옴팔로사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
옴팔로사우루스는 중생대 트라이아스기에 살았던 멸종된 해양 파충류의 한 속으로, 독특한 치아 구조가 특징이다. 단추 모양의 치아는 오렌지 껍질 질감의 에나멜 표면을 가지며, 턱 구조는 지속적인 마모에 적합하게 진화했다. 옴팔로사우루스는 경두성 동물로 추정되며, 암모나이트나 이매패류를 먹이로 삼았을 가능성이 있다. 1902년 미국 네바다에서 화석이 처음 발견되었으며, 현재까지 여러 종이 보고되었다.
더 읽어볼만한 페이지
- 미국의 화석 - 트리케라톱스
트리케라톱스는 북아메리카에서 발견된 각룡류 공룡의 한 속으로, 거대한 두개골과 뿔이 특징이며 백악기 말기에 번성했고, 현재는 두 종으로 분류된다. - 미국의 화석 - 티라노사우루스
티라노사우루스는 백악기 후기 북아메리카에 서식했던 몸길이 15m에 달하는 거대한 육식 공룡으로, 강력한 턱힘과 짧은 앞다리, 긴 꼬리를 가진 최상위 포식자였으며, '수'와 '스카티' 등의 표본과 대중문화 속 인기 덕분에 그 생태와 행동에 대한 연구가 지속되고 있다. - 어룡 - 운도로사우루스
운도로사우루스는 후기 쥐라기 시대에 살았던 해양 파충류로, 불완전하게 융합된 좌골치골과 강한 치열을 가지고 있으며, 삼각형 이빨로 해양 생물을 잡아먹는 육식성 포식자로 추정되지만, 계통 발생적 위치는 아직 논쟁 중이다. - 어룡 - 겐가사우루스
쥐라기에 이탈리아 겐가 지역에서 서식한 겐가사우루스는 약 2.4m의 몸길이를 가진 오프탈모사우루스과의 어룡으로, 비교적 완전한 화석 골격 특징으로 오프탈모사우루스와 구별된다.
| 옴팔로사우루스 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Omphalosaurus |
| 명명자 | Merriam, 1906 |
| 속 | Omphalosaurus |
| 시대 | 전기 삼첩기 - 중기 삼첩기, 2억 5200만 년 전 ~ 2억 3700만 년 전 |
| 종 | |
| O. nevadanus | Merriam, 1906 (모식종) |
| O. merriami | Maisch, 2010 |
| O. nettarhynchus | Mazin & Bucher, 1987 |
| O. peyeri | Maisch & Lehmann, 2002 |
| O. wolfi | Tichy, 1995 |
2. 특징
옴팔로사우루스(그리스어로 '단추 도마뱀'이라는 뜻)는 트라이아스기 초기부터 중기까지 살았던 해양 파충류의 한 속으로, 어룡의 일종으로 여겨진다.[2] 이름처럼 독특한 치아 구조가 가장 큰 특징이며, 이는 다른 어룡들과 구별되는 중요한 특징이다. 현재까지 알려진 옴팔로사우루스에 대한 정보는 주로 여러 턱 조각과 일부 갈비뼈, 척추뼈 화석에 기반한다. 이 화석들은 미국 서부, 독일, 오스트리아, 그리고 노르웨이 북부 해안의 스피츠베르겐 섬 등지에서 발견되었다.
