성구동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
성구동물은 바다에 사는 무척추동물로, 몸통과 내반으로 구성되며, 전 세계적으로 약 162종이 존재한다. 수정란은 발생 과정을 거쳐 트로코포라 유생이 되며, 일부 종은 무성 생식과 유성 생식을 모두 한다. 해부학적으로는 분절되지 않은 몸과 정수압골격, 촉수 등을 가지며, 소화계, 순환계, 신경계가 발달해 있다. 이들은 캄브리아기부터 존재했으며, 화석 기록이 드물다. 일부 종은 식용 또는 낚시 미끼로 사용된다.
더 읽어볼만한 페이지
- 성구동물 - 등촉수별벌레강
등촉수별벌레강은 바다 밑바닥에 서식하며 유기물을 섭취하는 벌레의 일종으로, 몸 표면의 방패 모양 유무에 따라 아스피도시폰목과 등촉수별벌레목으로 나뉜다. - 성구동물 - 성구강
성구강은 미끈이별벌레목과 별벌레목으로 구분되며, 2개의 목과 4개의 과로 분류되고, 체벽 근육의 차이로 식별하며, 2012년 분자계통학 연구에 기반하여 과 수준의 분류가 재편되었다. - 콘스턴틴 새뮤얼 라피네스크가 명명한 분류군 - 해마 (동물)
해마는 말과 유사한 생김새를 가진 물고기로, 수컷이 알을 품는 독특한 생식 방식을 가지며, 전 세계 열대 및 온대 해역에 분포하고 멸종 위기에 처해 CITES 규제를 받는다. - 콘스턴틴 새뮤얼 라피네스크가 명명한 분류군 - 전갱이과
전갱이과는 전갱이목에 속하는 물고기 과로, 몸길이와 형태가 다양하며 전 세계 열대 및 온대 해역에 분포하고, 식용으로 어획되는 종들이 있으며, 4개의 아과와 30여 개의 속으로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 성구동물 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
| 학명 | Sipuncula |
| 명명자 | Rafinesque, 1814 |
| 계 | 동물계 |
| 아계 | 진정후생동물아계 |
| 아계 계급 없음 | 선구동물 가지 |
| 상문 | 관륜동물 상문 |
| 문 | 환형동물문 |
| 강의 하위 분류 | 스지호시무시강 (Sipunculidea) 사메하다호시무시강 (Phascolosomatidea) |
| 하위 분류 | |
| 아강, 목, 과 | 등촉수별벌레아강 (Phascolosomatidea) 등촉수별벌레목 (Aspidosiphonida) 등촉수별벌레과 (Aspidosiphonidae) 별벌레목 (Phascolosomatida) 별벌레과 (Phascolosomatidae) 스지호시무시아강 (Sipunculidea) 골핑기아목 (Golfingiida) 골핑기아과 (Golfingiidae) 파스콜리온과 (Phascolionidae) 스지호시무시과 (Sipunculidae) 테미스티다과 (Themistidae) |
| 추가 정보 | |
| 관련 링크 | 생물학 생물 생물 분류 |
2. 발생
성구동물의 발생은 성숙한 정자나 알이 신관에 채워지면, 이들을 밤이나 이른 아침에 바닷물 속에 방출하여 수정이 이루어진다. 그 후 난할이 진행되어 수정 후 이틀 정도 지나면 트로코포라 유생이 된다. 이때 앞쪽 끝에는 긴 편모가 생기며, 한 쌍의 붉은 눈과 폭이 넓은 섬모대가 있어서 활발히 헤엄쳐 돌아다닌다. 그 후 변태하여 물 밑에 가라앉는데 일주일 정도가 지나면 한 쌍의 작은 촉수가 생기기 시작하여 약 2주 후에는 먹이를 취할 수 있게 된다.
별벌레는 대부분의 다른 환형동물과 유사하게 섬모가 없는 표피와 큐티클로 덮인 바깥쪽 고리층 및 안쪽 세로층의 근육으로 구성된 몸 벽을 가지고 있다. 몸 벽은 체액으로 채워진 체강을 둘러싸고 있으며, 이 체강은 몸 벽 근육이 동물을 확장하거나 수축시키는 정수압골격으로 작용한다. 위협을 받으면 몸을 땅콩 커널과 유사한 모양으로 수축할 수 있어 "땅콩 벌레"라는 이름이 붙었다.[10]
바닷물 속에서 수정된 알은 전할의 나선 난할을 거쳐 배 발생을 진행한다. 극히 일부 종은 직접 발생으로, 수정란은 젤리 모양의 물질에 덮인 채 발생이 진행되어 작은 벌레가 되어 부화한다. 그러나 대다수는 간접 발생으로, 수정란은 트로코포라 유생이 된다. 트로코포라 유생은 섭식하지 않고, 난황의 영양을 사용하여 발생을 진행한다. 트로코포라 유생에서 직접 변태하여 벌레가 되는 것도 있지만, 더 나아가 펠라고스페라 유생 단계를 거치는 것도 있다. 펠라고스페라 유생에는 난황 영양성의 것과, 플랑크톤을 섭식하는 것이 있다.
3. 해부학적 특징
체강은 열체강성의 진체강이며, 복막에 의해 구획되어 있지만 완전하지 않고 하나로 연결되어 있다. 뚜렷한 혈관계는 보이지 않으며, 몸의 운동에 의해 체강액이 섞이는 것이 순환계의 기능을 한다. 체강액에는 헤메리트린을 포함하는 적혈구가 있어 체표를 통해 얻은 산소를 운반한다. 가스 교환 장소는 종에 따라 다르며, 촉수와 주둥이 표면을 모두 사용하는 경우도 있지만, 바위에 구멍을 뚫는 종이나 산소가 부족한 진흙 등에 잠기는 종은 촉수 표면만으로 호흡한다. 대형 종은 체표 전체를 사용할 가능성이 있다. 체강액 내에는 적혈구 외에도 여러 종류의 세포가 포함되며, 압력을 통해 몸을 지지하거나 주둥이를 뻗는 역할을 한다.
신관은 한 쌍 또는 하나가 체간 전부 복면 쪽에 있으며, 외신공으로 체표에, 내신공으로 체강에 연결된다. 질소 노폐물은 신관을 통해 암모니아로 배설된다.
작은 뇌(뇌 신경절)와 하나의 복신경삭이 있으며, 이 둘을 잇는 식도신경환이 체벽을 따라 뻗어 있다. 이 구조는 환형동물과 비슷하지만, 별벌레에서는 복신경삭이 분절되지 않는다는 차이가 있다. 촉각수용기는 체표 전체에 있지만, 특히 촉수에 많다. 화학수용기는 주둥이 등쪽에 보이며, 뇌 신경절 등쪽에 한 쌍의 색소컵 단안을 가진 종도 많다.
