유공충
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
유공충은 단단한 껍질을 가진 원생생물로, 해양 환경에 널리 분포하며 화석 기록이 풍부하여 생물 층서 및 고기후 연구에 중요한 정보를 제공한다. 유공충은 껍질의 형태와 구성에 따라 분류되며, 다양한 환경 지표로 활용된다. 석유 탐사, 고고학, 환경 연구 등 다양한 분야에서 활용되며, 껍질은 퇴적되어 석회암을 형성하기도 한다.
더 읽어볼만한 페이지
- 원생생물 - 코아노조아
코아노조아는 칼라 구조를 특징으로 하는 깃편모충류와 동물을 포함하는 계통 분류군으로, 동물의 기원과 밀접한 관련이 있으며, 동물 진화 연구에 중요한 연결고리를 제공한다. - 원생생물 - 짚신벌레
짚신벌레는 담수 환경에 사는 단세포 섬모충류로, 섬모를 이용해 움직이며 세균을 섭취하고, 대핵과 소핵을 가지며 이분열과 접합으로 번식한다. - 생물 계통도에 관한 - 눈표범
눈표범은 고양이과 동물로 중앙아시아와 남아시아의 고산 지대에 서식하며, 길고 두꺼운 털, 큰 발, 긴 꼬리가 특징이며, 유전자 분석 결과 호랑이와 가장 가까운 종으로 밝혀졌으며, 밀렵과 서식지 파괴로 인해 개체 수가 감소하여 여러 국가에서 보호 노력이 진행 중이다. - 생물 계통도에 관한 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다.
| 유공충 - [생물]에 관한 문서 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
| 학명 | Foraminifera |
| 명명자 | d'Orbigny, 1826 |
| 로마자 표기 | fəˌræmənɪfərə |
| 발음 | /fə|ˈ|r|æ|m|ə|ˌ|n|ɪ|f|ə|r|ə/ |
| 별칭 | 유공충 Foraminifers Benthic Foraminifera Planktic Foraminifera |
| 생물학적 분류 | |
| 계 | 리자리아 |
| 문 | 유공충문 |
| 하위 분류 | |
| 하위 분류 | Monothalamea Allogromiida Astrorhizida Xenophyophorea Reticulomyxa Tubothalamea Miliolida Spirillinida Silicoloculinida Globothalamea Textulariida Rotaliida Globigerinida Carterinida Robertinida Fusulinida? – extinct incertae sedis Involutinida Lagenida |
| 특징 | |
| 크기 | 미세한 크기 |
| 서식지 | 해양 퇴적물 토양 담수 |
| 껍데기 | CaCO3 키틴질 규소질 모래 알갱이 등 |
| 특징 | 위족 단세포 아메바형 유공충은 단세포 생물이다. 퇴적물에 서식하는 유공충은 저서성으로 분류된다. |
| 생태 | |
| 식성 | 박테리아 조류 유기물 입자 작은 무척추동물 |
| 분포 | 해양 담수 토양 |
| 역할 | 생물 지표 고환경 연구 퇴적물 형성 |
| 화석 기록 | |
| 출현 시기 | 에디아카라기 후기 - 현재 |
2. 역사
펠릭스 뒤자르댕은 1835년에 유공충이 원생동물임을 처음으로 알아냈다.[18] 1852년 알시드 도르비니는 껍질 형태를 기준으로 72개의 유공충 속을 제시하는 분류 체계를 발표했으나, 동료들에게 심한 비판을 받았다.[17]
1884년 H.B. 브레이디는 챌린저호 탐험에서 발견한 유공충을 설명하며 10개 과와 29개 아과를 제시했다. 그의 분류 체계는 여러 특징을 고려하여 응집성과 석회질 속을 밀접하게 분류했다.
1920년대 후반 조셉 어거스틴 커시먼은 벽 구성을 가장 중요한 특징으로 보는 분류 체계를 제시하여 널리 받아들여졌으나, "생물학적으로 건전하지 않다"는 비판도 받았다. 지질학자 아이린 크레스핀은 유공충 연구를 통해 많은 논문을 발표했다.[21]
1964년 타판과 로블리히는 껍질 벽 미세 구조를 기반으로 유공충을 분류했으며, 이는 현재까지 사용되는 분류 체계의 기반이 되었다.[17] 2013년 파울로브스키의 분자 계통학적 연구는 타판과 로블리히의 분류를 대체로 확인하고 주요 유공충 그룹 간 상위 관계를 밝혀냈다.[22]
2. 1. 고대 기록
기원전 5세기에 헤로도토스는 기자 대피라미드를 구성하는 암석을 이루는 물질로 유공충을 언급했다. 이것들은 오늘날 ''Nummulites''으로 인식된다. 기원전 1세기 스트라보는 같은 유공충을 언급하면서 피라미드 건설 노동자들이 남긴 렌틸콩의 잔해라고 추측했다.[15]로버트 훅은 현미경으로 유공충을 관찰하여 1665년 그의 저서 ''현미경술(Micrographia)''에 그림으로 묘사하였다.[16]

안톤 판 레이우엔훅은 1700년 유공충의 껍질을 기술하고 그림으로 묘사하여 작은 조개라고 설명했는데, 그의 그림은 ''엘피디움속''으로 알아볼 수 있다.[17] 초기 연구자들은 유공충을 ''앵무조개속''으로 분류하면서 특정 두족류와의 유사성을 지적했다. 로렌츠 슈펭글러는 1781년 유공충의 격벽에 구멍이 있다는 것을 인지하여 결국 이 그룹에 이름을 부여하게 되었다.[18] 슈펭글러는 또한 유공충의 격벽이 앵무조개와 반대 방향으로 휘어져 있고 신경관이 없다는 점을 지적했다.[19]
알시드 도르비니는 1826년 그의 저서에서 유공충을 작은 두족류의 한 무리로 간주하고 그들의 이상한 형태를 언급하면서 위족을 촉수로 해석하고 매우 퇴화된(실제로는 없는) 머리를 지적했다.[20] 그는 이 그룹에 ''foraminifères'' 즉 "구멍이 있는 것들"이라는 이름을 붙였다.[14]
펠릭스 뒤자르댕은 1835년에 유공충의 원생동물적 특성을 처음으로 인지했다.[18]
2. 2. 초기 연구
헤로도토스는 기원전 5세기에 기자 대피라미드를 구성하는 암석에서 ''Nummulites''(Nummulite) 속으로 알려진 유공충을 최초로 기록했다. 기원전 1세기 스트라보는 이 유공충을 피라미드 건설 노동자들이 남긴 렌틸콩 잔해로 추정했다.[15]로버트 훅은 1665년 저서 ''현미경술(Micrographia)''에서 현미경으로 관찰한 유공충을 그림과 함께 묘사했다.
그는 유공충을 작은 물 달팽이 껍데기와 비슷하다고 설명했다.[16] 1700년 안톤 판 레이우엔훅은 유공충 껍질을 작은 조개라고 묘사하며 그림으로 남겼는데, 그의 그림은 ''엘피디움속''으로 확인된다.[17] 초기 연구자들은 유공충을 앵무조개속과 유사한 두족류로 분류했다. 1781년 로렌츠 슈펭글러는 유공충 격벽에 구멍이 있음을 발견하고, 앵무조개와 달리 격벽이 반대 방향으로 휘어져 있으며 신경관이 없음을 확인했다.[18][19]
1826년 알시드 도르비니는 유공충을 작은 두족류 무리로 간주하고, 위족을 촉수로 해석했다. 그는 껍질 구획 사이에 구멍이 있는 특징 때문에 앵무조개나 암모나이트와 구별하여 ''foraminifères''(구멍이 있는 것들)라는 이름을 붙였다.[14][20]
1835년 펠릭스 뒤자르댕은 유공충이 원생동물임을 처음으로 인지했다.[18] 1852년 도르비니는 껍질 형태에 기반한 72개 유공충 속 분류 체계를 제시했으나, 동료들로부터 비판받았다.[17]
H.B. 브레이디는 1884년 논문에서 ''챌린저''호 탐험을 통해 발견된 유공충을 설명하며 10개 과와 29개 아과를 제시했다. 그의 분류 체계는 응집성과 석회질 속을 밀접하게 분류하여 다양한 특징을 강조했다.
조셉 어거스틴 커시먼은 1920년대 후반 유공충 분류에서 벽 구성을 가장 중요한 특징으로 보았다. 그의 분류 체계는 널리 사용되었으나, "생물학적으로 건전하지 않다"는 비판도 있었다.
지질학자 아이린 크레스핀은 유공충 분야에서 광범위한 연구를 수행하여 독자적으로 약 90편, 공동으로 20편 이상의 논문을 발표했다.[21]
커시먼의 체계는 1964년 타판과 로블리히의 분류가 나올 때까지 지배적이었다. 이 분류는 껍질 벽 미세 구조를 기반으로 현재 사용되는 유공충 그룹을 제시했다.[17] 파울로브스키(2013)의 분자 계통학적 연구는 타판과 로블리히의 그룹을 대체로 확인했으며, 주요 유공충 그룹 간 상위 수준 관계를 밝혀냈다.[22]
2. 3. 19세기 이후 연구
펠릭스 뒤자르댕은 1835년에 유공충이 원생동물이라는 것을 처음으로 알아냈다.[18] 1852년, 알시드 도르비니는 껍질 형태를 기준으로 72개의 유공충 속을 제시하는 분류 체계를 발표했지만, 동료들에게 심한 비판을 받았다.[17]1884년 H.B. 브레이디는 챌린저호 탐험에서 발견한 유공충을 설명하는 논문을 발표했다. 브레이디는 지층 범위는 거의 고려하지 않고 10개 과와 29개 아과를 제시했다. 그의 분류 체계는 여러 가지 다른 특징이 분류군을 구분해야 한다는 생각을 강조하여, 응집성과 석회질 속을 밀접한 관계로 분류했다.
