육기어류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
육기어류는 살집이 있고 엽상 지느러미를 가지며, 사지동물의 다리와 유사한 관절을 가진 어류의 한 분류군이다. 육기어류는 크게 실러캔스류, 폐어류, 사지형류로 나뉘며, 실러캔스류는 현존하는 종이 있으며, 폐어류와 사지형류는 멸종된 종이 많다. 육기어류는 부레와 골화된 내골격을 진화시켰으며, 사르코프테리기의 비늘에 코스민 층이 존재하는 특징을 보인다. 육기어류의 진화는 데본기 시대에 시작되었으며, 사지동물로의 진화에 중요한 역할을 했다. 육기어류의 육상 진출에 대한 가설로는 물웅덩이 축소 가설, 조간대 적응 가설, 삼림 늪지 적응 가설 등이 있다.
육기어류는 살덩이 형태의 지느러미를 가진 경골어류의 한 분류군으로[9][13][14], 현생종은 크게 실러캔스류(총기어강 또는 복어류), 폐어류(폐어강 또는 폐어형류), 그리고 사지동물을 포함하는 사지형류로 나뉜다.
2. 특징

초기 육기어류는 경골어류의 일종으로, 살집이 있고 잎 모양(엽상)의 짝지느러미를 가진다. 이 지느러미는 하나의 뼈를 통해 몸통과 연결된다. 육기어류의 지느러미는 다른 모든 어류의 지느러미와 구별되는 특징을 보이는데, 각각 몸에서 뻗어 나온 살집이 있고 엽상이며 비늘로 덮인 줄기 위에 지느러미가 달려 있다는 점이다. 이러한 구조는 사지동물의 팔다리 원기인 사지 원기와 유사하다.
육기어류의 비늘은 진정한 골편(bony scale)으로, 판상골(lamellar bone) 위에 혈관이 분포하는 뼈층, 상아질과 유사한 코스민(cosmine) 층, 그리고 가장 바깥쪽에는 케라틴 층이 덮여 있는 복잡한 구조를 가진다.
조기어류가 보통 하나의 등지느러미를 가지는 것과 달리, 육기어류는 기저부가 분리된 두 개의 등지느러미를 가진다는 특징이 있다. 또한, 육기어류의 두개골은 원시적으로 머리뼈 중간에 움직일 수 있는 경첩선(hinge line)을 가지고 있었으나, 이 특징은 폐어와 사지동물로 진화하면서 사라졌다. 모든 육기어류는 진짜 법랑질로 덮인 이빨을 가진다. 초기 육기어류는 일반적으로 대칭적인 꼬리를 가지고 있었다.
실러캔스를 제외한 총기류는 내비공(choana, 코안)을 가지고 있기 때문에, 이들을 묶어 내비공류(Choanichthyes)라고도 부른다.[12] 그러나 총기류의 내비공이 오스테올레피스류나 사지동물의 내비공과 진정한 상동 기관인지에 대해서는 논란이 있다.[10][11]
특히 사지동물과 가까운 관계에 있는 어류인 사지형류(tetrapodomorph)의 신체 구조는 동물이 물에서 뭍으로 서식지를 옮기는 진화 과정을 이해하는 데 중요한 단서를 제공한다. 이들의 앞지느러미와 뒤지느러미는 사지동물의 팔다리와 유사한 관절 구조를 가지며, 최초의 사지동물인 초기 양서류는 이러한 지느러미에서 진화한 다리를 가졌다.
3. 분류
사지동물은 육기어류에서 진화했기 때문에[10], 사지동물을 제외하면 육기어류는 측계통군이 된다. 이 때문에 일부 분류 체계에서는 사지동물을 육기어류의 하위 분류군으로 포함시켜 단계통군으로 만들기도 한다[10][11].