3. 1. 몸 구조
별벌레는 길이가 2mm에서 720mm에 이르며, 대부분의 종은 10cm 미만이다. 별벌레의 몸은 분절되지 않은 둥근 몸통과 "내반(introvert)"이라고 불리는 더 좁고 앞쪽 부분으로 나뉜다. 별벌레는 대부분의 다른 환형동물과 다소 유사한 몸 벽을 가지고 있는데, 이는 섬모가 없는 표피와 큐티클로 덮인 바깥쪽 고리층과 안쪽 세로층의 근육으로 구성된다(분절되지 않음). 몸 벽은 몸 벽 근육이 동물을 확장하거나 수축시키는 정수압골격으로 작용하는 체액으로 채워진 체강을 둘러싸고 있다. 위협을 받으면 별벌레는 몸을 땅콩 커널과 유사한 모양으로 수축할 수 있는데, 이로 인해 "땅콩 벌레"라는 이름이 붙었다. 내반은 몸통 벽에서 내반의 부착 지점까지 좁은 리본처럼 뻗어 있는 두 쌍의 수축근에 의해 몸통 안으로 당겨진다. 몸통 벽의 근육을 수축시켜 몸통에서 튀어나오게 할 수 있으며, 이로 인해 체액이 앞으로 밀려나온다.[10] 내반은 몸통 길이의 절반에서 몇 배에 이르기까지 크기가 다양하지만, 비교 크기가 어떻든 완전히 수축할 수 있다.[11]
입은 동물의 앞쪽 끝에 위치해 있으며, 별벌레아강에서 입은 18~24개의 섬모가 있는 촉수 덩어리로 둘러싸여 있는 반면, Phascolosomatidea강에서는 촉수가 입 위쪽의 호를 이루며, 내반 끝에 위치한 목덜미 기관을 둘러싸고 있다. 촉수 각각에는 음식물이 섬모에 의해 입으로 이동하는 깊은 홈이 있다.[11] 이들은 물이나 기질에서 유기물 부스러기를 수집하는 데 사용되며, 아가미 기능도 한다. Themistidae과에서는 촉수가 정교한 왕관 모양의 구조를 형성하며, 이 그룹의 구성원은 다른 별벌레 그룹과 달리 여과 섭식에 특화되어 있다. 다른 별벌레 그룹은 부식자이다.[12] 촉수는 속이 비어 있으며 내반과 유사한 방식으로 정수압에 의해 확장되지만, 식도 옆에 있는 하나 또는 두 개의 수축성 낭에 일련의 도관을 통해 연결되어 있다는 점에서 내반의 나머지 부분과는 다른 메커니즘을 가지고 있다.[10] 입 근처의 내반에는 종종 갈고리가 존재한다. 이것들은 키틴이 아닌 표피의 단백질 특화로, 고리 모양으로 배열되거나 흩어져 있다.[11] 이들은 바위에서 조류를 긁어내는 데 관여하거나 닻 역할을 할 수 있다.[13]
Aspidosiphonidae과의 세 속(''Aspidosiphon'', ''Lithacrosiphon'' 및 ''Cloeosiphon'')은 항문 및 미부 방패라고 알려진 표피 구조를 가지고 있다. 이것들은 두껍고 단단한 판으로 구성된 덩어리이며, 바위에 구멍을 뚫는 데 사용된다. 항문 방패는 동물의 내반 바로 아래, 몸통의 앞쪽에 위치한 항문 근처에 있으며, 미부 방패는 몸의 뒤쪽에 있다.[13] ''Aspidosiphon''과 ''Lithacrosiphon''에서 항문 방패는 등쪽에 국한되어 내반이 각도를 이루며 나오지만, ''Cloeosiphon''에서는 앞쪽 몸통을 둘러싸고 내반이 중심에서 나온다. ''Aspidosiphon''에서 방패는 단단하고 뿔 모양의 구조이고, ''Lithacrosiphon''에서는 석회질 원뿔이며, ''Cloeosiphon''에서는 개별 판으로 구성되어 있다. 이러한 동물에서 내반이 수축되면 항문 방패가 굴로 들어가는 입구를 막는다. 몸통의 뒤쪽 끝에는 ''Aspidosiphon''에 단단한 미부 방패가 있는 경우도 있는데,[14] 이것은 동물이 굴에 고정되는 데 도움이 되거나 구멍을 뚫는 과정에서 사용될 수 있다.[11]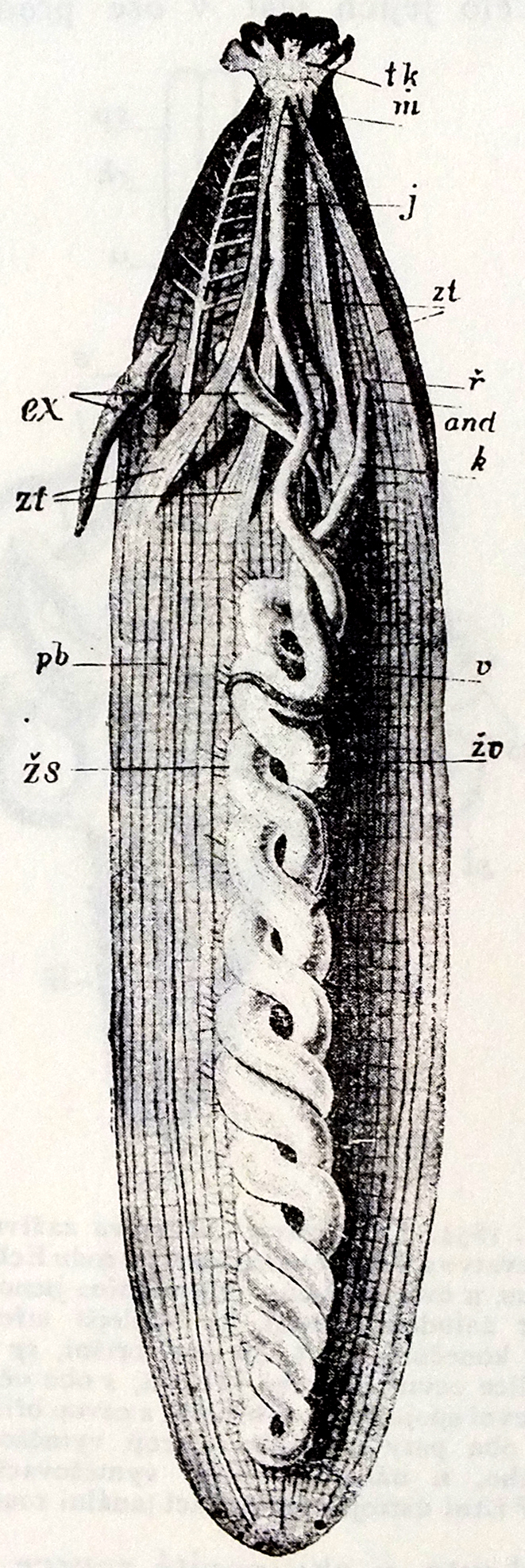
소시지 같은 체형의 벌레로, 체장은 3cm에서 10cm 정도인 것이 많지만, 1cm에 미치지 못하는 작은 종이나 약 50cm에 달하는 대형 종도 있다. 체색은 다양하지만, 칙칙한 색이 많다.