이러한 분류 체계는 1920년대 후반 조셉 어거스틴 커시먼의 연구가 나올 때까지 유지되었다. 커시먼은 유공충 분류에서 벽 구성을 가장 중요한 특징으로 보았다. 그의 분류 체계는 널리 받아들여졌지만, "생물학적으로 건전하지 않다"는 비판도 받았다. 아이린 크레스핀은 유공충 분야에서 광범위한 연구를 수행하여 독자적으로 약 90편의 논문(유공충에 관한 주목할 만한 연구 포함)과 다른 과학자들과 공동으로 20편 이상의 논문을 발표했다.[21]
커시먼의 체계는 1964년 타판과 로블리히의 분류가 나올 때까지 지배적인 분류 체계로 남아 있었는데, 이 분류는 껍질 벽의 미세 구조를 기반으로 오늘날에도 사용되는 일반적인 그룹으로 유공충을 분류했다.[17] 이러한 그룹들은 상위 수준의 분류 체계에 따라 다양하게 이동되었다. 파울로브스키(2013)의 분자 계통학적 연구는 일부가 다계통군 또는 단계통군으로 밝혀진 것을 제외하고는 대체로 타판과 로블리히의 그룹을 확인했으며, 이 연구는 주요 유공충 그룹 간의 상위 수준 관계를 확인하는 데에도 도움이 되었다.[22]
2. 4. 20세기 이후 연구
조셉 어거스틴 커시먼은 1920년대 후반에 유공충 분류에서 벽 구성을 가장 중요한 특징으로 보았다. 그의 분류 체계는 널리 받아들여졌지만 "생물학적으로 건전하지 않다"는 비판도 받았다.[17] 지질학자 아이린 크레스핀은 이 분야에서 광범위한 연구를 수행하여 독자적으로 약 90편의 논문(유공충에 관한 주목할 만한 연구 포함)과 다른 과학자들과 공동으로 20편 이상의 논문을 발표했다.[21]커시먼의 체계는 1964년 타판과 로블리히의 분류가 나올 때까지 지배적인 분류 체계로 남아 있었는데, 이 분류는 껍질 벽의 미세 구조를 기반으로 오늘날에도 사용되는 일반적인 그룹으로 유공충을 분류했다.[17] 이러한 그룹들은 상위 수준의 분류 체계에 따라 다양하게 이동되었다. 파울로브스키(2013)의 분자 계통학적 연구는 일부가 다계통군 또는 단계통군으로 밝혀진 것을 제외하고는 대체로 타판과 로블리히의 그룹을 확인했으며, 이 연구는 주요 유공충 그룹 간의 상위 수준 관계를 확인하는 데에도 도움이 되었다.[22]
3. 형태
대부분의 유공충에서 가장 눈에 띄는 것은 단단한 껍질, 즉 각(殼)이다. 각은 하나 또는 여러 개의 방으로 구성될 수 있으며, 단백질, 퇴적물 입자, 방해석, 아라고나이트, 또는 (드물게) 규산으로 구성될 수 있다.[24] 일부 유공충은 각이 전혀 없기도 하다.[32] 연체동물이나 산호와 같은 다른 껍질 분비 유기체와 달리, 유공충의 각은 세포막 내부, 즉 원형질 내부에 위치한다. 세포소기관은 각의 구획 내에 위치하며, 각의 구멍을 통해 위족에서 내부 세포로, 그리고 그 반대로 물질 이동이 가능하다.[33]
유공충 세포는 과립성 내형질과 투명한 외형질로 나뉘며, 이로부터 위족 그물이 단일 개구부 또는 각의 많은 구멍을 통해 나타난다. 개별 위족은 특징적으로 양방향으로 흐르는 작은 과립을 가진다.[34] 유공충은 ''과립망상 위족''을 가지는 것이 독특하며, 현미경으로 보면 위족이 과립으로 보인다. 이러한 위족은 종종 길쭉하고 서로 분리되고 다시 합쳐질 수 있으며, 세포의 필요에 따라 신장되거나 수축될 수 있다. 위족은 운동, 고정, 배설, 각 생성 및 규조류나 박테리아와 같은 작은 유기체를 포획하는 데 사용된다.[35][33]
각 외에도 유공충 세포는 미세소관의 세포골격에 의해 지지되는데, 이는 다른 아메바류에서 볼 수 있는 구조 없이 느슨하게 배열되어 있다. 유공충은 길쭉한 위족의 빠른 형성과 수축을 허용하는 미세소관을 빠르게 조립하고 분해하는 특수한 세포 메커니즘을 진화시켰다.[24]
배우체(유성형)에서 유공충은 일반적으로 핵이 하나뿐이지만, 무성생식체(무성형)는 여러 개의 핵을 가지는 경향이 있다. 적어도 일부 종에서는 핵이 이형성이며, 체세포핵은 생식핵보다 3배나 많은 단백질과 RNA를 포함하고 있다. 그러나 핵 해부학은 매우 다양하다.[36] 핵은 다방 유공충에서 반드시 하나의 방에 국한되지 않으며, 구형이거나 많은 엽을 가질 수 있다. 핵은 일반적으로 직경이 30~50 μm이다.[37]
일부 유공충 종은 세포 내에 크고 빈 공포를 가지고 있는데, 정확한 목적은 불분명하지만 질산염 저장소 역할을 하는 것으로 추정된다.[37]
미토콘드리아는 세포 전체에 고르게 분포되어 있지만, 일부 종에서는 기공 아래와 세포 바깥쪽 가장자리에 집중되어 저산소 환경에 적응한 것으로 추정된다.[37]
유공충류의 여러 종에서 세포 내에 비정상적으로 높은 농도의 방사성 동위원소가 발견되었는데, 이는 모든 진핵생물 중에서 가장 높은 수치에 속한다. 이 현상의 목적은 아직 밝혀지지 않았다.[38]
유전자 연구에 따르면, 나출된 아메바 ''Reticulomyxa''와 특이한 크세노피오포라가 껍데기가 없는 유공충으로 확인되었다. 다른 몇몇 아메바류는 그물 모양 위족을 생성하며, 이전에는 유공충과 함께 그라눌로레티쿨로사(Granuloreticulosa)로 분류되었지만, 이것은 더 이상 자연 그룹으로 간주되지 않으며, 대부분은 현재 세르코조아(Cercozoa)로 분류된다.[60]
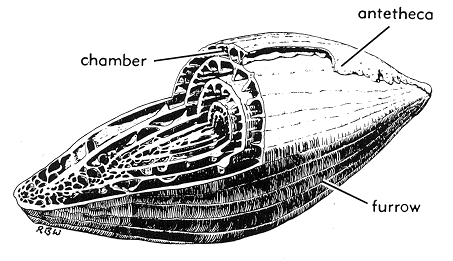
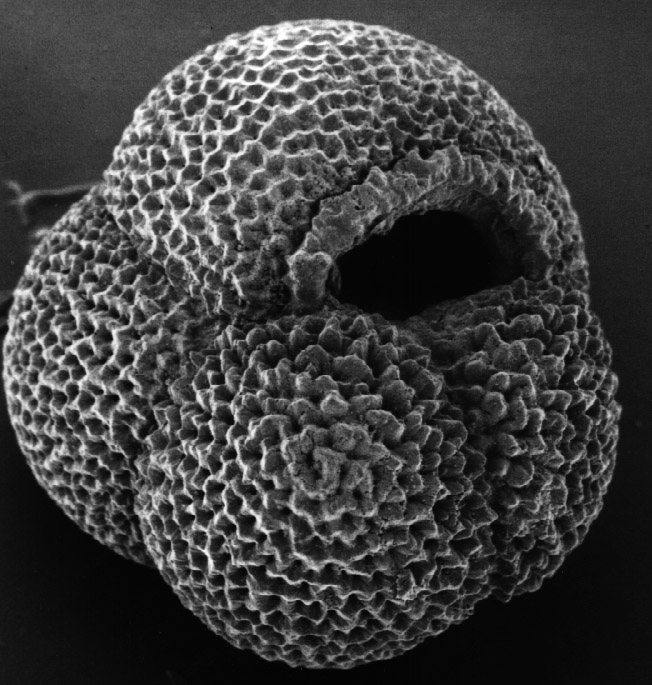
3. 1. 껍데기 (각, Test)
유공충의 껍데기는 내부의 생물체를 보호하는 역할을 한다. 다른 원생생물에 비해 일반적으로 단단하고 내구성이 강하기 때문에(비교적), 유공충 껍데기는 이 그룹에 대한 과학적 지식의 주요 원천이다.세포질이 외부로 뻗어나갈 수 있도록 껍데기에 있는 구멍을 구멍(aperture)이라고 한다.[56] 외부로 이어지는 ''주요'' 구멍(primary aperture)은 종에 따라 둥근 모양, 초승달 모양, 틈새 모양, 후드 모양, 방사상(별 모양), 수지상(나뭇가지 모양) 등 다양한 모양을 가진다. 어떤 유공충은 "톱니 모양", 플랜지 모양, 또는 입술 모양의 주요 구멍을 가지고 있다. 주요 구멍은 하나일 수도 있고 여러 개일 수도 있으며, 여러 개가 있는 경우에는 무리지어 있거나 적도상에 있을 수 있다. 주요 구멍 외에도 많은 유공충은 ''보조'' 구멍(supplemental apertures)을 가지고 있다. 이러한 구멍은 과거의 주요 구멍(이전 성장 단계의 주요 구멍)으로 남아 있거나 독특한 구조로 형성될 수 있다.
껍데기의 모양은 유공충 종류에 따라 매우 다양하며, 단방(unilocular) 또는 다방(multilocular)일 수 있다. 다방 형태에서는 생물체가 성장함에 따라 새로운 방이 추가된다. 단방 및 다방 형태 모두에서 나선형, 직렬형, 밀리올린형 등 다양한 껍데기 형태가 발견된다.[33]
많은 유공충은 껍데기에서 거대구형(megalospheric)과 미소구형(microspheric) 개체를 보이는 이형현상을 나타낸다. 이러한 이름은 전체 유기체의 크기를 가리키는 것이 아니라 첫 번째 방 또는 ''초시방(proloculus)''의 크기를 가리킨다. 화석으로서의 껍데기는 에디아카라기부터 알려져 있으며,[57] 많은 해양 퇴적물은 주로 이것으로 구성되어 있다. 예를 들어, 이집트 피라미드를 구성하는 석회암은 거의 전적으로 누물리트 저서성 유공충으로 구성되어 있다.[58] 산호초 유공충이 매년 약 4300만ton의 탄산칼슘을 생성하는 것으로 추정된다.[59]
껍질은 많은 방(chamber)으로 나뉘어 있는 것이 많고, 매우 정교한 구조가 되는 것도 있다. 이것은 '''초실'''(proloculus)을 핵으로 하여, 성장함에 따라 외측에 방을 만들어내기 때문이며, 그 전체 형태는 종종 소라껍질과 비슷하다. 껍질에는 구멍(aperture)이나 미세한 수많은 구멍이 있으며, 이를 통해 외부로 위족을 내민다.
3. 2. 세포 구조
유공충 세포의 가장 두드러진 특징은 단단한 껍질, 즉 각(殼)이다. 각은 하나 또는 여러 개의 방으로 구성되며, 단백질, 퇴적물 입자, 방해석, 아라고나이트, 또는 규산으로 구성될 수 있다.[24] 일부 유공충은 각이 전혀 없다.[32] 연체동물이나 산호와 달리 유공충의 각은 세포막 내부, 원형질 내부에 위치한다. 세포소기관은 각의 구획 내에 위치하며, 각의 구멍을 통해 위족에서 내부 세포로, 또는 그 반대로 물질이 이동할 수 있다.[33]유공충 세포는 과립성 내형질과 투명한 외형질로 나뉘며, 여기서 위족 그물이 단일 개구부 또는 각의 많은 구멍을 통해 나타난다. 개별 위족은 양방향으로 흐르는 작은 과립을 가진다.[34] 유공충은 ''과립망상 위족''을 가지는 것이 특징이며, 현미경으로 보면 위족이 과립으로 보인다. 위족은 길쭉하고 서로 분리되었다가 다시 합쳐질 수 있으며, 세포의 필요에 따라 늘어나거나 줄어든다. 위족은 운동, 고정, 배설, 각 생성 및 작은 유기체(예: 규조류, 박테리아) 포획에 사용된다.[35][33]
각 외에도 유공충 세포는 미세소관의 세포골격에 의해 지지된다. 유공충은 미세소관을 빠르게 조립하고 분해하는 특수한 세포 메커니즘을 진화시켜 위족을 빠르게 형성하고 수축시킨다.[24]
유성생식체(배우체)는 보통 핵이 하나뿐이지만, 무성생식체는 여러 개의 핵을 가진다. 일부 종에서는 핵이 이형성이며, 체세포핵은 생식핵보다 3배 많은 단백질과 RNA를 포함한다. 그러나 핵 해부학은 매우 다양하다.[36] 핵은 다방 유공충에서 반드시 하나의 방에 국한되지 않으며, 구형이거나 많은 엽을 가질 수 있다. 핵은 일반적으로 직경이 30~50 μm이다.[37]
일부 유공충 종은 세포 내에 크고 빈 공포를 가지고 있는데, 정확한 목적은 불분명하지만 질산염 저장소 역할을 하는 것으로 추정된다.[37]
미토콘드리아는 세포 전체에 고르게 분포하지만, 일부 종에서는 기공 아래와 세포 바깥쪽 가장자리에 집중되어 저산소 환경에 적응한 것으로 보인다.[37]
유공충류의 여러 종에서 세포 내에 비정상적으로 높은 농도의 방사성 동위원소가 발견되었는데, 이는 모든 진핵생물 중에서 가장 높은 수치에 속한다. 그 목적은 알려져 있지 않다.[38]
세포는 과립을 포함하는 내질과 투명한 외질로 나뉜다. 위족은 실 모양이며, 가지를 치거나 융합하여 그물 모양이 되고, 그 속을 과립이 양방향으로 흐른다. 이것은 이동, 부착, 포식 등에 사용된다. 세포내 공생체로서 녹조류, 홍조류, 황금색조류, 규조류, 와편모조류 등 다양한 조류를 가지고 있는 것이 상당히 많다. 또한 클렙토플라스트, 즉 섭취한 조류의 엽록체를 남겨 광합성을 하는 것도 있다.
껍질은 많은 방(chamber)으로 나뉘어 있는 것이 많고, 매우 정교한 구조가 되는 것도 있다. 이것은 '''초실'''(proloculus)을 핵으로 하여, 성장함에 따라 외측에 방을 만들어내기 때문이며, 그 전체 형태는 종종 소라껍질과 비슷하다. 껍질에는 구멍(aperture)이나 미세한 수많은 구멍이 있으며, 이를 통해 외부로 위족을 내민다.
