진용각류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
진용각류는 목이 길고 사족보행을 하며 초식성인 공룡 분류군이다. 큰 몸집에 맞춰 골격이 특화되었으며, "대량 섭식 모드"로의 진화를 거쳤다. 기저 진용각류인 파타고사우루스의 두개골은 약 60cm이며, 슈노사우루스는 긴 목을 특징으로 한다. 이들은 외부 콧구멍의 위치, 주둥이 형태, 두정골과 사각관골 사이의 접촉 부재 등의 특징을 갖는다. 발자국 증거를 통해 반-디지티그레이드 발 자세를 가졌을 것으로 추정되며, 중족골의 비대칭성을 보인다. 진용각류는 모든 주요 대륙에서 발견되며, 특히 남아메리카, 아프리카, 유럽, 아시아에서 다양한 종이 발견된다. 1995년 폴 업처치에 의해 명명되었으며, 불카노돈보다 티타노사우루스류에 더 가깝다. 진용각류의 진화에 대한 연구는 주로 북반구의 지층에 의존하지만, 탄자니아의 지층에서 더 다양한 고생물학적 특성이 발견된다.
더 읽어볼만한 페이지
| 진용각류 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Eusauropoda |
| 명명자 | 폴 업처치(Paul Upchurch), 1995 |
| 한국어 이름 | 진용각류 (신룡각류) |
| 로마자 표기 | Jin-yonggangnyu |
| 멸종 여부 | 멸종 |
| 화석 발견 시기 | 쥐라기 전기 - 백악기 후기 |
| 화석 연대 | 1억 8450만 년 전 - 6600만 년 전 |
| 분류 | |
| 상위 분류 | 용각하목 |
| 하위 분류 | 케티오사우루스과(Cetiosauridae) 마멘치사우루스과(Mamenchisauridae) 투리아사우루스류(Turiasauria) 신룡각류(Neosauropoda) |
| 불확실한 관계 및 기저 속 | |
| 속 | 알고아사우루스(Algoasaurus)? 아시아토사우루스(Asiatosaurus)? 아틀라사우루스(Atlasaurus)? 바구알리아(Bagualia) 바라파사우루스(Barapasaurus)? 벨루사우루스(Bellusaurus) 케티오사우리스쿠스(Cetiosauriscus) 체브사우루스(Chebsaurus) 콘드로스테오사우루스(Chondrosteosaurus)? 페르가나사우루스(Ferganasaurus) 라파렌토사우루스(Lapparentosaurus) 조바리아(Jobaria) 클라멜리사우루스(Klamelisaurus)? 코타사우루스(Kotasaurus)? 리우방고사우루스(Liubangosaurus) 네불라사우루스(Nebulasaurus) 옴덴사우루스(Ohmdenosaurus)? 킨링고사우루스(Qinlingosaurus)? 로에토사우루스(Rhoetosaurus)? 슈노사우루스(Shunosaurus) 스피노포로사우루스(Spinophorosaurus) 폴크하이메리아(Volkheimeria) |
2. 특징
진용각류는 목이 길고, 엄격하게 초식성이며, 반드시 사족보행을 한다.[3] 그들은 큰 몸집 때문에 고도로 특화된 골격 적응을 보이며, 중력보행을 한다.[4]
Yates와 Upchurch는 진용각류의 진화를 "대량 섭식 모드"로 이동하는 것으로 묘사했다. 그들은 치아를 지닌 뼈의 치조 가장자리에 측면 판의 발달을 설명했다. 이 판은 잎을 벗기는 데 사용될 수 있으며, 진용각류의 "U자형" 턱은 넓은 물기를 만들고, "살이 많은 뺨"의 손실은 입의 벌어짐을 증가시켰다.[4] 진용각류 치아의 치관은 또한 "주름진 에나멜 질감"을 가지고 있지만, 이것이 그들의 섭식 습관에 어떤 의미를 갖는지는 불분명하다.[5]
== 머리와 목 ==