몸은 체간부와 함입주둥이로 이루어진다. 함입주둥이의 선단에 입이 있고, 그 주위 또는 등쪽에 촉수가 다수 있다. 일본어 명칭인 '호시구치(星口)'는 이 촉수가 방사상으로 펼쳐지는 모습에서 유래했다. 체강 내에 1쌍 또는 2쌍의 견인근이 있으며, 이것에 의해 함입주둥이를 체간부에 끌어들일 수 있다. 종에 따라 주둥이를 집어넣고 몸을 부풀린 모습이 땅콩과 비슷하여 peanut worm|피넛 웜영어이라고 불린다. 함입주둥이에는 종종 가시나 갈고리가 늘어서 있다.
체표는 큐티클로 덮여 있으며, 그 표면에는 돌기나 가시가 있는 경우도 많다. 긴다리별벌레과의 것은 체표의 일부에 큐티클이 두꺼워지거나, 석회질이 침착되어 만들어진 방패 모양 부분을 갖는다.
큐티클 아래에는 표피, 이어서 진피가 있다. 그 안쪽에는 근육이 있으며, 바깥쪽에 환상근층, 안쪽에 종주근층이 있는 외에, 이 2층 사이에 얇은 사선근층이 보이는 경우도 있다. 종주근층 아래에는 체강막이 있으며, 그 안쪽이 체강이다.
체강은 열체강성의 진체강이다. 복막에 의해 구획되어 있지만 완전하지 않고, 하나로 연결되어 있다. 뚜렷한 혈관계는 보이지 않으며, 몸의 운동에 의해 체강액이 교반되는 것이 순환계의 기능을 한다. 체강액 중에는 헤메리트린을 포함하는 적혈구가 있으며, 체표를 통해 얻어진 산소를 수송한다. 가스 교환의 장소는 종에 따라 다르며, 촉수와 주둥이의 표면을 함께 사용하는 것도 있지만, 바위에 구멍을 뚫는 종이나, 산소가 부족한 진흙 등에 잠기는 종은, 촉수 표면만으로 호흡을 한다. 대형 종은 체표를 모두 사용할 가능성이 있다.
체강액 내에는 적혈구 외에도 몇 종류의 세포가 포함된다. 체강액은 으로, 압력에 의해 몸을 지지하거나, 주둥이를 뻗는 역할도 한다.
체강의 대부분은 소화관이 차지한다. 소화관은 선단의 입에서 후부로 뻗어, 후단에서 접어, 체간 전단의 배면에 있는 항문으로 이어진다. 따라서 소화관은 U자형이 되며, 뒤로 뻗는 장관과, 후단에서 되돌아오는 장관은 서로 나선형으로 감겨 있다. 이러한 굴곡된 소화관은 별벌레 동물 외에, 고착성인 동물에게 종종 보인다.
신관이 1쌍 또는 1개, 체간 전부의 복면 쪽에 있다. 신관은 외신공으로 체표에, 내신공으로 체강에 연결된다. 질소 노폐물은 신관을 통해 암모니아로 배설된다.
작은 뇌 (뇌 신경절)와 1개의 복신경삭이 있으며, 이 둘을 잇는 식도신경환이 체벽을 따라 뻗어 있다. 이 구조는 환형동물과 비슷하지만, 별벌레에서는 복신경삭이 분절되지 않는다는 차이가 있다. 촉각수용기는 체표 전체에 있지만, 특히 촉수에 많다. 화학수용기는 주둥이의 등쪽에 보인다. 뇌 신경절의 등쪽에 한 쌍의 색소컵 단안을 가진 종도 많다.
3. 2. 소화계
성구동물의 소화관은 함입주둥이 내근(retractor muscle) 사이의 식도에서 시작한다. 몸통에서 내장은 뒤쪽으로 뻗어 고리를 형성하고 다시 앞쪽으로 돌아간다. 창자의 하강 부분과 상승 부분은 서로 꼬여서 이중 나선을 형성한다. 창자 고리가 끝나면 직장이 나타나 몸통 앞쪽 1/3 지점에 위치한 항문에서 끝난다. 소화는 세포 외 소화로, 창자 내강에서 일어난다. 대부분의 종에서 나타나는 직장 맹낭은 창자와 직장의 전환점에 있는 막힌 주머니로, 기능은 알려져 있지 않다. 항문은 내반부가 몸통 안으로 들어갈 때 종종 보이지 않는다.[10]
체강의 대부분은 소화관이 차지한다. 소화관은 입에서 시작하여 뒤쪽으로 뻗어 몸통 후단에서 접혀 돌아와 체간 전단 배면에 있는 항문으로 이어진다. 소화관은 U자형이며, 뒤로 뻗는 부분과 되돌아오는 부분은 서로 나선형으로 감겨 있다. 이러한 굴곡된 소화관은 별벌레 동물 외에, 고착성인 동물에게서도 종종 관찰된다.
3. 3. 순환계
성구동물은 혈관계가 없다. 대신 체강에서 유체 수송과 기체 교환이 이루어지며, 여기에는 호흡 색소인 헤메리트린을 포함하는 별도의 촉수 계통이 있다. 이 두 계통은 격막으로 분리되어 있다. 체강액에는 혈구 세포, 과립구, 큰 다핵 세포, 섬모가 있는 항아리 모양 세포, 미성숙 세포 등 5가지 유형의 체강 세포가 있다. 섬모가 있는 항아리 세포는 복막에 붙어 체강액에서 노폐물을 걸러내는 데 도움을 줄 수 있다.[10]
촉수 체강은 내구 끝의 촉수를 기저부의 고리 관에 연결하며, 여기에서 수축성 혈관이 식도 옆을 따라 뒤쪽으로 뻗어 막힌 채로 끝난다. 일부 증거는 이러한 구조가 초미세 여과에 관여한다는 것을 보여준다.[15] 틈새에 사는 성구동물의 경우, 호흡은 주로 촉수 계통을 통해 이루어지며, 산소는 촉수 체강에서 몸통 체강으로 확산된다. 그러나 다른 종에서는 피부가 얇고 호흡은 주로 몸통의 큐티클을 통해 이루어지며, 여기서 산소 섭취는 표피 바로 아래에 있는 진피 체강관에 의해 촉진된다.[11][16]
뚜렷한 혈관계는 보이지 않으며, 몸의 운동에 의해 체강액이 섞이는 것이 순환계의 기능을 한다. 체강액에는 헤메리트린을 포함하는 적혈구가 있어 체표를 통해 얻은 산소를 운반한다. 가스 교환 장소는 종에 따라 다르며, 촉수와 주둥이 표면을 모두 사용하는 경우도 있지만, 바위에 구멍을 뚫는 종이나 산소가 부족한 진흙 등에 잠기는 종은 촉수 표면만으로 호흡한다. 대형 종은 체표 전체를 사용할 가능성이 있다. 체강액 내에는 적혈구 외에도 여러 종류의 세포가 포함된다. 체강액은 압력을 통해 몸을 지지하거나 주둥이를 뻗는 역할을 한다.