4. 생태
대부분의 플랑크톤성 유공충은 로탈리아목 내의 글로비게리니나에서 발견된다.[22] 그러나 다른 현생 로탈리아목 계통인 ''네오갈리텔리아''는 독립적으로 플랑크톤성 생활 방식을 진화시킨 것으로 보인다.[41][42] 일부 쥐라기 화석 유공충도 독립적으로 플랑크톤성 생활 방식을 진화시켰고, 로베르티니다목의 구성원일 수 있다는 제안도 있다.[43]
유공충은 무척추동물, 어류, 해안 조류, 다른 유공충 등 다양한 생물에게 먹힌다. 포식자가 유공충 자체보다 껍질의 칼슘을 노리는 경우도 있을 수 있다. 몇몇 수생 달팽이는 특정 종의 유공충을 선호하는 것으로 알려져 있다.[50]
유공충의 생활환에는 단상과 복상의 세대교번이 있지만, 두 세대의 외형은 매우 비슷하다. 단상 가모트는 단핵이며, 세포 분열하여 보통 편모 두 개를 가진 배우자를 많이 만든다. 편모가 없는 아메바 형태의 배우자를 생성하는 종도 있다. 복상 시존트는 다핵체이며, 감수분열 후 분열하여 새로운 가모트를 만든다. 생식 세대 사이에 여러 번 무성생식을 하는 경우도 드물지 않다.[104]
4. 1. 서식지
현생 유공충은 주로 해양 생물이지만, 기수, 담수[34], 심지어 육상 서식지에서도 발견된다.[7] 대부분의 종은 저서성이며, 50종 이상의 형태종은 플랑크톤성이다.[35] 그러나 많은 유전적으로 구별되는 종들이 형태학적으로 구별이 안 될 수 있으므로, 이 수치는 실제 다양성의 일부만을 나타낼 수 있다.[40]저서성 유공충은 일반적으로 미세 입자의 퇴적물에서 발견되며, 층 사이를 활발하게 이동한다. 그러나 많은 종들이 암석 기질, 해조류에 부착되거나, 퇴적물 표면 위에 서식한다.[24]
현생 유공충은 기본적으로 해산이지만 기수에서도 생존이 가능하다. 레만 호 등에는 담수종도 몇 종 있으며, 열대 다우림의 습한 토양에서 생활하는 종도 있다. 바다에서는 얕은 해저의 해조류 뿌리 근처에 부착하여 생활하고 있으며, 예를 들어 오키나와 기념품으로 유명한 별모래는 유공충의 일종인 호시즈나 (''Baculogypsina sphaerulata'')의 껍질이지만, 그러한 장소를 찾으면 살아있는 것을 발견할 수 있다. (참고로 "태양의 모래"라고도 불리는, 돌기 끝이 둥근 별모래는 ''Calcarina'' 속의 유공충이다.) 더욱이 심해 퇴적물 표층에도 유공충이 다수 서식하고 있는 것이 밝혀졌다.[103] 이와 같이 해저에 사는 것을 '''저서성 유공충'''(Benthic foraminifera)이라고 하며, 현생 유공충의 대부분이 여기에 해당한다.
4. 2. 섭식
유공충은 주로 해양 생물이지만, 기수, 담수,[34] 심지어 육상 서식지에서도 발견된다.[7] 대부분의 종은 저서성이며, 50종 이상의 형태종은 플랑크톤성이다.[35] 그러나 많은 유전적으로 구별되는 종들이 형태학적으로 구별이 안 될 수 있으므로, 이 수치는 실제 다양성의 일부만을 나타낼 수 있다.[40]대부분의 유공충은 종속영양성이며, 더 작은 유기체와 유기물을 섭취한다. 일부 작은 종들은 식물 부유물을 전문적으로 섭취하는 반면, 다른 종들은 규조류 섭취를 전문으로 한다. 일부 저서성 유공충은 위족을 사용하여 퇴적물과 유기 입자 내부에 스스로를 낭포화하는 먹이 낭포를 만든다.[24] 특정 유공충은 요각류 또는 쿠마세아류와 같은 작은 동물을 포식하며, 일부 유공충은 먹이의 각피에 구멍을 뚫어 다른 유공충을 포식하기도 한다.[48] 현탁물 여과 섭식 또한 이 그룹에서 흔하며, 적어도 일부 종들은 용존 유기 탄소를 이용할 수 있다.[24]
몇몇 유공충 종은 기생하며, 해면, 연체동물, 산호, 또는 다른 유공충을 감염시킨다. 기생 전략은 다양하며, 일부는 외기생으로 작용하여 위족을 사용하여 숙주로부터 먹이를 훔치는 반면, 다른 종들은 숙주의 껍질이나 체벽을 통해 파고 들어 부드러운 조직을 먹는다.[24]
저서성 및 플랑크톤성[44][45] 유공충 중 다수는 녹조류, 홍조류, 황금색조류, 규조류, 디노플라젤라류 등 다양한 계통의 단세포 조류를 내공생체로 가지고 있다.[35] 이러한 혼합영양성 유공충은 영양분이 부족한 대양 수역에서 특히 흔하다.[46] 일부 유공충은 클렙토플라스틱이며, 섭취한 조류로부터 엽록체를 보유하여 광합성을 수행한다.[47]
4. 3. 공생
현생 유공충은 주로 바다에서 살지만, 민물과 섞인 기수(汽水)나 담수,[34] 심지어 육지에서도 발견된다.[7] 대부분의 종은 바닥에서 살아가며, 50종 이상의 형태종은 플랑크톤처럼 물에 떠다닌다.[35] 하지만, 유전적으로는 다양하지만 겉모습은 구별하기 어려운 종들이 많아서, 실제 다양성은 훨씬 더 클 수 있다.[40]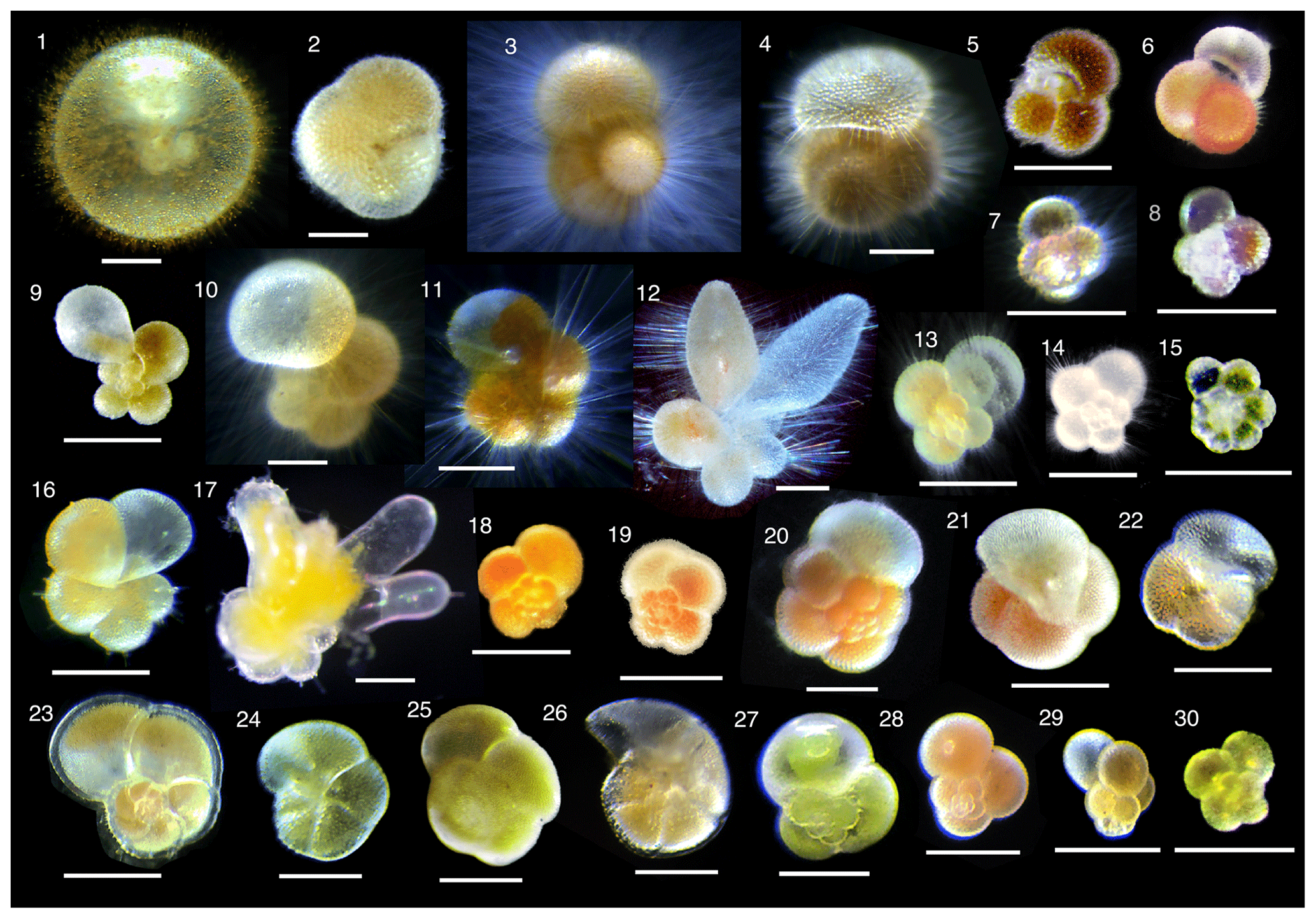
바닥에 사는 유공충은 보통 미세한 알갱이로 된 퇴적물 속에서 발견되며, 퇴적물 층 사이를 활발하게 이동한다. 하지만, 많은 종들이 암석, 해조류에 붙어 살거나, 퇴적물 표면 위에서 살기도 한다.[24]
많은 저서성 및 플랑크톤성 유공충들은[44][45] 녹조류, 홍조류, 황금색조류, 규조류, 디노플라젤라류 등 다양한 종류의 단세포 조류와 공생 관계를 맺는다.[35] 이러한 혼합영양성 유공충은 영양분이 부족한 바다에서 특히 흔하다.[46] 일부 유공충은 클렙토플라스틱 방식으로, 먹이인 조류의 엽록체를 빼앗아 광합성을 하기도 한다.[47]
4. 4. 극한 환경
현생 유공충은 주로 해양 생물이지만, 기수, 담수[34], 심지어 육상 서식지[7]에서도 발견된다. 일부 저서성 유공충은 24시간 이상 무산소성 조건에서 생존할 수 있는데, 이는 퇴적물-수계 경계면 근처의 변화하는 산소 조건에 적응한 결과로 해석된다.[51]
유공충은 마리아나 해구의 챌린저 해연과 같이 가장 깊은 해양 지역에서도 발견된다. 이러한 심해에서는 탄산염 보상 심도 아래에 위치하여 껍질의 탄산칼슘이 극심한 압력으로 인해 물에 용해된다. 따라서 챌린저 해연에서 발견되는 유공충은 탄산염 껍질 대신 유기물 껍질을 가지고 있다.[52]
비해양 유공충은 전통적으로 유공충 연구에서 간과되었으나, 최근 연구 결과 이전에 알려진 것보다 훨씬 다양하다는 사실이 밝혀졌다. 이들은 이끼, 강, 호수, 연못, 습지, 토양, 이탄 늪, 사구 등 다양한 생태적 틈새에 서식한다.[53]
5. 생식
유공충의 일반적인 생활사는 반수체와 이배체 세대가 번갈아 나타나는 형태이다. 하지만 두 세대의 외형은 대부분 유사하다.[23][54] 반수체(감수체, gamont)는 하나의 핵을 가지며, 분열하여 여러 생식세포를 만든다. 이 생식세포는 보통 두 개의 편모를 가진다. 이배체(무성생식체, agamont)는 다핵체이며, 감수분열 후 분열하여 새로운 감수체를 만든다. 저서성 유공충은 성적 세대 사이에 여러 번 무성생식을 하기도 한다.[34]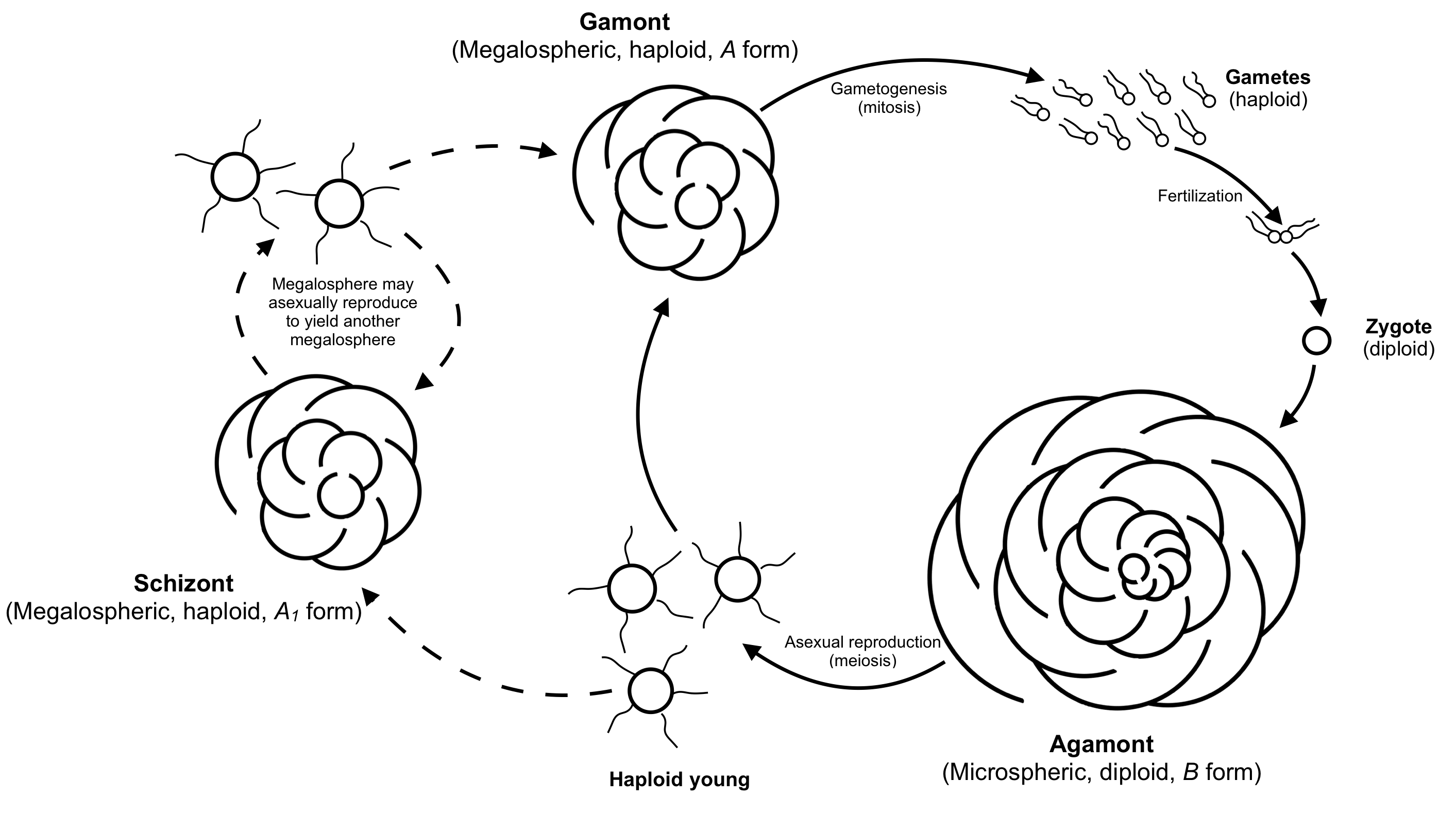
현생 유공충은 기본적으로 바다에 살지만, 기수에서도 생존이 가능하다. 레만호 등에는 담수종도 몇 종 있으며, 열대 다우림의 습한 토양에서 생활하는 종도 있다. 바다에서는 얕은 해저의 해조류 뿌리 근처에 부착하여 생활하고 있으며, 오키나와 기념품으로 유명한 별모래는 유공충의 일종인 호시즈나(''Baculogypsina sphaerulata'')의 껍질이다. 그러한 장소를 찾으면 살아있는 호시즈나를 발견할 수 있다. 참고로 "태양의 모래"라고도 불리는, 돌기 끝이 둥근 별모래는 ''Calcarina'' 속의 유공충이다. 심해 퇴적물 표층에도 유공충이 다수 서식하고 있는 것으로 밝혀졌다.[103]
5. 1. 세대교번
일반적인 유공충의 생활사는 반수체와 이배체 세대가 번갈아 나타나는 세대교번을 보이지만, 대부분 형태는 유사하다.[23][54] 반수체 또는 감수체(gamont)는 초기에는 단일 핵을 가지고 있으며, 여러 개의 생식세포를 생성하기 위해 분열하는데, 이 생식세포는 일반적으로 두 개의 편모를 갖는다. 이배체 또는 무성생식체(agamont)는 다핵체이며, 감수분열 후 새로운 감수체를 생성하기 위해 분열한다. 저서성 종에서는 성적 세대 사이에 여러 번의 무성생식이 드물지 않게 일어난다.[34]유공충은 생식주기와 관련된 형태적 이형 현상을 보인다. 감수체 또는 성적으로 번식하는 반수체 형태는 '거대구체형(megalospheric)'인데, 그 '원구(proloculus)' 또는 첫 번째 방이 비례적으로 크다. 감수체는 'A'형으로도 알려져 있다. 감수체는 일반적으로 원구가 더 크지만, 무성생식체보다 전체적인 껍질 지름이 일반적으로 더 작다.
