카마라사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
카마라사우루스는 후기 쥐라기에 살았던 중형 용각류 공룡으로, 카마라사우루스과에 속한다. 1877년 콜로라도에서 척추뼈 화석이 처음 발견되었으며, 에드워드 드링커 코프에 의해 명명되었다. 가장 흔한 종인 C. lentus는 최대 15m, C. supremus는 최대 18~23m까지 성장했다. 튼튼한 두개골과 주걱 모양의 이빨을 가지고 있으며, 연조직의 흔적도 발견되었다. 미국 서부의 초원에서 무리 생활을 했을 것으로 추정되며, 알로사우루스와 같은 육식 공룡의 공격을 받았을 가능성이 있다.
더 읽어볼만한 페이지
- 1877년 기재된 화석 분류군 - 캄프소사우루스
캄프소사우루스는 백악기 후기부터 팔레오세에 북아메리카와 유럽에 서식한 코리스토데라목 파충류로, 긴 주둥이와 유선형 몸체를 가진 수생 생물이며 1877년 코프에 의해 발견되었고 7종이 알려져 있다. - 1877년 기재된 화석 분류군 - 스테고사우루스
스테고사우루스는 쥐라기 후기에 살았던 초식 공룡으로, 등에 두 줄로 솟은 골판과 꼬리 끝의 골침이 특징이며, 골판 배열과 기능, 꼬리 가시 용도에 대한 논쟁이 있고 대중문화에 큰 영향을 미쳐 콜로라도주의 상징 공룡으로 지정되기도 했다. - 모리슨층의 공룡 - 마르쇼사우루스
마르쇼사우루스는 1976년 미국 콜로라도주에서 발견된 몸길이 약 2.5미터의 작은 수각류 공룡으로, 가볍고 민첩한 골격과 모리슨 층에서의 발견, 그리고 장골 변형 등의 특징을 통해 행동 및 생태 연구가 진행되었으며, 초기에는 메갈로사우루스류, 이후에는 피아트니츠키사우루스과와의 관계 연구가 이루어졌다. - 모리슨층의 공룡 - 드리오사우루스
쥐라기 후기 북아메리카에 서식한 드리오사우루스는 긴 목과 다리, 각질 부리를 가진 최대 3m 크기의 조각류 공룡으로, 모리슨 층 등 다양한 지역에서 화석이 발견되었으며, 1894년에 'Dryosaurus altus'로 재분류되었다. - 쥐라기 후기 북아메리카의 공룡 - 마르쇼사우루스
마르쇼사우루스는 1976년 미국 콜로라도주에서 발견된 몸길이 약 2.5미터의 작은 수각류 공룡으로, 가볍고 민첩한 골격과 모리슨 층에서의 발견, 그리고 장골 변형 등의 특징을 통해 행동 및 생태 연구가 진행되었으며, 초기에는 메갈로사우루스류, 이후에는 피아트니츠키사우루스과와의 관계 연구가 이루어졌다. - 쥐라기 후기 북아메리카의 공룡 - 드리오사우루스
쥐라기 후기 북아메리카에 서식한 드리오사우루스는 긴 목과 다리, 각질 부리를 가진 최대 3m 크기의 조각류 공룡으로, 모리슨 층 등 다양한 지역에서 화석이 발견되었으며, 1894년에 'Dryosaurus altus'로 재분류되었다.
| 카마라사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Camarasaurus |
| 명명자 | 코프, 1877a |
| 어원 | 고대 그리스어 καμαρα(kamara, 방) + σαυρος(sauros, 도마뱀) |
| 속 | 카마라사우루스속 |
| 명명 연도 | 1877년 |
| 타입 종 | 카마라사우루스 수프레무스 |
| 타입 종 명명자 | 코프, 1877a |
| 시대 | 쥐라기 후기 |
| 생존 시기 | 백만 년 전 |
| 동의어 | Caulodon (코프, 1877) Morosaurus (마시, 1878) Uintasaurus (홀랜드, 1924) 카테토사우루스? (젠슨, 1988) |
| 분류 | |
| 상목 | 공룡상목 |
| 목 | 용반목 |
| 아목 | 용각아목 |
| 하목 | 용각하목 |
| 과 | 카마라사우루스과 |
| 아과 | 카마라사우루스아과 |
| 종 | |
| 종 목록 | 카마라사우루스 그란디스 (마시, 1877) 길모어, 1925 카마라사우루스 렌투스 (마시, 1889) 묵, 1914 카마라사우루스 레위시? (젠슨, 1988) 밀러, 매킨토시, 슈타트만 & 길레트, 1992 |
2. 역사
1877년 오라멜 W. 루카스가 콜로라도주에서 카마라사우루스의 흩어진 척추뼈 몇 개를 발견했다. 고생물학자 에드워드 드링커 코프는 오트니엘 찰스 마시와의 치열한 경쟁(화석 전쟁) 중에 이 단편적인 화석을 구입하여 같은 해에 카마라사우루스라고 명명하고 기재했다.
1925년 찰스 W. 길모어는 처음으로 카마라사우루스의 완전한 골격을 복원했다. 그러나 이 골격은 어린 개체의 것이었기 때문에, 오늘날 알려진 것보다 훨씬 작은 공룡으로 묘사되었다.
로키 산맥 동쪽 면을 따라 위치한 모리슨 층에서는 쥐라기 후기의 암석이 풍부하게 발견된다. 이곳에서는 카마라사우루스를 비롯하여 디플로도쿠스, 아파토사우루스, 브라키오사우루스 등 매우 다양한 공룡 종들이 발견되었다. 그중에서도 카마라사우루스는 가장 많이 발견되는 공룡이며, 콜로라도 주, 뉴멕시코 주, 유타 주, 와이오밍 주에서 완전한 골격이 발견되었다.
