짚신벌레
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
짚신벌레는 17세기 후반에 현미경으로 관찰된 최초의 섬모충 중 하나로, 물에 사는 단세포 생물이다. 짚신벌레는 펠리클로 둘러싸인 타원형 또는 담배 모양의 세포를 가지며, 섬모를 사용하여 이동하고 세균, 조류, 효모 등을 섭취한다. 짚신벌레는 무성 생식과 유성 생식을 하며, 유성 생식인 접합을 통해 유전 물질을 교환한다. 짚신벌레는 클론 노화를 겪으며, 접합이나 자가수정을 통해 젊어질 수 있다. 짚신벌레는 다양한 종으로 분류되며, 학습 능력에 대한 연구도 진행되었다.
더 읽어볼만한 페이지
- 섬모충류 - 나팔벌레
나팔벌레는 전 세계 담수에서 발견되는 20종 이상의 원생생물 속으로, 뿔 모양 몸체와 섬모 고리가 특징이며, 일부 종은 녹조류와 공생하고 재생 능력이 뛰어나 블레파리스마속과 관련이 있다. - 섬모충류 - 빈막류
빈막류는 섬모충류의 한 그룹으로, 균일한 섬모 밀집과 작거나 퇴화된 구강 섬모를 가지며, 다양한 환경에서 자유 생활 또는 공생을 통해 작은 유기물을 섭취하는 미소포식성 생물이며, 사각구강류, 무구강류, 막구강류, 털말뚝류, 주모류, 각피섬모충류 등의 하위 분류군을 포함한다. - 원생생물 - 유공충
유공충은 단단한 껍질을 가진 원생생물로, 껍질 형태와 구성에 따라 분류되며, 해양 환경에서 생물 층서 및 고기후 연구, 석유 탐사 등 다양한 분야에 활용된다. - 원생생물 - 코아노조아
코아노조아는 칼라 구조를 특징으로 하는 깃편모충류와 동물을 포함하는 계통 분류군으로, 동물의 기원과 밀접한 관련이 있으며, 동물 진화 연구에 중요한 연결고리를 제공한다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 짚신벌레 - [생물]에 관한 문서 | |
|---|---|
| 특징 | |
| 분류 | 섬모충류 |
| 크기 | 약 0.2mm |
| 생태 | 민물 서식, 박테리아 등을 먹음 |
| 생식 | 세포 분열, 접합 |
| 형태 | |
| 모양 | 짚신 모양 |
| 섬모 | 전신에 섬모가 덮여 있음 |
| 세포 소기관 | 수축포 식포 대핵 소핵 |
| 생리학 | |
| 운동 | 섬모 운동으로 회전하며 이동 |
| 섭식 | 섬모 운동으로 음식물을 세포구로 모아 식포 형성 |
| 배설 | 수축포를 통해 삼투압 조절 및 노폐물 배출 |
| 호흡 | 세포막을 통한 기체 교환 |
| 유전 | |
| 유전 물질 | 대핵과 소핵에 유전 물질 존재 |
| 접합 | 두 개체가 일시적으로 결합하여 유전 물질 교환 |
| 연구 | |
| 중요성 | 세포 생물학 연구에 중요한 모델 생물 |
| 활용 | 세포 구조 및 기능 연구, 유전 연구 등에 활용 |
