엘라스모사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
엘라스모사우루스는 백악기 후기에 서부 내해에 서식했던 장경룡으로, 70개 이상의 목뼈를 가진 매우 긴 목이 특징이다. 가늘고 삼각형 모양의 두개골과 노처럼 생긴 사지를 가지고 있었으며, 작은 물고기나 해양 무척추동물을 먹었을 것으로 추정된다. 1867년 발견되었으며, 1868년 에드워드 드링커 코프에 의해 명명되었으나, 코프의 복원 오류와 조셉 레이디, 오스니엘 찰스 마시와의 논쟁은 "화석 전쟁"으로 알려져 있다. 엘라스모사우루스는 초기에 플레시오사우루스로 분류되었으나, 1869년 엘라스모사우루스과로 재분류되었고, 엘라스모사우루스과의 계통발생학적 위치는 연구에 따라 변화해왔다.
더 읽어볼만한 페이지
- 1868년 기재된 화석 분류군 - 이브리디오수쿠스
이브리디오수쿠스는 쥐라기 바토니안절에 살았던 멸종된 바다악어 속으로, 염분 조절 기능과 강력한 턱을 갖춘 대형 해양 포식자였다. - 백악기의 수장룡 - 크로노사우루스
크로노사우루스는 백악기 전기에 살았던 대형 플리오사우루스류 해양 파충류로, 강력한 턱과 이빨을 가진 최상위 포식자로 추정되나, 모식 표본의 단편성으로 인해 속의 유효성에 대한 논쟁이 있다. - 백악기의 수장룡 - 후타바사우루스
후타바사우루스는 백악기 후기 일본에 서식한 엘라스모사우루스과의 해양 파충류로, 일본 최초의 엘라스모사우루스류 화석 발견 사례이며, 일본 열도에서 중생대 대형 파충류 화석이 발견될 가능성을 제시했다. - 에드워드 드링커 코프가 명명한 분류군 - 수염고래
수염고래는 이빨 대신 케라틴 수염판을 가진 고래류 아목으로, 플랑크톤 등을 걸러 먹고, 저주파음을 내며 장거리 회유를 하며 멸종 위기에 처해 보호받고 있다. - 에드워드 드링커 코프가 명명한 분류군 - 곡경아목
곡경아목은 머리를 옆으로 구부려 껍질 속으로 넣는 특징을 가지며, 잠경아목과 구분되고, 경추 구조, 두개골 절흔, 껍질의 각질판 배열 등에서도 차이를 보이며, 남미늪거북과를 포함한 일부 종은 흡입 섭식을 하고, 주로 강, 호수, 늪 등에 서식하며, 남아메리카, 아프리카, 오스트레일리아 등지에 분포한다.
| 엘라스모사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
![[[로키 산맥 공룡 자원 센터]]에 복원된 골격](https://cdn.onul.works/wiki/noimage.png) | |
| 학명 | Elasmosaurus platyurus |
| 명명자 | 에드워드 드링커 코프, 1868 |
| 속 | 엘라스모사우루스속 |
| 종 | platyurus |
| 파생어 | IPA: /ɪˌlæzməˈsɔːrəs,_moʊ-/ |
| 어원 | '얇은 판'을 뜻하는 그리스어 단어에서 유래 |
| 생물학적 분류 | |
| 시대 | 백악기 후기 (샹파뉴절) |
| 생존 시기 | 약 8,350만 년 전 ~ 7,050만 년 전 |
| 분류 | 파충강 |
| 아강 | 쌍궁아강 |
| 상목 | 기룡류 |
| 목 | 수장룡목 |
| 아목 | 플레시오사우루스아목 |
| 과 | 엘라스모사우루스과 |
| 크기 | |
| 길이 | 약 10.3m |
| 높이 | 약 7.1m |
2. 특징
엘라스모사우루스는 70개가 넘는 목뼈를 가진 매우 긴 목을 가지고 있었으며, 이는 척추동물 중 가장 많은 수이다. 목은 몸길이의 절반 이상을 차지하며, 평소에는 S자 모양으로 구부리고 있다가 먹이를 발견하면 뻗어서 물고기나 오징어 같은 작은 바다 생물을 낚아챘을 것으로 추정된다.