2. 1. 최초 발견
1877년 봄, 콜로라도주 캐넌시티의 오라멜 윌리엄 루카스가 콜로라도주 가든 파크에서 몇 개의 큰 척추뼈를 발견한 것이 최초의 ''카마라사우루스'' 기록이다. 그는 이 척추뼈를 펜실베이니아주 필라델피아에 있던 에드워드 드링커 코프에게 보냈다.[1] 처음에 보내진 자료는 부분적인 경추 1개, 등쪽 척추뼈 3개, 꼬리 척추뼈 4개였으며, 이 경추가 분류군의 이름이 되었다.[3][1] 이 표본은 현재 여러 개체의 것으로 여겨진다.[3] 코프는 이러한 초기 단편적인 유해를 바탕으로 ''카마라사우루스 수프레무스''(“최고의 방뼈 도마뱀”)를 처음 기술하고 속을 설립했다. 이 유해는 현재 미국 자연사 박물관에 AMNH 560이라는 목록 번호로 소장되어 있다.[3] 코프는 원래 뼈를 받은 후, 수집가들을 고용하여 더 많은 자료를 수집했고, 이 자료는 1921년 헨리 페어필드 오스본과 찰스 무크에 의해 기술되었다.[3]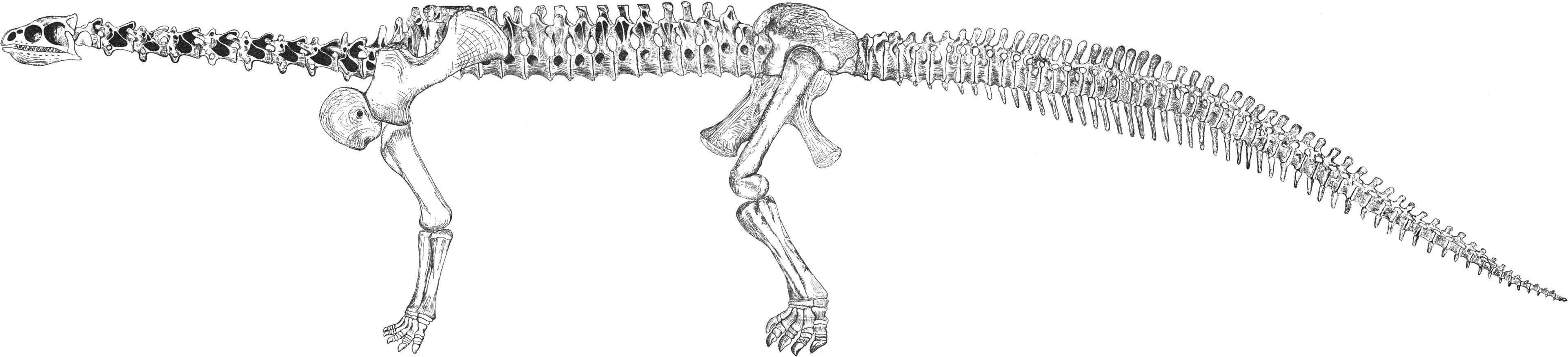
자료의 양은 방대하여 여러 개의 뒤섞인 부분 골격으로 구성되었다.[3] 한 번에 모두 준비된 것은 아니지만, 1870년대에서 1890년대까지 코프의 지시 아래 야곱 가이즈마르가 상당량을 정리했다.[3] 1877년, 존 라이더 박사는 코프 교수의 지시 아래 여러 캔버스에 ''카마라사우루스''의 골격 재구성을 그렸고, 코프는 이 그림들을 강연에 사용하여 청중에게 깊은 인상을 주었다.[3] 이 재구성은 용각류 공룡의 최초 재구성이었으며, 실물 크기였고 여러 개체의 유해를 나타냈다. 재구성의 길이는 50피트가 넘었다. 코프의 수집가들은 1877년부터 1878년까지 더 많은 자료를 보냈고,[3][2] 코프는 더 많은 자료를 얻으면서 새로 보내진 유해를 기반으로 분류군 이름을 지었다. 이러한 추가 분류군 중 대부분은 현재 의심스럽거나 ''카마라사우루스''와 동의어로 간주된다.[2] 가든 파크에서 수집이 끝날 무렵, 골격의 거의 모든 부분에서 최소 4개체와 수백 개의 뼈가 발견되었다.[3]
최초의 발견 기록은 1877년에 오라멜 W. 루카스에 의해 몇 개의 흩어진 척추뼈가 콜로라도주에서 발견된 것이다. 고생물학자 에드워드 드링커 코프는 오트니엘 찰스 마시와의 길고 신랄한 경쟁 (화석 전쟁으로 알려짐) 중에 이 단편적인 화석을 구입하여 같은 해에 명명 및 기재했다.
1925년, 찰스 W. 길모어에 의해 처음으로 완전한 골격이 복원되었다. 그러나 이 골격은 어린 개체의 것이었기 때문에, 오늘날 알려진 것보다 훨씬 작은 공룡으로 묘사되었다.
2. 2. 코모 블러프에서의 발견과 모로사우루스
1877년 후반, 오스니얼 찰스 마시를 위해 일하는 팀이 미국 와이오밍 주의 코모 블러프 채석장 1에서 단편적인 뒤쪽 두개골과 부분적인 몸통 뼈대를 발견하여 수집했다. 이 골격은 당시까지 가장 잘 보존된 단일 개체 ''카마라사우루스''였으며, 1877년에 새로운 종의 ''아파토사우루스''로 명명되었다. 이 표본은 1879년에 완전히 수집되었으며, 어린 개체의 골격 대부분(모식 표본 YPM 1901)을 포함하고 있다.[4] 한편, 에드워드 코프를 위해 가든 파크에서 일하는 팀은 대퇴골과 꼬리뼈 2개로 구성된 조각난 표본을 수집했으며, 코프는 이를 새로운 종의 ''암피코엘리아스''로 만들어 1877년에 ''암피코엘리아스 라투스''(Amphicoelias latus)라고 명명했다.[5] 이 종은 1921년에 잠정적으로 ''C. 수프레무스''(C. supremus)와 동의어로 여겨졌다. 1998년, 케네스 카펜터는 이 발견의 지층 위치를 볼 때 ''C. 그란디스''(C. grandis)와 동의어일 가능성이 더 높다고 주장했지만, 2005년 ''카마라사우루스''의 생물층서학적 분포에 대한 연구에서, 타케히토 이케지리는 이를 ''C. 수프레무스''와 동의어로 유지했다.1878년, 코모 블러프에서 다른 엉성한 용각류 몸통 뼈대 요소들과 함께 용각류 천골이 발견되었다. 유해는 마시에게 보내졌고, 1878년에 천골은 새로운 속과 종으로 지정되어 ''모로사우루스 임파르''(Morosaurus impar, "짝이 없는 어리석은 도마뱀")가 되었다. ''모로사우루스''는 19세기 후반에 걸쳐 여러 새로운 종을 받았으며, 심지어 1892년에는 새로운 과인 모로사우루스과의 일부가 되었다. 현재 ''모로사우루스'' 종의 대부분은 의심스럽거나[29] 재분류된 것으로 간주된다.[29] 1889년, 코모 블러프에서 발견된 부분 두개골과 골격을 바탕으로 새로운 종의 ''모로사우루스''가 명명되었다. ''모로사우루스 렌투스''(Morosaurus lentus)는 골격(모식 표본 YPM 1910)에 주어진 이름이었으며, 이 골격은 1930년에 예일 피바디 박물관 화석 전시관에 전시되었다.