| 학명 | |
| 속명 | Paramecium |
| 명명자 | Müller, 1773 |
| 이명 | Paramoecium Hermann, 1783 Paramœcium Hermann, 1783 Chloroparamecium Fokin, Przybos, Chivilev, Beier, Horn, Skotarczak, Wodecka & Fujishima, 2004 Viridoparamecium Kreutz, Stoeck & Foissner, 2012 |
| 분류학적 정보 | |
| 역 | 진핵생물 |
| 계 | 크로말베올라타계 |
| 상문 | 피하낭상문 |
| 문 | 섬모충문 |
| 강 | 빈막강 |
| 아강 | 주모아강 |
| 목 | 전모목 |
| 과 | 짚신벌레과 |
2. 역사적 배경
''짚신벌레''는 17세기 후반에 현미경을 이용해 관찰된 최초의 섬모충 중 하나였다. 이들은 네덜란드 원생생물학 개척자인 안토니 판 레이우엔훅에게도 알려졌을 가능성이 높으며, 그의 동시대인인 크리스티안 호이겐스가 1678년의 편지에서 명확하게 묘사했다.[4] ''짚신벌레'' 종의 가장 초기의 그림은 1703년 영국 왕립학회 철학 회보에 익명으로 게재되었다.[5]
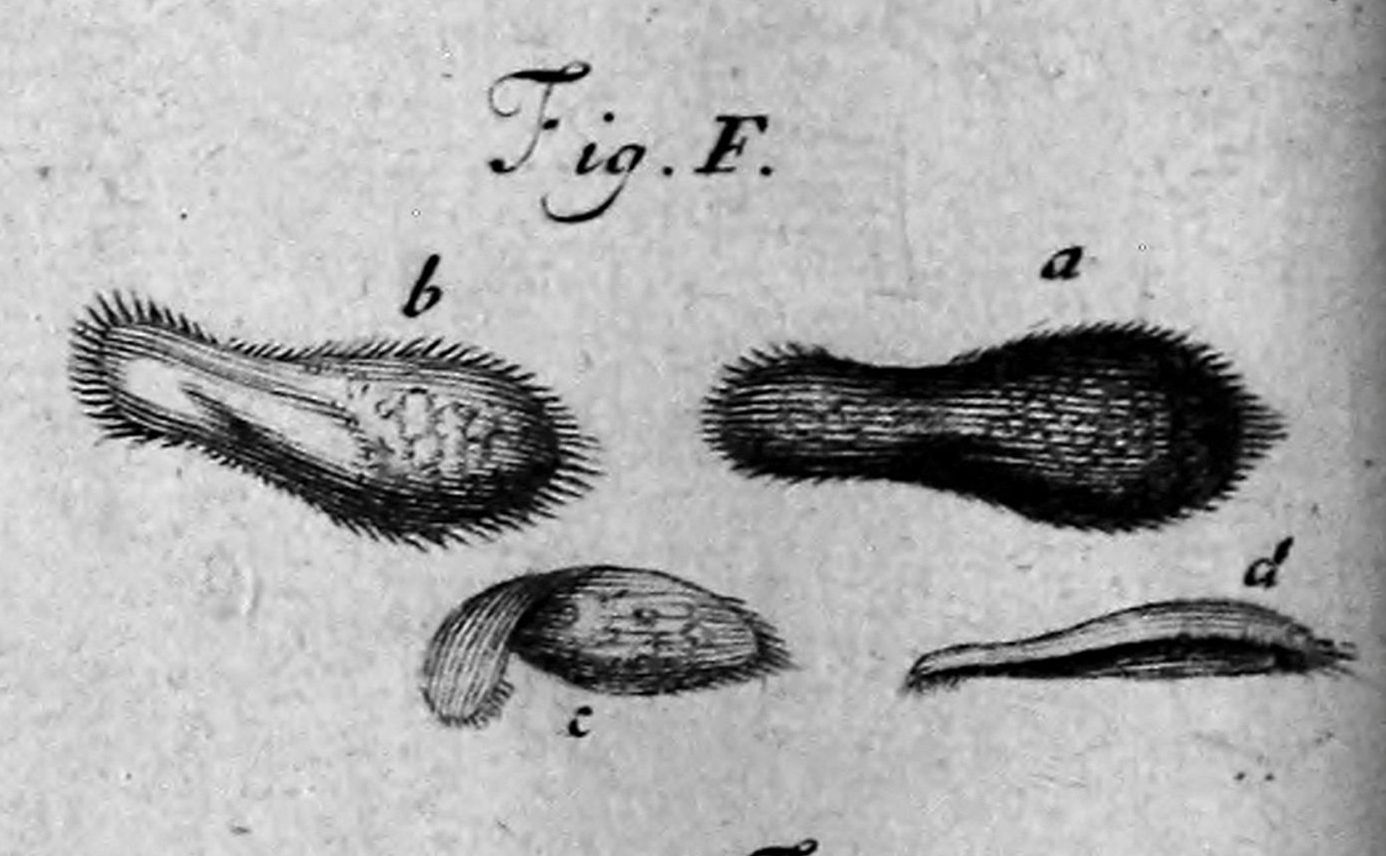
1718년, 프랑스 수학 교사이자 현미경 연구가인 루이 조블로는 물에 담근 참나무 껍질의 주입에서 발견한 미세한 물고기/poisson프랑스어에 대한 설명과 그림을 발표했다. 조블로는 이 생물에게 슬리퍼/"Chausson"프랑스어라는 이름을 붙였으며, "슬리퍼 벌레"라는 구어적인 별칭은 18세기와 19세기 내내 ''짚신벌레''에 사용되었다.[6]
"''짚신벌레''"라는 이름은 그리스어 (''paramēkēs'', "직사각형")에서 유래되었으며, 1752년 영국의 현미경 연구가인 존 힐이 "눈에 보이는 팔다리나 꼬리가 없고 불규칙한 직사각형 모양을 한 벌레"에 일반적으로 적용하면서 만들어졌다.[7] 1773년, O. F. 뮐러는 이 속을 린네의 분류 체계에 처음으로 포함시킨 연구자로서, ''짚신벌레''라는 이름을 채택했지만 철자를 ''Paramæcium''으로 변경했다.[8] 1783년, 요한 헤르만은 철자를 다시 한번 ''Paramœcium''으로 변경했다.[9] C. G. 에렌베르크는 1838년에 발표된 미세동물에 대한 주요 연구에서 힐의 원래 철자를 복원했으며, 대부분의 연구자들이 그의 방식을 따랐다.[10]
3. 형태 및 특징
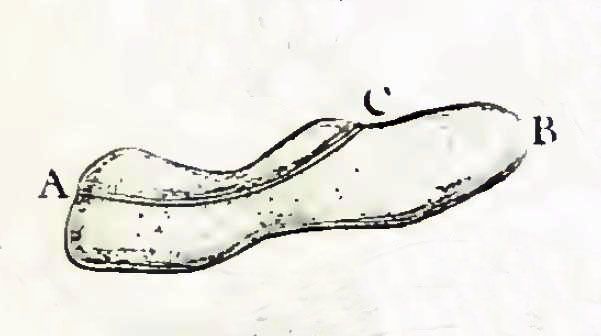
짚신벌레 종은 길이가 0.06mm에서 0.3mm에 이른다. 세포는 전형적으로 타원형, 길쭉하거나 발 또는 담배 모양이다.