엘라스모사우루스의 목은 길이가 7.1m로 추정된다.[17] 엘라스모사우루스와 그 친척인 알베르토넥테스는 지금까지 살았던 동물 중 목이 가장 긴 동물 중 하나였으며, 알려진 모든 척추동물 중에서 가장 많은 목 척추뼈를 가지고 있었다.[18][21]
다른 엘라스모사우루스과처럼 엘라스모사우루스는 가늘고 삼각형 모양의 두개골을 가지고 있었을 것이다. 주둥이는 둥글었고 위에서 보면 거의 반원을 이루었으며, 전상악골은 중앙선에 낮은 용골을 가지고 있었다. 아마도 각 전상악골에 6개의 이빨이 있었고, 거기에 보존된 이빨은 큰 송곳니처럼 형성되었다. 전상악골 이빨의 수는 일반적으로 더 적은 수를 가진 원시 플레시오사우루스류와 대부분의 다른 엘라스모사우루스과와 엘라스모사우루스를 구별했다. 아래턱 앞부분의 알려진 이빨은 큰 송곳니였고, 턱 뒤쪽의 이빨은 더 작아 보였다. 엘라스모사우루스과의 치아는 일반적으로 이형치아였으며, 앞쪽에서 뒤쪽으로 갈수록 이빨이 점점 작아졌다. 이빨은 서로 맞물렸고, 치관은 가늘고 단면이 둥글었다. 하악 결합은 잘 골화되었으며, 보이는 봉합이 없었다.[18][20]
어깨뼈는 융합되었고 중앙선에서 만났으며, 중간 막대는 전혀 없었다. 가슴뼈대는 가운데에 긴 막대를 가지고 있었는데, 이것은 어린 플레시오사우루스에게는 없다고 여겨지는 진보된 특징으로 생각된다. 좌골은 가운데에서 연결되어 골반을 따라 중간 막대가 존재했는데, 이는 플레시오사우루스에서는 일반적으로 발견되지 않는 특징이다.[20]
개별 목 척추뼈는 특별히 길지 않았지만, 척추뼈의 수가 매우 많아 극도로 긴 목 길이를 가지게 되었다.[17] 엘라스모사우루스는 72개의 목 척추뼈를 가지고 있다는 점에서 다른 모든 사경룡과 달랐다.[21][20]
아틀라스와 축뼈 복합체는 첫 번째 두 개의 목 척추뼈로 구성되어 있으며 두개골 뒤쪽에 연결되어 있었다. 이 척추뼈의 신경궁은 매우 얇고 상당히 높았으며, 신경관은 삼각형 윤곽을 갖도록 했다. 아틀라스와 축 척추뼈의 하단 중앙을 따라 뚜렷한 용골이 있었다.[20]
대부분의 목 척추뼈는 옆으로 압축되었으며, 특히 목의 중간 부분에서 압축되었다. 능선(ridge)이 목 척추뼈의 측면을 따라 세로로 이어져 있었으며(엘라스모사우루스과의 특징), 목 뒤쪽에서 세 번째 척추뼈에서 55번째 척추뼈까지 볼 수 있었다. 이 능선은 목의 근육을 고정하는 역할을 했을 것이다. 중심은 목 척추뼈의 위치에 따라 모양이 달랐다.[18][21][20]
목 척추뼈의 신경궁은 중심에 잘 융합되어 뚜렷한 봉합선이 없었으며, 신경관은 앞쪽 척추뼈에서 좁았고 뒤쪽 척추뼈에서 더 두드러지게 발달하여 너비와 높이가 같았으며 거의 원형이었다. 목 척추뼈의 전관절돌기와 후관절돌기는 인접한 척추뼈가 서로 맞물리도록 연결하는 돌기였으며, 길이가 같았다. 목 척추뼈의 신경 극은 낮았고 20번째 척추뼈 부근에서 거의 반원형인 것으로 보인다. 각 목 척추뼈의 바닥에는 중앙에 영양 구멍 쌍이 있었고, 능선으로 분리되었으며, 이는 목 뒤쪽으로 갈수록 점점 더 두드러지고 두꺼워졌다.[20]
엘라스모사우루스는 3개의 가슴 척추뼈를 가지고 있었는데, 이는 엘라스모사우루스과의 일반적인 숫자이다. 등 척추뼈는 신경관과 같은 높이에 갈비뼈 면을 가지고 있었고, 여기에서 횡돌기의 앞쪽과 뒤쪽 부분은 가장자리에 뚜렷한 능선을 가지고 있었다. 등 척추뼈는 엘라스모사우루스과를 구별하는 데 유용하지 않다.[21][20]
엘라스모사우루스는 4개의 천추를 가지고 있었는데, 이는 엘라스모사우루스과의 일반적인 숫자이다. 첫 번째 꼬리 척추뼈는 더 작은 갈비뼈 면을 가지고 있고 중심의 하반부에 위치해 있다는 점에서 앞선 천추와 구별할 수 있었다. 엘라스모사우루스과의 꼬리 척추뼈의 일반적인 숫자는 30개이다.[20]
2. 1. 신체 구조