옴팔로사우루스는 비교적 크고 통통한 체형을 가졌을 것으로 추정되며,[2] 고도로 특화된 치아 구조를 가지고 있었다. (자세한 내용은 #치아 구조 참조) 턱과 치아 외의 골격 구조에 대해서는 알려진 정보가 상대적으로 적지만, 발견된 화석을 통해 일부 특징을 알 수 있다. (자세한 내용은 #골격 구조 참조)
2. 1. 치아 구조
옴팔로사우루스는 다른 어룡에 비해 매우 특화된 치아 구조로 가장 잘 알려져 있다.[2] 치아는 단추 모양이며 측면에서 보면 돔 형태를 하고 있다. 거의 원형의 치관을 가지며,[4] 불규칙한 에나멜 표면은 오렌지 껍질의 질감과 비슷하다.[5] 개별 치아의 직경은 12mm를 넘지 않으며,[5] 전상악골과 치골에 있는 치판에만 배열되어 서로 90º 각도를 이룬다. ''O. nevadanus''의 잘 보존된 매끄러운 구개골을 근거로 볼 때, ''옴팔로사우루스''가 판치류와 유사한 구개 치아를 가졌을 가능성은 낮다.[2]''옴팔로사우루스''의 치아는 수백 개에 이를 수 있으며 두개골 정중선을 따라 집중되어 있다.[6] 각 종마다 치아 배열 정도가 다르지만, ''O. nevadanus''는 약간의 불균일함에도 불구하고 가장 깔끔하게 정돈된 치아 배열(뚜렷한 열 형태)을 보여준다. 다른 종의 치아는 대부분 교합면에서 불규칙한 패턴으로 배열되어 있다.[7] 상부 치판은 볼록한 표면을 형성하고 하부 치판은 오목하다.
과거에는 턱이 짧고 넓으며 강력한 무는 힘을 가졌다고 여겨졌으나, 최근의 복원에 따르면 치골 결합부가 길고 약 15º 각도로 연결되어 턱이 긴 "V"자 모양을 이룬다.[2] 복원된 ''O. nevadanus''의 아래턱 길이는 50cm를 넘을 수도 있다.[6]
턱 조각 연구를 통해 ''옴팔로사우루스''가 지속적인 마모에 적합한 치열을 가졌으며, 치아 교체율이 높았다는 사실이 밝혀졌다.[2][8] ''옴팔로사우루스''는 미성숙한 교체 치아와 성숙한 치아의 에나멜 미세 구조가 다르다는 점에서 특이하다. 다른 어룡처럼 성숙한 치아는 미세 단위 에나멜 구조를 가지지만, 교체 치아는 기둥형 에나멜 구조를 가진다. 이러한 구조적 변화가 어떻게 일어나는지는 아직 밝혀지지 않았다.[5]
2. 2. 골격 구조
''옴팔로사우루스''는 비교적 크고 통통한 해양 파충류로,[2] 길이는 5m이며 무게는 783kg 이상이다.[3]치아 구조 외에는 ''옴팔로사우루스''에 대해 알려진 바가 비교적 적으며, ''O. wolfi''에 속하는 소수의 갈비뼈와 척추 앞쪽의 척추뼈만이 알려져 있다.[9] 갈비뼈는 부풀어 있고 속이 비어 있는데,[7][9] 이는 물로 돌아가는 양막류에서 흔히 볼 수 있는 특징이다.[7] 척추뼈는 깊은 양쪽 오목 형태를 보인다.[7] 또한, ''옴팔로사우루스''는 척추뼈 몸통 위에 신경궁을 잃었다.[2] 뼈는 섬유상 골 조직으로 짜여져 있어 뼈의 성장 속도가 빨랐음을 나타낸다.[10]
3. 고생물학
옴팔로사우루스는 중생대 트라이아스기 초기부터 중기(약 2억 4500만 년 전 ~ 2억 년 전)에 살았던 파충류이다. 주로 현재의 북아메리카와 유럽을 중심으로 한 북부 대서양 지역에 서식했던 것으로 보인다.
최초의 화석은 1902년 미국 네바다에서 V. C. 오스몬트(Osmont)에 의해 발견되었고, 1906년 존 C. 메리암(John C. Merriam)이 처음으로 학계에 보고했다. 메리암은 이 화석을 당시 알려진 어룡(Ichthyosaur)으로 분류하지 않고, 오히려 플라코돈트(Placodont)나 린코사우루스(Rhynchosaur)와 유사할 것으로 추정했다.
1934년 쿤(Kuhn)이 처음 옴팔로사우루스를 어룡으로 분류했고, 1983년 마진(Mazin)이 이를 지지했다. 그러나 1997년과 2000년에 모타니(Motani)는 옴팔로사우루스가 어익형류(Ichthyopterygia)의 기본적인 특징이 부족하다며 어룡 분류에 반대하고, 기룡상목(Sauropterygia)과의 연관성을 제기했다. 이후 2010년 마이스치(Maisch)가 새로운 종을 기술하면서 다시 어룡과의 관계를 주장하며 분류에 대한 논쟁이 이어졌다.