기저 진용각류인 ''파타고사우루스''(Patagosaurus)의 두개골 길이는 약 60cm이다.[6] 가장 기저적인 진용각류 중 하나인 ''슈노사우루스''(Shunosaurus)는 진용각류의 특징인 긴 목을 가지고 있는데, 이는 척추의 첫 번째 등뼈에 해당하는 부분이 목 부위로 통합되었고, 목 척추 중앙에 두 개의 목 척추가 추가된 것이다.[7]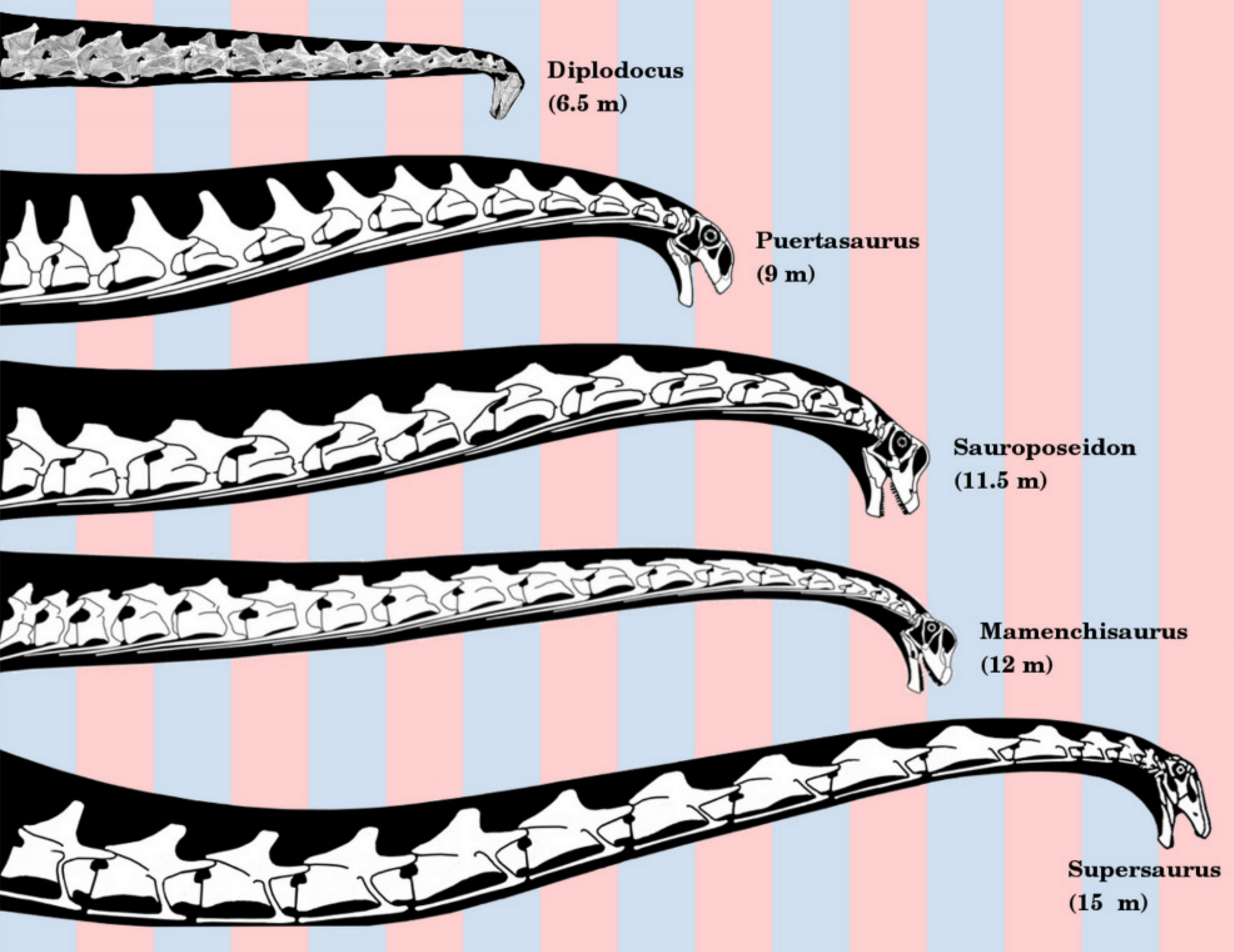
진용각류의 다른 공유파생형질로는 외부 콧구멍의 후퇴된 위치가 있다. 원용각류와 수각류와 달리, 진용각류는 매끄럽고 돌출되지 않은 치조와 비강 하부 부위를 가진 주둥이를 가지고 있지 않으며, "계단식 앞쪽 가장자리"를 가진 주둥이를 가지고 있다. 진용각류의 특징을 더 구분하면, 두정골과 사각관골 사이의 접촉 부재, 전두엽의 전방 돌기 부재, 사각관골의 원위부로 길어진 전방 가지 등이 있다. 또한, 진용각류는 비골의 전복부 돌기를 전상악골의 후외측 돌기에서 분리하고, 외부 콧구멍의 후복부 가장자리를 형성하는 긴 상악골을 가지고 있다.[5]
== 다리와 발 ==
유사우로포다는 발자국 증거를 통해 "반-디지티그레이드" 발 자세를 가졌을 것으로 추정된다.[7] 고생물학자 제프리 윌슨은 유사우로포드가 뒤꿈치와 중족골이 땅에서 떨어진 디지티그레이드 발을 가진 수각류와 용각류와 다르다고 설명한다. 유사우로포드는 중족골 I이 다른 중족골보다 넓어 중족골의 샤프트 직경에서 비대칭성을 보여주며, 이는 그들의 체중이 주로 발 안쪽에 실렸음을 시사한다.[5] 스티븐 샐즈버리와 제이 네이어에 따르면, 기초 유사우로포드는 4개의 발톱을 유지하지만, 네 번째 발가락의 마디 수를 세 단위로 줄인다.[8] 유사우로포드의 중족골은 경골 길이의 1/4 미만인데, 이는 긴 뒷다리와 경골 길이의 거의 절반에 달하는 중족골을 가진 용각류 외군과는 다르다.[7]
2. 1. 머리와 목
기저 진용각류인 ''파타고사우루스''(Patagosaurus)의 두개골 길이는 약 60cm이다.[6] 가장 기저적인 진용각류 중 하나인 ''슈노사우루스''(Shunosaurus)는 진용각류의 특징인 긴 목을 가지고 있는데, 이는 척추의 첫 번째 등뼈에 해당하는 부분이 목 부위로 통합되었고, 목 척추 중앙에 두 개의 목 척추가 추가된 것이다.[7]진용각류의 다른 공유파생형질로는 외부 콧구멍의 후퇴된 위치가 있다. 원용각류와 수각류와 달리, 진용각류는 매끄럽고 돌출되지 않은 치조와 비강 하부 부위를 가진 주둥이를 가지고 있지 않으며, "계단식 앞쪽 가장자리"를 가진 주둥이를 가지고 있다. 진용각류의 특징을 더 구분하면, 두정골과 사각관골 사이의 접촉 부재, 전두엽의 전방 돌기 부재, 사각관골의 원위부로 길어진 전방 가지 등이 있다. 또한, 진용각류는 비골의 전복부 돌기를 전상악골의 후외측 돌기에서 분리하고, 외부 콧구멍의 후복부 가장자리를 형성하는 긴 상악골을 가지고 있다.[5]
2. 2. 다리와 발
유사우로포다는 발자국 증거를 통해 "반-디지티그레이드" 발 자세를 가졌을 것으로 추정된다.[7] 고생물학자 제프리 윌슨은 유사우로포드가 뒤꿈치와 중족골이 땅에서 떨어진 디지티그레이드 발을 가진 수각류와 용각류와 다르다고 설명한다. 유사우로포드는 중족골 I이 다른 중족골보다 넓어 중족골의 샤프트 직경에서 비대칭성을 보여주며, 이는 그들의 체중이 주로 발 안쪽에 실렸음을 시사한다.[5] 스티븐 샐즈버리와 제이 네이어에 따르면, 기초 유사우로포드는 4개의 발톱을 유지하지만, 네 번째 발가락의 마디 수를 세 단위로 줄인다.[8] 유사우로포드의 중족골은 경골 길이의 1/4 미만인데, 이는 긴 뒷다리와 경골 길이의 거의 절반에 달하는 중족골을 가진 용각류 외군과는 다르다.[7]3. 분포