성숙기에 도달하면 감수체는 유사분열을 통해 수천 개의 생식세포를 생성하는데, 이들도 반수체이다. 이러한 생식세포는 모두 완전한 세포소기관 세트를 가지고 있으며, 껍질을 손상시키지 않고 환경으로 배출된다. 생식세포는 정자와 난자로 분화되지 않으며, 종의 어떤 두 개의 생식세포도 일반적으로 서로 수정할 수 있다.
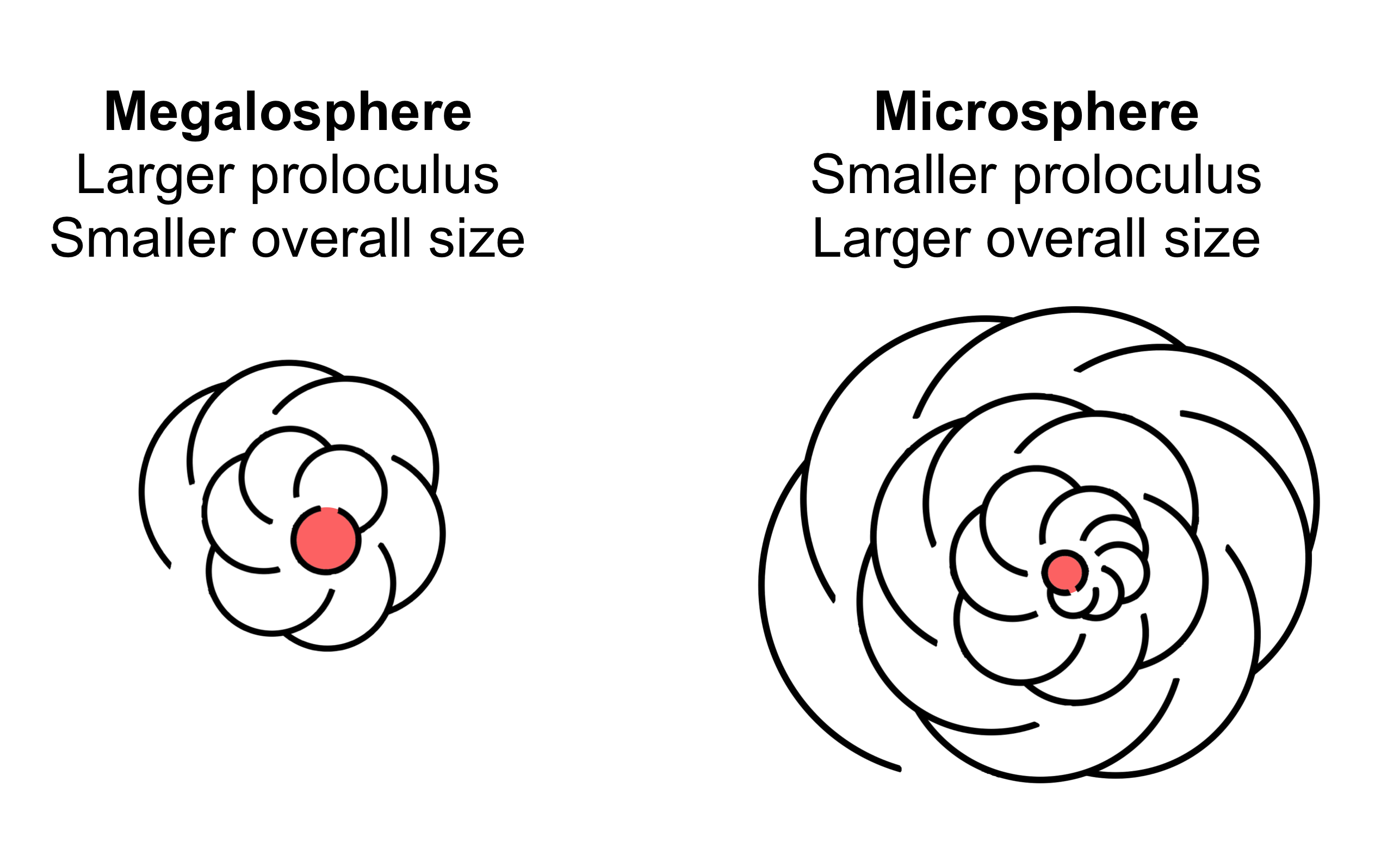
두 개의 생식세포가 결합하면 '무성생식체(agamont)' 또는 'B'형으로 알려진 이배체 다핵 세포를 생성한다. 감수체와는 달리, 무성생식체는 '미소구체형(microspheric)'이며, 비례적으로 작은 첫 번째 방을 가지지만 일반적으로 더 많은 방을 가진 더 큰 전체 지름을 갖는다. 무성생식체는 유공충의 무성생식 단계이다. 성체에 도달하면 원형질이 껍질을 완전히 비우고 세포질을 감수적으로 다분열을 통해 여러 개의 반수체 자손을 형성한다. 이 자손들은 산포되기 전에 거대구체형의 첫 번째 방을 형성하기 시작한다.
어떤 경우에는 반수체 어린 유공충이 거대구체형으로 성숙하여 무성생식을 통해 또 다른 거대구체형 반수체 자손을 생성할 수 있다. 이 경우, 첫 번째 거대구체형은 '분열체(schizont)' 또는 'A₁'형이라고 하며, 두 번째는 감수체 또는 'A₂'형이라고 한다.

성숙과 생식은 더 차갑고 깊은 물에서 더 느리게 일어나는데, 이러한 조건은 유공충이 더 크게 자라게 한다. 'A'형은 두 개의 생식세포가 만나 성공적으로 결합할 가능성이 줄어들기 때문에 'B'형보다 항상 훨씬 더 많은 것으로 보인다.[55][33]
5. 2. 형태적 이형성
유공충은 생식주기에 따라 형태가 달라지는 특징을 보인다. 감수체(gamont) 또는 반수체(haploid) 형태는 '거대구체형(megalospheric)'이라고 불리며, 첫 번째 방인 '원구(proloculus)'가 상대적으로 크다. 이를 'A'형이라고도 부른다.[55] 감수체는 원구가 크지만, 무성생식체에 비해 전체 껍질 지름은 보통 더 작다.감수체는 성숙하면 유사분열을 통해 수천 개의 반수체 생식세포를 만든다. 이 생식세포들은 모두 완전한 세포소기관을 갖추고 있으며, 껍질을 손상시키지 않고 외부로 방출된다. 생식세포는 정자와 난자로 분화되지 않고, 같은 종의 생식세포라면 일반적으로 서로 수정할 수 있다.[55]
두 생식세포가 결합하면 '무성생식체(agamont)' 또는 'B'형이라 불리는 이배체(diploid) 다핵 세포가 된다. 무성생식체는 감수체와 달리 '미소구체형(microspheric)'이며, 원구가 작지만 방의 개수가 많아 전체 지름은 보통 더 크다. 무성생식체는 유공충의 무성생식 단계이다. 성체가 되면 원형질이 껍질을 비우고 세포질이 감수 다분열을 통해 여러 개의 반수체 자손을 형성한다. 이들은 흩어지기 전에 거대구체형의 첫 번째 방을 만들기 시작한다.[55]
어떤 경우, 반수체 어린 유공충이 거대구체형으로 성숙하여 무성생식을 통해 또 다른 거대구체형 반수체 자손을 낳기도 한다. 이때 처음 거대구체형은 '분열체(schizont)' 또는 'A₁'형, 두 번째는 감수체 또는 'A₂'형이라고 한다.[55]
성숙과 생식은 더 차갑고 깊은 물에서 더 느리게 진행되며, 이는 유공충이 더 크게 자라는 조건이 된다. 'A'형은 두 생식세포가 만나 수정될 확률이 낮기 때문에 'B'형보다 항상 훨씬 많다.[55][33]
5. 3. 생식 전략
일반적으로 유공충의 생활사는 반수체와 이배체 세대가 번갈아 나타나는 세대교번을 보이지만, 대부분 형태는 유사하다.[23][54] 반수체 또는 감수체(gamont)는 처음에 하나의 핵을 가지며, 분열하여 여러 생식세포를 만든다. 이 생식세포는 보통 두 개의 편모를 갖는다. 이배체 또는 무성생식체(agamont)는 다핵체이며, 감수분열 후 분열하여 새로운 감수체를 생성한다. 저서성 유공충의 경우 성적 세대 사이에 여러 번의 무성생식이 드물지 않게 일어난다.[34]유공충은 생식주기와 관련하여 형태적 차이를 보인다. 감수체 또는 성적으로 번식하는 반수체 형태는 '거대구체형(megalospheric)'인데, 이는 첫 번째 방인 '원구(proloculus)'가 상대적으로 크다는 것을 의미한다. 감수체는 'A'형으로도 불린다. 감수체는 원구가 더 크지만, 무성생식체보다 전체적인 껍질 지름은 보통 더 작다.
성숙기에 도달하면 감수체는 유사분열을 통해 수천 개의 반수체 생식세포를 만든다. 이 생식세포들은 모두 완전한 세포소기관을 가지며, 껍질을 손상시키지 않고 외부로 배출된다. 생식세포는 정자와 난자로 분화되지 않으며, 종에 따라 다르지만 일반적으로 두 개의 생식세포가 서로 수정할 수 있다.
두 개의 생식세포가 결합하면 '무성생식체(agamont)' 또는 'B'형으로 알려진 이배체 다핵 세포가 만들어진다. 감수체와 달리, 무성생식체는 '미소구체형(microspheric)'이며, 상대적으로 작은 첫 번째 방을 가지지만, 일반적으로 더 많은 방을 가진 더 큰 전체 지름을 갖는다. 무성생식체는 유공충의 무성생식 단계이다. 성체가 되면 원형질이 껍질을 완전히 비우고 세포질이 감수적으로 다분열하여 여러 개의 반수체 자손을 형성한다. 이 자손들은 흩어지기 전에 거대구체형의 첫 번째 방을 만들기 시작한다.