현재 옴팔로사우루스는 중소형 어룡으로 간주된다. 다른 어룡들처럼 뚜렷한 횡돌기(transverse process)가 없는 원반형 척추뼈를 가지며, 척추 중심부(centrum)는 너비보다 길이가 짧다. 갈비뼈는 어룡이나 샤스타사우루스과(Shastasauridae)와 유사한 등쪽-배쪽(dorsoventral) 관절 형태를 보인다. 성숙한 개체 치아의 미세 에나멜 구조(microlamellar enamel)는 다른 파충류에서는 드물지만 어룡의 특징적인 형질이며, 이는 옴팔로사우루스를 어룡으로 분류하는 주요 근거 중 하나이다.
하지만 옴팔로사우루스의 독특한 치아 배열은 분류 논쟁의 핵심이 되었다. 다른 어룡들이 뚜렷한 열을 이루는 것과 달리, 옴팔로사우루스의 이빨은 불규칙하게 배열되어 마치 포장도로와 같은 형태를 이룬다. 이는 하위 섹션 '식성'에서 더 자세히 다루어진다. 또한, 속이 빈 갈비뼈와 독특하게 휘어진 상완골 등 다른 어룡에게서는 발견되지 않는 특징들 때문에 옴팔로사우루스의 정확한 분류학적 위치에 대한 연구가 계속되고 있다.
3. 1. 식성
''옴팔로사우루스''의 고도로 특화된 치아 구조는 이들이 경두성(딱딱한 껍질을 가진 먹이를 먹는) 동물이었음을 시사한다.[7] 이빨은 단추처럼 생겼고 옆에서 보면 돔 모양이며, 오렌지 껍질 질감과 유사한 에나멜 표면을 가진 거의 원형의 크라운(치관) 형태였다. 개별 치아의 직경은 12mm를 넘지 않았으며, 수백 개에 이를 수 있는 많은 이빨이 두개골 중간선을 따라 빽빽하게 배열되어 있었다. 이빨은 심한 마모에 적합하게 발달했으며, 컴퓨터 단층 촬영(CT) 스캔 결과 단단한 먹이를 처리하기 위해 이빨이 매우 빠르게 교체되었음을 보여준다.[7]하지만 먹이를 붙잡는 데 사용되는 앞니와 같은 이빨은 부족했다. 또한, 초기 연구와 달리 최근의 복원에 따르면 턱은 길고 좁은 'V'자 형태였으며, 이는 껍데기를 부수는 데 필요한 강력한 턱 힘을 가진 다른 경두성 동물들의 짧고 튼튼한 턱과는 다른 모습이다.[7]
이렇게 마모가 심한 이빨과 상대적으로 약한 턱 힘의 조합은 초식 동물이나 조각류 공룡과 유사한 특징이다. 조각류처럼 ''옴팔로사우루스''도 이빨 교체율이 높고 이차적인 교합면(맞물리는 면)이 발달했지만, 중기 트라이아스기 바다에는 섬유질 해양 식물이 거의 없었기 때문에 초식 동물이었을 가능성은 낮다.[7]
따라서 당시 흔했던 암모나이트나 껍질이 단단하지만 비교적 얇았던 의사-플랑크톤성 할로비아 이매패류 등이 주요 먹이였을 것으로 추정된다.[7] 샌더(Sander)와 파버(Faber)는 ''옴팔로사우루스''가 살집 있는 뺨을 이용해 물과 함께 먹이를 입안으로 빨아들인 후(흡입 섭식), 수많은 단추 모양 이빨로 껍질을 부수거나 갈아서 먹었을 것이라는 가설을 제시했다.[2] 이는 먹이를 잡는 이빨이 부족한 점을 보완하는 섭식 방식이었을 수 있다. 최근 연구는 이들이 플라코돈트가 선호했던 이매패류보다는 암모나이트를 주로 사냥했을 가능성을 더 높게 보고 있다.[11]
3. 2. 감압병
다른 초기 어룡과 마찬가지로, 옴팔로사우루스에서는 무혈성 괴사의 증거가 발견되지 않았다. 이는 옴팔로사우루스가 감압병에 걸리지 않았을 가능성이 높다는 것을 시사한다. 연구자들은 이러한 특징을 초기에서 중기 트라이아스기에 대형 수생 포식자가 부족했던 환경과 연관 짓는다. 즉, 옴팔로사우루스가 포식자를 피해 급하게 깊은 곳으로 잠수할 필요가 적었을 것이라는 추정이다. 게다가 초기 어룡들은 일반적으로 물속에서 천천히 위아래로 움직였거나, 급격한 수압 변화에 대응할 수 있는 생리적인 보호 능력을 갖추었을 가능성도 제기된다.4. 발견 및 분류