2. 3. 20세기 이후의 발견
1890년대 후반, 미국 자연사 박물관과 필드 자연사 박물관은 코모 절벽과 프루이타 (콜로라도)에서 추가적인 ''모로사우루스''(Morosaurus) 자료를 발견했다. 이 자료들은 주로 사지로 구성되어 용각류의 손과 발 구조를 새롭게 재구성하는 데 기여했다.[6][7] 1899년, 미국 자연사 박물관은 와이오밍 주의 본 캐빈 채석장에서 최초로 완전한 ''카마라사우루스''(Camarasaurus) 두개골과 턱뼈, 그리고 관련된 목 척추를 발견했다.[8]1901년, 엘머 S. 리그스는 ''모로사우루스''(Morosaurus)와 ''카마라사우루스''(Camarasaurus)에 대한 주요 재평가를 통해 마르시가 명명한 다섯 종의 ''모로사우루스''(Morosaurus) 중 세 종만이 유효하다고 판단했다.[7] ''모로사우루스 그란디스''(Morosaurus grandis), ''모로사우루스 렌투스''(Morosaurus lentus), ''모로사우루스 아질리스''(Morosaurus agilis) (현재는 ''스미타노사우루스(Smitanosaurus)''로 알려짐)가 유효한 종으로 인정되었고, ''모로사우루스 임파르''(Morosaurus impar)는 ''M. 그란디스''(M. grandis)와 같은 종으로 간주되었다. 리그스는 또한 ''모로사우루스''(Morosaurus)와 ''카마라사우루스''(Camarasaurus)가 같은 종일 가능성을 제기했다.[7] 1905년, 미국 자연사 박물관에서 최초의 용각류 골격 표본이 ''브론토사우루스''(Brontosaurus)로 조립되었는데, 이 두개골은 코모 절벽에서 발견된 ''카마라사우루스''(Camarasaurus) 자료를 기반으로 한 것이었다.[9]
카네기 자연사 박물관은 1909년에 거의 완전한 어린 개체의 골격(표본 번호 CM 11338)을 발견했는데, 이는 사망 자세 그대로 연결된 상태로 발견되어 박물관 홀에 전시되어 있다. 이 표본은 찰스 W. 길모어에 의해 1925년에 ''카마라사우루스 렌투스''(Camarasaurus lentus)로 분류되었으며, 취약한 목 척추를 포함하여 거의 모든 뼈가 연결된 상태로 보존되었다.
1918년, 공룡 국립 기념지에서 카네기 직원들이 발견한 또 다른 ''카마라사우루스''(Camarasaurus) 골격(USNM V 13786)은 국립 자연사 박물관으로 옮겨져 1936년 텍사스 센테니얼 엑스포지션에서 전시되었다.[10] 1947년까지 준비 작업이 계속되었고, 골격은 화석 홀에 사망 자세로 조립되었다. 이 ''카마라사우루스''(Camarasaurus) 역시 ''C. 렌투스''(C. lentus)로 분류되었다.
1919년, W. J. 홀랜드는 카네기 박물관이 1909년에 발견한 공룡 국립 기념지의 또 다른 용각류 표본을 바탕으로 ''우인타사우루스 도글라시''(Uintasaurus douglassi)를 명명했다.[11] 이 표본은 5개의 앞쪽 목 척추로 구성되어 있고,[11] ''카마라사우루스 렌투스''(Camarasaurus lentus)와 같은 종이다. 1940년대에는 오클라호마 서부의 블랙 메사 근처에서 발견된 ''카마라사우루스''(Camarasaurus) 자료가 ''카마라사우루스 수프레무스''(Camarasaurus supremus)로 분류되었다.[12] 이 자료는 많은 큰 척추와 일부 두개골 요소로 구성되어 있다.
1967년, 제임스 A. 젠슨은 콜로라도 서부 언컴파그레 힐에서 척추 대부분을 포함한 보존이 잘 된 부분적이고 연결된 두개골의 골격을 수집했다.[13][14] 이 골격(BYU 9740)은 브리검 영 대학교에 보관되었고,[15][13] 1988년에 카마라사우루스과의 새로운 속이자 종인 ''카테토사우루스(Cathetosaurus) lewisi''로 묘사되었다.[14] 1996년, 존 맥킨토시와 그의 동료들은 이 골격을 연구하여 ''카마라사우루스''(Camarasaurus)의 종으로 분류하며, ''C. supremus, C. grandis, C. lentus''와 ''C. lewisi''가 유효하다고 결론지었다.[13] 2013년, 옥타비오 마테우스와 이매뉴얼 츠초프는 ''C. lewisi''가 실제로는 자체 속이라고 주장했지만,[16] 츠초프의 추가 연구는 하우 채석장 표본이 ''카마라사우루스''(Camarasaurus)일 가능성이 높다고 결론지었다. 2019년 현재, 대부분의 연구자들은 ''C. lewisi''를 ''카마라사우루스''(Camarasaurus)의 종으로 간주한다.
1992년, 제프리 파커와 그의 동료들은 본 캐빈 채석장 근처의 코모 블러프에서 또 다른 ''카마라사우루스 그란디스''(Camarasaurus grandis) 골격(GMNH-PV 101)을 발견했다. 이 골격은 현재 군마 현립 자연사 박물관에 소장되어 있다. 같은 해, 하우 채석장에서 아탈 공룡 박물관 팀에 의해 또 다른 ''카마라사우루스''(Camarasaurus) 골격(SMA 002)이 발견되었다. 이 골격은 거의 모든 요소가 연결되어 있고 두개골과 뒷다리에 피부 인상이 남아 있으며,[17][22] ''C. lewisi''의 표본일 가능성이 제기되었다.[22] 1996년에는 사우스다코타 서부[18]와 뉴멕시코에서 ''카마라사우루스''(Camarasaurus)의 여러 단편적 유해가 묘사되어[19] 이 속의 분포 범위를 확장했다. 2005년, 몬태나 중부에서 ''카마라사우루스''(Camarasaurus)의 가장 북쪽 표본이 발견되었는데, 거의 완전한 두개골과 몇 개의 두개골 후 요소를 포함하고 있다.
3. 특징


''카마라사우루스'' 화석은 매우 흔하게 발견되며, 500개 이상의 표본이 알려져 있다. 여기에는 많은 고립된 뼈와 약 50개의 부분 골격이 포함된다.[3] 이들은 미국 서부의 광범위한 지역, 북쪽으로는 몬태나 주에서 남쪽으로는 뉴멕시코 주에 이르기까지 모리슨 층의 암석에서 발견된다.[3] 이러한 풍부함 때문에 ''카마라사우루스''는 매우 잘 알려진 용각류 중 하나이다. 특히 어린 개체 CM 11338은 지금까지 발견된 가장 완전한 용각류 골격이다.[4] 용각류에서 완전한 목뼈가 드물게 발견됨에도 불구하고, 다섯 개의 ''카마라사우루스'' 표본은 모든 또는 거의 모든 경추를 보존하고 있다.[5]
전반적으로 9~18미터, 추정 체중 20톤에 달하지만, 동시대의 용각류로서는 그다지 크지 않아 크게 번성한 이유로 생각된다.
각 발에는 5개의 발가락이 있었고, 안쪽 발가락에는 방어를 위한 크고 날카로운 갈고리 발톱이 있었다. 대부분의 용각류와 마찬가지로 앞다리는 뒷다리보다 짧았지만, 어깨의 높이는 거의 수평이며 약간 뒤로 기울어져 있었다. 몇몇 용각류에는 강력한 근육 부착부가 되는 비교적 긴 극돌기가 각 척추뼈에 있지만, 카마라사우루스에는 이것이 없어 뒷다리만으로 일어설 수 없었던 것으로 보인다. 척추뼈는 경량화 장치로서의 공포가 많이 있는 등 특화되어 있으며, 여기에서 "공동을 가진 도마뱀"이라는 학명이 붙여졌다. 현생 코끼리와 같이 몸이 무거운 생물에 특유한, 발꿈치 뿌리에 있는 쐐기 모양의 해면 조직을 가지고 있었다. 이 크기의 용각류로서는 목과 그에 상응하는 꼬리는 비교적 짧았다.