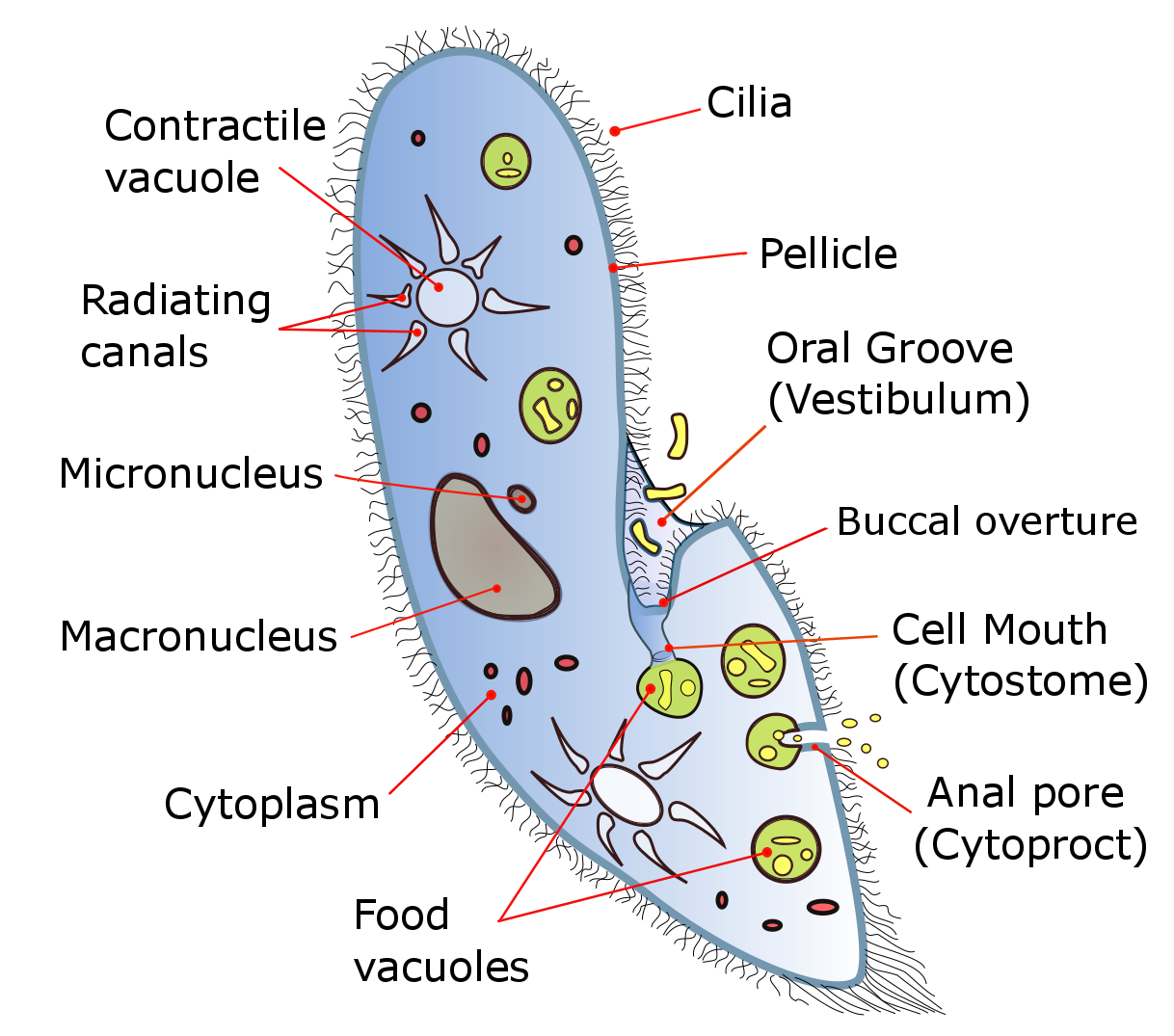
세포의 몸체는 펠리클이라고 하는 딱딱하지만 탄력적인 구조로 둘러싸여 있다. 펠리클은 바깥쪽 세포막 (원형질막), ''alveoli''라고 하는 납작한 막으로 둘러싸인 낭포층, 그리고 ''epiplasm''이라고 하는 안쪽 막으로 구성되어 있다. 펠리클은 매끄럽지 않고 육각형 또는 직사각형의 함몰부로 질감이 있다. 이러한 다각형 각각은 하나의 섬모가 튀어나오는 중앙 구멍으로 관통된다. 펠리클의 alveolar sac 사이에는 대부분의 짚신벌레 종에서 가깝게 간격을 둔 방추형 자포체가 있는데, 이는 종종 방어 목적으로 사용되는 얇고 비독성 실을 배출하는 폭발성 세포 소기관이다.[11][12]
일반적으로 항문공 (세포항문)은 세포의 복부 표면, 세포 후방 반에 위치해 있다. 모든 종에서 세포의 전방에서 중간 지점까지 이어지는 깊은 구강 홈이 있다. 이 홈은 끊임없이 박동하여 음식을 세포 안으로 끌어들이는 눈에 띄지 않는 섬모로 덮여 있다.[13] 짚신벌레는 주로 종속영양생물로, 세균 및 기타 작은 유기체를 먹고 산다. 일부 종은 세포의 세포질에서 운반되는 공생체 조류 (클로렐라)로부터 일부 영양분을 얻는 혼합영양생물이다.[14]
삼투 조절은 수축포에 의해 수행되며, 주변 환경에서 삼투에 의해 흡수된 액체를 보충하기 위해 세포에서 물을 능동적으로 배출한다.[15] 수축포의 수는 종에 따라 다르다.[15]
''짚신벌레''는 세균, 조류, 효모와 같은 미생물을 먹고 산다. 먹이를 섭취하기 위해 짚신벌레는 섬모를 움직여 먹이 생물과 약간의 물을 구강구(전정 또는 현관)을 통해 세포 안으로 쓸어 넣는다. 섬모로 덮인 구강구에서 먹이는 협곡이라고 불리는 더 좁은 구조로 들어간다. 거기에서 음식 입자는 세포구 또는 세포 입이라고 불리는 작은 구멍을 통과하여 세포 내부로 이동한다. 음식이 세포로 들어가면 음식 액포에 모이게 되는데, 이 액포는 주기적으로 닫히고 세포질로 방출되어 세포 내용물의 흐름 운동, 즉 세포질 유동이라고 불리는 과정을 통해 세포체를 순환하기 시작한다. 음식 액포가 이동하면서 세포질의 효소가 들어가 내용물을 소화시킨다. 효소 소화가 진행되면서 액포 내용물은 더욱 산성화된다. 액포가 형성된 지 5분 이내에 내용물의 pH는 7에서 3으로 떨어진다.[20] 소화된 영양분이 세포질로 들어가면서 액포는 수축한다. 완전히 소화된 액포가 배설공에 도달하면 파열되어 세포 밖으로 노폐물 내용물을 배출한다.[21][22][23]