3. 4. 신경계
신경계는 등쪽 뇌 신경절(dorsal cerebral ganglion), 식도 위에 위치한 뇌, 그리고 뇌를 몸통 전체를 따라 뻗어 있는 단일 복측 신경삭과 연결하는 식도 주위 신경환으로 구성된다.[11] 측면 신경은 이 신경삭에서 갈라져 나와 몸 벽의 근육에 분포한다.[11]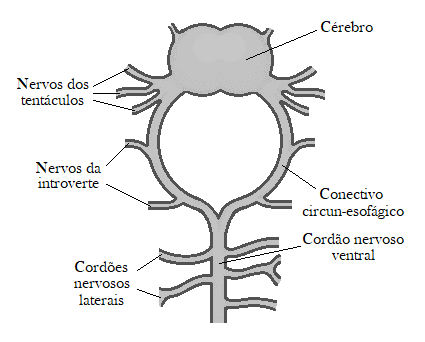
일부 종에서는 뇌와 관련된 단순한 빛 감지 눈점이 있다. 화학수용체 기능을 하는 것으로 보이는 두 기관, 즉 쌍극 신경 세포를 가진 비섬모성 뇌 기관과 뇌 뒤쪽에 위치한 경추 기관이 앞쪽 가장자리 근처에 있다.[10] 비슷한 빛 감지 기관이 파우벨리오프시스과 환형동물에서도 보고되었다.[17] 모든 성구동물은 몸 벽, 특히 주변 환경을 탐색하는 데 사용되는 구기 앞쪽 끝부분에 많은 감각 신경 종말을 가지고 있다.[10]
작은 뇌(뇌 신경절)와 하나의 복신경삭이 있으며, 이 둘을 잇는 식도신경환이 체벽을 따라 뻗어 있다. 이 구조는 환형동물과 비슷하지만, 별벌레에서는 복신경삭이 분절되지 않는다는 차이가 있다. 촉각수용기는 체표 전체에 있지만, 특히 촉수에 많다. 화학수용기는 주둥이 등쪽에 보인다. 뇌 신경절 등쪽에 한 쌍의 색소컵 단안을 가진 종도 많다.
4. 생태
성구동물은 촉수에 있는 섬모와 점액을 이용하여 해저 표면의 데트리투스를 먹는다. 또한 아주 작은 원생동물, 물벼룩, 갑각류, 다모류 유생, 규조류 등을 점액으로 굳혀 입으로 들여보내기도 한다.[19] 이 외에도 퇴적물을 직접 섭취하거나 수중 유기물을 걸러 먹는 종도 있으며, 석회암에 사는 종은 구기에 있는 갈고리나 가시를 이용해 데트리투스를 긁어 먹는다.
4. 1. 서식지
성구동물은 모두 바다에서 서식하며, 주로 얕은 바다의 돌 밑, 바위틈, 바닷말 뿌리 사이, 산호초 사이, 죽은 조개껍질 속, 모래펄 속 깊은 구멍 등에서 발견된다. 갯지렁이에 기생하는 종도 있다.
모든 성구동물은 해양 저서성 생물이며, 극지방, 적도, 심해저를 포함한 전 세계 바다에서 발견되지만, 대부분의 종은 얕은 물에서 비교적 흔하게 발견된다. 이들은 모래, 진흙, 점토 및 자갈 속에 굴을 파고, 돌 밑, 바위 틈새, 속이 빈 산호, 나무, 빈 조개껍데기, 죽은 고래 뼈 내부 등 다양한 서식지에 서식한다. 일부는 다시마 부착기 아래, 갯줄풀 얽힘 속, 해면동물 내부, 다른 유기체의 빈 관 속에 숨어 살며, 일부는 인공 구조물에 있는 오손생물 사이에서 산다.[5][19] 일부는 단단한 바위에 구멍을 뚫어 자신만의 은신처를 만든다.[30]
조간대 퇴적물 표면 아래에서 흔하게 발견되며, 하루 10시간에서 18시간 동안 해저에 잠겨 있을 수 있다. 염분 농도가 낮으면 민감하게 반응하므로 하구 근처에서는 흔히 발견되지 않는다. 산호암에서도 풍부하게 발견될 수 있으며, 하와이에서는 바위 속 굴에서 제곱미터당 최대 700마리의 개체가 발견되기도 했다.[11]

온난한 해역에 분포하는 종이 많지만, 북극해나 남극해를 포함한 전 세계 바다의 조간대에서 수심 5,000m가 넘는 심해까지 폭넓은 수심에 서식한다. 대부분 모래 속이나 돌 밑, 해조류 뿌리 등에 서식하며, 산호초나 연안에는 딱딱한 석회암에 구멍을 뚫는 종이 많다. 고둥 껍질이나 다모류 등이 만든 관을 보금자리로 삼는 종도 있다.
부드러운 기질에 굴을 파거나 해조류 뿌리, 자갈 속을 움직일 때는 세로 근육과 환상 근육, 구기를 움직여 연동 운동을 한다. 딱딱한 기질에 구멍을 뚫는 종은 체표의 가시나 방패 모양 부위를 줄처럼 사용하거나 분비물로 화학적으로 녹이기도 한다. 삿갓혹성동물류는 방패 모양 부위를 뚜껑으로 하여 굴 입구를 막는다.
삿갓혹성동물속의 어떤 종은 스츠보산호속이나 무시노스쵸지가이속의 단체성 산호와 공생한다. 산호 골격에 뚫린 가는 구멍 안에 혹성동물이 사는데, 산호 골격 안에서 조개 껍질이 발견되는 것으로 보아, 혹성동물은 처음에 조개 껍질에 살다가 조개 껍질에 산호가 부착되어 껍질을 덮을 정도로 성장하면서 공생 관계가 성립되는 것으로 추정된다. 이들은 모래 바닥에 서식하지만, 산호는 스스로 거의 움직일 수 없기 때문에 뒤집히거나 모래에 파묻힐 위험이 있다. 혹성동물이 산호를 끌어 움직임으로써 산호는 모래 바닥을 안전하게 이용할 수 있게 된다. 혹성동물 입장에서는 산호가 조개 껍질보다 크게 성장함으로써 자신이 커져도 새로운 보금자리를 찾을 필요가 없어진다. 또한 산호는 광범위하게 먹이를 찾을 수 있고, 혹성동물은 산호의 자포에 의해 외부 적으로부터 몸을 지킬 수 있다는 이점도 있다. 따라서 혹성동물과 산호의 공생은 양쪽 모두에게 이익이 되는 상리 공생 관계라고 생각된다.