엘라스모사우루스는 유선형 몸체와 긴 지느러미발을 가진 전형적인 장경룡의 형태를 띠고 있었다. 목 길이는 6.1-7.1미터[98]로 수장룡 중에서도 매우 길었으며, 전체 몸길이는 10.3미터에 달했다.[99] 목은 76개의 경추로 이루어져 있어, 몸 측면까지 자유자재로 움직일 수 있었다고 한다[100]. 하지만 물의 저항이 커지기 때문에 헤엄치면서 목을 구부리는 동작은 하지 않았을 것으로 추정된다.몸통은 짧고 복늑골을 가지고 있어 유연성이 떨어졌으며, 꼬리는 짧고 지느러미가 없었다. 따라서 몸통을 구불거려 헤엄치는 방식은 불가능했다. 사지는 지느러미발이었으며, 앞쪽 한 쌍이 더 길었다.
2. 2. 목의 움직임과 기능
엘라스모사우루스의 목은 아래쪽으로 75–177도, 위쪽으로 87–155도, 옆으로 94–176도 움직였을 것으로 추정된다.[102] 1869년 코프가 추정한 것처럼 백조처럼 목을 수면 위로 내는 자세는 불가능했을 것으로 여겨진다.[102] 긴 목의 역할은 불분명하지만,[103] 목을 아래로 움직여 해저 가까이에 사는 먹이를 포식하는 것이 가능했을 것으로 추측된다.[104] 3D 모델을 이용한 추측에 따르면, 긴 목을 가진 수장룡은 짧은 목을 가진 수장룡에 비해 항력이 보통 증가하지 않지만, 목을 옆으로 굽힌 자세에서는 증가하는 것으로 나타났다.[105] 또한, 다른 연구에서는 긴 목은 앞으로 헤엄칠 때 저항이 증가하지만, 큰 몸통으로 상쇄된다고 추측된다.[106]
2. 3. 식성
엘라스모사우루스는 작은 경골어류와 해양 무척추동물을 주로 먹었을 것으로 추정된다. 엘라스모사우루스는 먹이를 발견하면 평소 S자 모양으로 구부리고 있던 긴 목을 뻗어서 물고기와 오징어 같은 작은 바다 생물을 낚아채 먹었을 것으로 보인다.[90] 작고 운동성이 없는 두개골은 먹을 수 있는 먹이의 크기를 제한했을 것이며, 먹이를 잡는 데 적합하고 찢는 데 적합하지 않은 길고 가느다란 이빨을 가진 엘라스모사우루스는 먹이를 통째로 삼켰을 것이다.엘라스모사우루스의 모식 표본 척추 바로 아래에서는 여섯 종의 물고기 비늘과 이빨이 발견되었는데, 코프는 이 물고기들이 엘라스모사우루스의 먹이가 되었을 것이라고 추론했다.[4] 근연종인 스틱소사우루스의 화석에서는 엔코두스 등을 포식했다는 증거가 발견되었다.[108]
엘라스모사우루스는 위석을 가지고 있었는데, 이는 소화를 돕거나, 음식 내용물을 혼합하고, 미네랄을 보충하며, 저장 및 부력을 조절하는 데 사용되었을 수 있다.[93] 스틱소사우루스의 한 표본은 가슴띠 뒤 복부 부위에 조각난 물고기 뼈와 돌을 포함하고 있었는데, 이 돌은 표본이 발견된 곳에서 600km 떨어진 곳의 암석과 일치하는 것으로 밝혀졌다.[92]
3. 연구사
1867년 초, 미국 군의 외과 의사 더필러스 헌트 터너와 군 정찰병 윌리엄 콤스톡은 캔자스주 포트 윌리스 주변에서 화석을 탐험하던 중 큰 화석 파충류의 뼈를 발견했다.[20][1] 터너는 피에르 셰일 지층의 협곡에서 발견한 이 뼈들이 "멸종된 괴물"의 것이라고 생각하고, 존 르콩트를 통해 필라델피아 자연과학 아카데미의 고생물학자 에드워드 드링커 코프에게 척추뼈 표본을 보냈다. 코프는 이 화석이 유럽에서 본 것보다 더 큰 플레시오사우루스의 유해임을 확인하고, 터너에게 나머지 표본을 보내달라고 요청했다.[20][1]
터너와 포트 윌리스의 다른 사람들은 현장으로 돌아가 더 많은 척추뼈와 뼈들을 약 362.87kg 가량 회수했다. 이 화석들은 말과 마차를 통해 포트 윌리스로 운송되었고, 코프의 지시에 따라 포장되어 철도를 통해 필라델피아로 보내졌다. 1868년 3월, 코프는 표본을 검사한 후 ANSP 회의에서 ''엘라스모사우루스 플라티우루스''(Elasmosaurus platyurus)라고 명명했다. 속명 ''엘라스모사우루스''는 "얇은 판 파충류"를, 종명 ''플라티우루스''는 "납작한 꼬리"를 의미한다.[1][2][3]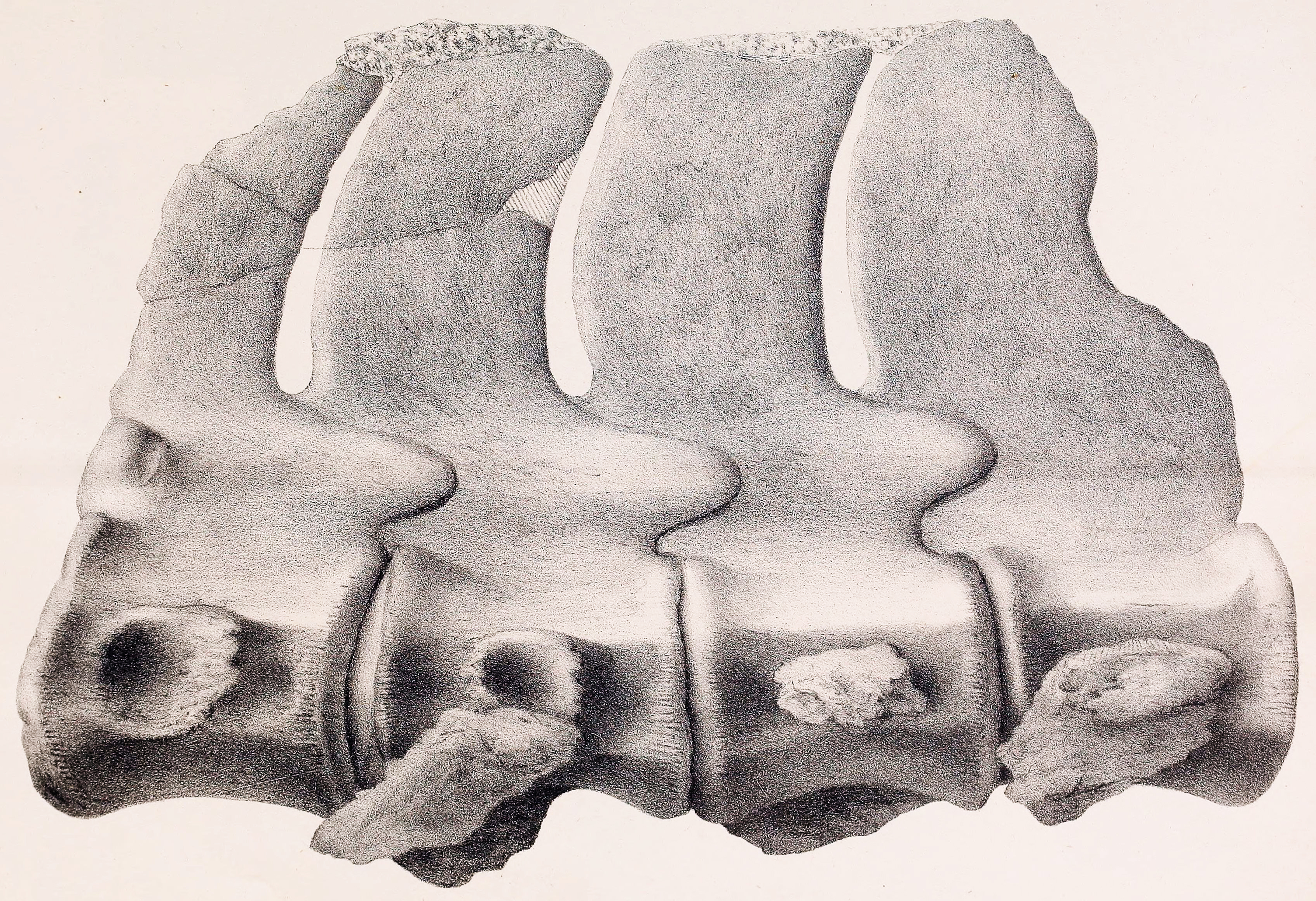