과거에는 실러캔스목과 멸종한 오스테올레피스목 등을 묶어 총기류(Crossopterygii)로 분류했으나, 현재는 실러캔스목이 다른 육기어류와 분리되어 복어류(Actinistia)를 형성하고, 오스테올레피스목 등은 사지동물과 함께 사지형류(Tetrapodomorpha)를 구성하는 것으로 본다[10][11]. 또한, 멸종한 폴로레피스목(Porolepiformes)은 폐어류(Dipnoi)와 함께 폐어형류(Dipnomorpha)를 이루며[11], 폐어형류와 사지형류는 엽상지느러미류(Rhipidistia)라는 더 큰 그룹으로 묶인다[11][17].
3. 1. 현존하는 종
라티메리아속 (''Latimeria'')
* 서인도양실러캔스 (''Latimeria chalumnae'')
* 인도네시아실러캔스 (''Latimeria menadoensis'')
오스트레일리아폐어속 (''Neoceratodus'')
* 오스트레일리아폐어 (''Neoceratodus forsteri'')
남아메리카폐어속 (''Lepidosiren'')
* 남아메리카폐어 (''Lepidosiren paradoxa'')
아프리카폐어속 (''Protopterus'')
* 표범폐어 또는 프로톱테루스 에이티오피쿠스 (''Protopterus aethiopicus'')
* 서아프리카폐어 또는 프로톱테루스 안넥텐스 (''Protopterus annectens'')
* 동아프리카폐어 또는 프로톱테루스 암피비우스 (''Protopterus amphibius'')
* 반점아프리카폐어 또는 프로톱테루스 돌로이 (''Protopterus dolloi'')
3. 2. 계통 분류
육기어류의 계통 분류는 연구자에 따라 세부적인 부분에서 차이가 있으나[18][19], 주요 분류군과 그 관계는 다음과 같이 정리할 수 있다. 척추동물이 육기어류로부터 직접 진화했다는 점을 반영하여, 일부 분류 체계(예: Benton, 2004)에서는 상위 분류군인 사지동물을 폐어강에 포함시키기도 한다.총기어강 (Actinistia) 
총기어강은 육기어류의 한 아강으로, 대부분 화석으로만 알려져 있으며 현생종은 단 2종(서인도양 실러캔스, 인도네시아 실러캔스)만이 남아있다. 실러캔스가 이 강에 속한다. 폐어강 (Dipnoi) 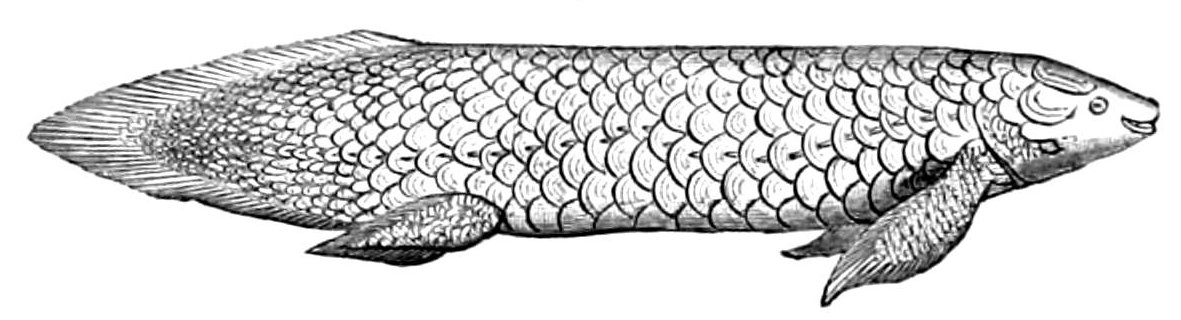
폐어강은 일반적으로 폐어라고 불리며, 민물고기의 한 아강이다. 폐어는 공기 호흡 능력과 같은 경골어류의 원시적 특징과 잘 발달된 내골격을 가진 엽상 지느러미와 같은 육기어류의 원시적 구조를 함께 가지고 있는 것으로 유명하다. 오늘날 폐어는 아프리카, 남아메리카, 오스트레일리아 대륙에만 분포하는데, 이는 중생대의 초대륙 곤드와나 시절의 분포를 반영하는 것으로 해석되기도 한다. 그러나 화석 기록을 보면 과거에는 전 세계적으로 널리 분포했으며, 현재의 분포는 판게아, 곤드와나, 로라시아 대륙 분열 이후 많은 계통이 멸종한 결과로 보인다. 사지형류 (Tetrapodomorpha) 