어떤 경우에는 반수체 어린 유공충이 거대구체형으로 성숙하여 무성생식을 통해 또 다른 거대구체형 반수체 자손을 생성할 수 있다. 이 경우, 첫 번째 거대구체형은 '분열체(schizont)' 또는 'A₁'형이라고 하며, 두 번째는 감수체 또는 'A₂'형이라고 한다.
성숙과 생식은 더 차갑고 깊은 물에서 더 느리게 진행되는데, 이러한 조건은 유공충이 더 크게 자라도록 유도한다. 'A'형은 두 개의 생식세포가 만나 성공적으로 결합할 가능성이 낮기 때문에 'B'형보다 항상 훨씬 더 많이 발견된다.[55][33]
유공충 그룹에 따라 다양한 생식 전략이 나타난다.
단극종에서는 A형과 B형이 존재한다. 다극 유공충의 미소구형과 마찬가지로, 무성생식을 하는 B형이 유성생식을 하는 A형보다 크다.
나선유공충과에 속하는 유공충은 편모 대신 아메바 형태의 배우자를 갖는다. 이 그룹의 다른 생식 방식은 일반적으로 다른 유공충 그룹과 유사하다.
석회질 나선유공충류인 ''Patellina corrugata''는 대부분의 다른 유공충과 약간 다른 생식 전략을 보인다. 무성생식을 하는 B형은 세포 전체를 둘러싸는 낭을 생성하고, 그 안에서 분열하여 어린 세포가 모세포의 각질 껍질을 섭취하여 자신의 각질 껍질의 첫 번째 방을 형성한다. 성숙한 A형은 최대 9개체까지 그룹을 이루고, 그 주위에 보호용 낭을 형성한다. 배우자 형성은 이 낭 안에서 일어나며, 매우 적은 수의 배우자가 생성된다. B형 유생은 낭 안에서 생성되며, 세포로 결합되지 않은 핵은 발달하는 유생의 먹이로 사용된다. A형 ''Patellina''는 자웅이주이며, "플러스"와 "마이너스"로 불리는 성이 존재한다. 이 성들은 핵의 수가 다르며, "플러스"형은 3개의 핵을, "마이너스"형은 4개의 핵을 가지고 있다. B형은 다시 A형보다 크다.[33][55][48]
6. 분류
유공충은 1854년 슐체(Schultze)가 유공충목(Foraminiferida)으로 처음 언급한 이후[23] 다양한 분류 체계를 거쳐왔다. 1992년 로블리히와 태펀은 유공충을 현재와 같이 강(Class)으로 재분류했다.[24]
전통적으로 유공충은 껍질의 형태와 구성에 따라 분류되었다. 탄산칼슘으로 이루어진 석회질 껍질은 다시 투명한 유리질과 불투명한 자기질로 나뉜다. 유기질 껍질이나 퇴적물 파편을 접착한 사질 껍질을 가진 유공충도 있으며, 규산질 껍질을 가진 속도 하나 존재한다. 그러나 이러한 껍질 구성에 따른 분류는 실제 계통을 반영하지 못하는 것으로 밝혀졌다.
분자계통 분석 결과, 유공충은 리자리아(Rhizaria)에 속하며[2], 켈코조아(Cercozoa)와 방산충과 가깝지만 이들과의 계통적 관계는 아직 명확하지 않다. 과거에는 원생동물 근족류로 분류되기도 했다.[107][108]
아래는 2019년 리자리아의 계통 분류이다.[111]
- Globothalamea
- * 카르테리나목 (Carterinida)
- * 로베르티나목 (Robertinida)
- * 수레바퀴유공충목 (Rotaliida)
- * 애직공유공충아강 (Textulariia)
- ** 나팔유공충목 (Lituolida)
상처유공충아목 (Hormosinina)
나팔유공충아목 (Lituolina)
Nezzazatina
Rzehakinina
Spiroplectamminina
바퀴매듭유공충아목 (Trochamminina)
베르네우일리나아목 (Verneuilinina)
- ** 로프투시아목 (Loftusiida)
- ** 애직공유공충목 (Textulariida)
- Monothalamea
- * 알로그로미아목 (Allogromiida)
- * 아스트로리자목 (Astrorhizida)
- * 미분류 목
- ** Xenophyophoroidea
- ** 미분류 상과
미분류 과
* ''Haplomyxa''
* ''Nellya''
* ''Reticulomyxa''
- Tubothalamea
- * 기장구멍벌레목 (Miliolida)
- * 스피릴리나목 (Spirillinida)
- 미분류 강
- * 병유공충목 (Lagenida)
6. 1. 껍질 구성에 따른 분류 (전통적 분류)
껍질의 형태와 구성은 유공충 분류 및 동정의 기본적인 수단이다. 탄산칼슘을 주성분으로 하는 석회질(calcareous) 껍질을 가진 것이 많으며, 이는 다시 투명한 유리질(hyaline)과 불투명한 자기질(porcelaneous)로 나뉜다. 유기질(또는 의사키틴질, pseudochitinious) 껍질이나 퇴적물 파편을 접착하여 만들어지는 사질(또는 교착질, agglutinated) 껍질을 가진 유공충도 있으며, 규산질 껍질을 가진 속도 하나 있다. 그러나 이러한 껍질 구성을 이용한 분류 체계는 자연 분류가 아닌 것으로 밝혀졌다.[107][108]6. 2. 분자 계통 분류 (최근 분류)
2019년 리자리아의 계통 분류는 다음과 같다.[111]분자계통 분석 결과, 유공충은 리자리아(Rhizaria)에 속하며[2], 켈코조아(Cercozoa)와 방산충과 가깝지만 이들과의 계통적 관계는 아직 불명확한 부분이 있다. 과거에는 원생동물 근족류로 분류되기도 했다.[107][108]
6. 3. 계통 분류
유공충의 계통 분류는 1854년 슐체(Schultze)가 유공충목(Foraminiferida)으로 언급한 이후 다양하게 변화해 왔다.[23] 1992년 로블리히와 태펀은 유공충을 강(Class)으로 재분류했다.[24]유공충은 일반적으로 원생동물[25][26][27] 또는 프로티스타[28][29]에 포함된다. 분자 계통 발생학적 증거에 따르면 유공충은 리자리아(Rhizaria)에 속한다.[2] 리자리아는 분류 계급상 문(Phylum)과 같은 용어를 사용하지 않아 문제가 있지만, 카발리어-스미스는 리자리아를 원생동물계 내의 아계(Subkingdom)로 정의했다.[2]
일부 학자들은 유공충을 자체 문(Phylum)으로 분류하기도 한다. 분자 데이터에 따르면 유공충은 세르코조아(Cercozoa) 및 방산충류(Radiolaria)와 관련이 있으며, 이 세 그룹이 리자리아를 구성한다.[2] 그러나 정확한 관계는 아직 불분명하며, 피낭성 아메바와도 가깝다.[30]
2019년 리자리아의 계통 분류는 다음과 같다.[111]
- 사족충류
- * 필로사(Filosa)
- ** 레티쿨로필로사(Reticulofilosa)
- ** 모나도필로사(Monadofilosa)
- * 엔도믹사(Endomyxa)
- 유공충류
- * 유공충아문
- * 다공낭충류
- * 방사극충강(Acantharea)
2013년 Pawlowski 등의 유공충 계통 발생도는 다음과 같다.[22]
- "단세포 유공충류" (다계통군)
- * 유공충목(Foraminiferida)
- * 튜보탈라메아(Tubothalamea)
- ** 밀리올리다(Miliolida)
- ** 나선유공충목(Spirillinida)
- * "단세포 유공충류"
- ** 크세노피오포라(Xenophyophorea)
- * 글로보탈라메아(Globothalamea)
- ** "사각유공충목" (다계통군)
- ** 로베르티나목(Robertinida)
- ** 수레바퀴유공충목(Rotaliida)
분자계통 분석 결과, 껍질이 없는 유공충인 ''Reticulomyxa''와 크세노피오포라(Xenophyophorea)가 밝혀졌다. 과거 유공충과 함께 과립근족류(Granuloreticulosa)로 묶였던 망상 위족을 가진 아메바상 생물들은 대부분 세르코조아(Cercozoa)에 포함된다.
Mikhalevich (2013)의 분류[31]는 다음과 같다.
과거에는 원생동물 근족류로 분류되었으나, 2007년 현재는 리자리아(Rhizaria)에 포함된다.[107][108] 켈코조아(Cercozoa)와 방산충(Radiolaria)이 리자리아에 속하지만, 이들과 유공충의 계통적 관계는 불분명하다.
껍질 형태와 구성은 유공충 분류 및 동정의 기본 수단이다. 석회질 껍질은 투명한 유리질과 불투명한 자기질로 나뉜다. 유기질 또는 사질 껍질을 가진 유공충도 있으며, 규산질 껍질을 가진 속도 하나 있다. 그러나 이러한 껍질 구성을 이용한 분류는 자연 분류가 아닌 것으로 밝혀졌다.
7. 화석 기록
분자 시계 연구에 따르면 유공충의 현생종 분류군은 신원생대(9억 년에서 6억 5천만 년 전 사이)에 진화했을 가능성이 높다.[61][62] 이는 밀접하게 관련된 필로사(Filosa) 아메바의 신원생대 화석과도 일치한다. 에디아카라기 말기 이전의 유공충 화석은 발견되지 않았는데, 이는 이 시기 원생대 유공충 대부분이 단단한 껍질(test)을 가지지 않았기 때문일 가능성이 높다.[61][62] 무기질 껍질이 없는 알로그로미이목(Allogromiida)은 화석 기록이 없다.[61]
에디아카라기 시대의 신비로운 벤도조아(Vendozoa)가 화석 크세노피오포레아(Xenophyophorea)를 나타낸다는 제안이 있었지만,[63] ''Dickinsonia'' 유해와 관련된 C27 스테롤(sterol)의 발견으로 인해 이 주장은 의문을 갖게 되었고, 대신 동물일 가능성이 제기되었다.[64] 다른 연구자들은 흔적 화석 ''Paleodictyon''과 그 친척인 그라포글립티다이(Graphoglyptidae)가 화석 크세노피오포레아(xenophyophore)를 나타낼 수 있다고 제안했으며,[65] 현생 크세노피오포레아(xenophyophore) ''Occultammina''와의 유사성을 지적했다.[66] 그러나 현대의 ''Paleodictyon'' 사례는 이 문제를 해결하지 못했고, 이 흔적은 굴이나 해면동물(Hexactinellid)일 수도 있다.[67] 살아있는 크세노피오포레아(xenophyophores)의 서식지가 화석 그라포글립티다이(graphoglyptids)의 추정 서식지와 유사하다는 점이 이러한 견해를 뒷받침한다. 그러나 많은 그라포글립티다이(graphoglyptids)의 큰 크기와 규칙성, 그리고 그 화석에서 크세노피아(xenophyae)가 보이지 않는다는 점이 그 가능성에 의문을 제기한다.[66] 2017년 기준으로 확실한 크세노피오포레아(xenophyophore) 화석은 발견되지 않았다.[68]
껍질을 가지고 있기 때문에 캄브리아기 이후부터 화석으로 많이 발견되어 시상화석, 시준화석으로 중요하게 여겨진다. 고생대 석탄기의 푸줄리나(방추충)나 화폐석은 현재는 볼 수 없는 대형 종이다.[109]
7. 1. 주요 화석 유공충
껍질을 가진 유공충은 현생누대 전반에 걸쳐 우수한 화석 기록을 가지고 있다. 가장 오래된 확실한 유공충은 에디아카라기의 바로 끝 무렵 화석 기록에 나타난다. 이러한 형태는 모두 응집성 껍질을 가지고 있으며, 단실(unilocular)이다. 여기에는 ''Platysolenites''와 ''Spirosolenites''와 같은 형태가 포함된다.[69][57]
단실 유공충은 캄브리아기 전반에 걸쳐 다양화되었다. 일반적으로 발견되는 형태에는 ''Ammodiscus'', ''Glomospira'', ''Psammosphera'', ''Turritellella''가 포함된다. 이러한 종은 모두 응집성이다. 이들은 아모디스키나(Ammodiscina)의 일부를 구성하며, 이는 여전히 현생 형태를 포함하는 스피릴리니다(Spirillinida)의 계통이다.[70][22] 후기 스피릴리니다(spirillinids)는 다실(multilocularity)과 방해석 껍질을 진화시켰으며, 이러한 최초의 형태는 트라이아스기에 나타났다. 이 그룹은 K-Pg 멸종으로 인한 다양성에 거의 영향을 받지 않았다.[71]
가장 오래된 다실 유공충은 응집성 종이며, 중기 캄브리아기 시대의 화석 기록에 나타난다. 보존 상태가 좋지 않아 주요 유공충 그룹에 확실하게 배정할 수 없다.[70]
가장 오래된 석회질 벽을 가진 유공충은 푸줄리나(Fusulinida)이며, 초기 실루리아기의 란도베리절에 화석 기록에 나타난다. 이 중 가장 초기의 것은 현미경적이며, 평면 나선형으로 감겨 있고, 나선이 풀려 있다. 후기 형태는 렌즈형, 구형, 그리고 길쭉한 쌀알 모양의 다양한 형태를 진화시켰다.[72][73]
후기 푸줄리나(fusulinids) 종은 훨씬 더 큰 크기로 성장했으며, 일부 형태는 길이가 5cm에 달했다. 보고에 따르면 일부 표본은 최대 14cm에 달하며, 이는 현존하거나 멸종된 가장 큰 유공충 중 하나이다. 푸줄리나(Fusulinids)는 광합성 유기체와 공생 관계를 진화시킨 것으로 생각되는 가장 오래된 유공충 계통이다. 푸줄리나(fusulinids)의 화석은 남극 대륙을 제외한 모든 대륙에서 발견되었다. 이들은 석탄기의 비제절에 가장 큰 다양성에 도달했다. 그 후 그룹의 다양성은 점차 감소하여 페름기-트라이아스기 멸종 사건 동안 최종적으로 멸종되었다.[33][71][74]
석탄기의 투르네시절 동안, 밀리올리다(Miliolida) 유공충이 처음으로 화석 기록에 나타났으며, 투보탈라메아(Tubothalamea) 내에서 스피릴리니다(Spirillinida)에서 갈라져 나왔다. 밀리올리다(Miliolids)는 페름기-트라이아스기 및 K-Pg 멸종 모두에서 약 50%의 희생을 입었지만 오늘날까지 생존했다. 일부 화석 밀리올리다(miliolids)는 직경이 최대 2cm에 달했다.[71]
가장 오래된 라게니다(Lagenida) 화석은 석탄기의 모스코비절에 나타난다. 페름기-트라이아스기 또는 K-Pg 멸종으로 인한 영향이 거의 없었던 이 그룹은 시간이 지남에 따라 다양화되었다. 이차적으로 단실 분류군은 쥐라기와 백악기 동안 진화했다.
