우루스의 경추는 19개에 달하며,[52] 추안제사우루스와 함께 용각류 중에서도 가장 많다. 경추가 16개를 넘는, 특히 목이 긴 타입의 용각류 화석은 중국에서 많이 발견되고 있다.[52] 고환경 연구에 따르면, 당시 동남아시아에서는 키 큰 나무의 숲이 발달했다고 하며, 목이 길어진 것도 이러한 나무의 잎을 먹는 것에 대한 적응으로 보는 의견도 있었다. 그러나 경추의 구조는 마멘키사우루스가 그렇게 머리를 높이 들 수 없었음을 보여준다. 긴 목은 높은 곳의 잎을 먹기 위해서가 아니라, 먹이를 찾아 돌아다니는 에너지 손실을 최대한 줄이기 위해 목을 수평 방향으로 움직여 먹이가 되는 식물을 대량으로 얻기 위한 적응으로 생각된다.[52] 또한 긴 목은 성 선택 때문일 수도 있다.[53]
용각류는 대형화와 병행하여 목과 꼬리도 신장하는 경향을 보인다. 하지만 꼬리는 수각류나 조각류에도 길게 발달한 예가 있어, 용각류만의 특징이라고는 할 수 없다. 또한 극단적으로 긴 꼬리의 소유자는 용각류 중에서도 디플로도쿠스류에 한정된다. 하지만 목의 길이에서는 마멘키사우루스가 비할 데가 없다. 목의 신장을 진화로 본다면, 슈노사우루스처럼 소형이고 원시적인 용각류의 목이 짧은 것은 납득하기 쉽다. 그러나 디크라에오사우루스과는 진화 단계가 높음에도 불구하고 목은 비교적 짧다. 후기 쥐라기의 아르헨티나에 서식했던 브라키트라케로판은, 몸에 대한 목의 비율이 디크라에오사우루스과의 다른 종보다 40% 가까이 짧아, 간신히 땅에 닿는 정도이다. 반면 중기~후기 쥐라기의 중국에서는, 계통상의 관계에 관계없이 극도로 목이 긴 용각류가 집중적으로 분포했다. 용각류의 목에는 냉각 기관으로서의 역할도 있지만, 당시 중국만 유독 더웠다는 증거는 없다. 나무의 높이에 의한 것이 아니었다 하더라도, 역시 목의 장단에는 어떤 식생의 차이가 영향을 미치고 있다고 생각해야 할 것이다.
4. 고생태
마멘키사우루스는 네 발 걷기를 하고 몸집이 크며 머리가 작고 꼬리가 긴, 전형적인 용각류의 특징을 보인다. 마멘키사우루스과의 가장 독특한 특징은 전체 길이의 절반에 가까운 매우 긴 목이다.[4] 'M. youngi'와 'M. hochuanensis'의 모식 표본은 각각 18개와 19개의 척추뼈로 구성된 완전한 목을 보존하고 있다.[10][27] 마멘키사우루스의 경추는 길고 가볍게 구성되었으며 매우 공기화되어 있다.[46]
마멘키사우루스는 꼬리 중간 부분부터 디플로도쿠스류에서 볼 수 있는 것과 유사한 갈라진 쉐브론(또는 썰매 쉐브론)을 가지고 있는데, 이는 뒤로 강하게 굽어 앞으로 튀어나오게 한다.[35][36] 폴은 이러한 유형의 쉐브론이 일어선 행동과 관련된 적응이며, 삼각 자세에서 꼬리가 받침대 역할을 할 때 갈라진 쉐브론이 무게를 균등하게 분산시키는 데 도움이 된다고 주장했다. 또한 마멘키사우루스의 골반이 뒤집혀(기울어져) 있을 수 있으며, 이는 이족 보행을 하는 동안 느리게 걸을 수 있게 했을 수 있다고 지적한다.[36][37]
'M. hochuanensis'에 속할 수 있는 표본인 ZDM0126은 꼬리 끝 근처에 4개의 융합된 미추골을 가지고 있었다. 이 척추뼈는 확장된 신경궁과 더 높은 신경 가시를 가지고 있어 꼬리 곤봉을 나타낼 수 있다. 리다 싱과 동료들의 연구는 곤봉에 대한 유한 요소 분석을 수행했는데, 저자들은 곤봉이 방어 무기로서의 능력은 제한적이지만 감각 기관으로도 기능했을 수 있다고 결론지었다. 다른 중국 용각류인 '수노사우루스'와 '오메이사우루스'도 꼬리 곤봉을 가지고 있는 것으로 알려져 있지만, ZDM0126과는 모양이 다르다.[38][16]