사지형류는 사지동물(네 발 달린 척추동물)과, 현생 폐어보다는 현생 사지동물과 더 가까운 관계에 있는 육기어류 친척들을 포함하는 분지군이다. 틱타알릭과 같이 물고기와 초기 사지동물 사이의 전이적 특징을 보이는 고등 사지형류는 발견자들에 의해 "피쉬어포드(fishapods)"라고 불리기도 했다. 사지형류에는 사지동물 계통군(현생 사지동물의 마지막 공통 조상과 그 모든 후손)과 여러 그룹의 초기 줄기 사지동물, 그리고 관련 육기어류 그룹(집합적으로 오스테올레피스목으로 알려짐)이 포함된다. 사지동물 계통군을 제외한 사지형류는 물고기에서 사지동물로의 진화 과정을 보여주는 측계통군이다. 사지형류를 정의하는 특징 중 하나는 지느러미 골격의 변화인데, 특히 볼록한 머리를 가진 상완골이 어깨 관절의 관절와와 연결되는 구조를 가진다. 사지형류 화석은 초기 데본기부터 발견되며, 오스테올레피스, 판데리크티스, 케니크티스, 퉁세니아 등이 포함된다.
계통 발생학적 연구(Janvier 외 1997, Mikko's Phylogeny Archive, Swartz 2012 등)에 따르면, 육기어강 내의 주요 분기 관계는 다음과 같이 나타낼 수 있다. 육기어강은 크게 †오니코돈티과(Onychodontidae)와 실러캔스목(Coelacanthiformes), 그리고 이들과 자매군을 이루는 육지어류(Rhipidistia)로 나뉜다. 육지어류는 다시 폐어형류(Dipnomorpha)와 사지형류(Tetrapodomorpha)로 구분된다. 폐어형류에는 †포롤레피스목(Porolepiformes)과 현생 폐어류(Dipnoi)가 포함된다. 사지형류는 매우 다양한 그룹을 포함하는데, †리조돈티과(Rhizodontidae), †오스테올레피스목(Osteolepiformes), 엘피스토스테갈리아(Elpistostegalia) 등을 거쳐 최종적으로 사지동물(Tetrapoda)로 이어진다. †''틱타알릭''과 같은 중요한 전이 화석은 엘피스토스테갈리아 내에 위치한다.
현생 종만을 고려한 분류 체계(야노 2006, 카이 2018 등 기반)[13][14][15]는 다음과 같다. 폐어아강과 사지동물아강을 묶어 폐어사지동물아강(Dipnotetrapodomorpha)으로 분류하기도 한다.[13][16] 또한, 폐어류의 두 목을 아목으로 취급하여 하나의 목(오스트레일리아폐어목, Ceratodontiformes)으로 통합하는 견해도 있다.[9][16]
현존하는 3개의 주요 계통 외에도 다양한 화석 분류군이 알려져 있다.[10] 과거에는 실러캔스목과 오스테올레피스목 등을 묶어 총기류(Crossopterygii)라고 불렀으나, 현재는 실러캔스목(총기어아강 Actinistia)이 다른 현생 육기어류 전체의 자매군이며, 오스테올레피스목, 리조두스형류(Rhizodontiformes), 엘피스토스테갈리아(Elpistostegalia, =판데릭티스목 Panderichthyida) 등은 사지동물과 함께 사지형류(Tetrapodomorpha)라는 단계통군을 형성하는 것으로 여겨진다.[10][11] 폴로레피스목(Porolepiformes)은 폐어류(Dipnoi)의 자매군으로 간주되어 함께 폐어형류(Dipnomorpha)를 구성한다.[11] 폐어형류와 사지형류를 합쳐 엽상지느러미류(Rhipidistia)라고 부른다.[11][17] †오니코두스목(Onychodontiformes)은 엽상지느러미류와 총기어아강보다 더 일찍 분기한 기저 그룹으로 생각되지만[11], 이들을 엽상지느러미류의 자매군으로 보고 함께 폐어사지동물아강에 포함시키는 설도 있다.[16]
다음은 계통 내 위치가 불분명한(''incertae sedis'') 일부 육기어류 분류군이다.