개체에 따라 체표 후부에 돌기가 확인되는데, 이는 이 생물에 기생하는 이매패류의 일종으로, 'sisoy'라고 불리며 Erycinacea과에 속하는 것으로 추정된다.[31]
4. 2. 먹이 섭취
대부분의 성구동물은 다양한 방법으로 먹이를 얻는 퇴적물 섭식자이다. 굴에 사는 종은 촉수를 퇴적물 표면 위로 뻗는다. 음식 입자는 점액 분비물에 갇히고 섬모의 운동으로 입까지 운반된다. 모래를 뚫고 들어가는 종들 중에는 촉수가 주름진 형태로 바뀌어 퇴적물과 음식 입자를 긁어모은다. 이 물질의 대부분은 삼켜지지만 더 큰 입자는 버려진다. 틈새에 서식하는 종은 체내를 철회하여 굵어진 몸통으로 틈새 입구를 막고, 동시에 덫에 걸린 음식을 섭취하는 것으로 보인다. 한 종인 ''Thysanocardia procera''는 육식동물로 여겨지며, 어떤 방식으로든 바다쥐의 내부로 들어가 액화된 내용을 빨아낸다.[19]성구동물은 입술을 바다밑 개펄 위에 대고 촉수 표면에 나 있는 섬모를 이용하여 아주 작은 원생동물이나 물벼룩, 갑각류, 다모류의 유생이나 규조류 등을 점액으로 굳혀 입으로 들여보낸다.
채식 행동에는 불명확한 점이 많지만, 대부분의 종은 촉수의 섬모와 점액을 사용하여 해저 표면의 데트리투스를 먹는다고 생각된다. 그 외에 퇴적물을 섭식하는 종과, 수중의 유기물을 걸러 먹는 현탁물 식자도 있다. 석회암에 사는 종은 구기에 있는 갈고리나 가시로 데트리투스를 긁어 먹는다.
5. 생식과 발생
성구동물은 유성 생식과 무성 생식이 모두 가능하지만, 무성 생식은 드물게 나타난다. 성구동물의 발생은 정자나 난자가 성숙하여 신관에 채워지면 밤이나 이른 아침에 바닷물 속에 방출되어 수정이 이루어진다. 수정 후 이틀 정도 지나면 트로코포라 유생이 되며, 이후 변태 과정을 거쳐 성체가 된다.
5. 1. 유성 생식
대부분의 성구동물 종은 자웅이체이다. 이들의 생식 세포는 체강 내벽에서 생성되며, 여기서 체강으로 방출되어 성숙한다. 이 생식 세포는 신장관계를 통해 수집되어 수생 환경으로 방출되어 수정이 일어난다.[10] 적어도 한 종인 ''Themiste pyroides''에서는 산란 직전에 성체가 바위 사이에서 빽빽한 덩어리를 만드는 떼 지어 행동하는 현상이 나타난다.[19]일부 종은 성체 형태로 직접 부화하지만, 많은 종은 종에 따라 하루에서 한 달에 걸쳐 성체로 변태하는 트로코포라 유충을 가지고 있다. 몇몇 종에서 트로코포라는 성체로 직접 발달하지 않고, 확대된 후구(섬모 띠)를 가진 중간 단계인 ''pelagosphaera'' 단계로 발달한다.[20] 변태는 적합한 서식 조건에서만 발생하며, 성체의 존재에 의해 유발된다.[21][22]
온대에서는 여름부터 초겨울에 번식하는 종이 가장 많지만, 예외도 있다. 번식 행동은 야간에 행해지는 경우가 많다. 배우자는 신관을 통해 체외로 방출되어, 해수 중에서 수정(체외수정)한다. 수컷이 먼저 정자를 방출하면, 암컷이 물 속의 정자에 반응하여 난자를 방출한다는 관찰이 있지만, 종에 따라서는 반대로 난자가 먼저 방출되는 경우도 있다.
거의 모든 종이 암수딴몸이지만, 예외적으로 주름벌레과의 Nephasoma minutum|네파소마 미누툼la는 자웅동체이다. 다만 자가수정은 하지 않는다. 이 종에서는 수정란이 어미의 굴 속에서 육아된다.
5. 2. 무성 생식
''Aspidosiphon elegans''와 ''Sipunculus robustus''에서만 드물게 무성 생식이 관찰되었다. 이들은 횡적 분열을 통해 무성 생식을 하며, 필수적인 신체 구성 요소의 재생이 뒤따른다.[18] ''S. robustus''는 출아로도 번식한다.[18] 성구동물의 한 종인 ''Themiste lageniformis''는 처녀생식으로 번식하는 것으로 기록되었는데, 정자가 없는 상태에서 생산된 난자는 정상적인 단계를 거쳐 발달했다.[18]일부 종에서는 높은 재생 능력에 의한 무성 생식도 확인되었다. 이 경우 몸이 가로 분열에 의해 후부의 작은 단편과 전부의 큰 단편으로 나뉘어, 각각 잃어버린 부분을 재생시켜 두 개체가 된다.
플로리다에 많이 서식하는 별벌레속의 Themiste lageniformis|테미스테 라게니포르미스la는 암컷만으로 단성 생식을 하는 경우가 있다. 이 종은 수컷이 암컷에 비해 수가 적다.