일부 짚신벌레 종은 다른 유기체와 상호 공생 관계를 형성한다. ''Paramecium bursaria''(부르사리아 짚신벌레)와 ''Paramecium chlorelligerum''(클로렐리게럼 짚신벌레)은 내생 공생 녹조류를 품고 있으며, 이로부터 영양분을 얻고 ''Didinium nasutum(디디니움 나수툼)''과 같은 포식자로부터 어느 정도 보호를 받는다.[24][25] 짚신벌레 종에서 수많은 세균 내생 공생체가 확인되었다.[26] 카파 입자로 알려진 일부 세포 내 세균은 짚신벌레에게 카파 입자가 없는 다른 짚신벌레 균주를 죽일 수 있는 능력을 부여한다.[26]
세포 길이는 90-150µm, 폭은 40µm 정도이다. 이름은 납작한 인상을 주지만 실제로는 원통형에 가깝고, 배 부분에는 세포구라는 움푹 들어간 부분이 약간 비틀린 듯이 들어가 있다. 세포 표면에는 약 3,500개의 섬모를 가지고 있으며, 그 섬모를 사용하여 헤엄친다. 섬모는 체표의 섬모열을 따라 나 있다. 섬모는 세포 전체에 거의 균일하게 나 있지만, 세포구 안쪽 부분에서는 세포 인두를 향해 특수한 배치와 움직임이 보인다. 섬모는 5% 에탄올 처리 등의 스트레스에 의해 탈리될 수 있다(탈섬모). 세포구 안쪽에는 세포 내로 먹이를 섭취하는 세포 인두가 있으며, 먹이는 이곳을 통과하여 식포로 섭취된다. 식포 내에서 소화가 이루어지고, 유용한 성분은 세포 내로 흡수되면서 식포는 세포 내를 돌도록 이동한다. 배설물은 세포 후방의 세포항문에서 방출된다.
세포 앞뒤에는 크고 별 모양 또는 꽃으로 보이는 방사형 세포 기관이 있다. 이를 수축포라고 하며, 세포 내 삼투압 조절을 담당한다. 수축포는 중앙의 원형 부분과 주변에 꽃잎처럼 늘어선 물방울형 부분으로 구성된다. 물 배출 시에는 먼저 물방울형 부분에 물이 모이고, 여기에서 중앙의 원형 부분으로 물이 옮겨져 세포 외로 물이 방출된다.