코프는 터너에게 더 많은 화석을 요청했고, 1868년 8월 또는 9월에 추가 화석이 보내졌다. ANSP는 터너의 "매우 귀중한 선물"에 감사를 표했지만, 터너는 1869년 7월에 예기치 않게 사망했다. 터너가 표본을 발견한 상황은 1987년에 그의 편지가 출판되기 전까지 알려지지 않았다.[1] ''엘라스모사우루스''는 캔자스에서 최초로 발견된 주요 화석이었으며, 이후 캔자스에서 수천 개의 화석이 미국 동부 해안의 박물관으로 보내지는 화석 수집 러시의 시작을 알렸다. 또한 당시 신세계에서 알려진 몇 안 되는 플레시오사우루스 중 하나였으며, 긴 목을 가진 플레시오사우루스과의 최초로 확인된 구성원이었다.[20]
1869년, 코프는 ''엘라스모사우루스''를 과학적으로 기술하고 그림으로 나타냈으며, 프리프린트 버전의 원고에는 다른 플레시오사우루스와 달리 짧은 목과 긴 꼬리를 가진 ''엘라스모사우루스''를 보여주는 골격 재구성이 포함되어 있었다. 그러나 1870년 3월, 조셉 레이디는 코프의 재구성이 척추 기둥의 잘못된 끝에 두개골을 배치한 오류를 지적했다. 레이디는 또한 ''엘라스모사우루스''가 1851년에 그가 명명한 플레시오사우루스 ''디스코사우루스''와 동일하다고 결론 내렸다.[4][6][5][1]
3. 1. 코프의 복원 오류와 화석 전쟁
에드워드 드링커 코프는 엘라스모사우루스를 복원하는 과정에서 두개골을 꼬리 끝에 붙이는 실수를 저질렀다. 이를 오스니엘 찰스 마시가 지적하면서 두 학자 사이에 "화석 전쟁"이라 불리는 치열한 경쟁이 시작되었다.[1][6][7][8][9][64]코프는 자신의 실수를 숨기기 위해 발표했던 논문을 회수하려 했으나, 마시가 이 실수를 공개적으로 비판하면서 두 학자 간의 갈등은 더욱 심화되었다.
코프가 이러한 명백한 해부학적 실수를 저지른 이유에 대해서는 여러 추측이 제기되었다. 당시 엘라스모사우루스는 독특한 표본이었기 때문에 해석하기 어려웠을 것이라는 주장도 있고, 코프가 젊고 경험이 부족했기 때문이라는 분석도 있다. 그러나 코프는 당시 플레시오사우루스 해부학에 대해 충분한 지식을 가지고 있었기 때문에 이러한 실수는 쉽게 피할 수 있었다는 반론도 존재한다.[6]
3. 2. 분류
코프는 원래 엘라스모사우루스를 플레시오사우루스로 인식했지만, 1869년 논문에서 ''Cimoliasaurus''와 ''Crymocetus''와 함께 새로운 사우롭테리기아 파충류목에 배치했다. 그는 이 그룹에 스트렙토사우리아(Streptosauria) 또는 "거꾸로 된 도마뱀"이라는 이름을 붙였는데, 이는 다른 척추동물에서 보이는 것과 비교하여 개별 척추뼈의 방향이 반대이기 때문이라고 추정했다.[64][65] 그는 이후 1869년 ''Elasmosaurus''의 설명에서 이 아이디어를 포기했는데, 여기서 그는 Leidy의 ''Cimoliasaurus''에 대한 잘못된 해석에 근거했다고 말했다.[4] 이 논문에서 그는 또한 새로운 과인 엘라스모사우루스과(Elasmosauridae)에 ''Elasmosaurus''와 ''Cimoliasaurus''를 포함시켰다. 이 과 내에서 그는 전자는 압축된 척추뼈를 가진 더 긴 목으로, 후자는 사각형의 움푹 들어간 척추뼈를 가진 더 짧은 목으로 구별되는 것으로 간주했다.[4]