유사우로포다는 모든 주요 대륙에서 발견되며, 디플로코이드는 북반구에 널리 분포하고, 티타노사우루스는 남반구에서 발견된다. 그러나 이 두 그룹에 속하지 않는 기저 유사우로포다도 꽤 잘 알려져있다.[9]
''볼케하이메리아''와 ''아미그달로돈''과 같은 초기 유사우로포다, 그리고 ''파타고사우루스''와 같은 더 파생된 유사우로포다가 남아메리카에서 발견되었다.[6] ''볼케하이메리아''는 기저 유사우로포다로 분류되지만, 2004년 폴 업처치는 "뒤쪽이 오목한 경추 중심체, 대퇴골 전방 전자 부재, 그리고 경골의 측면으로 튀어나온 경골능선" 때문에 그 분류에 의문을 품었으며, 대신 일반 용각류일 수 있다고 생각했다.[9]
아프리카 유사우로포다에는 니제르에서 발견된 ''스피노포로사우루스''가 포함될 수 있지만, 그 분류군은 대신 ''불카노돈''에 더 가깝고 유사우로포다 밖에 있을 수 있다.[10][8] 또한, ''아틀라사우루스''는 모로코에서 발견되었고, ''요바리아''는 니제르에서 발견되었다. 그러나 두 속 모두 마크로나리아로 가능성이 있는 것으로 밝혀졌지만, 2012년 씽의 계통 발생 분석에 따르면 ''아틀라사우루스''는 투리아사우루스류로, ''요바리아''는 유사우로포다로 밝혀졌다.[10][9]
유럽에서는, 투리아사우리아가 프랑스, 스페인, 그리고 아마도 잉글랜드에서 발견되었으며, 스페인에서는 동일한 지역에서 여러 속이 발견되었다.[9][11] ''세티오사우루스'' 골격도 잉글랜드에서 발견되었으며, 이빨만 알려진, 유사우로포다일 가능성이 있는 속인 ''카르디오돈''과 ''오플로사우루스''도 발견되었다.[9]
마멘키사우루스과는 아시아 전역에서 널리 발견된다. 대부분의 속은 중국에서 발견되지만, ''마멘키사우루스''의 가능성 있는 표본이 태국에서 발견되었다.[10][9] 또한 중국에서는, 기저 유사우로포드인 ''네불라사우루스''가 ''스피노포로사우루스''의 자매 분류군으로, 마멘키사우루스과보다 더 파생되었지만 ''파타고사우루스''보다 덜 파생된 것으로 밝혀졌으며, ''슈노사우루스'' 속은 가장 기저적인 유사우로포드 중 하나일 가능성이 있다.[10][9] ''바라파사우루스'' 속은 인도에서 발견되었으며, 세티오사우루스류, 기저 유사우로포드 또는 유사우로포다 밖의 속을 나타낼 수 있다.[10][8][9]
4. 분류
1995년에 폴 업처치(Paul Upchurch)는 불카노돈과 같은 초기의 용각류보다 후기의 티타노사우루스류에 계통상 더 가까운 속들을 진용각류로 분류했다.[17]
다음은 용각류 내의 가설적 관계를 보여주는 분기도이다.[12] 기저 용각류에는 투리아사우리아(''투리아사우루스'', ''즈비'' 등)가 포함된다.[13]
살타사우루스와 같은 불카노돈과 같은 원시적인 용각류에 비해 더 티타노사우루스에 근연 관계에 있는 모든 용각류로 구성된 용각류 공룡의 분기군이다. 이는 케임브리지 대학교의 폴 업처치에 의해 명명되었다.
4. 1. 하위 분류
1995년에 폴 업처치(Paul Upchurch)는 불카노돈과 같은 초기의 용각류보다 후기의 티타노사우루스류에 계통상 더 가까운 속들을 진용각류로 분류했다.[17]다음은 용각류 내의 가설적 관계를 보여주는 분기도이다.[12] 기저 용각류에는 투리아사우리아(''투리아사우루스'', ''즈비'' 등)가 포함된다.[13]
살타사우루스와 같은 불카노돈과 같은 원시적인 용각류에 비해 더 티타노사우루스에 근연 관계에 있는 모든 용각류로 구성된 용각류 공룡의 분기군이다. 이는 케임브리지 대학교의 폴 업처치에 의해 명명되었다.