5. 3. 발생 과정
성구동물의 발생은 성숙한 정자나 알이 신관에 채워지면, 이들을 밤이나 이른 아침에 바닷물 속에 방출하여 수정이 이루어진다. 그 후 난할이 진행되어 수정 후 이틀 정도 지나면 트로코포라 유생이 된다. 이때 앞쪽 끝에는 긴 편모가 생기며, 한 쌍의 붉은 눈과 폭이 넓은 섬모대가 있어서 활발히 헤엄쳐 돌아다닌다. 그 후 변태하여 물 밑에 가라앉는데 일주일 정도가 지나면 한 쌍의 작은 촉수가 생기기 시작하여 약 2주 후에는 먹이를 취할 수 있게 된다.[18]무성 생식과 유성 생식 모두 성구동물에서 발견될 수 있지만, 무성 생식은 드물고, ''Aspidosiphon elegans''와 ''Sipunculus robustus''에서만 관찰되었다. 이들은 횡적 분열을 통해 무성 생식을 하며, 필수적인 신체 구성 요소의 재생이 뒤따른다. ''S. robustus''는 또한 출아로 번식한다.[18] 성구동물의 한 종인 ''Themiste lageniformis''는 처녀생식으로 번식하는 것으로 기록되었다. 정자가 없는 상태에서 생산된 난자는 정상적인 단계를 거쳐 발달했다.[18]
대부분의 성구동물 종은 자웅이체이다. 그들의 생식 세포는 체강 내벽에서 생성되며, 여기서 체강으로 방출되어 성숙한다. 이 생식 세포는 다음으로 신장관계를 통해 수집되어 수생 환경으로 방출되어 수정이 일어난다.[10] 적어도 한 종인 ''Themiste pyroides''에서는 산란 직전에 성체가 바위 사이에서 빽빽한 덩어리를 만드는 떼 지어 행동하는 현상이 나타난다.[19]
일부 종은 성체 형태로 직접 부화하지만, 많은 종은 종에 따라 하루에서 한 달에 걸쳐 성체로 변태하는 트로코포라 유충을 가지고 있다. 몇몇 종에서 트로코포라는 성체로 직접 발달하지 않고, 확대된 후구(섬모 띠)를 가진 중간 단계인 ''pelagosphaera'' 단계로 발달한다.[20] 변태는 적합한 서식 조건에서만 발생하며, 성체의 존재에 의해 유발된다.[21][22]
바닷물 속에서 수정된 알은 전할의 나선 난할을 거쳐 배 발생을 진행한다. 극히 일부 종은 직접 발생으로, 수정란은 젤리 모양의 물질에 덮인 채 발생이 진행되어 작은 벌레가 되어 부화한다. 그러나 대다수는 간접 발생으로, 수정란은 트로코포라유생이 된다. 트로코포라 유생은 섭식하지 않고, 난황의 영양을 사용하여 발생을 진행한다. 트로코포라 유생에서 직접 변태하여 벌레가 되는 것도 있지만, 더 나아가 펠라고스페라 유생 단계를 거치는 것도 있다. 펠라고스페라 유생에는 난황 영양성의 것과, 플랑크톤을 섭식하는 것이 있다.
6. 분류
Sipunculala는 현재 사용하지 않는 속 이름 Sipunculusla의 여성형 변형으로, siphunculusla("작은 관")의 변형이며, 이는 다시 라틴어 siphola에서 유래되었고, 고대 그리스어 (''síphōn'', "관, 파이프")의 지소사이다.[1]
칼 린네는 1767년 그의 저서 Systema Naturaela에서 벌레 Sipunculus nudusla를 처음으로 기술했다.[2] 1814년, 프랑스 동물학자 콩스탕틴 새뮤얼 라피네스크는 "Sipuncula"라는 단어를 과 (현재는 Sipunculidae)를 설명하는 데 사용했으며,[3] 시간이 지남에 따라 이 용어는 전체 강을 지칭하는 데 사용되었다.[4] 이들은 비교적 연구가 덜 된 그룹이며, 전 세계적으로 약 162종이 있는 것으로 추정된다.[5]
별벌레류는 린네의 『』 제12판(1767년)에 기재되어 있지만, 잡다한 무척추동물과 함께 의 일원으로 여겨져 독립적인 분류군이 되지는 않았다. 라마르크와 퀴비에는 별벌레류가 해삼에 가깝다고 생각했다. 1828년에 블랑빌은 별벌레류를 "Sipunculida"라고 명명했지만, 이 그룹에는 일부 기생성 벌레도 포함되어 있었다. 는 별벌레와 갯지렁이, 짚벌레가 환형동물과 극피동물의 중간 형태라고 생각하여 이 동물들을 묶어 위환형동물류(Gephyrea, 그리스어로 다리를 의미하는 gephyrael에서 유래)라고 불렀다.
6. 1. 하위 분류
- 등촉수별벌레강 (Phascolosomatidea)
- * Aspidosiphoniformes
- ** Aspidosiphonidae
- * 등촉수별벌레목 (Phascolosomatiformes)
- ** 등촉수별벌레과 (Phascolosomatidae) - 상어껍질별벌레(''Phascolosoma scolops'') 포함
- 성구강 (Sipunculidea)
- * 미끈이별벌레목 (Golfingiiformes)
- ** 미끈이별벌레과 (Golfingiidae)
- ** Phascolionidae
- ** 수지별벌레과 (Themistidae)
- * 별벌레목 (Sipunculiformes)
- ** 별벌레과 (Sipunculidae)
1959년, 하이먼은 별벌레를 독립적인 동물문으로 분류했다. 하이먼은 별벌레문을 강이나 목으로 나누지 않았지만, 그 후의 연구에 의해, 다음과 같은 2강 4목 6과로 분류되어 있다.
- 줄별벌레강 (Sipunculidea) - 촉수는 입을 둘러싸는 형태로 배열된다.
- 줄별벌레목 (Sipunculiformes) - 세로줄은 분리된다. 신관은 2개 있다.
- ** 줄별벌레과 (Sipunculidae)
- 주머니별벌레목 (Golfingiiformes) - 세로줄은 분리되지 않는다.
- ** 주머니별벌레과 (Golfingiidae) - 촉수는 가지치지 않고, 신관은 2개 있다.
- ** 고둥별벌레과 (Phascolionidae) - 촉수는 가지치지 않고, 신관은 1개뿐이다.
- ** 가지별벌레과 (Themistidae) - 촉수는 가지치고, 신관은 2개 있다.
- 가시별벌레강 (Phascolosomatidea) - 촉수는 입을 둘러싸지 않는다.
- 가시별벌레목 (Phascolosomatiformes) - 체간에 방패 모양 부분을 가지지 않는다.
- ** 가시별벌레과 (Phascolosomatidae)
- 방패별벌레목 (Aspidosiphoniformes) - 체간에 딱딱한 방패 모양 부분을 가진다.
- ** 방패별벌레과 (Aspidosiphonidae)
위의 분류 체계는 형태에 의한 계통 추정의 결과와 거의 일치하지만, 분자 계통학에 근거하여 일부 재검토가 필요하다는 연구자도 있다. 2012년에는 역시 분자 계통에 기초하여 과 수준의 분류를 재편하는 논문이 발표되었다. 그에 따르면, 별벌레는 다음과 같은 6과로 분류된다.