스시구 형성에서 '마멘키사우루스' sp.로 언급된 길이 96cm의 큰 자뼈(GPIT SGP 2006/10)는 뼈 조직학 분석 결과, 사망 당시 나이가 43세로 추정되었다.[40]
4. 1. 서식 환경
마멘키사우루스는 극도로 긴 목을 가진 용각류이다. 마멘키사우루스 속은 총 5종이 기재되어 있지만, 종에 따라서는 전체 길이의 절반 이상을 목이 차지하기도 한다. 아시아 최대의 공룡 중 하나로 꼽힌다. 용각류의 경추는 보통 15개 이하이지만 마멘키사우루스의 경추는 19개에 달하며[52], 용각류 중에서도 가장 많다. 경추가 16개를 넘는, 특히 목이 긴 타입의 용각류 화석은 중국에서 많이 발견되고 있다[52]。고환경 연구에 따르면, 당시 동남아시아에서는 키 큰 나무의 숲이 발달했다고 하며, 목의 장대화도 이러한 나무의 잎을 먹는 것에 대한 대응으로 보는 의견도 있었다. 그러나 경추의 구조는 마멘키사우루스가 그렇게 머리를 높이 들 수 없었음을 보여준다. 긴 목은 높은 곳의 잎을 먹기 위해서가 아니라, 먹이를 찾아 돌아다니는 에너지 손실을 최대한 줄이기 위해 목을 수평 방향으로 움직여 먹이가 되는 식물을 대량으로 얻기 위한 적응으로 생각된다[52]。 또한 긴 목은 성 선택 때문일 수도 있다[53]。
용각류는 대형화와 병행하여 목과 꼬리도 신장하는 경향을 보이지만, 당시 중국만 유독 더웠다는 증거는 없다. 나무의 높이에 의한 것이 아니었다 하더라도, 목의 장단에는 어떤 식생의 차이가 영향을 미치고 있다고 생각해야 할 것이다.
4. 2. 식생 및 먹이 섭취
마멘키사우루스는 목이 매우 긴 용각류 공룡이다. 목 길이는 종에 따라 다르지만, 전체 길이의 절반 이상을 차지하는 경우도 있다. 경추 (목뼈)의 개수가 19개로, 일반적인 용각류(15개 이하)보다 많다.[52]긴 목은 높은 곳의 나뭇잎을 먹기 위한 적응으로 여겨졌지만, 경추 구조상 머리를 높이 들기는 어려웠을 것으로 보인다. 대신 목을 수평으로 움직여 넓은 범위의 식물을 섭취하는 데 유리했을 것이라는 견해가 있다.[52] 긴 목은 성 선택의 결과일 수도 있다.[53]
안드레아스 크리스티안과 동료들은 'M. youngi'의 목을 분석하여, 중립적인 자세에서 목이 거의 곧게 뻗어 있었고, 기저부에 약간의 위쪽 굴곡과 머리 쪽으로 약간의 아래쪽 굴곡이 있었음을 발견했다. 목의 기저부는 위쪽 유연성이 높지만 아래쪽은 제한적이었고, 머리 근처는 아래쪽 유연성이 좋고 위쪽은 낮았다. 중간 부위는 아래쪽 유연성이 높아, 'M. youngi'가 낮은 곳에서 자주 먹이를 먹었을 것으로 추정했다.[39]
당시 동남아시아에는 키 큰 나무들이 숲을 이루고 있었는데, 이는 긴 목의 진화와 관련이 있을 수 있다. 그러나 중국에서 발견되는 긴 목을 가진 용각류들은 목의 구조상 높은 곳의 먹이를 먹는 것이 아니라, 수평 방향으로 목을 움직여 최대한 에너지 손실을 줄이면서 많은 먹이를 얻기 위해 긴 목을 가지게 된 것으로 생각된다.[52]
4. 3. 타포노미 (화석화 과정)
마멘키사우루스의 화석에서는 초기 포유류(뾰족뒤쥐와 비슷했을 것으로 추정)의 이빨 자국이 발견되었다.[54] 또한 같은 장소에서 대형 수각류와 소형 수각류의 단리된 이빨(빠진 이빨)도 발견되었으며, 이러한 크고 작은 육식 동물들에 의해 마멘키사우루스의 시체가 분해되었을 것으로 보인다.[54]5. 분류