세포 내에는 크고 작은 두 개의 세포핵이 있으며, 각각 대핵과 소핵이라고 불린다. 대핵은 평소 활동에 관여한다. 소핵은 생식핵이라고도 불리며, 유성 생식에 관해 작용한다고 여겨진다. 세포 내에 기능적으로 분화된 핵을 갖는 것은 섬모충류의 특징이다.
3. 1. 섬모 운동
짚신벌레는 몸 바깥쪽에 촘촘하게 배열된 섬모의 채찍 같은 움직임으로 이동한다. 각 섬모의 움직임은 두 단계로 이루어진다. 섬모가 비교적 뻣뻣한 상태로 움직이는 빠르고 "유효 타격" 단계와, 섬모가 느슨하게 한쪽으로 구부러지고 시계 반대 방향으로 앞으로 휩쓸리는 느린 "회복 타격" 단계가 그것이다. 촘촘하게 배열된 섬모들은 조화롭게 움직이며, "섬모 카펫"을 가로질러 활동의 파동을 만들어낸다.
짚신벌레는 전진하면서 물속에서 나선형으로 움직인다. 장애물을 만나면 섬모의 "유효 타격"이 반전되어 잠시 뒤로 헤엄친 다음 다시 앞으로 나아가는데, 이를 회피 반응이라고 한다.
짚신벌레는 물속을 추진하는 데 에너지의 절반 이상을 소비하는 것으로 계산되었다. 이러한 섬모 운동 방식의 효율은 1% 미만인 것으로 밝혀졌다. 그럼에도 불구하고 이 낮은 비율은 짚신벌레류의 구성원과 같이 짧은 섬모를 가진 유기체가 달성할 수 있는 최대 이론적 효율에 가깝다.
세포 표면에는 약 3,500개의 섬모를 가지고 있으며, 그 섬모를 사용하여 헤엄친다. 섬모는 체표의 섬모열을 따라 나 있다. 섬모는 세포 전체에 거의 균일하게 나 있지만, 세포구 안쪽 부분에서는 세포 인두를 향해 특수한 배치와 움직임이 보인다. 섬모는 5% 에탄올 처리 등의 스트레스에 의해 탈리될 수 있다(탈섬모).
3. 2. 세포 구조
짚신벌레는 세균, 조류, 효모와 같은 미생물을 먹고 산다. 섬모를 움직여 먹이와 물을 세포 안으로 넣고, 세포구와 세포 인두를 통해 식포를 형성한다. 식포는 세포질 내에서 순환하며 효소에 의해 소화되고, 영양분은 흡수된다. 소화된 5분 이내에 내용물의 pH는 7에서 3으로 떨어진다.[20] 소화 후 남은 찌꺼기는 세포항문을 통해 배출된다.[21][22][23]
세포 주변을 둘러싼 섬모는 이동에 사용된다. 세포 오른쪽 아래와 오른쪽에 별 모양의 수축포는 삼투압 조절을 담당한다. 중앙 아래의 밋밋한 회색 부분은 대핵이다.
세포 길이는 90-150µm, 폭은 40µm 정도이며, 원통형에 가깝다. 세포 표면에는 약 3,500개의 섬모가 있으며, 세포구 안쪽에는 세포 인두가 있어 먹이를 섭취한다. 세포 앞뒤의 방사형 세포 기관인 수축포는 세포 내 삼투압을 조절한다. 수축포는 중앙의 원형 부분과 주변의 물방울형 부분으로 구성되며, 물 배출 시 물방울형 부분에 물이 모였다가 중앙의 원형 부분을 통해 세포 밖으로 방출된다.
세포 내에는 대핵과 소핵 두 종류의 세포핵이 있다. 대핵은 평소 활동에, 소핵은 유성 생식에 관여한다.