그 후 수년에 걸쳐 엘라스모사우루스과는 플레시오사우르스가 분류되는 세 그룹 중 하나가 되었으며, 다른 그룹은 플리오사우루스과와 플레시오사우루스과였다(때로는 하나의 그룹으로 병합됨).[66] 찰스 앤드루스는 1910년과 1913년에 엘라스모사우루스과와 플리오사우루스과의 차이점을 자세히 설명했다. 그는 엘라스모사우루스과를 긴 목과 작은 머리, 그리고 앞다리 구동 운동을 위한 튼튼하고 잘 발달된 어깨뼈(그러나 위축되거나 없는 빗장뼈 및 쇄골 사이 뼈)로 특징지었다. 한편, 플리오사우루스과는 짧은 목을 가지고 있었지만 큰 머리를 가지고 있었으며 뒷다리 구동 운동을 사용했다.[67][68]
1940년에 시어도어 화이트는 서로 다른 플레시오사우르 과 사이의 상호 관계에 대한 가설을 발표했다. 그는 엘라스모사우루스과를 플리오사우루스과와 가장 가깝다고 생각했는데, 이는 상대적으로 좁은 갈고리뼈뿐만 아니라 쇄골 사이 뼈나 빗장뼈가 없다는 점을 지적했다. 엘라스모사우루스과에 대한 그의 진단은 또한 중간 길이의 두개골(즉, 중간 두개골), 목 갈비뼈가 한두 개의 머리를 가짐, 어깨뼈와 갈고리뼈가 정중선에서 접촉, 갈고리뼈의 둔한 뒤쪽 외부 각도, 그리고 플리오사우루스과와 비교하여 어깨뼈-갈고리뼈 복합체에 있는 한 쌍의 구멍(창)이 더 좁은 뼈로 구분된다는 점을 지적했다.[70]
웰스는 1943년 플레시오사우루스에 대한 그의 수정에서 화이트의 분류에 이의를 제기하며, 화이트의 특징은 보존과 개체 발생 모두에 의해 영향을 받는다고 지적했다. 그는 목 길이, 머리 크기, 궁둥뼈 길이, 상완골과 대퇴골(지느러미)의 가늘음에 따라 플레시오사우루스를 플레시오사우루스상과와 플리오사우루스상과로 나눴다. 각 상과는 갈비뼈의 머리 수와 상지 지느러미(epipodialia)의 비율에 따라 더 세분화되었다. 따라서 엘라스모사우루스과는 긴 목, 작은 머리, 짧은 궁둥뼈, 튼튼한 지느러미, 단일 머리 갈비뼈, 짧은 상지 지느러미를 가지고 있었다.[34]
1997년 카펜터의 플레시오사우르스의 계통 발생학적 분석은 목 길이에 따른 플레시오사우르스의 전통적인 세분화에 도전했다. 폴리코틸리드는 이전에 플리오사우루스상과에 속했지만, 카펜터는 유사성에 기초하여 폴리코틸리드를 엘라스모사우루스과의 자매군으로 이동시켜 폴리코틸리드와 플리오사우르가 짧은 목을 독립적으로 진화했음을 암시했다.[61]
엘라스모사우루스과 내에서, ''Elasmosaurus'' 자체는 매우 다양한 관계를 가진 "와일드카드 분류군"으로 간주되어 왔다.[79] 2020년 오고먼은 ''Elasmosaurus''를 그룹 내에 포함시켜 Styxosaurinae를 공식적으로 Elasmosaurinae와 동의어로 만들었으며, 또한 이 분류군에 대한 진단적 특성의 목록을 제공했다.[80]

4. 고생태
엘라스모사우루스는 약 8천 5백만 년 전 후기 백악기 캄파니아절 시대에 북아메리카 중부를 덮고 있던 얕은 바다인 서부 내해에 서식했다.[95][94] 엘라스모사우루스과는 유선형 몸체와 긴 지느러미를 가지고 있어 바다 생활에 완전히 적응했으며,[18] 지속적인 헤엄을 위해 항온성(일정한 체온)을 유지했을 가능성도 있다.[81] 지느러미는 매우 뻣뻣하고 특화되어 있어 육지에 올라와 알을 낳는 대신 바다뱀처럼 새끼를 낳는 방식(태생)으로 번식했을 가능성이 높다.[84]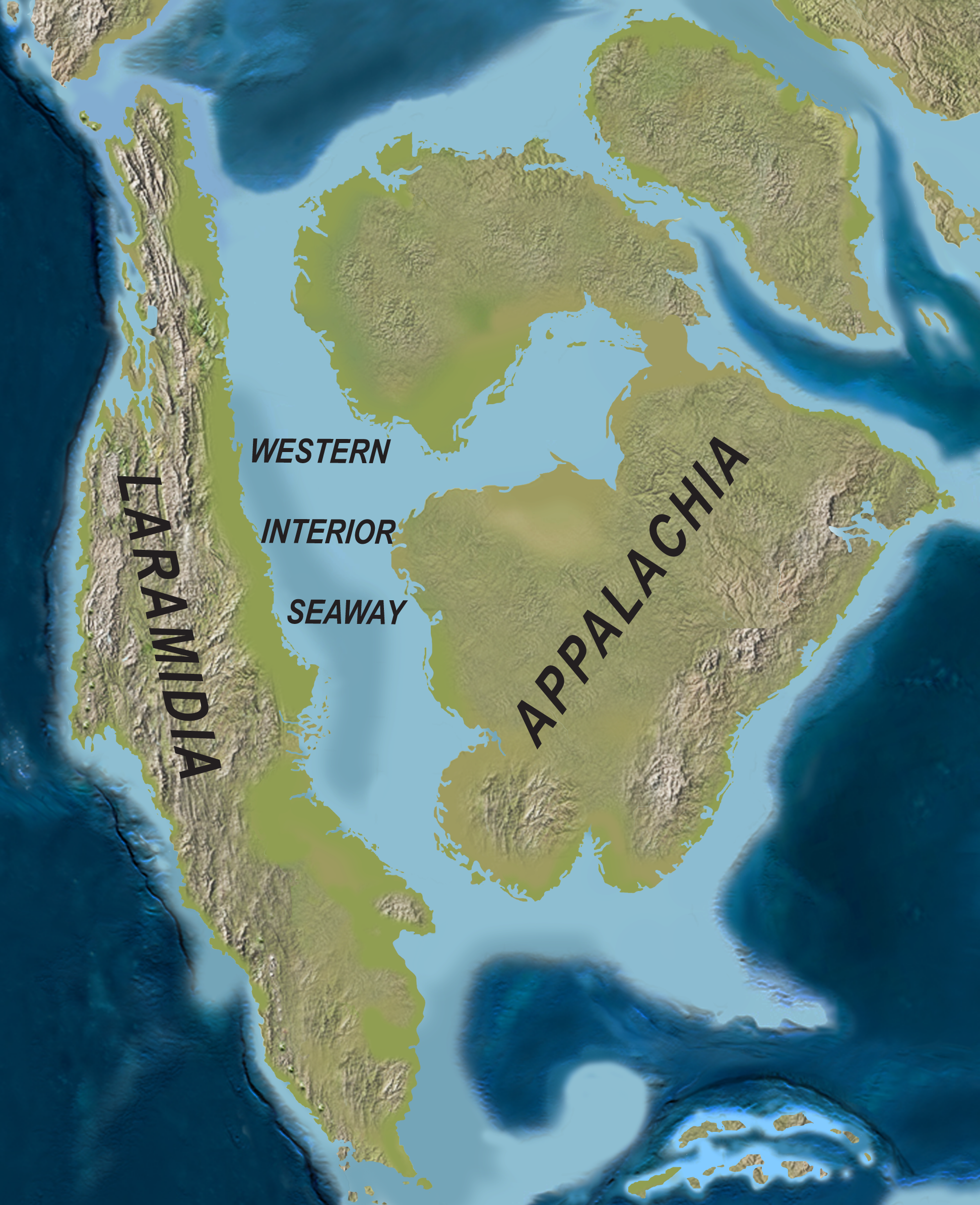
서부 내해는 최대 규모일 때 로키 산맥에서 동쪽 애팔래치아 산맥까지 약 1000km 폭으로 뻗어 있었고, 가장 깊었을 때는 800m 깊이였다.[95] 이 바다에는 다양한 종류의 해양 생물들이 서식했는데, 바닥에는 굴로 덮인 큰 ''Inoceramus'' 조개가 주를 이루었고, 루디스트, 바다나리, 두족류(오징어 및 암모나이트 포함) 등도 서식했다.