Mamenchisaurus|마멘키사우루스중국어는 수년에 걸쳐 여러 종이 명명되었다. 다른 속의 종이 Mamenchisaurus|마멘키사우루스중국어로 이전되기도 했지만, 이러한 이전의 유효성에 대해서는 이견이 존재한다.[3][16] 이들 중 일부 종은 단편적인 유해를 바탕으로 하여 진단이 불가능한 것으로 간주되기도 하고, 다른 종들은 무효이거나 ''노멘 누둠''으로 간주되기도 한다.[3][16]
1976년, 허우, 차오, 추는 새로운 속인 ''지공고사우루스 푸시엔시스''를 명명했다.[11][17] 상샤시마오 지층에서 최소 4개의 표본이 발견되었는데, 모식 표본(CV 02501[18])에는 두개골 재료, 상악, 치골 및 기저후두골이 포함되었다.[11][17] 추가 재료로는 등쪽 척추, 치골 및 좌골이 있다. ''지공고사우루스''가 기술된 이후, 다른 연구자들은 이 속의 유효성에 대해 이견을 보였다. 1983년, 둥, 저우, 장은 일부 유해를 유사하게 명명된 ''오메이사우루스 푸시엔시스''에 할당했고, 나머지는 ''오메이사우루스 중시엔시스''에 할당했다. 장과 천은 1996년에 유해를 ''마멘키사우루스''에 ''M. 푸시엔시스''로 할당했다.[19] 리와 차이는 1997년에 이를 ''노멘 누둠''으로 간주했다.[11] 1999년, 발레리 마틴-롤란드는 ''지공고사우루스''를 유효한 속으로 간주했다. 왕과 동료들은 2019년에 이를 진단할 수 없는 것으로 간주했다.[3][26][16]
"마멘키사우루스 광위안엔시스"는 공식적으로 기술되지 않아 ''노멘 누둠''이다. 상샤시마오 지층에서 유년기부터 성체에 이르기까지 여러 개체의 유해가 발견되었으며, 가장 큰 개체는 16m의 길이로 추정되었다. 이 종은 원래 1981년 장수핑의 논문에서 "오메이사우루스 광위안엔시스"로 기술되었지만,[34] 1997년 리와 차이는 이를 ''마멘키사우루스''의 종으로 등재했다. ''마멘키사우루스''와 마찬가지로, ''오메이사우루스''와 달리 이 종은 갈라진 신경 가시와 전강 꼬리 척추를 보여준다.[34] 왕과 동료들은 2019년에 이를 진단할 수 없는 것으로 간주했다.[3]
''M. 윈난엔시스''는 2004년 팡과 동료들에 의해 쓰촨 분지 윈난의 안닝 지층에서 기술되었다. 모식 표본은 분리된 앞다리, 뒷다리 및 골반 재료로 구성되었다.[20] 왕과 동료들은 2019년에 ''마멘키사우루스''로의 할당에 의문을 제기했다.[3]
1988년, 허와 동료들은 꼬리 척추의 특징을 바탕으로 ''오메이사우루스 창쇼우엔시스''가 ''마멘키사우루스''에 더 가깝다고 간주했다. 2019년, 탄과 동료들은 이러한 평가에 동의했다.[21] 1996년, 장과 천은 ''오메이사우루스 창쇼우엔시스''와 ''오메이사우루스 쿵지엔엔시스''를 ''마멘키사우루스''에 할당할 수 있다고 간주했다.[19] 그러나 2004년, 업처치는 ''O. 창쇼우엔시스''를 진단할 수 없다고 간주했다.[26]
1978년, 일본 초기 백악기 시대 미야코 군 지층에서 발견된 불완전한 용각류 상완골(NSM PV17656)은 아마도 ''마멘키사우루스''에 속하는 것으로 간주되었다. 1991년, 하세가와와 동료들은 이를 ''?마멘키사우루스 sp''에 할당했다.[22][23] 그러나 상완골은 1998년 아즈마 & 토미다와 2002년 바렛과 동료들에 의해 재평가되었다. 이들 저자는 상완골을 특정 용각류 그룹에 배치할 수 있는 구별되는 특징을 찾을 수 없었고, 불확정 용각류로 간주했다.[24][25] 이 유해에는 "모시류"라는 비공식적인 이름이 붙여졌는데, "모시"는 발견 장소의 지역 이름이고, "류"는 용을 의미한다.[22] "모시사우루스"라고도 불린다.[23]