다른 용각류처럼 골반 근처에서 척추가 비대해졌는데, 한때 고생물학자들은 거대한 몸을 제어하기 위해 필요한 두 번째 뇌라고 믿었다. 현대의 견해에서는 뇌가 아니라 큰 신경의 활동 영역이었다고 여겨진다. 이 비대해진 신경은 두개골 내에 존재하는, 현저하게 작은 뇌보다 컸다.
''카마라사우루스''는 닭처럼 위에서 물리적인 음식 소화를 돕기 위한 위석을 삼켰으며, 그 돌이 매끄러워져 쓸모없어지면 토하거나 배설했던 것으로 보인다. 이 설을 뒷받침하듯, 모리슨 층에서는 종종 표면이 매우 매끄러운 돌 무더기가 고립되어 다수 발견되고 있다.
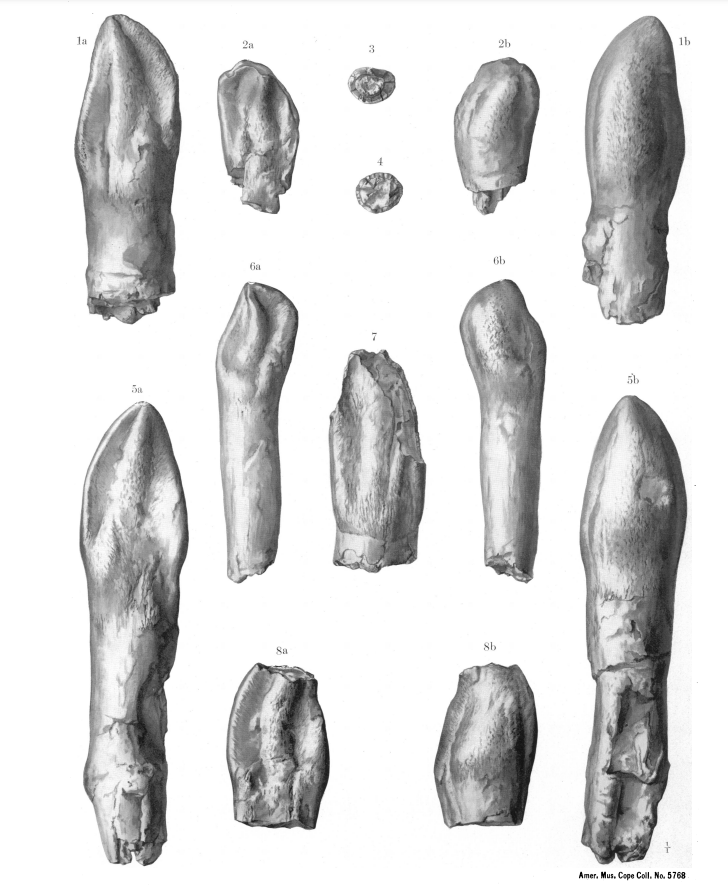
3. 1. 두개골 및 치아
''카마라사우루스''의 아치형 두개골은 눈에 띄게 사각형이었고, 무딘 주둥이에는 많은 창(fenestrae)이 있었다. ''카마라사우루스''의 튼튼한 두개골은 모리슨 층에서 발견되는 디플로도쿠스과의 가늘고 섬세한 두개골과 달리 다른 많은 사우로포드보다 훨씬 더 잘 보존된다.[21] 19cm 길이의 이빨은 끌 모양(주걱 모양)이었으며 턱을 따라 고르게 배열되어 있었다. 이빨의 강도는 ''카마라사우루스''가 가늘고 긴 이빨을 가진 디플로도쿠스과보다 더 거친 식물 재료를 먹었을 가능성이 있음을 시사한다.와이오밍의 호-스티븐스 채석장에서 발견된 SMA 0002(''Cathetosaurus''로도 분류됨)라는 ''카마라사우루스'' 표본은 연조직의 증거를 보여준다.[22] 턱선을 따라 동물의 잇몸이었던 것으로 보이는 골화된 잔해가 발견되었는데, 이는 깊숙이 자리 잡은 이빨이 잇몸으로 덮여 있고, 치아의 팁만 튀어나와 있음을 나타낸다. 죽은 후에는 잇몸이 뒤로 물러나고, 건조되고, 부패하면서 이빨이 소켓에서 더 밀려 나왔다. 표본 검사 결과 이빨이 질긴 바깥쪽 비늘과 다양한 종류의 부리로 덮여 있었을 가능성이 있지만, 이는 확실하지 않다.[22]
두개골은 매우 특징적이며 머리는 비교적 크고 현저한 사각형이며, 평평한 코끝에는 많은 구멍이 있었다. 꽤 튼튼하며, 종종 상태가 좋은 화석이 발견된다. 눈 앞에 배치된 거대한 콧구멍은 아마도 큰 침윤성 막을 가지고, 쥐라기의 고온 기후에서 뇌를 냉각시키는 역할을 했을 것으로 생각된다.
입에는 길이 19cm의 주걱 모양의 튼튼한 이빨이 턱을 따라 균등하게 배치되어 있었다. 이빨의 강도는 카마라사우루스가 가늘고 부서지기 쉬운 이빨을 가진 디플로도쿠스보다 거친 식물을 먹었음을 나타낸다.
3. 2. 골격
카마라사우루스는 발견된 용각류 공룡 중 가장 흔하고 보존 상태가 좋은 공룡 중 하나이며, 수많은 출판물에서 잘 묘사되어 있다.[22][3] 다른 마크로나리아와 마찬가지로, 전형적인 큰 콧구멍, 긴 앞다리, 그리고 동시대의 디플로도쿠스과에 비해 짧은 꼬리를 가지고 있었다.[3][21] 카마라사우루스는 동시대의 같은 지층에 속하는 종들에 비해 중간 크기의 용각류였지만, 티토니아 시기에는 ''C. supremus''가 큰 크기에 도달했다.[3] 가장 흔한 종인 ''C. lentus''의 최대 크기는 약 15m였다. 가장 큰 종인 ''C. supremus''는 최대 18m에서 23m의 길이와 최대 추정 체중 47톤에 도달했다.[20]카마라사우루스의 아치형 두개골은 현저하게 사각형이었고, 무딘 주둥이에는 많은 창(fenestrae)이 있었다. 카마라사우루스의 튼튼한 두개골은 모리슨 층에서 발견되는 디플로도쿠스과의 가늘고 섬세한 두개골과 달리 다른 많은 용각류보다 훨씬 더 잘 보존된다.[21] 19cm 길이의 이빨은 끌 모양(주걱 모양)이었으며 턱을 따라 고르게 배열되어 있었다. 이빨의 강도는 카마라사우루스가 가늘고 긴 이빨을 가진 디플로도쿠스과보다 더 거친 식물 재료를 먹었을 가능성이 있음을 시사한다.