가장 오래된 인볼루티니다(Involutinida) 화석은 페름기 동안 나타난다. 이 계통은 유라시아의 중생대 전반에 걸쳐 다양화되었지만, 세노마눔-투로눔 경계 사건 이후 화석 기록에서 사라진 것으로 보인다. 현생 그룹 플라니스피릴리니다이(planispirillinidae)는 인볼루티니다(involutinida)로 언급되었지만, 이는 여전히 논쟁의 여지가 있다.[75][71]
로베르티니다(Robertinida)는 트라이아스기의 아니시절 동안 화석 기록에 처음 나타난다. 이 그룹은 화석 역사 전반에 걸쳐 낮은 다양성을 유지했다. 모든 살아있는 대표는 팔레오세에 처음 나타난 로베르티니다이(Robertinidae)에 속한다.[71]
가장 오래된 확실한 로탈리다(Rotaliida) 화석은 트라이아스기-쥐라기 멸종 사건 이후 쥐라기의 플리엔스바키안절까지 화석 기록에 나타나지 않는다.[76] 이 그룹의 다양성은 세노마눔-투로눔 사건 이후까지 낮게 유지되었으며, 그 후 이 그룹은 급속한 다양화를 보였다. 이 그룹 중 플랑크톤성 글로비게리나(Globigerinina)—가장 오래된 플랑크톤성 유공충 그룹—는 토아르시안 전환 이후에 처음 나타났다. 이 그룹은 K-Pg 멸종과 에오세-올리고세 멸종 사건 모두에서 큰 손실을 보았지만, 오늘날까지 현존하고 다양하다.[71] 플랑크톤성 생활 방식의 추가 진화는 마이오세 또는 플라이오세에 발생했는데, 로탈리다(rotaliid) ''Neogallitellia''가 독립적으로 플랑크톤성 생활 방식을 진화시켰다.[41][42]
껍질을 가지고 있기 때문에 캄브리아기 이후부터 화석으로 많이 발견되어 시상화석, 시준화석으로 중요하게 여겨진다. 고생대 석탄기의 푸줄리나(방추충)나 화폐석은 현재는 볼 수 없는 대형 종이다. 그 외에도 ''Dentalina'', ''Endothyra'', ''Globigerina'', ''Hyperammina'', ''Triticites'' 등이 있다. 저서성 유공충은 캄브리아기 이전에 출현하였고, 부유성 유공충이 출현한 것은 쥐라기 이후이다.[109]
7. 2. 진화 역사
분자 시계 연구에 따르면 유공충의 현생종 분류군은 신원생대(9억 년에서 6억 5천만 년 전 사이)에 진화했을 가능성이 높습니다.[61][62] 이는 밀접하게 관련된 필로사(Filosa) 아메바의 신원생대 화석과도 일치합니다. 에디아카라기 말기 이전의 유공충 화석은 발견되지 않았는데, 이는 이 시기 원생대 유공충 대부분이 단단한 껍질(test)을 가지지 않았기 때문일 가능성이 높습니다.[61][62] 무기질 껍질이 없는 알로그로미이목(Allogromiida)은 화석 기록이 없습니다.[61]에디아카라기 시대의 벤도조아(Vendozoa)가 화석 크세노피오포레아(Xenophyophorea)를 대표한다는 주장이 있었지만,[63] ''Dickinsonia'' 유해와 관련된 C27 스테롤(sterol)의 발견으로 인해 이 주장은 의문을 갖게 되었고, 대신 동물일 가능성이 제기되었습니다.[64] 다른 연구자들은 흔적 화석 ''Paleodictyon''과 그 친척인 그라포글립티다이(Graphoglyptidae)가 화석 크세노피오포레아(xenophyophore)를 나타낼 수 있다고 제안했으며,[65] 현생 크세노피오포레아(xenophyophore) ''Occultammina''와의 유사성을 지적했습니다.[66] 그러나 현대의 ''Paleodictyon'' 사례는 이 문제를 해결하지 못했고, 이 흔적은 굴이나 해면동물(Hexactinellid)일 수도 있습니다.[67] 살아있는 크세노피오포레아(xenophyophores)의 서식지가 화석 그라포글립티다이(graphoglyptids)의 추정 서식지와 유사하다는 점이 이러한 견해를 뒷받침합니다. 그러나 많은 그라포글립티다이(graphoglyptids)의 큰 크기와 규칙성, 그리고 그 화석에서 크세노피아(xenophyae)가 보이지 않는다는 점이 그 가능성에 의문을 제기합니다.[66] 2017년 기준으로 확실한 크세노피오포레아(xenophyophore) 화석은 발견되지 않았습니다.[68]
껍질을 가진 유공충은 현생누대(Phanerozoic) 전반에 걸쳐 풍부한 화석 기록을 가지고 있습니다. 가장 오래된 유공충 화석은 에디아카라기 말기에 나타나며, 이들은 모두 응집성 껍질을 가진 단실(unilocular) 유공충입니다. 여기에는 ''Platysolenites''와 ''Spirosolenites''와 같은 형태가 포함됩니다.[69][57]
단실 유공충은 캄브리아기 동안 다양해졌으며, ''Ammodiscus'', ''Glomospira'', ''Psammosphera'', ''Turritellella''와 같은 종들이 흔하게 발견됩니다. 이들은 모두 응집성 껍질을 가지고 있으며, 아모디스키나(Ammodiscina)에 속합니다. 아모디스키나는 현생 형태를 포함하는 스피릴리니다(Spirillinida)의 한 계통입니다.[70][22] 후기 스피릴리니다(spirillinids)는 다실(multilocularity)과 방해석 껍질을 진화시켰으며, 최초의 형태는 트라이아스기에 나타났습니다. 이 그룹은 K-Pg 멸종으로 인한 다양성 감소에 거의 영향을 받지 않았습니다.[71]
가장 오래된 다실 유공충은 응집성 종이며, 중기 캄브리아기 시대의 화석 기록에 나타납니다. 하지만 보존 상태가 좋지 않아 주요 유공충 그룹에 확실하게 분류하기는 어렵습니다.[70]
가장 오래된 석회질 벽을 가진 유공충은 푸줄리나(Fusulinida)이며, 초기 실루리아기 란도베리절(Llandovery)에 화석 기록에 나타납니다. 초기 푸줄리나(fusulinids)는 현미경적 크기였으며, 평면 나선형으로 감겨 있고, 나선이 풀려 있었습니다. 후기에는 렌즈형, 구형, 길쭉한 쌀알 모양 등 다양한 형태로 진화했습니다.[72][73] 후기 푸줄리나(fusulinids) 종은 훨씬 더 큰 크기로 성장했으며, 일부는 길이가 5cm에 달했고, 어떤 표본은 최대 14cm에 달해 현존하거나 멸종된 가장 큰 유공충 중 하나입니다. 푸줄리나(Fusulinids)는 광합성 유기체와 공생 관계를 진화시킨 것으로 추정되는 가장 오래된 유공충 계통입니다. 푸줄리나(fusulinids) 화석은 남극 대륙을 제외한 모든 대륙에서 발견되었습니다. 이들은 석탄기 비제절(Viséan)에 가장 큰 다양성을 보였고, 이후 점차 다양성이 감소하여 페름기-트라이아스기 멸종 사건 때 멸종했습니다.[33][71][74]
석탄기 투르네시절(Tournaisian) 동안, 밀리올리다(Miliolida) 유공충이 처음으로 화석 기록에 나타났으며, 투보탈라메아(Tubothalamea) 내에서 스피릴리니다(Spirillinida)에서 분리되었습니다. 밀리올리다(Miliolids)는 페름기-트라이아스기 및 K-Pg 멸종에서 약 50%의 손실을 입었지만, 오늘날까지 생존했습니다. 일부 화석 밀리올리다(miliolids)는 직경이 최대 2cm에 달했습니다.[71]
가장 오래된 라게니다(Lagenida) 화석은 석탄기 모스코비절(Moscovian)에 나타납니다. 페름기-트라이아스기 또는 K-Pg 멸종의 영향을 거의 받지 않은 이 그룹은 시간이 지남에 따라 다양화되었습니다. 이차적으로 단실 분류군은 쥐라기와 백악기 동안 진화했습니다.
가장 오래된 인볼루티니다(Involutinida) 화석은 페름기에 나타납니다. 이 계통은 유라시아의 중생대(Mesozoic) 전반에 걸쳐 다양화되었지만, 세노마눔-투로눔 경계 사건(Cenomanian-Turonian boundary event) 이후 화석 기록에서 사라진 것으로 보입니다. 현생 그룹 플라니스피릴리니다이(planispirillinidae)가 인볼루티니다(involutinida)로 언급되기도 하지만, 이는 여전히 논란의 여지가 있습니다.[75][71]
로베르티니다(Robertinida)는 트라이아스기 아니시절(Anisian) 동안 화석 기록에 처음 나타납니다. 이 그룹은 화석 역사 전반에 걸쳐 낮은 다양성을 유지했습니다. 모든 살아있는 대표는 팔레오세에 처음 나타난 로베르티니다이(Robertinidae)에 속합니다.[71]
가장 오래된 확실한 로탈리다(Rotaliida) 화석은 트라이아스기-쥐라기 멸종 사건 이후 쥐라기의 플리엔스바키안절(Pliensbachian)까지 화석 기록에 나타나지 않습니다.[76] 이 그룹의 다양성은 세노마눔-투로눔 사건(Cenomanian-Turonian event) 이후까지 낮게 유지되었으며, 그 후 급속한 다양화를 보였습니다. 이 그룹 중 플랑크톤성 글로비게리나(Globigerinina) (가장 오래된 플랑크톤성 유공충 그룹)는 토아르시안 전환(Toarcian Turnover) 이후에 처음 나타났습니다. 이 그룹은 K-Pg 멸종과 에오세-올리고세 멸종 사건(Eocene-Oligocene extinction) 모두에서 큰 손실을 보았지만, 오늘날까지 현존하고 다양합니다.[71] 플랑크톤성 생활 방식의 추가 진화는 마이오세(Miocene) 또는 플라이오세(Pliocene)에 발생했는데, 로탈리다(rotaliid) ''Neogallitellia''가 독립적으로 플랑크톤성 생활 방식을 진화시켰습니다.[41][42]
껍질을 가지고 있기 때문에 캄브리아기 이후부터 화석으로 많이 발견되어 시상화석, 시준화석으로 중요하게 여겨진다. 고생대 석탄기의 푸줄리나(Fusulina)나 화폐석은 현재는 볼 수 없는 대형 종입니다.[109] 그 외에도 ''Dentalina'', ''Endothyra'', ''Globigerina'', ''Hyperammina'', ''Triticites'' 등이 있습니다. 저서성 유공충은 캄브리아기 이전에 출현하였고, 부유성 유공충이 출현한 것은 쥐라기 이후입니다.[109]
8. 응용
유공충은 생층서, 고기후학, 고해양학, 석유 탐사, 고고학 등 다양한 분야에서 활용된다.