1902년 미국 네바다에서 V. C. 오스몬트에 의해 최초의 ''옴팔로사우루스'' 화석이 발견되었으며, 1906년 존 C. 메리엄에 의해 처음으로 기술되었다.[13] 메리엄은 이 화석(''O. nevadanus'')을 어룡으로 분류하지 않고, 대신 플라코돈트나 린코사우루스와의 유사성을 제시했다.[4] 이후 1934년 쿤(Kuhn)이 처음으로 ''옴팔로사우루스''를 어룡으로 분류했으며, 1983년 마진(Mazin)이 이러한 분류를 지지하는 근거를 제시했다. 그러나 1997년과 2000년에 모타니(Motani)는 어룡의 주요 공유 파생 형질(synapomorphy)이 부족하다는 점을 지적하며 이 분류에 반대하고, 사우롭테리기아와의 연관성을 주장했다.[2][7][8] 하지만 2010년 마이쉬(Maisch)는 새로운 종을 기술하면서 ''옴팔로사우루스''가 어룡에 속한다는 분류를 다시 한번 확인했다.[14]
4. 1. 어룡 내 분류 특징
''옴팔로사우루스''는 현재 소형에서 중형 크기의 어룡으로 여겨진다. 다른 어룡과 마찬가지로, 옴팔로사우루스는 뚜렷한 가로 돌기(transverse processeseng)가 없는 깊은 양쪽 오목 척추(amphicoelous vertebraeeng)를 가지며, 척추 중심부(centrumeng)는 너비보다 짧다. ''옴팔로사우루스''의 갈비뼈는 어룡 및 샤스타사우루스과와 등쪽-배쪽(dorsoventraleng) 관절을 공유한다.[7] 또한, ''O. wolfi'' 종은 해면질 뼈(cancellous bone) 구조를 가지고 있는데, 이는 어룡과 동일하지만 다른 여러 수생 동물에서도 흔하게 나타나는 특징이다.[7] ''옴팔로사우루스''를 어룡으로 분류하는 가장 뚜렷한 근거 중 하나는 성숙한 치아에서 발견되는 미세 단위 법랑질(microunit enamel) 구조이다. 이는 어룡이 공통으로 가지는 특징으로 알려져 있으며, 다른 파충류에서는 드물게 나타난다.[9]하지만 ''옴팔로사우루스''의 분류에는 몇 가지 논란이 있는데, 이는 다른 어룡들과 구별되는 독특한 특징들 때문이다. 가장 두드러진 차이점은 치아의 배열 방식으로, 다른 어룡들이 뚜렷한 열을 이루어 치아가 배열되는 것과 달리, ''옴팔로사우루스''의 치아는 불규칙한 포장도로처럼 배열되어 있다. 또한, 다른 어룡에서는 발견되지 않는, 서로 직각을 이루는 상악(윗턱) 연삭면(grinding surfaceeng)을 가지고 있다. ''옴팔로사우루스''의 치아 크라운(crowneng)은 단단한 먹이를 먹는 다른 어룡들보다 낮고 불규칙하며, 법랑질 표면은 어룡의 전형적인 특징인 세로 주름 대신 오렌지 껍질 같은 질감을 보인다. 이 외에도 ''옴팔로사우루스''는 다른 어룡에서는 발견되지 않는 속이 빈 갈비뼈와, 위팔뼈(humerus)에 두드러진 삼각근 융기(deltopectoral cresteng)를 가지고 있다는 점도 독특한 특징이다.[7]
5. 종
옴팔로사우루스 속에는 여러 화석 종이 포함된다. 이들 종은 주로 발견된 지역의 지층과 트라이아스기 내에서의 시기, 그리고 턱과 치아의 형태 등 해부학적 특징을 바탕으로 분류된다. 일부 종은 분류학적 위치에 대해 논쟁이 있기도 하다. 각 종에 대한 자세한 설명은 하위 문서를 참고한다.