이 바다에 서식했던 큰 물고기로는 경골어류인 ''Pachyrhizodus'', ''Enchodus'', ''Cimolichthys'', ''Saurocephalus'', ''Saurodon'', ''Gillicus'', ''Ichthyodectes'', ''Xiphactinus'', ''Protosphyraena'', ''Martinichthys'' 등이 있었고,[95] 상어인 ''Cretoxyrhina'', ''Cretolamna'', ''Scapanorhynchus'', ''Pseudocorax'', ''Squalicorax'' 등이 있었다. 엘라스모사우루스 외에도 ''Libonectes'', ''Styxosaurus'', ''Thalassomedon'', ''Terminonatator'', ''Polycotylus'', ''Brachauchenius'', ''Dolichorhynchops'', ''Trinacromerum'' 등의 플레시오사우루스류와[95] ''Mosasaurus'', ''Halisaurus'', ''Prognathodon'', ''Tylosaurus'', ''Ectenosaurus'', ''Globidens'', ''Clidastes'', ''Platecarpus'', ''Plioplatecarpus'' 등의 모사사우루스류, 그리고 ''Archelon'', ''Protostega'', ''Porthochelys'', ''Toxochelys'' 등의 바다거북이 있었다. 또한, 날지 못하는 수생 조류인 ''Hesperornis''와 익룡인 ''Pteranodon''과 ''Nyctosaurus'', 조류인 ''Ichthyornis''도[96] 발견된다.
엘라스모사우루스는 경골어류와 해양 무척추동물을 주로 먹었으며,[90] 원양에서 활동적인 사냥꾼이었을 가능성이 있다.[90] 이들은 물고기 떼를 은밀히 따라가 머리를 위로 움직여 사냥하거나, 아래에서 어두운 물 속에 숨어 먹이를 사냥했을 것으로 추정된다. 엘라스모사우루스와 같은 엘라스모사우루스과 해양 파충류들은 상어와 같은 다른 포식자들의 먹이가 되기도 했다. 화석 증거를 통해 엘라스모사우루스과가 ''Cretoxyrhina''[85] 상어에게 공격받은 흔적을 확인할 수 있다.
참조
[1]
논문
Captain Theophilus H. Turner and the Unlikely Discovery of ''Elasmosaurus platyurus''
2017
[2]
논문
Scattered and Shattered: A Brief History of the Early Methods of Digging, Preserving and Transporting Kansas Fossils
2017
[3]
논문
Remarks on a new enaliosaurian, ''Elasmosaurus platyurus''
https://archive.org/[...]
[4]
논문
Synopsis of the extinct Batrachia, Reptilia and Aves of North America, Part I
https://www.biodiver[...]
2017-11-08
[5]
논문
Remarks on Elasmosaurus platyurus
http://oceansofkansa[...]
2017-11-21
[6]
논문
Bonehead mistakes: The background in scientific literature and illustrations for Edward Drinker Cope's first restoration of ''Elasmosaurus platyurus''
[7]
뉴스
Wrong End Foremost
http://www.oceansofk[...]
New York Herald
2009-02-02
[8]
논문
On ''Elasmosaurus platyurus'' Cope
http://oceansofkansa[...]
2017-09-26
[9]
논문
Synopsis of the extinct Batrachia, Reptilia and Aves of North America, Part I
https://www.biodiver[...]
2017-11-11
[10]
논문
How elongated? The pattern of elongation of cervical centra of ''Elasmosaurus platyurus'' with comments on cervical elongation patterns among plesiosauromorphs
2024
[11]
논문
North American plesiosaurs ''Elasmosaurus'', ''Cimoliasaurus'', and ''Polycotylus''
https://zenodo.org/r[...]
2022-12-09
[12]
논문
The curious case of Central Park's dinosaurs: The destruction of Benjamin Waterhouse Hawkins' Paleozoic Museum revisited
2023
[13]
논문
The Mystery of ''Elasmosaurus platyurus'' Cope 1868 – Where is the rest of the type specimen?
2018
[14]
논문
Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of ''Elasmosaurus platyurus'' Cope 1868?
http://www.palarch.n[...]
2017-09-26
[15]
논문
Postscript to Everhart, M.J. 2005. "Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of ''Elasmosaurus platyurus'' Cope 1868?" – PalArch's Journal of Vertebrate Palaeontology 4, 3: 19–32
http://www.palarch.n[...]
2017-11-13
[16]
논문
Reste eines Elasmosauriers aus der Oberkreide von Schleswig-Holstein in der Sammlung des Naturkunde-Museums Bielefeld
https://www.research[...]
2017
[17]
논문
Why sauropods had long necks; and why giraffes have short necks
2013
[18]
웹사이트
Fossil Focus: Elasmosaurs
http://www.palaeonto[...]
Palaeontology Online
2018-01-18
[19]
논문
A Small Body Sized Non-Aristonectine Elasmosaurid (Sauropterygia, Plesiosauria) from the Late Cretaceous of Patagonia with Comments on the Relationships of the Patagonian and Antarctic Elasmosaurids
[20]
논문
Redescription of ''Elasmosaurus platyurus'', Cope 1868 (Plesiosauria: Elasmosauridae) from the Upper Cretaceous (lower Campanian) of Kansas, U.S.A
https://www.research[...]
[21]
논문
Revised vertebral count in the "longest-necked vertebrate" ''Elasmosaurus platyurus'' Cope 1868, and clarification of the cervical-dorsal transition in Plesiosauria
[22]
논문
Revision of North American elasmosaurs from the Cretaceous of the western interior
https://www.research[...]