4. 생태
짚신벌레는 주로 논, 늪, 연못 등 담수 정체역에 분포한다.[24] 세포 표면의 섬모에 의해 유영하기 때문에 단세포 생물로서는 이동력이 크다. 장애물 등에 접촉하면 섬모 역전에 의해 유영 방향을 반전시킨다(후퇴 유영).[24]
짚신벌레는 주로 진정세균을 먹이로 하는 세균 식성 생물이다. 다만, 미즈색 짚신벌레(Paramecium bursariala)는 체내에 녹조류인 클로렐라를 공생시키고 있어, 광합성 산물의 환류를 받아 생활하는 것도 가능하다. 짚신벌레의 포식자는 대형의 아메바나, 디디니움(Didiniumla, 소금벌레)과 같은 다른 섬모충이다. 디디니움은 세포 전단의 구吻부에 익스트루솜(extrusome)이라고 불리는 사출 기관을 가지고 있으며, 이것을 짚신벌레에 쏘아 움직임을 멈추고 삼켜서 소화한다.[24]
일부 짚신벌레 종은 다른 유기체와 상호 공생 관계를 형성한다. ''Paramecium bursaria''(부르사리아 짚신벌레)와 ''Paramecium chlorelligerum''(클로렐리게럼 짚신벌레)은 내생 공생 녹조류를 품고 있으며, 이로부터 영양분을 얻고 ''Didinium nasutum(디디니움 나수툼)''과 같은 포식자로부터 어느 정도 보호를 받는다.[24][25] 짚신벌레 종에서 수많은 세균 내생 공생체가 확인되었다.[26] 카파 입자로 알려진 일부 세포 내 세균은 짚신벌레에게 카파 입자가 없는 다른 짚신벌레 균주를 죽일 수 있는 능력을 부여한다.[26]
5. 생식
무성 생식은 분열에 의해 이루어진다. 다른 섬모충과 마찬가지로, 체축 방향의 앞뒤 부분으로 나뉘어 분열한다. 유성 생식으로는 세포의 접합이 이루어지는데, 그 방법은 다소 특이하다. 접합에 앞서 대핵이 소실됨과 동시에 생식핵인 소핵이 감수 분열을 하여 4개의 핵으로 나뉜다. 이 중 3개는 소실되고, 남은 1개가 다시 2개로 분열되며, 이 중 하나의 핵을 서로 교환한다. 그 후, 각 세포 내의 2개의 핵이 융합함으로써 접합이 완료된다. 이 동안 2개체의 짚신벌레는 서로 같은 방향을 향해 가까이 붙어 있지만, 세포 간에 연락을 가질 뿐 세포 자체가 융합되지는 않는다. 접합 후에는 대핵이 소핵을 바탕으로 다시 형성된다.
5. 1. 무성 생식
짚신벌레의 생식은 무성 생식으로, 이분열에 의해 이루어진다.[3] 이분열 동안 대핵은 일종의 무사분열로 분열하고, 소핵은 분열을 겪는다.[3] 그런 다음 세포는 가로로 분열되고, 각 새로운 세포는 소핵과 대핵의 복사본을 얻는다.[3]이분열은 영양 세포 주기 동안 자발적으로 발생할 수 있다. 특정 조건에서는 자가 수정 (자가수정)이 선행될 수 있거나,[33] 호환 가능한 교배 유형의 짚신벌레가 일시적으로 융합하여 유전 물질을 교환하는 접합 직후에 발생할 수 있다.
5. 2. 유성 생식 (접합)
짚신벌레류와 같은 섬모충류에서 접합은 세포 내에서 유전자 재조합과 핵 재조직화를 초래하는 성적 현상이다.[31][26] 접합 동안 호환 가능한 짝짓기 유형의 두 짚신벌레가 함께 모여 세포질 사이에 다리를 형성한다. 각 미세핵은 감수 분열을 거치고, 반수체 미세핵이 다리를 통해 교환된다. 접합 후 세포는 분리된다. 오래된 대핵은 파괴되고, 두 접합 후 세포는 미세핵에서 DNA 증폭을 통해 새로운 대핵을 형성한다.[31] 접합 후 하나 이상의 "접합 후 분열"이 이어진다.[34]무성 생식은 분열에 의해 이루어진다. 다른 섬모충과 마찬가지로, 체축 방향의 앞뒤 부분으로 나뉘어 분열한다. 유성 생식으로는 세포의 접합이 이루어지는데, 그 방법은 다소 특이하다. 접합에 앞서 대핵이 소실됨과 동시에 생식핵인 소핵이 감수 분열을 하여 4개의 핵으로 나뉜다. 이 중 3개는 소실되고, 남은 1개가 다시 2개로 분열되며, 이 중 하나의 핵을 서로 교환한다. 그 후, 각 세포 내의 2개의 핵이 융합함으로써 접합이 완료된다. 이 동안 2개체의 짚신벌레는 서로 같은 방향을 향해 가까이 붙어 있지만, 세포 간에 연락을 가질 뿐 세포 자체가 융합되지는 않는다. 접합 후에는 대핵이 소핵을 바탕으로 다시 형성된다.
6. 노화
짚신벌레는 성장의 무성 분열 단계에서 유사 분열을 통해 세포 분열을 반복하면서 점차 활력을 잃는 클론 노화를 겪는다.[35] ''Paramecium tetraurelia''와 같은 종은 자가수정 또는 접합을 거치지 않으면 약 200번의 분열 후 소멸한다.[35] 클론 노화의 원인은 대핵에 있으며, 클론 젊은 ''짚신벌레''의 대핵을 이식하면 수명이 연장되지만, 세포질 이식은 그렇지 않다.[35] 클론 노화 동안 DNA 손상이 크게 증가하며,[39] 대핵 내 DNA 손상이 ''P. tetraurelia'' 노화의 주요 원인으로 보인다.[39] 이는 노화의 DNA 손상 이론에서 설명하는 다세포 진핵생물의 노화와 유사하다.