- 생층서: 유공충은 작고 단단한 껍질, 풍부한 개체수, 종 구별의 용이성 덕분에 생층서 연구에 유용하다. 화석 유공충 군집을 분석하여 암석의 연대를 추정할 수 있다.
- 고기후학 및 고해양학: 유공충 껍질의 안정 동위원소 비율과 미량 원소 함량은 과거 기후 및 해양 환경을 재구성하는 데 사용된다.
- 석유 탐사: 유공충은 퇴적층의 연대와 고환경을 해석하고, 열 성숙도를 추정하여 석유 매장량을 찾는 데 활용된다. 유공충 색채 지수(FCI)는 석유 생성 초기 단계(약 100°C)에서 유용하다.
- 고고학: 석재에 포함된 유공충 화석을 분석하여 석재의 기원을 추정할 수 있다.
- 환경 지표: 유공충은 특정 환경 조건에 민감하게 반응하므로, 고대 해양 환경을 추적하고 변화하는 기후 조건을 연구하는 데 사용된다. 또한, 현생 유공충 군집은 해안 환경의 생물지표로 활용된다.
이처럼 유공충은 다양한 분야에서 유용한 정보를 제공하며, 특히 미화석으로서 그 가치가 높다.
8. 1. 생물 층서
유공충은 생층서 분야에서 매우 유용하게 활용된다. 유공충의 작고 단단한 껍질은 보존 상태가 좋고 풍부하게 발견되며, 복잡한 형태 덕분에 종을 쉽게 구별할 수 있다. 화석 기록에 나타나는 유공충 종들은 최초 진화와 소멸 시기가 명확하게 구분되어, 팬에로조익 시대 대부분에 걸쳐 유공충 군집의 변화를 연구하여 암석의 연대를 좁힐 수 있다. 이는 방사성 동위원소 연대 측정이 불가능한 퇴적암의 연대를 해석하는 데 유용하다.[91] 이러한 유공충의 활용은 1920년 알바 C. 엘리서에 의해 발견되었다.[92]유공충 껍질은 미화석으로 산출되며, 다양한 형태와 복잡성, 그리고 다량으로 산출되는 특징 덕분에 화석 유공충 군집을 통해 암석의 상대적 연대를 정확하게 파악할 수 있다. 현생 유공충 군집은 해안 환경의 생물지표로도 사용되는데, 이는 유공충의 서식지가 수온, 수심, 정수압, 빛, 산소, 염분 등 다양한 요인에 의해 결정되고 환경 변화에 민감하게 반응하기 때문이다. 하지만 유공충의 탄산칼슘 껍질은 산성 조건에서 쉽게 용해되어 화석화 이후 변형될 수 있으며, 기후 변화나 해양 산성화의 영향을 받거나 다른 광물로 치환될 가능성도 있다.[105]
석유 산업에서는 유층 후보를 찾는 데 유공충과 같은 미화석을 활용하기도 했다. 유공충 껍질의 일부는 퇴적물로 쌓여 유공충 연니를 형성하며, 시간이 흘러 굳어지면 석회암이 된다. 이집트 피라미드 건축 재료로 사용된 누메룰라이트 석회암이 대표적인 예시이다.
8. 2. 고기후 및 고해양학
죽은 플랑크톤성 유공충은 엄청난 수로 계속해서 해저로 떨어지며, 그들의 광물질 껍질(test)은 쌓이는 퇴적암 속에 화석으로 보존된다. 1960년대부터, 심해 시추 계획, 해양 시추 계획, 국제 해양 시추 프로그램(International Ocean Drilling Programmes)의 후원 아래, 그리고 석유 탐사 목적으로 발전된 심해 시추 기술은 유공충 화석을 포함한 퇴적물 코어를 채취해 왔다.[77] 이러한 화석 껍질의 사실상 무한한 공급과 코어에 대해 이용 가능한 비교적 정밀한 연대 측정 모델은 중기 쥐라기까지 거슬러 올라가는 매우 질 높은 플랑크톤성 유공충 화석 기록을 만들어냈으며, 과학자들이 진화 과정을 시험하고 기록하는 데 비할 데 없는 기록을 제공한다.[77] 이 화석 기록의 탁월한 질은 종 간의 상호 관계에 대한 인상적으로 상세한 그림을 화석을 기반으로 개발할 수 있게 했으며, 많은 경우 이후 현존 표본에 대한 분자 유전 연구를 통해 독립적으로 검증되었다.[78]특정 유형의 유공충은 특정 환경에서만 발견되기 때문에, 그들의 화석은 고대 해양 퇴적물이 퇴적된 환경의 종류를 알아내는 데 사용될 수 있다. 염도, 수심, 산소 조건 및 광 조건과 같은 조건은 다양한 종의 유공충의 서식지 선호도를 통해 결정될 수 있다. 이를 통해 연구자들은 유공충에 대한 정보를 종합하여 시간에 따른 변화하는 기후와 환경 조건을 추적할 수 있다.[79] 암석에서 발견되는 플랑크톤성 유공충과 저서성 유공충 화석의 상대적 비율은 암석이 퇴적될 당시 특정 지역의 수심을 나타내는 지표로 사용될 수 있다.[80]


최소 1997년부터 팔레오세-에오세 최고온 시기(PETM)는 지구 온난화의 영향과 해양과 대기로의 대량 탄소 유입, 해양 산성화를 포함한 것을 이해하기 위한 유추로 조사되어 왔다.[85] 오늘날 인류는 매년 약 10GT의 탄소(약 37GT CO2e)를 배출하며, 그 속도로 약 1000년 후에 PETM과 비슷한 양을 배출할 것이다. 주요 차이점은 PETM 당시 지구에는 빙하가 없었고, 드레이크 해협이 아직 열리지 않았고 중앙아메리카 해로가 아직 닫히지 않았다는 점이다.[86] PETM은 현재 일반적으로 지구 온난화와 대량 탄소 배출에 대한 사례 연구로 여겨지지만, 이 사건의 원인, 세부 사항 및 전반적인 중요성은 불확실하다.[87][88][89][90]

석회질 화석 유공충은 서식했던 고대 바다에서 발견되는 원소로 형성된다. 따라서 이들은 고기후학 및 고해양학에서 매우 유용하다. 껍질(test)의 안정 동위원소 비율과 미량 원소 함량을 조사하여 과거 기후를 재구성하는 기후 대용 지표로 사용할 수 있다. 산소 동위원소는 지구 온도와 빙량을 나타낼 수 있으며, 탄소의 안정 동위원소 비율을 조사하여 탄소 순환과 해양 생산성의 역사를 알 수 있다; δ18O 및 δ13C 참조. 스트론튬(Sr),[94] 마그네슘(Mg),[95] 리튬(Li)[96] 및 붕소(B)[97]와 같은 미량 원소의 농도는 지구 온도 순환, 대륙 풍화 및 지구 탄소 순환에서 해양의 역할에 대한 풍부한 정보를 제공한다. 플랑크톤성 유공충 화석 기록에서 볼 수 있는 지리적 패턴은 고대 해류를 재구성하는 데에도 사용된다.

유공충 화석은 고기후학이나 고해양학에서도 유용하다. 유공충 화석의 산소 안정 동위원소비(18O의 16O에 대한 비)를 조사함으로써 과거의 기후를 재구성할 수 있다. 또한 부유성 유공충 화석의 지리적 분포 및 탄소 안정 동위원소비는 과거 해류를 재구성하는 데 사용된다. 즉, 화석 유공충은 시준화석으로서도 시상화석으로서도 중요하다.[106]
8. 3. 석유 탐사
석유 산업은 잠재적인 탄화수소 매장량을 찾는 데 유공충과 같은 미화석에 크게 의존한다.[98] 유공충은 석유 탐사에 여러 용도로 사용되며, 유정의 퇴적층 연대와 고환경을 해석하는 데 일상적으로 사용된다.[99] 퇴적 분지에 깊이 매몰된 응집성 화석 유공충은 석유 생성의 주요 요소인 열 성숙도를 추정하는 데 사용될 수 있다. 유공충 색채 지수[100](FCI)는 색 변화를 정량화하고 매몰 온도를 추정하는 데 사용된다. FCI 데이터는 석유 생성 초기 단계(약 100°C)에서 특히 유용하다.8. 4. 고고학
석유 산업은 잠재적인 탄화수소 매장량을 찾는 데 유공충과 같은 미세 화석에 크게 의존한다.[98] 유공충은 생층서 지표로서 연안 환경의 생물지표나 산호초 건강 지표로 사용되어 왔다. 탄산칼슘은 산성 조건에서 용해되기 쉽기 때문에 유공충은 기후 변화와 해양 산성화의 영향을 특히 많이 받을 수 있다.유공충은 석유 탐사에 여러 용도로 사용되며, 유정의 퇴적층의 연대와 고환경을 해석하는 데 일상적으로 사용된다.[99] 퇴적 분지에 깊이 매몰된 응집성 화석 유공충은 석유 생성의 주요 요소인 열 성숙도를 추정하는 데 사용할 수 있다. 유공충 색채 지수[100](FCI)는 색 변화를 정량화하고 매몰 온도를 추정하는 데 사용된다. FCI 데이터는 석유 생성 초기 단계(약 100°C)에서 특히 유용하다.
유공충은 일부 석재 원료의 기원 추정에도 사용될 수 있다. 석회암과 같은 일부 석재 유형에는 일반적으로 화석화된 유공충이 포함되어 있는 것으로 나타난다. 석재 샘플 내 이러한 화석의 유형과 농도를 사용하여 해당 샘플을 동일한 "화석 시그니처"를 포함하는 것으로 알려진 원천과 일치시킬 수 있다.[101]
8. 5. 환경 지표
특정 유형의 유공충은 특정 환경에서만 발견되기 때문에, 이들의 화석은 고대 해양 퇴적물이 퇴적된 환경을 알아내는 데 사용될 수 있다. 염도, 수심, 산소 및 광 조건과 같은 조건은 다양한 종의 유공충의 서식지 선호도를 통해 결정될 수 있다.[79] 이를 통해 연구자들은 유공충에 대한 정보를 종합하여 시간에 따른 변화하는 기후와 환경 조건을 추적할 수 있다.암석에서 발견되는 플랑크톤성 유공충과 저서성 유공충 화석의 상대적 비율은 암석이 퇴적될 당시 특정 지역의 수심을 나타내는 지표로 사용될 수 있다.[80]
팔레오세-에오세 최고온 시기(PETM)는 지구 온난화의 영향과 해양과 대기로의 대량 탄소 유입, 해양 산성화를 포함한 것을 이해하기 위한 유추로 조사되어 왔다.[85] 오늘날 인류는 매년 약 10Gt의 탄소(약 37Gt CO2e)를 배출하며, 그 속도로 약 1000년 후에 PETM과 비슷한 양을 배출할 것이다. 주요 차이점은 PETM 당시 지구에는 빙하가 없었고, 드레이크 해협이 아직 열리지 않았고 중앙아메리카 해로가 아직 닫히지 않았다는 점이다.[86] PETM은 현재 일반적으로 지구 온난화와 대량 탄소 배출에 대한 사례 연구로 여겨지지만, 이 사건의 원인, 세부 사항 및 전반적인 중요성은 불확실하다.[87][88][89][90]
석회질 화석 유공충은 서식했던 고대 바다에서 발견되는 원소로 형성된다. 따라서 이들은 고기후학 및 고해양학에서 매우 유용하다. 껍질(test)의 안정 동위원소 비율과 미량 원소 함량을 조사하여 과거 기후를 재구성하는 기후 대용 지표로 사용할 수 있다. 산소 동위원소는 지구 온도와 빙량을 나타낼 수 있으며, 탄소의 안정 동위원소 비율을 조사하여 탄소 순환과 해양 생산성의 역사를 알 수 있다.[93] ( δ18O 및 δ13C 참조) 스트론튬(Sr),[94] 마그네슘(Mg),[95] 리튬(Li)[96] 및 붕소(B)[97]와 같은 미량 원소의 농도는 지구 온도 순환, 대륙 풍화 및 지구 탄소 순환에서 해양의 역할에 대한 풍부한 정보를 제공한다. 플랑크톤성 유공충 화석 기록에서 볼 수 있는 지리적 패턴은 고대 해류를 재구성하는 데에도 사용된다.
현생 유공충 군집은 해안 환경의 생물지표로 사용된다. 이는 유공충의 서식지(혹은 형태)가 수온, 수심, 정수압, 빛, 산소, 염분 등 다양한 요인에 의해 결정되고, 환경 변화에 민감하게 반응한다는 성질 때문이다. 단, 유공충의 탄산칼슘 껍질은 산성 조건에서는 용해되기 쉽기 때문에, 화석화 후 변형으로 기후 변화나 해양 산성화의 영향을 받거나, 다른 광물로의 치환 등이 일어날 가능성도 있다.[105]
유공충 화석은 고기후학이나 고해양학에서도 유용하다. 유공충 화석의 산소 안정 동위원소비(18O의 16O에 대한 비)를 조사함으로써 과거의 기후를 재구성할 수 있다. 또한 부유성 유공충 화석의 지리적 분포 및 탄소 안정 동위원소비는 과거 해류를 재구성하는 데 사용된다. 즉, 화석 유공충은 시준화석으로서도 시상화석으로서도 중요하다.[106]
참조
[1]
Wikidata
[2]
논문
Foraminifers (Benthic)
https://doi.org/10.1[...]