5. 1. 유효종
- ''옴팔로사우루스 네바다누스''(Omphalosaurus nevadanusla)는 이 속의 모식종이며, 미국 네바다 주 훔볼트 산맥의 해양 프리다 지층에서 발견된 두 종 중 하나이다. ''O. nevadanus''는 중기 트라이아스기 아니시안 지층에서 유래되었으며, 1906년 메리엄(Merriam)에 의해 처음 기술되었다.[7] 화석은 전방 경추와 하악의 일부를 가진 두개골의 아랫부분으로 구성되어 있으며, 관절 부위가 유실되고 앞쪽의 결합 부위 앞에서 부러져 있다. 각골, 상각골, 치골 및 설골이 모두 하악에서 보인다.[13] 다른 종과 달리 치골 결합 부위의 후방 가장자리가 둥글다. ''O. nevadanus''는 ''O. nettarhynchus''보다 훨씬 크고 이빨 수도 더 많다.[7]
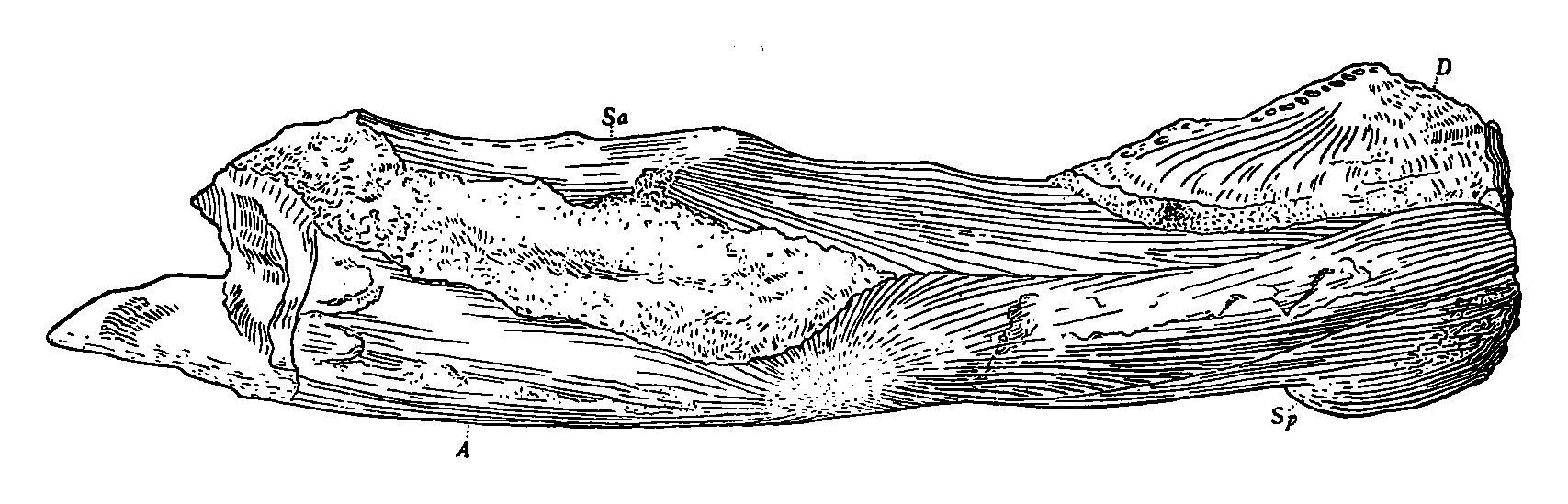
- ''옴팔로사우루스 네타린쿠스''(Omphalosaurus nettarhynchusla)는 프리다 지층에서 발견된 두 번째 종으로, 중기 트라이아스기 스파시안 지층에서 유래되었다. 1987년 마진(Mazin)과 부처(Bucher)에 의해 처음 기술되었다. 화석은 단편적인 하악만으로 구성되어 있지만, ''O. nettarhynchus''는 작은 크기, 상대적으로 적지만 큰 이빨, 그리고 측면으로 확장된 턱 결합 부위로 인해 다른 종과 구별된다.[7]
- ''옴팔로사우루스 메리아미''(Omphalosaurus merriamila)는 스발바르 제도에 있는 해양 스티키 키프(Sticky Keep) 지층에서 유래했으며 2010년 마이슈(Maisch)에 의해 기술되었다. 트라이아스기 하류 시대에 살았다. 이 화석은 원래 페소프테릭스 니세리(''Pessopteryx nisseri'')의 일부라고 생각되었던 턱 파편들로 이루어져 있는데, 나중에 턱이 옴팔로사우루스의 것임이 밝혀졌다. 턱 파편에는 부드러운 에나멜과 플리시덴틴을 함유한 치아뿌리가 3줄로 나 있다.
- ''옴팔로사우루스 sp.''(Omphalosaurus sp.la)의 왼쪽 상완골이 중기 트라이아스기의 독일 프랑코니아 무쉘칼크 지층, 특히 가른베르크(Garnberg) 채석장의 호엔로에 지층에서 발견되었다. 후기 라디니안에서 유래되었으며 1984년 하그도른(Hagdorn)에 의해 발견되었고, 1998년 산더(Sander)와 파버(Faber)에 의해 기술되었다. 상완골은 전형적인 어룡류의 해면골 구조를 나타냈지만, 특정 종은 아직 불분명하다.[7]
5. 2. 경합종
- ''Omphalosaurus wolfi'': 중기 트라이아스기 라디니아 초기 종으로, 북부 알프스의 뒤른베르크 산, 레르체크 석회암에서 발견되었다.[7] 1991년 G. 볼프가 발견하고 1995년 티히가 기술하였다.[15] 표본은 여러 개의 척추 앞부분, 갈비뼈, 부분적으로 연결된 하악을 가진 분리된 두개골 등으로 구성된다.[7] 치아는 ''O. nisseri''와 유사하며, 하악 요소는 ''O. nevadanus''와 비슷하다.[7] 2003년 잔더와 파버는 이 종이 실제로는 ''O. cf. nevadanus''일 수 있다고 제안했지만, 마이시는 두개골 물질에 대한 재조사가 필요하며 그때까지는 독립된 종으로 유지되어야 한다고 주장하여 분류에 논란이 있다.[14]