4. 진화
- 진흙투성이 얕은 곳에 적응한 ''판데리히티스'';
* 육지로 이동할 수 있는 지느러미 같은 다리를 가진 ''틱타알릭'';
* 잡초가 무성한 늪지대의 초기 사지동물들, 예를 들어:
여덟 개의 발가락을 가진 발을 가진 ''아칸토스테가'',
다리를 가진 ''이크티오스테가''.
후손에는 실러캔스 종과 같은 중층성 엽상 지느러미 어류도 포함되었다.]]

육기어류는 자매군인 조기어류와 함께 상강 경골어류를 구성한다. 이들은 연골어류와 달리 골화된 내골격과 폐와 공통 조상을 가진 부레를 가진 것이 특징이다. 육기어류의 가장 초기 화석은 약 4억 1800만 년 전, 고생대 실루리아기 최상층에서 발견되었다. 이 초기 육기어류는 당시 살았던 극어류와 매우 유사했다.
초기-중기 데본기(약 4억 1600만 년 전~3억 8500만 년 전) 동안, 일부 육기어류는 바다를 떠나 담수 환경으로 이동했다. 초기 데본기(약 4억 1600만 년 전~3억 9700만 년 전)에 육기어류는 크게 실러캔스와 리피디스티아류(Rhipidistia)의 두 주요 계통으로 나뉘었다.
실러캔스는 바다에 남아 진화했으며, 후기 데본기와 석탄기(약 3억 8500만 년 전~2억 9900만 년 전)에 가장 번성했다. 실러캔스류에 속하는 액티니스티아류(Actinistia)는 약 3억 8000만 년 동안 존재해 왔으며, 시간이 지나면서 47개 속에 121종이 확인되었다. 이들의 다양성은 트라이아스기 초기에 가장 크게 증가했다. 오늘날까지 살아남은 라티메리아 속의 실러캔스는 고대 육기어류의 원시적 특징을 많이 간직하고 있어 '살아있는 화석'으로 유명하다.
리피디스티아류는 담수 환경으로 이동하여 폐어와 사지형류라는 두 그룹으로 다시 나뉘었다. 이 두 그룹 모두 부레를 공기를 호흡할 수 있는 폐로 진화시켰다. 폐어는 트라이아스기에 가장 다양하게 진화했으며, 일부는 데본기 중기(약 3억 9700만 년 전~3억 8500만 년 전)에 물 밖에서도 어느 정도 생활할 수 있도록 적응했다. 오늘날에는 소수의 폐어만이 남아있다.
반면, 사지형류는 지느러미를 다리로 진화시켜 육상 생활에 적응해 나갔다. 이들은 데본기 후기(약 3억 8500만 년 전~3억 5900만 년 전)에 최초의 사지동물(네 발 달린 척추동물)로 진화했다.[3][4] 후기 데본기 멸종 사건 이후 살아남은 사지동물은 육상 환경에서 빠르게 적응 방산하여 석탄기와 페름기 동안 육상 생태계의 주요 구성원이 되었다. 사지동물이 아닌 육기어류는 고생대 말까지 계속 존재했지만, 페름기-트라이아스기 멸종 사건(약 2억 5100만 년 전) 동안 큰 타격을 입었다.
4. 1. 육상 진출 가설
폐어류의 짧은 지느러미(원시적인 다리)가 어떻게 진화했는지에 대해서는 크게 세 가지 주요 가설이 있다.1. 물웅덩이 축소 가설: 첫 번째이자 전통적인 설명은 미국의 고생물학자 알프레드 로머가 제시한 "물웅덩이 축소 가설" 또는 "사막 가설"이다. 그는 살던 물웅덩이가 말라붙자, 새로운 물을 찾아 이동해야 하는 필요성 때문에 다리와 폐가 진화했을 것이라고 보았다.