[23]
논문
A taxonomic reappraisal of the families Elasmosauridae and Cryptoclididae (Reptilia: Plesiosauroidea)
[24]
논문
On the reptilian orders Pythonomorpha and Streptosauria
https://www.biodiver[...]
2017-11-12
[25]
논문
April 5th
1870
[26]
논문
The Fossil Reptiles of New Jersey (Continued)
1869
[27]
논문
Additional note on ''Elasmosaurus''
http://oceansofkansa[...]
2017-11-21
[28]
서적
A History of British Fossil Reptiles
Cassell & Company Ltd
1850
[29]
논문
A new species of elasmosaur from the Aptian of Columbia and a review of the Cretaceous plesiosaurs
https://books.google[...]
1962
[30]
논문
Cenomanian–Turonian marine amniote remains from the Saxonian Cretaceous Basin of Germany
2017
[31]
논문
A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group
http://paleoarchive.[...]
2017-11-17
[32]
논문
''Plesiosaurus houzeaui'' Dollo, 1909 from the Upper Campanian of Ciply (Belgium) and a review of the Upper Cretaceous plesiosaurs from Europe
http://biblio.natura[...]
2017-11-17
[33]
논문
Report on the Geology of the Region of the Judith River, Montana: And on Vertebrate Fossils Obtained on Or Near the Missouri River
https://www.biodiver[...]
2017-11-25
[34]
논문
Elasmosaurid plesiosaurs with description of new material from California and Colorado
1943
[35]
논문
Taxonomic reassessment of ''Hydralmosaurus'' as ''Styxosaurus'': new insights on the elasmosaurid neck evolution throughout the Cretaceous
2016
[36]
서적
Water Reptiles of the Past and Present
https://archive.org/[...]
University of Chicago Press
1914
[37]
논문
A specimen of ''Elasmosaurus serpentinus''
https://www.biodiver[...]
1939
[38]
간행물
Re-description of ''Thalassomedon haningtoni'' – an elasmosaurid from the Cenomanian of North America
https://www.research[...]
Museum of Evolution, Uppsala University
2016
[39]
논문
On the Structure of the Skull in the Plesiosaurian Reptilia, and on Two New Species from the Upper Cretaceous
1894
[40]
논문
A review of North American Cretaceous elasmosaurs
1952
[41]
논문
The occurrence of elasmosaurids (Reptilia: Plesiosauria) in the Niobrara Chalk of Western Kansas
https://www.research[...]
2006
[42]
논문
Structure of the Plesiosaurian Skull
https://zenodo.org/r[...]
1890
[43]
논문
An examination of Plesiosauria (Diapsida: Sauropterygia) from the Niobrara Chalk (Upper Cretaceous) of central North America
https://www.academia[...]
1999
[44]
논문
North American Plesiosaurs: ''Trinacromerum''
https://zenodo.org/r[...]
2019-06-26
[45]
논문
Reptiles from the Senonian (U. Cret.) of Scania (S. Sweden)
http://paleoarchive.[...]
2017-11-19
[46]
논문
Геологическое распространенiе эласмозавровъ
http://www.mathnet.r[...]
2017-11-18
[47]
논문
Sur la présence de l#REDIRECT
http://paleoglot.org[...]
2017-11-18
[48]
서적
Каталог местонахождений остатков морских рептилий в юр ских и меловых отложениях Нижнего Поволжья
Saratov State University Press
1999
[49]
서적
The Age of Dinosaurs in Russia and Mongolia
https://books.google[...]
Cambridge University Press
2000
[50]
논문
The first plesiosaur from the Primorye krai
http://doc.rero.ch/r[...]
2005
[51]
서적
Zoologia, Vol. 2
Imprenta Maulde y Renou
1848
[52]
서적
Catalogue of the fossil Reptilia and Amphibia in the British Museum. Part II
https://www.biodiver[...]
Printed by order of the Trustees
1889
[53]
논문
Ueber Saurierreste aus den Quiriquina−Schichten
https://books.google[...]
1895
[54]
논문
A new Cretaceous plesiosaur from Venezuela
1949
[55]
논문
Presence of ''Mauisaurus'' in the Maastrichtian (Late Cretaceous) of central Chile
2010
[56]
논문
Postcranial morphology of ''Aristonectes'' (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of Patagonia and Antarctica
2013
[57]
논문
A reappraisal of the Late Cretaceous Weddellian plesiosaur genus ''Mauisaurus'' Hector, 1874
2017
[58]
논문
Notice of some Saurian Fossils discovered by J. H. Hood, Esq., at Waipara, Middle Island, New Zealand
https://zenodo.org/r[...]
1870
[59]
논문
Late Cretaceous reptiles (Families Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand
1986
[60]
논문
A new elasmosaur from the Eagle Ford Shale of Texas
https://sites.smu.ed[...]
2017-11-19
[61]
서적
Ancient Marine Reptiles
https://archive.org/[...]
Academic Press
1997
[62]
논문
Postcranium of the paradigm elasmosaurid plesiosaurian ''Libonectes morgani'' (Welles, 1949)
2015
[63]
논문
Food resources and habitat selection of a diverse vertebrate fauna from the upper lower Campanian of the Kristianstad Basin, southern Sweden
2013
[64]
논문
''Elasmosaurus platyurus'' and a page from the Cope-Marsh war
https://www.research[...]
1984
[65]
논문
On the reptilian orders, Pythonomorpha and Streptosauria
http://oceansofkansa[...]
2017-11-23
[66]
서적
A Cladistic Analysis and Taxonomic Revision of the Plesiosauria (Reptilia: Sauropterygia)
http://mds.marshall.[...]
Acta Zoologica Fennica
2017-11-26
[67]
서적
A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay
British Museum (Natural History)
1910
[68]
서적
A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay
British Museum (Natural History)
1913
[69]
서적
The Osteology of the Reptiles
Harvard University Press
1925
[70]
논문
Holotype of ''Plesiosaurus longirostris'' Blake and Classification of the Plesiosaurs
1940
[71]
논문
Preliminary description and phylogenetic position of a new plesiosaur (Reptilia: Sauropterygia) from the Toarcian of Holzmaden, Germany
http://doc.rero.ch/r[...]