- ''Omphalosaurus peyeri'': 중기 트라이아스기 아니시안 시대의 종이다. 독일 뢰더스도르프의 샤움칼크 층에서 발견되었는데, 이는 얕은 연안 환경에 서식했을 가능성을 시사한다.[16] 화석은 왼쪽 위턱의 일부로, 3개의 성숙한 버섯 모양 치아와 소수의 교체 치아를 가지고 있다. 다른 ''Omphalosaurus'' 종과 달리 기능적인 치아 줄이 하나뿐이고 교체 치아도 적었다.[16] 이 때문에 처음에는 판상치류로 여겨졌으나, 2002년 마이시와 레만은 특징적인 치아 모양과 에나멜 질감을 근거로 ''Omphalosaurus''로 기술하였다.[17] 가장 원시적인 ''Omphalosaurus'' 종으로 생각되기도 하지만,[16] 윈트리히와 잔더는 판상치류와 유사한 원주형 에나멜 구조를 들어 이 분류에 의문을 제기하였다.[5][11]
- ''Omphalosaurus merriami'': 초기 트라이아스기에 살았던 종으로, 스발바르의 스티키킵 층에서 발견되었으며 2010년 마이시가 기술하였다.[14] 화석은 턱 조각으로 구성되어 있으며, 처음에는 함께 발견된 어룡 ''Pessopteryx nisseri''의 일부로 생각되었으나 나중에 옴팔로사우루스의 특징을 가진 것으로 밝혀졌다. 턱 조각에는 세 줄의 치아가 있다.[14] 그러나 2013년 에린 맥스웰과 벤자민 키어는 이 표본에서 종을 구별할 만한 뚜렷한 특징을 찾기 어렵고 보존 상태도 좋지 않다고 판단하여, ''O. merriami''를 ''nomen dubium'' (의문명)으로 간주하였다.[18]
5. 3. 수정종
페소프테릭스 니세리(Pessopteryx nisseri)는 1910년 비만(Wiman)이 스피츠베르겐에서 발견하고 기술한 여러 화석들을 포함한다. 이 중에는 현재 O. 메리암미(O. merriami)로 분류되는 턱 파편도 포함되어 있었다.[14] 처음에는 비만(Wiman)과 마진(Mazin)에 의해 O. 니세리(O. nisseri)로 여겨졌으나, 이후 지느러미와 가슴띠 화석이 어룡의 특성을 보이는 것으로 밝혀져 현재는 별도의 종으로 인정받고 있다. 이에 따라 페소프테릭스(Pessopteryx)는 고유의 속으로 인정받고 있다.[19]참조
[1]
논문
Preliminary report on ichthyopterygian elements from the Early Triassic (Spathian) of Spitsbergen
https://www.research[...]
2018-10-04
[2]
논문
The Triassic marine reptile ''Omphalosaurus'': osteology, jaw anatomy, and evidence for ichthyosaurian affinities
2003-12-24
[3]
논문
Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans
[4]
논문
Notes on the Dentition of ''Omphalosaurus''
[5]
서적
The microstructure of reptilian tooth enamel : terminology, function, and phylogeny
F. Pfeil
1999-01-01
[6]
논문
Early Triassic Marine Biotic Recovery: The Predators' Perspective
2014-03-19
[7]
논문
New finds ofOmphalosaurus and a review of Triassic ichthyosaur paleobiogeography
[8]
논문
Is ''Omphalosaurus'' Ichthyopterygian?: A Phylogenetic Perspective