''마멘키사우루스''는 때때로 '쓰레기통 분류군'으로 언급되며, 연구자들은 이 속에 할당된 종의 수와 파편적인 유해에 대해 의문을 제기한다.[5][41][26][46][42] 이 속은 계통 발생 관계를 이해하기 어렵게 만드는, 점점 더 혼란스러운 분류법으로 제대로 정의되지 않았다. 여러 분석에서 ''마멘키사우루스''가 단 계통군임을 보여주지 못했으며, 이 속을 수정해야 할 필요성을 시사한다.[46] 모식종인 ''M. constructus''에 대한 추가 연구가 이 속을 더 잘 이해하기 위해 필요하다.[26]
''M. constructus''가 처음 기술되었을 때, 영은 갈매기뼈가 디플로도쿠스과와의 친연성을 나타낸다고 언급했지만, 정확한 위치에 대해서는 확신하지 못했다.[7] 1958년, 영은 ''마멘키사우루스''를 티타노사우루스과에 할당했다.[9] ''M. hochuanensis''의 기술과 함께, 영과 자오는 1972년에 마멘키사우루스과를 만들었다.[10] 1978년, ''마멘키사우루스''의 두개골이 알려지지 않았을 때, 버먼과 맥킨토시는 분기된 갈매기뼈와 같은 디플로도쿠스류의 척추 특징을 기반으로 이 속을 디플로도쿠스과에 할당했다. 1990년, 맥킨토시는 ''마멘키사우루스''를 디플로도쿠스과 내 마멘키사우루스아과에 할당했다.[23]
1995년 업처치의 분석은 ''마멘키사우루스''를 유헬로포두스과에서 발견했다. 유헬로포두스과는 먼저 명명되었으므로 마멘키사우루스과보다 우선순위를 갖는다.[43] 이후 여러 분석에서는 ''유헬로푸스''가 더 멀리 관련된 마크로나리아로 밝혀졌으며, ''마멘키사우루스''는 네오사우로포다 바로 바깥에 있는 마멘키사우루스과에 속한다.[41][46]
2011년 세키야와 2020년 무어와 동료들의 분석에서는 ''M. constructus, M. hochuanensis,'' ZDM 0126 ''(M. hochuanensis'' 관련''), M. sinocandadorum, 및 M. youngi''를 개별적으로 취급했다.[46][44] 무어와 동료들의 분석에 따르면 ''M. constructus''의 위치는 불확실했는데, 아마도 설명에 담긴 특징 정보가 제한적이기 때문일 것이다. 사용된 데이터 세트에 따라 ''유헬로포두스''는 다른 연구에서 발견된 것처럼 마크로나리아 내에 있거나, 네오사우로포다 외부에 전통적인 위치에 있을 것이며, 다른 ''마멘키사우루스'' 유사 분류군과 함께 그룹화될 것이다. 후자의 시나리오는 마멘키사우루스류를 유헬로포두스과의 구성원으로 만들 것이다.[46] 세키야(2011)와 무어와 동료들(2020)의 분석에서는 ZDM 0126이 ''M. hochuanensis''의 모식표본의 자매 분류군으로 회복되지 않아, 이 종에 대한 언급에 의문을 제기했다.[46][44]
아래의 분지학적 나무는 2008년 알랑과 아퀘스비의 용각류 내에서 이 속의 가능한 계통 발생 위치를 보여준다:[45]
{{clade| style=font-size:90%;line-height:90%
|label1=용각류
|1={{clade
|1=''안테토니트루스''
|2={{clade
|1=''공시안사우루스''
|2={{clade
|1=''이사노사우루스''
|label2=그라비사우리아
|2={{clade
|label1=불카노돈과
|1={{clade
|1=''불카노돈''
|2=''타주다사우루스''
}}
|label2=진용각류