카마라사우루스의 목은 용각류 기준으로는 중간 길이였으며 12개의 척추뼈로 구성되어 있었다. 대부분의 경추 신경 가시는 이분되었고, 동물이 성장함에 따라 더 많은 척추뼈가 이분된 신경 가시를 발달시켰다. 다른 용각류와 마찬가지로 목과 몸통의 척추뼈에는 생전에 호흡계통에 연결된 공기 주머니로 채워진 방이 있었다. 카마라사우루스가 "방이 있는 도마뱀"이라는 이름을 얻게 된 것은 바로 이 방들 때문이다.
카마라사우루스의 꼬리는 53개의 척추뼈로 구성되어 있었다.
3. 3. 연조직
와이오밍 주 호-스티븐스 채석장에서 발견된 ''카마라사우루스'' 표본 SMA 0002 (''Cathetosaurus''로도 분류됨)는 연조직의 증거를 보여준다.[22] 턱선을 따라 동물의 잇몸이었던 것으로 보이는 골화된 잔해가 발견되었는데, 이는 깊숙이 자리 잡은 이빨이 잇몸으로 덮여 있고, 치아의 끝부분만 튀어나와 있음을 나타낸다. 죽은 후에는 잇몸이 뒤로 물러나고, 건조되고, 부패하면서 이빨이 이빨 소켓에서 더 밀려 나왔다. 표본 검사 결과 이빨이 질긴 바깥쪽 비늘과 다양한 종류의 부리로 덮여 있었을 가능성이 있지만, 이는 확실하지 않다.[22]4. 생태


유타주 공룡 국립 기념물에서 회수된 ''카마라사우루스'' 골반에서 ''알로사우루스''에 의한 것으로 추정되는 홈파기가 발견되었으며, ''C. lewisi''의 모식표본 장골에서도 수각류의 큰 물린 자국이 발견되었다.[15]
1992년, 콜로라도주 캐논시티 근처 모리슨 층의 브라이언 스몰 스테고사우루스 채석장에서 부분적인 ''C. grandis'' 골격이 발견되었다. 이 표본은 DMNH 2908로 분류된 부분적인 오른쪽 상완골과 등 및 꼬리에서 유래한 연관된 척추뼈를 보존하고 있었다. 2001년, 로리 맥위니, 케네스 카펜터, 브루스 로스차일드는 상완골에서 관찰된 병리학에 대한 설명을 발표했다. 그들은 25 x 18 cm 넓이의 골피질 근접 병변을 관찰했는데, 이는 엮인 섬유와 유사한 뼈로 이루어져 있었다. 엮인 뼈는 부상이나 질병의 징후이다. 엮인 뼈의 "물결 모양의 섬유 다발"은 ''m. brachialis''의 방향으로 향하는 것으로 관찰되었다. 병변의 융합과 그 근처와 먼 끝 부분에서 다공성의 부족은 골막염이 비활성 상태이거나 치유되었음을 나타낸다.
맥위니와 다른 연구자들은 이 부상이 동물에게 지속적인 고통의 원인이 되었을 것이라고 주장했다. 그것은 근육에 압력을 가했을 것이며, 이 압력은 근육의 혈관과 신경을 압박하여 사지의 굴근과 신근의 운동 범위를 감소시켰을 것이다. 이러한 효과는 상완골의 병변 위치에 따라 ''M. brachialis'', ''상완요골근'', 그리고 덜한 정도의 ''상완이두근''을 방해했을 것이다. 연구자들은 근육과 골막의 염증이 앞다리 아래쪽 부위에도 추가적인 합병증을 유발했을 것이라고 추론했다. 병변은 또한 장기적인 근막염과 근염을 유발했을 것이다. 이러한 병리학적 과정의 누적 효과는 사지의 움직임 능력에 중등도에서 심각한 영향을 미쳤으며 "음식 섭취와 포식자로부터의 도피와 같은 일상 활동을 수행하기 어렵게 만들었다."
병리학의 원인을 결정하기 위해 맥위니와 다른 연구자들은 3mm 간격으로 CT 스캔을 수행했다. CT 스캔 결과, 덩어리는 일관된 방사선 밀도를 가지고 있었고 뼈의 피질과 방사선 투과성 선으로 분리되어 있었다. 골수염이나 감염성 골막염과 같은 스트레스 골절이나 감염 과정의 증거는 발견되지 않았다. 그들은 또한 뼈 돌기의 축이 상완골의 수직축에 대해 25°인 반면, 골연골종은 상완골의 축에 대해 90°로 형성되기 때문에 골연골종을 배제했다.
과학자들이 뼈 돌기 병변의 기원으로 식별한 다른 후보는 다음과 같다.
- 비대성 골관절병증 – 뼈 돌기 같은 과정의 존재로 인해 배제되었다.
- 골성 골종 – 뼈 돌기나 골아세포성 반응을 설명하지 못한다.
- 정강이 통증 또는 경골 스트레스 증후군 – 많은 증상이 공통적으로 나타날 수 있으므로 가능한 기원이나, 정강이 통증은 뼈 돌기를 설명하지 못한다.
- 외상성 근육 골화증 (국소적) – 가능성이 있지만, 그럴 가능성은 낮다.
- 견열 손상 – 맥위니와 다른 연구자들은 "근육의 반복적인 과도한 활동"에 의해 발생한 견열 손상을 상완골의 병변에 대한 가장 유력한 원인으로 간주했다. 연구자들은 병변이 ''m. brachialis''의 견열로 시작되어 "아래로 경사진 타원형 덩어리"를 형성했다고 믿었다. 뼈 돌기는 굴근 운동에 의해 ''m. brachioradialis'' 기저부에서 파열된 후 골아세포 반응에 의해 발생했다.
로키 산맥 동쪽 측면에 위치한 모리슨 층은 후기 쥐라기 지층이 화석으로 풍부하게 발견되는 곳이다. 이곳에서는 ''카마라사우루스''의 친척인 ''디플로도쿠스'', ''아파토사우루스'', ''브라키오사우루스''를 포함한 많은 공룡 종을 발견할 수 있지만, 카마라사우루스는 이 지층에서 가장 흔한 공룡이다.[43] ''카마라사우루스'' 화석은 거의 모든 주요 지역에서 발견되었으며, 뉴멕시코에서 몬태나, 유타에서 오클라호마에 이르는 지역에서 화석이 발견되면서 모리슨 공룡 중 가장 넓은 분포를 보인다. 방사성 연대 측정에 따르면, 모리슨 퇴적층은 기저부가 1억 5630만 년 전(Mya)에서 상부가 1억 4680만 년 전으로, 이는 후기 옥스포드 절, 킴메리지절, 그리고 전기 티톤절 동물상 단계에 해당한다.[44][45] 그 환경은 뚜렷한 우기와 건기가 있는 반건조 기후로 해석된다.