2000-01-01
[9]
논문
Ichthyosauria: their diversity, distribution, and phylogeny
[10]
서적
Mechanics and physiology of animal swimming
Cambridge University Press
1994-01-01
[11]
논문
Two Records of ''Omphalosaurus'' from the Muschelkalk - The Reconstruction of Triassic Marine Ecosystems Based on Teeth
https://www.research[...]
[12]
논문
Adaptations for marine habitat and the effect of Triassic and Jurassic predator pressure on development of decompression syndrome in ichthyosaurs
2012-05-10
[13]
논문
Preliminary note on a new marine reptile from the middle triassic of Nevada
[14]
논문
Phylogeny, systematics, and origin of the Ichthyosauria-the state of the art
[15]
논문
Ein früher, durophager Ichthyosaurier (Omphalosauridae) aus der Mitteltrias der Alpen
[16]
논문
A new basal omphalosaurid from the Middle Triassic of Germany
[17]
웹사이트
Marine reptile – GSUB
http://www.geosammlu[...]
2017-03-04
[18]
논문
Triassic ichthyopterygian assemblages of the Svalbard archipelago: a reassessment of taxonomy and distribution
https://www.research[...]
[19]
논문
A New Look at Ichthyosaur Long Bone Microanatomy and Histology: Implications for Their Adaptation to an Aquatic Life
2014-04-21
[20]
논문
The Triassic marine reptile Omphalosaurus: osteology, jaw anatomy, and evidence for ichthyosaurian affinities
https://doi.org/10.1[...]
2003-12-24
[21]
논문
Is Omphalosaurus ichthyopterygian?—A phylogenetic perspective
https://doi.org/10.1[...]
2000-06-27
[22]
논문
A globally distributed durophagous marine reptile clade supports the rapid recovery of pelagic ecosystems after the Permo-Triassic mass extinction
https://www.nature.c[...]
2022-11-14
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com