Springer Netherlands
2021-05-13
[3]
논문
Planktic Foraminifera
https://doi.org/10.1[...]
Springer Japan
2021-05-13
[4]
논문
Taxonomic revision of freshwater foraminifera with the description of two new agglutinated species and genera
2017-08-01
[5]
논문
Benthonic Foraminifera of Brackish, Hypersaline, Ultrasaline, and Fresh Waters
https://doi.org/10.1[...]
Springer Netherlands
2021-05-13
[6]
서적
Meiobenthology: the microscopic motile fauna of aquatic sediments
Springer
[7]
논문
Molecular evidence for widespread occurrence of Foraminifera in soils
2010-09-01
[8]
서적
Neogene planktonic foraminifera: a phylogenetic atlas
https://books.google[...]
Hutchinson Ross
[9]
논문
Next-Generation Environmental Diversity Surveys of Foraminifera: Preparing the Future
https://www.journals[...]
2014-10-01
[10]
논문
Diversity, Nomenclature, and Taxonomy of Protists
https://wayback.arch[...]
2007
[11]
논문
Next-generation environmental diversity surveys of foraminifera: preparing the future
http://www.biolbull.[...]
[12]
웹사이트
World Foraminifera Database
http://www.marinespe[...]
[13]
뉴스
Zoologger: 'Living beach ball' is giant single cell
https://www.newscien[...]
2010-02-03
[14]
논문
What Should We call the Foraminifera
http://www.ucmp.berk[...]
2018-04-10
[15]
웹사이트
Foraminifera Fossil Focus Time Discovering Geology British Geological Survey (BGS)
https://www.bgs.ac.u[...]
2020-07-20
[16]
웹사이트
Micrographia, or, Some physiological descriptions of minute bodies made by magnifying glasses ?with observations and inquiries thereupon /by R. Hooke ... : Hooke, Robert, : Free Download, Borrow, and Streaming
https://archive.org/[...]
2020-07-20
[17]
논문
Modern Foraminifera
Springer Netherlands
2003
[18]
논문
Biology and Evolutionary History of Larger Benthic Foraminifera
UCL Press
2018
[19]
웹사이트
On Lorentz Spengler and a neotype for the foraminifer Calcarina spengleri
https://www.research[...]
1981-01-01
[20]
논문
Tableau Méthodique de la Classe des Céphalopodes
https://www.biodiver[...]
1826
[21]
웹사이트
Kennedy, Margaret Moore (1896–1967)
http://dx.doi.org/10[...]
Oxford University Press
2022-11-03
[22]
논문
New supraordinal classification of Foraminifera: Molecules meet morphology
http://www.sciencedi[...]
2013-04-01
[23]
서적
Part C, Protista 2
Geological Society of America
[24]
서적
Modern Foraminifera
https://books.google[...]
Springer
[25]
논문
Only Six Kingdoms of Life
http://www.cladocera[...]
[26]
논문
Protist phylogeny and the high-level classification of Protozoa
[27]
웹사이트
Tolweb Cercozoa
http://tolweb.org/Ce[...]
[28]
웹사이트
European Register of Marine Species
http://www.marbef.or[...]
[29]
웹사이트
eForams-taxonomy
http://www.eforams.i[...]
[30]
웹사이트
Testate amoebae as environmental indicators
http://www6.plymouth[...]
[31]
논문
New insight into the systematics and evolution of the foraminifera
https://www.research[...]
2013
[32]
논문
Molecular Evidence That Reticulomyxa Filosa Is A Freshwater Naked Foraminifer
1999
[33]
논문
Calcareous-Walled Microfossils
Springer International Publishing
2016
[34]
서적
Foraminifera: notes for a short course organized by M.A. Buzas and B.K. Sen Gupta
University of Tennessee, Dept. of Geological Sciences
[35]
서적
Modern Planktonic Foraminifera
https://books.google[...]
Springer-Verlag
[36]
논문
Cytogenetic systems and evolution in foraminifera
http://dx.doi.org/10[...]
1979-01-01
[37]
논문
An overview of cellular ultrastructure in benthic foraminifera: New observations of rotalid species in the context of existing literature
2018-01-01
[38]
논문
Natural 226Ra and 232Th radionuclides in xenophyophores of the Pacific Ocean
http://link.springer[...]
2015-07-01
[39]
논문
Characterizing photosymbiosis in modern planktonic foraminifera
2019-01-01
[40]
논문
Cryptic species of planktonic foraminifera: their effect on palaeoceanographic reconstructions
2002-04-01
[41]
논문
Molecular evidence for an independent origin of modern triserial planktonic foraminifera from benthic ancestors
2008-12-01
[42]
논문
Substitute names for some unicellular animal taxa (Protozoa
https://www.munisent[...]
2009-06-01
[43]
논문
Chamber arrangement versus wall structure in the high-rank phylogenetic classification of Foraminifera
2019-01-01
[44]
웹사이트
Symbiosis and microbiome flexibility in calcifying benthic foraminifera of the Great Barrier Reef
https://microbiomejo[...]
[45]
웹사이트
Characterizing photosymbiosis in modern planktonic foraminifera - BG
https://bg.copernicu[...]
[46]
서적
Advances in Microbial Ecology, Volum 11
https://books.google[...]
[47]
논문
Benthic Foraminifera of dysoxic sediments: chloroplast sequestration and functional morphology
[48]
서적
Modern Foraminifera
Springer Netherlands
2003
[49]
논문
Rapid response of the giant protist xenophyophores (Foraminifera, Rhizaria) to organic matter supply at abyssal depths revealed by an in situ dual stable isotope labeling experiment
https://www.scienced[...]
2021-10-01
[50]
서적
Predation on and by Foraminifera
http://link.springer[...]
Springer US
2020-09-30
[51]
논문
Tolerance of Infaunal Benthic Foraminifera for Low and High Oxygen Concentrations
https://www.biodiver[...]
1992-08-01
[52]
논문
New organic-walled Foraminifera (Protista) from the ocean's deepest point, the Challenger Deep (western Pacific Ocean)
https://zenodo.org/r[...]
2008-07-01
[53]
논문
Review: Freshwater and Soil Foraminifera – A Story of Long-Forgotten Relatives
https://doi.org/10.2[...]
2021-10-29
[54]
서적
Invertebrate Fossils
McGraw-Hill
[55]
서적
Foraminifera
https://books.google[...]
Springer
1981-06-18
[56]
논문
Cretaceous Carterina (Foraminifera)
[57]
논문
A section of Vendian in the east of West Siberian Plate (based on data from the Borehole Vostok 3)
http://www.sciencedi[...]
2008-12-01
[58]
웹사이트
Foraminifera: History of Study
http://www.ucl.ac.uk[...]
2007-09-20
[59]
논문
Global ocean carbonate and carbon dioxide production: The role of reef Foraminifera
http://jfr.geoscienc[...]
[60]
논문
The new higher level classification of Eukaryotes with emphasis on the taxonomy of Protists
[61]
논문
The evolution of early Foraminifera
2003-09-30
[62]
논문
Bayesian relaxed clock estimation of divergence times in foraminifera
http://www.sciencedi[...]
2011-10-01
[63]
논문
The nature of vendobionts
https://sp.lyellcoll[...]
2007-01-01
[64]
논문
Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals
2018-09-21
[65]
논문
Piaeodicton: The Traces of Infaunal Xenophyophores?
https://www.science.[...]
1982-10-01
[66]
논문
Paleoecology and Ecology of Xenophyophores
1994-01-01
[67]
논문
Paleodictyon nodosum: A living fossil on the deep-sea floor
http://www.sciencedi[...]
2009-09-01
[68]
논문
Giant protists (xenophyophores, Foraminifera) are exceptionally diverse in parts of the abyssal eastern Pacific licensed for polymetallic nodule exploration
2017-03-01
[69]
논문
Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms
2001-01-01
[70]
논문
Foraminifera from the Cambrian of Nova Scotia: The oldest multichambered foraminifera
https://pubs.geoscie[...]
2003-06-01
[71]
논문
Foraminiferal Evolution, Diversification, and Extinction
1988
[72]
서적
The Carboniferous of the World: China, Korea, Japan & S.E. Asia
https://books.google[...]
IGME
1983
[73]
서적
The Biology of Reefs and Reef Organisms
https://books.google[...]
University of Chicago Press
2023-01-10
[74]
웹사이트
Fusulinids GeoKansas
http://geokansas.ku.[...]
2020-05-16
[75]
웹사이트
PBDB Navigator
https://paleobiodb.o[...]
2020-05-16
[76]
논문
Benthic foraminifers and palaeoenvironment in the Lower and Middle Jurassic of the Western Basque-Cantabrian Basin (Northern Spain)
2005
[77]
논문
Nature debates
http://www.nature.co[...]
1998-11-19
[78]
논문
Using the Multiple Analysis Approach to Reconstruct Phylogenetic Relationships among Planktonic Foraminifera from Highly Divergent and Length-polymorphic SSU rDNA Sequences
http://www.la-press.[...]
[79]
논문
Cenomanian to Turonian foraminifera from Ashaka (NE Nigeria): quantitative analysis and palaeoenvironmental interpretation
http://www.sciencedi[...]
1997-02-01
[80]
논문
Badenian (Middle Miocene) basin development in SW Hungary: subsidence history based on quantitative paleobathymetry of foraminifera
2002-05-01
[81]
웹사이트
Planktic Foraminifera from Southern Maryland
https://www.usgs.gov[...]
United States Geological Survey
2021
[82]
논문
Climate Response to Orbital Forcing Across the Oligocene-Miocene Boundary
http://doc.rero.ch/r[...]
American Association for the Advancement of Science (AAAS)
2001-04-13
[83]
논문
A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records
2005
[84]
논문
Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica
http://www.escholars[...]
Springer Science and Business Media LLC
[85]
논문
A blast of gas in the latest Paleocene; simulating first-order effects of massive dissociation of oceanic methane hydrate
[86]
웹사이트
PETM Weirdness
http://www.realclima[...]
RealClimate
2016-02-03
[87]
논문
A perturbation of carbon cycle, climate, and biosphere with implications for the future
http://www.whoi.edu/[...]
2016-02-03
[88]
논문
Carbon dioxide forcing alone insufficient to explain Palaeocene–Eocene Thermal Maximum warming
[89]
웹사이트
Ancient microfossils are the key to future climate recovery
https://www.usgs.gov[...]
United States Geological Survey
2022
[90]
논문
Surface ocean warming and acidification driven by rapid carbon release precedes Paleocene-Eocene Thermal Maximum
https://lirias.kuleu[...]
American Association for the Advancement of Science (AAAS)
2022-03-18
[91]
웹사이트
Biostratigraphy
https://www.ga.gov.a[...]
2014-05-15
[92]
논문
The Foraminiferal Fauna of the Anahuac Formation
1945-01-01
[93]
논문
Trends, Rhythms, and Aberrations in Global Climate, 65 Ma to Present
http://doc.rero.ch/r[...]
[94]
논문
Exploring foraminiferal Sr/Ca as a new carbonate system proxy
https://epic.awi.de/[...]
2017-04
[95]
논문
The coordination of Mg in foraminiferal calcite
2013-12
[96]
논문
Lithium Isotope History of Cenozoic Seawater: Changes in Silicate Weathering and Reverse Weathering
2012-01-26
[97]
논문
Boron isotopic composition and concentration in modern marine carbonates
1992-01
[98]
서적
Fossil Invertebrates
Wiley
[99]
서적
Micropalaeontology in petroleum exploration
https://books.google[...]
Clarendon Press
[100]
서적
Colour Alteration, Thermal Maturity, and Burial Diagenesis in Fossil Foraminifers
https://books.google[...]
Geological Survey of Canada
[101]
논문
The application of microfossils in assessing the provenance of chalk used in the manufacture of Roman mosaics at Silchester
http://www.sciencedi[...]
2008-08-01
[102]
웹사이트
Zoologger: 'Living beach ball' is giant single cell
https://www.newscien[...]
[103]
논문
Simple foraminifera flourish at the ocean's deepest point
[104]
서적
Protistology
E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart
[105]
서적
微古生物学(上巻)
朝倉書店
[106]
저널
The deep ocean during the last interglacial period
[107]
저널
Polyubiquitin insertions and the phylogeny of Cercozoa and Rhizaria
[108]
저널
Monophyly of Rhizaria and multigene phylogeny of unicellular bikonts
[109]
저널
古環境指標としての浮遊性有孔虫
九州大学大学院比較社会文化学府
[110]
저널
Estimating the timing of early eukaryotic diversification with multigene molecular clocks
http://www.pnas.org/[...]
2011-08-16
[111]
저널
Phylogenomics supports the monophyly of the Cercozoa
2019-01-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com