- 줄별벌레과 (Sipunculidae)
- 주머니별벌레과 (Golfingiidae)
- 방패별벌레과 (Aspidosiphonidae)
- 가시별벌레과 (Phascolosomatidae)
- Siphonosomatidae
- Antillesomatidae
6. 2. 계통 진화
성구동물의 계통 진화는 오랫동안 논란의 대상이었다. 처음에는 체절, 강모 등 환형동물의 특징이 없었음에도 불구하고 환형동물로 분류되었다. 이후 형태형성 및 유충 특징을 바탕으로 연체동물과 연관되었으며, 더 나아가 환형동물, 유악동물 등을 포함하는 큰 그룹인 관륜동물(Lophotrochozoa)에 포함되었다.[6][7]79개의 리보솜 단백질을 사용한 계통발생학적 분석 결과, 성구동물은 환형동물 내에 위치하는 것으로 나타났다.[6][7] 미토콘드리아 DNA 분석에서도 환형동물과의 밀접한 관계가 확인되었으며, 초기 유충 단계에서는 환형동물과 유사한 원시적인 신경 분절이 발생한다는 사실도 밝혀졌다.[8][9]
성구동물은 전구동물이며, 그 중에서도 환형동물, 연체동물 등과 함께 관륜동물(Lophotrochozoa)에 속한다. 분자계통학 연구는 성구동물이 환형동물과 유연관계가 있으며, 심지어 환형동물 내에 포함된다는 결과를 보여준다. 스트럭 등의 연구에 따르면 성구동물은 환형동물 중에서 날개갯지렁이류, 흡구충류 다음으로 초기에 분기한 계통으로 추정된다 (단, 흡구충의 위치는 불확실하다).
성구동물은 체절이 없는 것이 특징인데, 이는 조상으로부터 물려받은 형질이 아니라, 한 번 체절을 진화시킨 후에 잃어버린 것으로 보인다. 야마토사메하다별벌레의 신경계 발생 과정은 환형동물과 공통되며, 이는 성구동물의 조상이 체절을 가지고 있었음을 시사한다.
과거 분류 체계는 다음과 같이 2강 4목 6과로 구성되었다.
| 강 | 목 | 과 | 특징 |
|---|---|---|---|
| 줄별벌레강 (Sipunculidea) | 줄별벌레목 (Sipunculiformes) | 줄별벌레과 (Sipunculidae) | 촉수는 입을 둘러싸는 형태, 세로줄 분리, 신관 2개 |
| 주머니별벌레목 (Golfingiiformes) | 주머니별벌레과 (Golfingiidae) | 촉수 가지 없음, 신관 2개 | |
| 고둥별벌레과 (Phascolionidae) | 촉수 가지 없음, 신관 1개 | ||
| 가지별벌레과 (Themistidae) | 촉수 가지, 신관 2개 | ||
| 가시별벌레강 (Phascolosomatidea) | 가시별벌레목 (Phascolosomatiformes) | 가시별벌레과 (Phascolosomatidae) | 체간에 방패 모양 부분 없음 |
| 방패별벌레목 (Aspidosiphoniformes) | 방패별벌레과 (Aspidosiphonidae) | 체간에 딱딱한 방패 모양 부분 존재 |
그러나 분자 계통학 연구 결과를 바탕으로, 2012년에는 과 수준의 분류를 재편하는 논문이 발표되었다. 이에 따르면, 별벌레는 다음과 같은 6과로 분류된다.
- 줄별벌레과 (Sipunculidae)
- 주머니별벌레과 (Golfingiidae)
- 방패별벌레과 (Aspidosiphonidae)
- 가시별벌레과 (Phascolosomatidae)
- (Siphonosomatidae)
- (Antillesomatidae)
7. 화석 기록


별벌레는 부드러운 몸 구조를 가져 화석은 극히 드물며, 몇몇 속에서만 알려져 있다. ''Archaeogolfingia''와 ''Cambrosipunculus''는 중국의 캄브리아기 청장 생물군에서 발견된다. 이 화석들은 왕관군에 속하는 것으로 보이며,[23][24] 별벌레가 약 5억 2천만 년 전인 캄브리아기 초부터 형태학적으로 거의 변하지 않았음을 보여준다.[23]
캐나다 앨버타의 버제스 셰일에서 캄브리아기의 이름 없는 별벌레가 발견되었으며,[25] ''Lecthaylus''는 스코틀랜드 에든버러 근처의 그랜턴 새우층(실루리아기)에서 발견되었다.[26] 별벌레가 만들었을 가능성이 있는 굴의 생흔 화석은 고생대에서 발견되었다.[23]
일부 과학자들은 한때 별벌레와 멸종된 히올리테스(나선형 창자를 공유하는 고생대의 뚜껑 껍데기) 사이에 밀접한 관계가 있다고 가설을 세웠지만, 이 가설은 이후 부인되었다.[27]
화석 기록은 빈약하지만, 중국 남서부 마오톈산의 캄브리아기 지층에서 발견된 Archaeogolfingia|la와 Cambrosipunculus|la 화석은 성구동물로 여겨진다. 이 두 속은 현생 주머니벌레과와 매우 유사하며, 이 그룹의 형태가 장기간에 걸쳐 큰 변화를 겪지 않았음을 시사한다.
그 외에도 성구동물로 여겨졌던 화석들이 있지만, 일부는 다른 분류군임이 밝혀졌고, 나머지도 의문이 있다. 버제스 셰일에서 발견된 오토이아는 처음에는 갯지렁이로 여겨졌으나, 나중에 새예벌레임이 판명되었다. 실루리아기와 석탄기의 Lecthaylus|la, 쥐라기의 Epitrachys|la 등을 성구동물로 간주하는 의견도 있지만, 의심하는 의견도 있다. 소화관 구조가 유사하다는 점에서 원뿔형 껍질을 가진 히오리테스가 성구동물에 가깝다고 주장하는 연구자도 있지만, 이 역시 의문시되고 있다.
성구동물 자체의 화석은 아니지만, 프랑스의 백악기 초기 지층에서 발견된 벌레고둥류의 산호 화석은 갯지렁이와 공생한 흔적을 남기고 있다. 그 외에 성구동물의 생흔화석으로 보이는 굴의 흔적과 바다나리의 혹도 발견되었지만, 그것들이 실제로 갯지렁이의 생흔인지는 불분명하다.
8. 인간과의 관계


성구동물은 식용이나 낚시 미끼로 이용되는 경우가 있다.