|2={{clade
|1=''바라파사우루스''
|2=''파타고사우루스''
|3={{clade
|label1=오메이사우루스과
|1={{clade
|1=''오메이사우루스''
|2='''''마멘키사우루스'''''
}}
|2=네오사우로포다
}}
}}
}}
}}
}}
}}
아래 두 개의 계통수는 곤잘레스 리가 데이터 세트의 함축 가중치 및 베이지안 분석과 같이 무어와 동료들이 가장 호의적으로 간주한 두 분석에서 유헬로포두스과/마멘키사우루스과의 내부 관계를 보여준다.[46]
'''토폴로지 A:''' 함축 가중치 분석, 곤잘레스 리가 데이터 세트[46]
{{clade|{{clade
|1=''톈산사우루스 치타이엔시스''
|2={{clade
|label2=핵심 ''마멘키사우루스'' 유사 분류군
|1=''오메이사우루스 융시엔시스''
|2={{clade
|1=''왐웨라카우디아 케란제이''
|2={{clade
|1={{clade
|1=''치쟝롱 구오크르''
|2='''''마멘키사우루스 시노칸나도룸'''''}}
|2={{clade
|1={{clade
|1=''추안지에사우루스 아나엔시스''
|2=''아날롱 추안지에엔시스''}}
|2={{clade
|1='''''마멘키사우루스 호추안엔시스''''' (모식표본)
|2='''''마멘키사우루스 호추안엔시스''''' (관련)
|3={{clade
|1='''''마멘키사우루스 영이'''''
|2={{clade
|1=시슈고우 경추 척추
|2={{clade
|1=푸 크라둥 분류군
|2=''클라멜리사우루스 고비엔시스''}} }} }} }} }} }} }}|style=font-size:90%; line-height:90%;|label1=마멘키사우루스과}}
'''토폴로지 B:''' 시간 보정 베이지안 분석, 곤잘레스 리가 데이터 세트[46]
{{clade|{{clade
|1={{clade
|1=''오메이사우루스 융시엔시스''
|2={{clade
|1=''세티오사우리스쿠스 스튜어티''
|2={{clade
|1=''오메이사우루스 마오이아누스''
|2=''오메이사우루스 톈푸엔시스''}} }} }}
|2={{clade
|label2=핵심 ''마멘키사우루스'' 유사 분류군
|1=''톈산사우루스 치타이엔시스''
|2={{clade
|1={{clade
|1=''추안지에사우루스 아나엔시스''
|2={{clade
|1=''아날롱 추안지에엔시스''
|2={{clade
|1=''치쟝롱 구오크르''
|2={{clade
|1=시슈고우 경추 척추
|2='''''마멘키사우루스 시노칸나도룸'''''}} }} }} }}
|2={{clade
|1={{clade
|1={{clade
|1='''''마멘키사우루스 호추안엔시스''''' (모식표본)
|2='''''마멘키사우루스 콘스트럭투스'''''}}
|2={{clade
|1=푸 크라둥 분류군
|2=''클라멜리사우루스 고비엔시스''}} }}
|2={{clade
|1='''''마멘키사우루스 호추안엔시스''''' (관련)
|2='''''마멘키사우루스 영이'''''
|3={{clade
|1=''왐웨라카우디아 케란제이''
|2={{clade
|1={{clade
|1=''시안산사우루스 시지아고우엔시스''
|2=''다시아티탄 빙링이''}}
|2={{clade
|1=''유헬로푸스 즈단스키''
|2=''동베이티탄 동기''}} }} }} }} }} }} }}|style=font-size:90%; line-height:90%;|label1=유헬로포두스과}}