2. 조간대 적응 가설: 니에치비에츠키(Niedźwiedzki), 슈렉(Szrek), 나르키에비츠(Narkiewicz) 등은 2010년에 두 번째 가설인 "조간대 가설"을 제안했다. 이들은 약 3억 9500만 년 전의 것으로 추정되는 자헬미(Zachełmie) 발자국 화석을 근거로 들었다. 이 발자국 화석은 사지동물의 가장 오래된 증거 중 하나로, 총지느러미류가 내륙의 물웅덩이가 아닌 바닷가의 조간대에서 처음 육지로 올라왔을 가능성을 시사한다.
3. 삼림 늪지 적응 가설: 그레고리 레탈랙은 2011년에 세 번째 가설인 "삼림 가설"을 제시했다. 레탈랙은 다리가 나무뿌리나 식물이 우거진 얕은 물, 즉 삼림 지대의 늪과 같은 환경에서 이동하기 위한 수단으로 발달했을 것이라고 주장했다. 그는 이러한 주장의 근거로, 육상으로 진출하던 과도기적 사지동물 화석들이 주로 과거 습하고 나무가 많았던 충적 평야 지역에서 발견된다는 점을 들었다.
4. 육지로의 습관적 탈출 가설: 네 번째 가설은 소수 의견으로, 육지로 나옴으로써 얻는 이점 때문에 진화가 이루어졌다고 본다. 육지는 물속보다 포식자로부터 안전하고, 먹이 경쟁이 덜하며, 산소 농도 및 온도 조절 과 같은 특정 환경적 이점이 있었다는 것이다. 즉, 다리를 발달시킨 생물은 물 밖에서 시간을 보내는 생활에도 적응했다는 의미이다. 그러나 연구에 따르면, 총지느러미류는 실제로 육지로 진출하기 훨씬 이전에 이미 걷기에 적합한 사지동물과 유사한 다리를 발달시켰다. 이는 그들이 육지로 올라오기 전에 물속 바닥을 걸어 다니는 것에 먼저 적응했음을 시사한다.
참조
[1]
논문
The Silurian-Devonian boundary in East Yunnan (South China) and the minimum constraint for the lungfish-tetrapod split
https://www.research[...]
[2]
서적
Vertebrate Palaeontology
Blackwell
[3]
서적
When the Invasion of Land Failed: The Legacy of the Devonian Extinctions
https://books.google[...]
Columbia University Press
2013-11-12
[4]
웹사이트
Research project: The Mid-Palaeozoic biotic crisis: Setting the trajectory of Tetrapod evolution
http://www.southampt[...]
2014-05-31
[5]
논문
A megalichthyid sarcopterygian fish from the Lower Permian (Autunian) of the Saar-Nahe Basin, Germany
[6]
논문
The Silurian-Devonian boundary in East Yunnan (South China) and the minimum constraint for the lungfish-tetrapod split
https://www.research[...]
[7]
논문
Herpetichthyes, Amphibioidei, Choanichthyes or Sarcopterygii?
https://doi.org/10.1[...]
[8]
기타
[9]
논문
Phylogenetic classification of bony fishes
https://doi.org/10.1[...]
[10]
서적
バイオディバーシティ・シリーズ 7 脊椎動物の多様性と系統
裳華房
[11]
서적
バイオディバーシティ・シリーズ 7 脊椎動物の多様性と系統
裳華房
[12]
논문
脊椎動物における鼻器の進化と多様性
https://doi.org/10.1[...]
[13]
서적
脊椎動物の多様性と系統
[14]
서적
動物学の百科事典
丸善出版
[15]
서적
バイオディバーシティ・シリーズ 7 脊椎動物の多様性と系統
裳華房
[16]
서적
Fishes of the World
John Wiley & Sons
[17]
논문
A re-examination of sarcopterygian interrelationships, with special reference to the Porolepiformes
https://doi.org/10.1[...]
[18]
논문
Phylogenetic classification of bony fishes
https://bmcevolbiol.[...]
2017-07-06
[19]
기타
The tree of life and a new classification of bony fishes
http://currents.plos[...]
PLoS Currents Tree of Life
2013-04-18
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com