공룡과 생흔 화석은 특히 뉴멕시코에서 앨버타와 서스캐처원까지 뻗어 있으며, 로키 산맥의 전구체가 서쪽으로 솟아오르기 시작하면서 형성된 모리슨 분지에서 발견된다. 동쪽을 향한 배수 분지에서 침식된 물질은 개울과 강에 의해 운반되어 늪지 저지대, 호수, 강 채널 및 범람원에 퇴적되었다.[46] 이 지층은 포르투갈의 로우리냐 지층과 아르헨티나의 카냐돈 칼카레오 지층과 연대가 비슷하며, 이 두 지층에서 카마라사우루스과 화석이 발견되었다.[37][47] 1877년, 이 지역은 초기 고생물학자 오트니엘 찰스 마쉬와 에드워드 드링커 코프 사이의 화석 수집 경쟁인 뼈 전쟁의 중심지가 되었으며, ''카마라사우루스'' 자체는 이 갈등 기간 동안 후자의 고생물학자에 의해 발견되고 명명되었다.
4. 1. 식성
카마라사우루스는 초식 공룡으로, 튼튼하고 숟가락 모양의 이빨을 가지고 있었다. 이 이빨은 다른 용각류 공룡들의 이빨보다 더 강하고 튼튼해서 질긴 식물을 먹기에 적합했다.4. 2. 성장

4. 3. 대사
이전에는 과학자들이 카마라사우루스와 다른 용각류들이 위에서 음식을 갈아내는 데 도움을 주기 위해 위석을 삼켰다가 매끄러워지면 토하거나 배설했을 것이라고 제안했다. 그러나 최근 위석에 대한 증거를 분석한 결과, 이는 사실이 아닌 것으로 보인다. 카마라사우루스의 강하고 튼튼한 이빨은 대부분의 용각류보다 더 발달했으며, 평균 62일마다 교체되었다. 이는 카마라사우루스가 음식을 삼키기 전에 어느 정도 입 안에서 저작했을 수 있음을 시사한다. 다른 연구 결과에 따르면 카마라사우루스 종은 다른 용각류와는 다른 식물을 선호하여, 경쟁 없이 동일한 환경을 공유할 수 있었다.[39]4. 4. 무리 생활
골 조직학을 통해 연구자들은 특정 개체가 도달한 나이를 추정할 수 있다. Griebeler 등(2013)의 연구에서는 장골 조직학적 데이터를 조사한 결과, ''카마라사우루스''(Camarasaurus) sp. '''CM 36664'''는 14247kg의 무게였으며, 20세에 성적 성숙에 도달하여 26세에 사망했다.[40]성체와 어린 개체가 함께 죽어있는 화석이 발견되어 무리 지어 이동했을 가능성이 시사된다. 건조한 곳을 선호했으며, 와이오밍주부터 뉴멕시코주에 걸쳐 널리 퍼진 미국 서부의 초원에 살았다.[53]
다른 공룡과 같은 둥지에 줄 지어 있는 형태가 아닌, 열을 지어 놓인 알의 발견 기록이 있다. 카마라사우루스는 자식 양육을 하지 않았다고 생각된다.
일부 고생물학자들은 카마라사우루스의 수명이 100년까지 살았을 가능성을 지적하기도 한다.
5. 고병리학
Camarasaurus영어는 포식자에게 취약한 기간을 줄이기 위해 빠르게 성장했으며, 이는 어린 개체일 때 상대적으로 높은 신진대사율을 가졌음을 시사한다.[42] 이글 외 다수는 클럼프 동위원소 열역학 기술을 이용하여, Camarasaurus영어를 포함한 다양한 쥐라기 용각류의 치아를 덮고 있는 에나멜을 분석했는데, 32.4°C~36.9°C도의 온도가 측정되었으며 이는 현대 포유류의 온도와 유사한 수준이다.[41]
6. 분류
카마라사우루스과(Camarasauridae)의 기준 속인 ''카마라사우루스''(Camarasaurus)는 후기 쥐라기에 살았던 중형 마크로나리아(Macronaria) 용각류이다. 카마라사우루스과는 뒷다리보다 앞다리가 짧고, 큰 어깨뼈와 가슴뼈를 가지고 있었으며, 목보다 긴 꼬리를 가지고 있었다.[3]
1877년 에드워드 코프(Edward Cope)는 ''카마라사우루스''를 세티오사우루스(Cetiosaurus), 보트리오스폰딜루스(Bothriospondylus), 오르니토프시스(Ornithopsis), 안키사우루스(Anchisaurus) (''메가닥틸루스''(Megadactylus)) 및 ''뉴마토스테우스''(Pneumatosteus)와 밀접한 관련이 있는 공룡이라고 보았지만,[1] ''암피코엘리아스''(Amphicoelias)가 묘사될 때까지 이러한 분류군에 대한 그룹을 명명하지 않았고, 카마라사우루스과를 세웠다.[5]
''카마라사우루스''는 논쟁의 여지없이 카마라사우루스과의 유효한 속으로 여겨지는 유일한 분류군이며, 다음 네 개의 종을 포함한다.
- ''C. 그란디스(C. grandis)''
- ''C. 렌투스(C. lentus)''
- ''C. 루이시(C. lewisi)''
- ''C. 수프레무스(C. supremus)''
''C. 루이시''는 별개의 속인 ''카테토사우루스''(Cathetosaurus)를 나타낼 수 있다. 종의 모식종이 이전에 ''카마라사우루스''에 할당되었던 ''로우리냐사우루스(Lourinhasaurus)''는 대부분의 연구에서 카마라사우루스과로 간주되지만, 기저 유사우로포드로도 간주되었다.[23][24]
대부분의 연구자들은 카마라사우루스에 '''C. grandis''', '''C. lentus''', '''C. lewisi''', '''C. supremus'''의 4종이 유효하다고 본다. 1877년 코프가 명명한 '''C. supremus'''는 모식종이다. '''C. grandis'''는 1877년에, '''C. lentus'''는 1889년에 명명되었다. '''C. lewisi'''는 원래 1988년 별개의 속인 '''Cathetosaurus'''로 기재되었지만, 1996년 '''Camarasaurus'''의 한 종으로 재분류되었다. 일부 연구자들은 '''Cathetosaurus'''를 별개의 속으로 복원해야 한다고 주장하는 반면,[16] 다른 연구자들은 '''C. lewisi'''가 다른 '''Camarasaurus''' 종과 동의어일 수 있다고 제안했다.

'''C. supremus'''는 이름에서 알 수 있듯이 알려진 '''Camarasaurus''' 종 중 가장 크며,[3] 후기 쥐라기 모리슨 층에서 발견된 가장 거대한 용각류 중 하나이다. '''C. supremus'''는 속 전체의 전형적인 모습과는 다르며, 층의 최상층부에서만 알려져 있으며 극히 드물다. '''C. grandis,'', '''C. lentus,''', 그리고 '''C. lewisi'''는 더 작았으며, 모리슨 층의 초기 단계에서도 발견되었다.