2023년 ''M. sinocanadorum''의 재기술은 그것과 ''신장티탄'' 사이에 일관된 자매 분류군 관계를 발견했으며, ''후디에사우루스''와 관련 자료 ''M. hochuanensis''(ZDM 0126)도 일관된 친척으로 밝혀졌다.[12]
{{clade|style=font-size:90%; line-height:90%;
|label1=마멘키사우루스과
|1={{clade
|1=''톈산사우루스''
|2={{clade
|1=''오메이사우루스 융시엔시스''
|2={{clade
|1=''왐웨라카우디아''
|2={{clade
|1=''치쟝롱''
|2={{clade
|1='''''마멘키사우루스 콘스트럭투스'''''
|2=''벨루사우루스''
|3={{clade
|1={{clade
|1=''로말레오파쿠스''
|2={{clade
|1=''추안지에사우루스''
|2=''아날롱'' }} }}
|2={{clade
|1='''''마멘키사우루스 영이'''''
|2={{clade
|1={{clade
|1='''''마멘키사우루스 호추안엔시스''''' (모식표본)
|2={{clade
|1=시슈고우 경추 척추
|2={{clade
|1=푸 크라둥 분류군
|2=''클라멜리사우루스'' }} }} }}
|2={{clade
|1={{clade
|1='''''마멘키사우루스 시노칸나도룸'''''
|2=''신장티탄'' }}
|2={{clade
|1=''후디에사우루스''
|2={{clade
|1=''다아노사우루스''
|2='''''마멘키사우루스 호추안엔시스''''' (관련) }} }} }} }} }}
}} }} }} }} }} }}
마멘키사우루스는 이전에 디플로도쿠스과에 분류되었던 적이 있는데, 이는 거의 일반적인 특징을 가진 용각류라는 점과 두개골이 발견되지 않았다는 점에 기인한다. 그러나 최근 두개골이 갖춰진 골격이 발견되었고, 그 오메이사우루스나 카마라사우루스와도 유사한 앞뒤로 짧은 특징을 보임에 따라 새롭게 분류하게 되었다. 최근 연구에서는 디플로도쿠스, 카마라사우루스와 같은 북미의 유명한 용각류(신용각류)의 조상과 초기에 분기된 원시적인 계통의 일원으로 여겨지고 있다.
6. 고연대
마멘키사우루스는 원래 중기~후기 쥐라기에 걸쳐 서식한 것으로 생각되었다.[47][3] 그러나 많은 마멘키사우루스 종이 발견되는 상부 샤시먀오 층군에 대한 신뢰할 만한 연대 측정 자료는 없다.[3]
2018년에 발표된 한 연구에서는 기존에 중기 쥐라기에 속한다고 여겨졌던 하부 샤시먀오 층군의 오메이사우루스 화석이 발견된 지층에 우라늄-납 연대 측정법을 사용했다. 그러나 방사성 연대 측정 결과 하부 샤시먀오 층군은 후기 쥐라기, 옥스포드절에 해당하며 약 1억 5900만 년 전으로 밝혀졌다. 이 발견은 상부 샤시먀오 층군의 마멘키사우루스 화석이 발견된 암석이 더 젊다는 것을 시사하며, 옥스포드절보다 오래되지 않았음을 의미한다.[3][48]
''M. 시노카나도룸''은 중부~상부 시슈고우 층군에서 발견되었다. 방사성 연대 측정 결과 이 지층은 옥스포드절에 해당하며, 1억 5870만 년에서 1억 6120만 년 전으로 추정된다.[3][48] ''M. 아니우엔시스''는 원래 중기~후기 쥐라기로 여겨졌던 수이닝 층군에서 발견되었다. 2019년 연구에 따르면 이 암석은 초기 백악기, 압트절에 해당하며 평균 연령은 약 1억 1440만 년 전이다. 이는 마멘키사우루스과가 이전에 생각했던 것보다 약 3천만 년 더 오래 생존했을 수 있음을 나타낸다.[3]
참조
[1]
서적
The Complete Dinosaur
https://archive.org/[...]