5. 고생물학
노바스가 지적하듯이, 진용각류의 진화에 대한 자료는 대부분 북반구의 몇몇 지층에 근거하고 있다. 하지만 탄자니아, 특히 카나돈 아스팔토와 카나돈 칼세레오 지층과 같은 지역의 다른 지층들은 후기 쥐라기 시대에 진용각류의 더욱 다양하고 광범위한 고생물학적 특성을 드러낸다.[6]
참조
[1]
논문
The evolutionary history of sauropod dinosaurs
http://doc.rero.ch/r[...]
[2]
논문
A basal sauropod dinosaur from the Early Jurassic of Morocco
http://cr-palevol.fr[...]
2004-05
[3]
논문
Why sauropods had long necks; and why giraffes have short necks
[4]
논문
A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism
[5]
서적
The Sauropods: evolution and paleobiology
University of California Press
[6]
서적
The age of dinosaurs in South America
Indiana University Press
[7]
논문
Early Evolution and Higher-Level Phylogeny of Sauropod Dinosaurs
http://doc.rero.ch/r[...]
[8]
논문
New anatomical information on ''Rhoetosaurus'' Longman, 1926, a gravisaurian sauropodomorph dinosaur from the Middle Jurassic of Queensland, Australia
[9]
서적
The Dinosauria
https://archive.org/[...]
University of California Press
[10]
논문
A new basal eusauropod from the Middle Jurassic of Yunnan, China, and faunal compositions and transitions of Asian sauropodomorph dinosaurs
2013
[11]
논문
High European sauropod dinosaur diversity during Jurassic-Cretaceous transition in Riodeva (Teruel, Spain)
2009-09
[12]
논문
Biology of the sauropod dinosaurs: the evolution of gigantism
[13]
논문
a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal
2014-05-06
[14]
문서
"Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages,"
http://www.geol.umd.[...]
[15]
문서
"The evolutionary history of sauropod dinosaurs."
[16]
문서
"Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages,"
http://www.geol.umd.[...]
[17]
문서
"The evolutionary history of sauropod dinosaurs."
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com