지층 증거는 세 종 사이의 물리적 차이와 일치하는 연대기적 순서를 제시하며, 모리슨 층 내의 진화적 진행을 설명한다. '''C. grandis'''는 가장 오래된 종이며, 모리슨 층의 가장 낮은 암석층에서 발견되었다. '''C. lewisi'''는 상부 모리슨 층의 가장 낮은 지층에서 '''C. grandis'''와 잠시 공존하다가 멸종되었지만, 이는 '''C. lewisi'''의 표본 부족 때문일 가능성이 있다. '''C. lentus'''는 그 후에 나타나서 수백만 년 동안 '''C. grandis'''와 공존했으며, 이는 두 종의 척추 해부학적 차이에서 시사되듯이 서로 다른 생태적 지위 때문일 수 있다. 나중 단계에서 '''C. grandis'''는 암석 기록에서 사라지고, '''C. lentus'''만 남았다. 그 후, '''C. lentus'''도 사라졌고, 동시에 '''C. supremus'''가 최상층에서 나타났다. 이 종들의 직접적인 계승과 두 종의 매우 유사하다는 점은 '''C. supremus'''가 '''C. lentus'''로부터 직접 진화하여 더 크고 더 오래 생존한 개체군을 나타낼 수 있음을 시사한다.[25]
코프(Cope)가 발견한 최초의 종은 ''C. supremus''(종소명의 의미는 "가장 큰")이다. 이 외에도 같은 1877년에 ''C. grandis'', 1889년에 ''C. lentus'', 그리고 1988년에 ''C. lewisi''(원래는 카테토사우루스''Cathetosaurus'')가 발견되었다.
6. 1. 계통도
카마라사우루스는 브라키오사우루스와 같은 더 파생된 형태보다는 모든 마크로나리아의 공통 조상과 더 밀접한 관련이 있는 기저(base) 마크로나리아이다.| 계통 |
|---|
카마라사우루스과에는 카마라사우루스(Camarasaurus)와 스페인의 아라고사우루스(''Aragosaurus'')가 포함된다. 과거에는 에우헬로푸스(Euhelopus) 등 사각형 두개골과 짧은 목을 가진 몇몇 용각류가 여기에 분류되었으나, 오늘날에는 다른 곳으로 분류된다.
탄 외(Tan et al) (2020) 이후의 기저 마크로나리아의 단순화된 분기도는 다음과 같다.
진용각류
:투리아사우리아
:신용각류
::디플로도쿠스상과
::마크로나리아
:::'''카마라사우루스'''
:::''벨루사우루스''
::::''유로파사우루스''
::::''테후엘체사우루스''
::::''갈베사우루스''
:::::''유헬로푸스''
:::::티타노사우루스형류
7. 분포
로키 산맥 동쪽 측면에 위치한 모리슨 층은 후기 쥐라기 지층이 화석으로 풍부하게 발견되는 곳이다. 이곳에서는 ''카마라사우루스''의 친척인 ''디플로도쿠스'', ''아파토사우루스'', ''브라키오사우루스''를 포함한 많은 공룡 종을 발견할 수 있지만, 카마라사우루스는 이 지층에서 가장 흔한 공룡이다.[43] ''카마라사우루스'' 화석은 거의 모든 주요 지역에서 발견되었으며, 뉴멕시코 주에서 몬태나 주, 유타 주에서 오클라호마 주에 이르는 지역에서 화석이 발견되면서 모리슨 공룡 중 가장 넓은 분포를 보인다.
방사성 연대 측정에 따르면, 모리슨 퇴적층은 기저부가 1억 5630만 년 전(Mya)에서 상부가 1억 4680만 년 전으로, 이는 후기 옥스포드 절, 킴메리지절, 그리고 전기 티톤절 동물상 단계에 해당한다.[44][45] 그 환경은 뚜렷한 우기와 건기가 있는 반건조 기후로 해석된다.
8. 공존했던 생물
모리슨 층은 로키 산맥 동쪽 측면에 위치한 후기 쥐라기 지층으로 화석이 풍부하게 발견되는 곳이다. 이곳에서는 ''카마라사우루스''의 친척인 ''디플로도쿠스'', ''아파토사우루스'', ''브라키오사우루스''를 포함한 많은 공룡 종을 발견할 수 있지만, 카마라사우루스는 이 지층에서 가장 흔한 공룡이다.[43] ''카마라사우루스'' 화석은 뉴멕시코에서 몬태나, 유타에서 오클라호마에 이르는 지역에 걸쳐 발견되어 모리슨 공룡 중 가장 넓은 분포를 보인다.
방사성 연대 측정에 따르면, 모리슨 퇴적층은 1억 5630만 년 전(Mya)에서 1억 4680만 년 전 사이로, 후기 옥스포드 절, 킴메리지절, 전기 티톤절 동물상 단계에 해당한다.[44][45] 이 환경은 뚜렷한 우기와 건기가 있는 반건조 기후로 해석된다.
모리슨 분지는 뉴멕시코에서 앨버타와 서스캐처원까지 뻗어 있으며, 로키 산맥의 전구체가 서쪽으로 솟아오르기 시작하면서 형성되었다. 동쪽을 향한 배수 분지에서 침식된 물질은 개울과 강에 의해 운반되어 늪지 저지대, 호수, 강 채널 및 범람원에 퇴적되었다.[46] 이 지층은 포르투갈의 로우리냐 지층과 아르헨티나의 카냐돈 칼카레오 지층과 연대가 비슷하며, 이 두 지층에서도 카마라사우루스과 화석이 발견되었다.[37][47]
모리슨 층은 ''마라푸니스aurus'', ''암피코엘리아스'', 바라사우루스, ''디플로도쿠스'', ''아파토사우루스'', ''브론토사우루스'', ''브라키오사우루스''와 같은 거대한 용각류 공룡이 지배했던 환경을 기록하고 있다. ''카마라사우루스''와 함께 살았던 공룡으로는 초식성 조반류인 ''캄프토사우루스'', ''가고일로사우루스'', ''드리오사우루스'', ''스테고사우루스'', ''나노사우루스''가 있었다.
이 고생물 환경의 포식자로는 수각류인 ''사우로파가낙스'', ''토르보사우루스'', ''케라토사우루'', ''마르소사우루스'', ''스토케소사우루스'', ''오르니톨레스테스'',[48] 그리고 알로사우루스가 있었는데, 알로사우루스는 수각류 표본의 최대 75%를 차지하며 모리슨 먹이 그물의 최상위 영양 단계에 있었다.[49][50] ''카마라사우루스''는 ''알로사우루스'', ''아파토사우루스'', ''스테고사우루스'', ''디플로도쿠스''와 같은 장소에서 발견된다.[51]
이 지역의 다른 생물체로는 이매패류, 달팽이, 조기어, 개구리, 도롱뇽, 거북이, 이빨도마뱀, 도마뱀, 육상 및 수생 악어류, 그리고 ''하르팍토그나투스''와 ''메사닥틸루스''와 같은 몇몇 종의 익룡이 있었다. 당시 존재했던 초기 포유류는 도코돈트 (예: ''도코돈''), 다구치류, 대칭치류, 삼관치류였다. 이 시기의 식물상은 녹조류, 균류, 이끼, 속새, 소철, 은행나무, 그리고 몇몇 과의 구과식물 화석으로 알 수 있다. 식생은 고사리와 양치류의 강변 숲 (갤러리 숲)에서부터 ''아라우카리아''와 유사한 구과식물 ''브라키필룸''과 같은 나무가 드문드문 있는 양치류 사바나까지 다양했다.[52]
9. 대중 문화
일본에서 출시된 공룡을 모티브로 한 로봇 플레오(PLEO)는 카마라사우루스의 생후 일주일 정도 된 새끼라는 설정이다.