Indiana University Press
[2]
서적
The Dinosauria
https://archive.org/[...]
University of California Press
[3]
간행물
Surprisingly young age for the mamenchisaurid sauropods in South China
[4]
문서
A large mamenchisaurid from the Junggar Basin, Xinjiang, People Republic of China.
1993
[5]
서적
The Princeton Field Guide to Dinosaurs
Princeton University Press
2016
[6]
간행물
Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals
http://www.gspauldin[...]
2019
[7]
문서
On a new sauropod from Yiping, Szechuan, China.
1954
[8]
웹사이트
Origin of the Mamenchisaurus name
http://www.bmnh.org.[...]
2007-09-27
[9]
간행물
New Sauropods from China
1958
[10]
문서
Mamenchisaurus hochuanensis sp. nov.
1972
[11]
서적
Dinosaurs: The Encyclopedia: Supplement 1
Jefferson: McFarland & Company
[12]
간행물
Re-assessment of the Late Jurassic eusauropod Mamenchisaurus sinocanadorum Russell and Zheng, 1993, and the evolution of exceptionally long necks in mamenchisaurids
https://www.tandfonl[...]
[13]
간행물
Determining the Largest Known Land Animal: A Critical Comparison of Differing Methods for Restoring the Volume and Mass of Extinct Animals
http://dx.doi.org/10[...]
2019-12-31
[14]
간행물
'A new species of sauropod, Mamenchisaurus anyuensis sp. nov.'
http://www.paleoglot[...]
[15]
간행물
'A new species of sauropod from the Late Jurassic of the Sichuan Basin (''Mamenchisaurus jingyanensis'' sp. nov.)'
[16]
간행물
Structure, orientation and finite element analysis of the tail club of Mamenchisaurus hochuanensis.
[17]
간행물
New discovery of sauropod dinosaurs from Sichuan
http://www.ivpp.cas.[...]
1976
[18]
간행물
Jurassic dinosaur faunas in Zigong
https://www.yumpu.co[...]
2005
[19]
서적
The Continental Jurassic
Museum of Northern Arizona
[20]
간행물
Discovery of Late Jurassic Mamenchisaurus in Yunnan, southwestern China
2004
[21]
간행물
Discovery of Omeisaurus (Dinosauria: Sauropoda) in the Middle Jurassic Shaximiao Formation of Yunyang, Chongqing, China
2019
[22]
간행물
A diplodocid dinosaur from the Early Cretaceous Miyako group of Japan
1991
[23]
서적
Dinosaurs, the encyclopedia
https://www.worldcat[...]
McFarland & Co
1997
[24]
문서
Japanese dinosaurs. In: Curie PJ, Padian K, eds. Encyclopaedia of dinosaurs. San Diego: Academic Press, 375–379.
1998
[25]
간행물
Barrett, Paul M., et al. "Sauropod dinosaurs from the Lower Cretaceous of eastern Asia: taxonomic and biogeographical implications
2002
[26]
서적
The Dinosauria
https://www.worldcat[...]
University of California Press
[27]
문서
A new species of sauropod from Zigong, Sichuan, Mamenchisaurus youngi. 87–91. In DEPARTMENT OF SPATIAL PLANNING AND REGIONAL ECONOMY (ed.), Publication in Geoscience Contributed to the 30th International Geological Congress. China Economic Publishing House, Beijing.
1996
[28]
간행물
The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world's largest dinosaurs
[29]
간행물
Biology of the sauropod dinosaurs: the evolution of gigantism
2011
[30]
간행물
A review and reappraisal of the specific gravities of present and past multicellular organisms, with an emphasis on tetrapods
[31]
간행물
Re-assessment of the Late Jurassic eusauropod dinosaur Hudiesaurus sinojapanorum Dong, 1997, from the Turpan Basin, China, and the evolution of hyper-robust antebrachia in sauropods
2021-07-04
[32]
간행물
Why sauropods had long necks; and why giraffes have short necks
2013-02-12
[33]
문서
"Osteology, paleobiology, and relationships of the sauropod dinosaur ''Sauroposeidon''"
2000
[34]
문서
'The First Mamenchisaurian Skeleton with Complete Skull: Mamenchisaurus youngi (in Chinese with English summary).'