8. 1. 식용
성구동물 젤리(토순동)는 중국 동남부의 별미로, 원래 취안저우 인근 안하이에서 유래되었다.[28] 필리핀 비사야 제도에서도 별미로 여겨진다. 근육은 먼저 향신료를 넣은 식초에 담가 준비한 다음, 세비체와 유사한 요리로 다른 재료와 함께 제공된다. 현지 어부들에게는 기본적인 음식이며, 때로는 도시의 레스토랑에서 전채 요리로 볼 수 있다. 이러한 음식 준비 방식은 현지에서 킬라윈 또는 키닐라(kinilaw)라고 불리며, 생선, 소라, 채소에도 사용된다.[29]말린 형태의 성구동물은 베트남에서도 별미로 여겨지며, 반돈 지구 민쩌우 섬 해안에서 잡힌다.[29] 성구동물의 비교적 높은 시장 가격은 지역 어부 가족에게 중요한 수입원이 되었다.[30]

중국 푸젠성의 요리인 토순동은 토순이라고 불리는 별벌레의 일종을 젤리로 만들어 소스를 뿌린 것이다. 산둥성 옌타이시에서는 Sipunculus nudus영어와 갯지렁이류를 구분하지 않고 하이창쯔라고 부르며, 부추와 함께 볶은 것을 찌차이차이하이창이라고 칭하여 먹는다. Sipunculus nudus영어는 베트남에서는 지충이라고 불리며, 채소와 함께 볶아 먹고, 미크로네시아에서도 식용으로 사용되며, 일본의 세토 내해나 큐슈에서는 낚시 미끼로 이용되고 있다.
필리핀 막탄 섬 등 일부 지역에서는 Sipunculus robustus영어를 살포라고 부르며, 식초와 향신료를 곁들여 생식하거나 햇볕에 말린 것을 구워 먹기도 한다. 황돔과 물고기를 낚기 위한 미끼로 사용하기도 한다. 모래 속에 있는 살포의 굴을 발견하고 채집하기 위해서는 숙련된 기술이 필요하며, 전문적인 채집자가 있다. 막탄 섬에서는 Sipunculus영어도 채집되지만, 쓴맛이 나기 때문에 식용으로는 선호되지 않으며, 낚시 미끼나 자양강장을 위한 민간 요법으로 이용된다. 카오하간 섬에서는 살포 외에 더 작은 별벌레류를 바토난, 더 작은 것을 바트레라고 부르며, 채소와 식초로 무쳐 먹는다. 단, 바토난(Holothuria inhabilis영어)과 바트레(Holothuria arenicola영어)는 해삼의 일종이다.[32]
일본에서는 1920년대부터 1930년대 경에 히로시마현 연안에서 채취된 Sipunculus nudus영어나 Sipunculus nudus영어모도키가 낚시 미끼로 이용되었다는 기록이 있지만, 2010년 전후 서일본에서의 조사에 따르면, 유통되는 것은 일본 국외에서 수입된 Sipunculus nudus영어뿐이다.
8. 2. 낚시 미끼
일본의 세토 내해나 큐슈에서는 낚시 미끼로 성구동물을 이용하고 있다. 필리핀의 막탄 섬 등 일부 지역에서는 황돔과 물고기를 낚기 위한 미끼로 사용하기도 한다. 막탄 섬에서는 스지호시무시도 채집되지만, 쓴맛이 나기 때문에 식용으로는 선호되지 않으며, 낚시 미끼나 자양강장을 위한 민간 요법으로 이용된다.1920년대부터 1930년대 경 히로시마현 연안에서 채취된 스지호시무시나 스지호시무시모도키가 낚시 미끼로 이용되었다는 기록이 있지만, 2010년 전후 서일본에서의 조사에 따르면, 유통되는 것은 일본 국외에서 수입된 스지호시무시뿐이다.
참조
[1]
웹사이트
Sipuncula
https://en.oxforddic[...]
Oxford University Press
2019-02-10
[2]
WoRMS
"''Sipunculus (Sipunculus) nudus'' Linnaeus, 1766"
2019-02-27
[3]
서적
Précis des découvertes et travaux somiologiques de m.r C. S. Rafinesque-Schmaltz entre 1800 et 1814 ou Choix raisonné de ses principales découvertes en zoologie et en botanique
https://archive.org/[...]
Royale typographie militaire, aux dépens de l'auteur
[4]
WoRMS
Sipuncula
2019-02-27
[5]
웹사이트
The phylum Sipuncula in the Mediterranean Sea
https://okeanos-dspa[...]
National Centre for Marine Research
2019-02-10
[6]
논문
Phylogenomic analyses unravel annelid evolution
2011-03-03
[7]
논문
Spiralian Phylogenomics Supports the Resurrection of Bryozoa Comprising Ectoprocta and Entoprocta
[8]
논문
A close phylogenetic relationship between Sipuncula and Annelida evidenced from the complete mitochondrial genome sequence of Phascolosoma esculenta
[9]
논문
Sipunculans and segmentation
2009-01
[10]
서적
Invertebrate Zoology
Holt-Saunders International
[11]
서적
Invertebrate Zoology, 7th edition
Cengage Learning
[12]
서적
The Sipuncula: Their Systematics, Biology, and Evolution
https://books.google[...]
Cornell University Press
[13]
서적
Polychaetes & Allies: The Southern Synthesis
https://books.google[...]
Csiro Publishing
[14]
논문
Reconstructing the phylogeny of the Sipuncula
2005-03
[15]
논문
Ultrastructural evidence for the contractile vessel of sipunculans as a possible ultrafiltration site
[16]
서적
Polychaetes & Allies: The Southern Synthesis
https://books.google[...]
Csiro Publishing
[17]
논문
Sipunculid-like ocellar tubes in a polychaete, ''Fauveliopsis cf. adriatica'' (Annelida, Fauveliopsidae): implications for eye evolution
[18]
웹사이트
Sipuncula
https://repository.s[...]
2019-02-06
[19]
서적
Encyclopedia of Marine Invertebrates
https://archive.org/[...]
TFH Publications
[20]
논문
Larval development and metamorphosis in Sipuncula
[21]
논문
Influence of delayed metamorphosis on postsettlement survival and growth in the sipunculan ''Apionsoma misakianum''
[22]
논문
Factors influencing larval metamorphosis in ''Golfingia misakiana'' (Sipuncula)
https://www.ingentac[...]
[23]
논문
Early Cambrian sipunculan worms from southwest China
2004-08-22
[24]
논문
Reconstructing the ancestral annelid
2012-02
[25]
논문
A new Burgess Shale-type assemblage from the "thin" Stephen Formation of the southern Canadian Rockies
https://scholarship.[...]
[26]
논문
A Lower Carboniferous sipunculan from the Granton Shrimp Bed, Edinburgh
[27]
논문
Hyoliths are Palaeozoic lophophorates
http://dro.dur.ac.uk[...]
[28]
웹사이트
Peanut worm jelly: It's what's for dinner
https://neveryetmelt[...]
Never yet melted
2009-12-16
[29]
웹사이트
Women earn a living digging for peanut worms in northern Vietnam
https://tuoitrenews.[...]
Tuoi Tre News
2018-06-16
[30]
간행물
The distribution of peanut-worm (''Sipunculus nudus'') in relation with geo-environmental characteristics.
[31]
논문
Sipunculids from the reef off Mactan Island (Visayas, Central Philippines)
[32]
논문
タツナミガイ (Dolabella auricularia)の内臓の採取活動と食用慣行
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com