클론 노화된 ''P. tetraurelia''는 접합 또는 자동교배를 통해 감수 분열을 하면 유전적 후손이 젊어져 더 많은 유사 분열을 할 수 있게 된다.[40] 접합 또는 자동교배 동안 소핵은 감수 분열을 하고, 오래된 대핵은 분해되며, 감수 분열을 거친 소핵 DNA 복제로 새로운 대핵이 형성된다.[40] 새 대핵은 DNA 손상이 거의 없어, 클론 노화가 DNA 손상 축적 때문이며, 젊어짐은 감수 분열 동안 소핵에서 손상 복구 때문이라는 것을 뒷받침한다. 감수 분열은 ''P. tetraurelia''에서 DNA 복구와 젊어짐을 위한 적응으로 보인다.[40] ''P. tetraurelia''에서 CtlP 단백질은 유성 생식 동안 감수 분열 완료와 생존 가능한 유성 자손 회복에 필수적이며,[1] Mre11 뉴클레아제 복합체는 상동 재조합 동안 이중 가닥 절단의 정확한 처리 및 복구에 필수적이다.[1]
기아 반응으로 나타나는 감수 분열 및 자웅동체의 적응적 이점은 ''P. tetraurelia''에서 새로운 유전적 변이 생성과는 무관한 것으로 보인다.[41] 이는 감수 분열의 분자 메커니즘이 유전적 다양성에 대한 성의 부수적 효과와 관계없이 적합성 이점을 제공함을 시사한다.[1][42]
7. 배양
짚신벌레는 삼투 영양을 하는 것이 아니라 포식성이기 때문에, 배양 시에는 세균이나 효모 등을 먹이로 준비해야 한다. 일반적으로는 짚이나 상추 등을 삶아 그 액체를 배지로 하여 고초균을 늘려 먹이로 한다. 고초균 배양이 어려운 경우에는 시판되는 건조 효모 등을 이용하는 방법이 있다. 가루 형태의 녹차도 먹이로 사용할 수 있다.
8. 하위 종
- ''Paramecium aurelia''
- ''Paramecium bursaria''
- ''Paramecium caudatum''
- ''Paramecium tetraurelia''
짚신벌레속 aurelia 종 복합체
- ''Paramecium primaurelia''
- ''Paramecium biaurelia''
- ''Paramecium triaurelia''
- ''Paramecium tetraurelia''
- ''Paramecium pentaurelia''
- ''Paramecium sexaurelia''
- ''Paramecium septaurelia''
- ''Paramecium octaurelia''
- ''Paramecium novaurelia''
- ''Paramecium decaurelia''
- ''Paramecium undecaurelia''
- ''Paramecium dodecaurelia''
- ''Paramecium tredecaurelia''
- ''Paramecium quadecaurelia''
- ''Paramecium sonneborni''
기타 종
- ''Paramecium buetschlii''
- ''Paramecium bursaria''
- ''Paramecium calkinsi''
- ''Paramecium caudatum''
- ''Paramecium chlorelligerum''
- ''Paramecium duboscqui''
- ''Paramecium grohmannae''
- ''Paramecium jenningsi''
- ''Paramecium multimicronucleatum''
- ''Paramecium nephridiatum''
- ''Paramecium polycaryum''
- ''Paramecium putrinum''
- ''Paramecium schewiakoffi''
- ''Paramecium woodruffi''
8. 1. ''Paramecium aurelia'' 종 복합체
''Paramecium primaurelia'', ''Paramecium biaurelia'', ''Paramecium triaurelia'', ''Paramecium tetraurelia'', ''Paramecium pentaurelia'', ''Paramecium sexaurelia'', ''Paramecium septaurelia'', ''Paramecium octaurelia'', ''Paramecium novaurelia'', ''Paramecium decaurelia'', ''Paramecium undecaurelia'', ''Paramecium dodecaurelia'', ''Paramecium tredecaurelia'', ''Paramecium quadecaurelia'', ''Paramecium sonneborni''는 모두 짚신벌레속 ''aurelia'' 종 복합체에 속한다.8. 2. 기타 종
짚신벌레속에는 짚신벌레(Paramecium caudatum), Paramecium aurelia, Paramecium bursaria, Paramecium tetraurelia 외에도 다양한 종들이 존재한다. 짚신벌레속 aurelia 종 복합체에는 Paramecium primaurelia, Paramecium biaurelia, Paramecium triaurelia, Paramecium tetraurelia, Paramecium pentaurelia, Paramecium sexaurelia, Paramecium septaurelia, Paramecium octaurelia, Paramecium novaurelia, Paramecium decaurelia, Paramecium undecaurelia, Paramecium dodecaurelia, Paramecium tredecaurelia, Paramecium quadecaurelia, Paramecium sonneborni 등이 있다.그 외에도, ''Paramecium buetschlii'', ''Paramecium bursaria'', ''Paramecium calkinsi'', ''Paramecium chlorelligerum'', ''Paramecium duboscqui'', ''Paramecium grohmannae'', ''Paramecium jenningsi'', ''Paramecium multimicronucleatum'', ''Paramecium nephridiatum'', ''Paramecium polycaryum'', ''Paramecium putrinum'', ''Paramecium schewiakoffi'', ''Paramecium woodruffi'' 등이 있다.