Sichuan Science and Technology Press, Chengdu
2002
[35]
간행물
Sauropod haemal arches: morphotypes, new classification and phylogenetic aspects
https://doi.org/10.1[...]
2012-06-01
[36]
서적
The Sauropod Dinosaurs : Life in the Age of Giants
https://www.worldcat[...]
Johns Hopkins University Press
[37]
논문
Restoring Maximum Vertical Browsing Reach in Sauropod Dinosaurs
2017
[38]
논문
New material of ''Mamenchisaurus hochuanensis'' from Ziging China
[39]
논문
Biomechanical Reconstructions and Selective Advantages of Neck Poses and Feeding Strategies of Sauropods with the Example of Mamenchisaurus youngi
2013-10-30
[40]
서적
Biology of the Sauropod Dinosaurs : Understanding the Life of Giants.
https://www.worldcat[...]
Indiana University Press
[41]
논문
A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids
http://www.tandfonli[...]
2015-01-02
[42]
논문
New sauropod material from the Late Jurassic part of the Shishugou Formation (Junggar Basin, Xinjiang, NW China)
http://www.schweizer[...]
2011-11-01
[43]
논문
The evolutionary history of sauropod dinosaurs
http://doc.rero.ch/r[...]
1995
[44]
문서
Sekiya, T. (2011). Re-examination of ''Chuanjiesaurus anaensis'' (Dinosauria: Sauropoda) from the Middle Jurassic Chuanjie Formation, Lufeng County, Yunnan Province, southwest China." ''Memoir of the Fukui Prefectural Dinosaur Museum'', '''10''': 1-54.
[45]
논문
Anatomy and phylogenetic relationships of Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco
http://cat.inist.fr/[...]
2021-03-13
[46]
논문
Osteology of ''Klamelisaurus gobiensis'' (Dinosauria, Eusauropoda) and the evolutionary history of Middle–Late Jurassic Chinese sauropods
https://discovery.uc[...]
[47]
논문
Dinosaur assemblages from the middle jurassic shaximiao formation and chuanjie formation in the sichuan-yunnan basin, China
2011
[48]
논문
Age of Jurassic basal sauropods in Sichuan, China: A reappraisal of basal sauropod evolution
https://pubs.geoscie[...]
2018-09-01
[49]
문서
학명의 읽는 방법은 연구자에 따라서도 라틴어 읽기나 영어 읽기 등 복수의 발음이 혼재하고 있는 경우가 많고, 일본어에 전사할 때는 혼란하기 쉽다. 학명은 본래 라틴어로 붙여지기 때문에, 고전 라틴어 읽기를 정식으로 하는 연구자나, (사어이며 모어 화자가 없는 라틴어의) 올바른 읽는 방법은 정의할 수 없다는 상대주의자도 적지 않다. 그렇지만 중국어로 모음의 길이를 구별하지 않고, 마멘치ー 아니면 마멘치라고 불리는 지역에서 발굴되었기 때문에, 마멘치사우루스가 지명의 발음에 충실하다고 할 수 있다.
[50]
논문
Surprisingly young age for the mamenchisaurid sauropods in South China
https://www.scienced[...]
2019-12-01
[51]
논문
Re-assessment of the Late Jurassic eusauropod Mamenchisaurus sinocanadorum Russell and Zheng, 1993, and the evolution of exceptionally long necks in mamenchisaurids
https://www.tandfonl[...]
2023-01-01
[52]
서적
絶滅したふしぎな巨大生物
PHP研究所
2011-06-10
[53]
문서
Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation(Phil Senter:2007)
[54]
문서
The smallest eating the largest: the oldest mammalian feeding traces on dinosaur bone from the Late Jurassic of the Junggar Basin (northwestern China)(Felix J. Augustin:2020)
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com