[72]
논문
The taxonomic and phylogenetic position of the Plesiosauroidea from the Lower Jurassic Posidonia Shale of south-west Germany
2007
[73]
논문
Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses
[74]
논문
A new leptocleidid (Sauropterygia, Plesiosauria) from the Vectis Formation (Early Barremian–early Aptian; Early Cretaceous) of the Isle of Wight and the evolution of Leptocleididae, a controversial clade
2013
[75]
논문
Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition
2014
[76]
논문
A new elasmosaurid plesiosaur from the Lower Jurassic of southern France
http://doc.rero.ch/r[...]
1999
[77]
논문
The elasmosaurid plesiosaur ''Aristonectes'' Cabreta from the Latest Cretaceous of South America and Antarctica
[78]
서적
A phylogeny of Plesiosauria (Sauropterygia) and its bearing on the systematic status of ''Leptocleidus'' Andrews, 1922
http://mapress.com/z[...]
Zootaxa
2017-12-09
[79]
논문
A new elasmosaurid (Sauropterygia, Plesiosauria) from the Bearpaw Shale (Late Cretaceous, Maastrichtian) of Montana demonstrates multiple evolutionary reductions of neck length within Elasmosauridae
https://ora.ox.ac.uk[...]
2017
[80]
논문
Elasmosaurid phylogeny and paleobiogeography, with a reappraisal of ''Aphrosaurus furlongi'' from the Maastrichtian of the Moreno Formation
https://figshare.com[...]
[81]
논문
Bone histology of aquatic reptiles: what does it tell us about secondary adaptation to an aquatic life?
2013-01-01
[82]
논문
Computer Simulations Imply Forelimb-Dominated Underwater Flight in Plesiosaurs
[83]
논문
The four-flipper swimming method of plesiosaurs enabled efficient and effective locomotion
[84]
논문
Viviparity and K-selected life history in a Mesozoic marine plesiosaur (Reptilia, Sauropterygia)
https://www.research[...]
[85]
논문
Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, ''Cretoxyrhina mantelli''
https://www.research[...]
[86]
논문
An elasmosaur bitten by a pliosaur
https://www.research[...]
[87]
논문
An integrated approach to understanding the role of the long neck in plesiosaurs
https://www.app.pan.[...]
2017-07-24
[88]
논문
Functional morphology and hydrodynamics of plesiosaur necks: Does size matter?
http://researchonlin[...]
2019
[89]
논문
Large size in aquatic tetrapods compensates for high drag caused by extreme body proportions
[90]
논문
Elasmosaur (Reptilia: Sauropterygia) neck flexibility: Implications for feeding strategies
http://doc.rero.ch/r[...]
[91]
서적
Ancient Sea Reptiles: Plesiosaurs, Ichthyosaurs, Mosasaurs, and More
Smithsonian Books
2022
[92]
논문
An elasmosaur with stomach contents and gastroliths from the Pierre Shale (Late Cretaceous) of Kansas
[93]
논문
A review of gastrolith function with implications for fossil vertebrates and a revised classification
http://www.dinosaurh[...]
2017-12-29
[94]
서적
The Changing Earth: Exploring Geology and Evolution
https://archive.org/[...]
Brooks/Cole, Cengage Learning
[95]
서적
Earth System History
W.H. Freeman and Company
[96]
서적
High-resolution Approaches in Stratigraphic Paleontology
Kluwer Academic Publishers
[97]
PBDB
Elasmosaurus
https://paleobiodb.o[...]
[98]
논문
Why sauropods had long necks; and why giraffes have short necks
https://www.ncbi.nlm[...]
2013-02-12
[99]
논문
A Small Body Sized Non-Aristonectine Elasmosaurid (Sauropterygia, Plesiosauria) from the Late Cretaceous of Patagonia with Comments on the Relationships of the Patagonian and Antarctic Elasmosaurids
http://sedici.unlp.e[...]
2016-06-01
[100]
서적
絶滅したふしぎな巨大生物
PHP研究所
2011-06-10
[101]
문서
A - かつて示された白鳥のように首をもたげる姿勢
B - 正面に首を伸ばした姿勢
C - 底生の獲物を捕食するときの姿勢
D - 水平方向に大きく首を曲げた姿勢
E - 首を蛇行させた姿勢
[102]
논문
Elasmosaur (Reptilia: Sauropterygia) neck flexibility: Implications for feeding strategies
https://www.scienced[...]
2008-06-01
[103]
웹사이트
PALAEONTOLOGY online {{!}} Article: Fossil Focus > Fossil Focus: Elasmosaurs
https://www.palaeont[...]
2015-02-01
[104]
논문
An integrated approach to understanding the role of the long neck in plesiosaurs
https://www.app.pan.[...]
2017-07-24
[105]
논문
Functional morphology and hydrodynamics of plesiosaur necks: does size matter?
https://www.tandfonl[...]
2019-03-04
[106]
논문
Large size in aquatic tetrapods compensates for high drag caused by extreme body proportions
https://www.ncbi.nlm[...]
2022-04-28
[107]
논문
Synopsis of the extinct Batrachia, Reptilia and Aves of North America, Part I
https://www.biodiver[...]
2017-11-08
[108]
논문
An elasmosaur with stomach contents and gastroliths from the Pierre Shale (Late Cretaceous) of Kansas
[109]
논문
Postscript to Everhart, M.J. 2005. "Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of ''Elasmosaurus platyurus'' Cope 1868?" – PalArch's Journal of Vertebrate Palaeontology 4, 3: 19–32
http://www.palarch.n[...]
2017-11-13
[110]
문서
2011年、1987年に発掘された首長竜の一種である[[ポリコティルス]]の化石の体内に[[胎児]]の骨格が残っていることがアメリカの研究チームによって確認された。このことから、首長竜が胎生(卵胎生)であったことは間違いないと見られている。
[111]
서적
The Princeton Field Guide to Mesozoic Sea Reptiles
Princeton University Press
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com