참조
[1]
논문
On a gigantic saurian from the Dakota Epoch of Colorado
Paleontological Bulletin
1877
[2]
논문
On reptilian remains from the Dakota beds of Colorado
Paleontological Bulletin
1877
[3]
웹사이트
Camarasaurus, Amphicoelias, and other sauropods of Cope: Memoirs of the American Museum of Natural History, v. 3, p. 247-387.
https://digitallibra[...]
[4]
뉴스
Notice of New Dinosaurian Reptiles from the Jurassic Formation
https://www.ajsonlin[...]
American Journal of Science
1877
[5]
논문
On Amphicoelias, a new genus of saurian from the Dakota Epoch of Colorado
Paleontological Bulletin
1877
[6]
서적
Fore and hind limbs of Sauropoda from the Bone Cabin Quarry
https://books.google[...]
Vol. 3
1901
[7]
간행물
The Fore Leg And Pectoral Girdle Of Morosaurus
1901
[8]
웹사이트
AMNH 467 - Camarasaurus sp. entry
http://research.amnh[...]
[9]
뉴스
Bully for ''Camarasaurus''
https://dinosours.wo[...]
Dinosours
2014-10-30
[10]
웹사이트
Diplodocus and Camarasaurus (NMNH Series)
https://extinctmonst[...]
2016-12-29
[11]
간행물
Description of the Type of Uintasaurus douglassi HOLLAND
Annals of the Carnegie Museum
1924
[12]
웹사이트
OMNH 1811, a Camarasaurus dorsal from Black Mesa
https://svpow.com/20[...]
2018
[13]
간행물
The osteology of Camarasaurus lewisi (Jensen, 1988)
https://pubs.er.usgs[...]
Brigham Young University Geology Studies
1996
[14]
간행물
A fourth new sauropod dinosaur from the Upper Jurassic of the Colorado Plateau and sauropod bipedalism
1988
[15]
웹사이트
Camarasaurus Paleofile
http://www.paleofile[...]
[16]
간행물
Cathetosaurus as a valid sauropod genus and comparisons with Camarasaurus.
https://docentes.fct[...]
2013
[17]
문서
Articulated bone sets of manus and pedes of Camarasaurus (Sauropoda, Dinosauria)
2015
[18]
간행물
Sauropod dinosaurs of the Morrison Formation (Upper Jurassic), Black Hills, South Dakota and Wyoming.
https://pubs.geoscie[...]
1996
[19]
간행물
Jurassic dinosaurs in New Mexico.
https://books.google[...]
1993
[20]
서적
Prehistoric Life
Dorling Kindersley
[21]
논문
Description of the palate and lower jaw of the sauropod dinosaur Diplodocus (Reptilia: Saurischia) with remarks on the nature of the skull of Apatosaurus
1975
[22]
간행물
The dentition of a well-preserved specimen of Camarasaurus sp.: implications for function, tooth replacement, soft part reconstruction, and food intake.
https://link.springe[...]
2017
[23]
간행물
Phylogenetic reassessment of ''Lourinhasaurus alenquerensis'', a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal
https://doi.org/10.1[...]
2014-02-19
[24]
서적
Sauropoda
https://archive.org/[...]
University of California Press
2004
[25]
문서
Camarasaurus grandis
2007
[26]
간행물
On Amphicoelias, a genus of Saurians from the Dakota epoch of Colorado
1877
[27]
간행물
A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)
2015-04-07
[28]
논문
Notice of new American Dinosauria
1889
[29]
간행물
The Late Jurassic sauropod dinosaur 'Morosaurus' agilis Marsh, 1889 reexamined and reinterpreted as a dicraeosaurid
https://doi.org/10.1[...]
2020-12-10
[30]
간행물
Appendix; Comparison of the principal forms of the Dinosauria of Europe and America
http://marsh.dinodb.[...]
1889
[31]
서적
Die fossil Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte
Monographien zur Geologie und Palaeontologie (Ser. 1)
1932
[32]
간행물
The Anatomy and Phylogenetic Relationships of "Pelorosaurus" becklesii (Neosauropoda, Macronaria) from the Early Cretaceous of England
https://doi.org/10.1[...]
2015-06-03
[33]
서적
Vertébrés fossiles du Portugal: contributions à l'étude des poissons et des reptiles du jurassique et du crétacique
l'Académie royale des Sciences
1898
[34]
간행물
A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia
1991-10-24
[35]
논문
Skull and atlas-axis complex of the Upper Jurassic sauropod Camarasaurus Cope (Reptilia: Saurischia)
1995
[36]
논문
Upper Jurassic sauropod record in the Lusitanian Basin (Portugal): Geographical and lithostratigraphical distribution
[37]
논문
Découverte d'une riche faune de Reptiles Dinosauriens dans le Jurassique supérieur du Portugal
[38]
서적
Sauropoda
University of California Press
[39]
웹사이트
High tooth replacement rates in largest dinosaurs contributed to their evolutionary success
https://www.scienced[...]
[40]
논문
Aging, Maturation and Growth of Sauropodomorph Dinosaurs as Deduced from Growth Curves Using Long Bone Histological Data: An Assessment of Methodological Constraints and Solutions
[41]
논문
Dinosaur Body Temperatures Determined from Isotopic (13C-18O) Ordering in Fossil Biominerals
2011-07-22
[42]
논문
Growth record and histological variation in the dorsal ribs ofCamarasaurussp. (Sauropoda)
http://dx.doi.org/10[...]
2014-06-07
[43]
문서
"Camarasaurus supremus"
[44]
논문
Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations
[45]
서적
The Morrison Formation: An Interdisciplinary Study
Taylor and Francis Group
[46]
서적
An Odyssey in Time: Dinosaurs of North America
NorthWord Press
[47]
논문
Osteology and phylogenetic relationships of Tehuelchesaurus benitezii (Dinosauria, Sauropoda) from the Upper Jurassic of Patagonia
2011-08-24
[48]
서적
Jurassic West: The Dinosaurs of the Morrison Formation and Their World
Indiana University Press
[49]
서적
Jurassic West: The Dinosaurs of the Morrison Formation and Their World
Indiana University Press
[50]
간행물
Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.
New Mexico Museum of Natural History and Science
[51]
논문
Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation
[52]
간행물
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[53]
웹사이트
JWE2 ツールボックス データべース カマラサウルス
https://www.paleo.gg[...]
2024-10-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com