9. 학습
짚신벌레의 학습 능력에 대한 문제는 많은 실험의 대상이 되었으나, 그 결과는 불확실했다. 그러나 2006년에 발표된 연구에 따르면, ''Paramecium caudatum''은 6.5 볼트의 전류를 가하여 밝기 수준을 구별하도록 훈련될 수 있다.[29] 이 실험은 신경계가 없는 유기체에서 세포 기억, 또는 후생유전적 학습의 가능한 사례로 인용되었다.[30]
참조
[1]
웹사이트
GBIF
https://www.gbif.org[...]
[2]
사전
paramecium
Merriam-Webster
[3]
서적
The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature
https://books.google[...]
Springer Science & Business Media
[4]
서적
Antony van Leeuwenhoek and his "Little Animals"
Dover
[5]
논문
Unmasking "The Eldest Son of The Father of Protozoology": Charles King
https://hal.sorbonne[...]
2019-08-01
[6]
서적
Description et usages de Plusieurs Nouveaux Microscopes, tant simple que composez
Jacques Collombat
[7]
서적
An History of Animals
https://archive.org/[...]
Thomas Osborne
[8]
서적
Vermivm terrestrium et fluviatilium, seu, Animalium infusoriorum, helminthicorum et testaceorum, non marinorum, succincta historia
https://www.biodiver[...]
apud Heineck et Faber
1773
[9]
웹사이트
Paramoecium Hermann, 1783
https://www.gbif.org[...]
[10]
논문
The structure, life history, and intrageneric relationships of Paramecium calkinsi, sp. nov.
2021-09-01
[11]
서적
The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature
https://www.springer[...]
Springer Netherlands
2008
[12]
서적
The Biology of Paramecium
https://books.google[...]
Springer Science & Business Media
2012-12-06
[13]
서적
British and other freshwater ciliated protozoa
Cambridge University Press
[14]
논문
Mixotrophy in Ciliates
[15]
서적
Campbell Biology
https://archive.org/[...]
Pearson Education
[16]
논문
Mechanics of ciliary locomotion
1974-02-01
[17]
논문
Artificial deciliation causes loss of calcium-dependent responses in Paramecium
[18]
논문
Substantial energy expenditure for locomotion in ciliates verified by means of simultaneous measurement of oxygen consumption rate and swimming speed
[19]
논문
Finding the ciliary beating pattern with optimal efficiency
http://www.pnas.org/[...]
2011-09-20
[20]
서적
The Biology of Paramecium
https://www.springer[...]
Springer US
1986
[21]
서적
The Biology of Paramecium
Plenum Press
[22]
서적
Campbell Biology
https://archive.org/[...]
Pearson Education
[23]
논문
The food-vacuole in Paramecium
https://www.biodiver[...]
1947-02-01
[24]
논문
Feeding Behaviour of Didinium nasutum on Paramecium bursaria with Normal or Apochlorotic Zoochlorellae
[25]
논문
Morphological and Molecular Characterization of Paramecium (Viridoparamecium nov. subgen.) chlorelligerum Kahl (Ciliophora)
[26]
논문
Kappa and Other Endosymbionts in Paramecium aurelia
1974-06-01
[27]
논문
Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia
2006-11-01
[28]
논문
Different modes of stop codon restriction by the Stylonychia and Paramecium eRF1 translation termination factors
2007-06-26
[29]
논문
Discrimination Learning in Paramecia (P. caudatum)
http://opensiuc.lib.[...]
2006-09-01
[30]
논문
Epigenetic learning in non-neural organisms
[31]
논문
DNA of ciliated protozoa
[32]
논문
Nuclear phenomena during conjugation and autogamy in ciliates
http://www.sciencedi[...]
1972
[33]
논문
Autogamy in Paramecium cell cycle stage-specific commitment to meiosis
1986-10-01
[34]
논문
Nuclear phenomena during conjugation and autogamy in ciliates
http://www.sciencedi[...]
1972
[35]
논문
Clonal aging in Paramecium tetraurelia. II. Evidence of functional changes in the macronucleus with age
[36]
논문
DNA repair and longevity assurance in Paramecium tetraurelia
[37]
논문
Accumulation of DNA damages in aging Paramecium tetraurelia
1986-07
[38]
논문
Lack of telomere shortening during senescence in Paramecium
http://www.pnas.org/[...]
[39]
서적
Aging, Sex, and DNA Repair
Academic Press
[40]
논문
Identification of a miniature Sae2/Ctp1/CtIP ortholog from Paramecium tetraurelia required for sexual reproduction and DNA double-strand break repair
https://hal.archives[...]
[41]
논문
What's Genetic Variation Got to Do with It? Starvation-Induced Self-Fertilization Enhances Survival in Paramecium
2020-05
[42]
논문
Genetic damage, mutation, and the evolution of sex
1985-09
[43]
백과사전
짚신벌레 - 원생동물
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com