코틸로린쿠스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
코틸로린쿠스는 컵 또는 구멍을 뜻하는 그리스어 'kotyle'와 부리를 의미하는 'rhynchos'에서 유래된 이름으로, 컵 모양의 뼈 표면으로 둘러싸인 콧구멍을 가진 카세이드 단궁류의 한 속이다. 이 속에는 크기가 다른 세 종, 즉 C. 로메리, C. 핸코키, C. 브랜소니가 포함되며, C. 로메리는 성적 이형성을 나타내는 두 개의 크기 그룹을 가진 것으로 추정된다. 코틸로린쿠스는 앞쪽으로 경사진 주둥이, 매우 큰 비강 개구부, 수많은 작은 함몰부가 있는 두개골 지붕, 매우 큰 송과선 구멍을 특징으로 하며, 튼튼한 몸통 골격과 짧고 튼튼한 사지를 가지고 있다. 이들은 초식 동물로, 긴 갈비뼈와 강력한 턱을 통해 식물을 소화했다. 화석은 북아메리카 남부, 텍사스 주, 오클라호마 주 등지에서 발견되며, 시기에 따라 쿵구리안 또는 로디안 시대로 추정된다.
더 읽어볼만한 페이지
- 카세아아목 - 카세아과
카세아과는 페름기 초중기에 번성한 단궁류 분류군으로, 작은 두개골과 통 모양 몸통, 다양한 형태의 치아를 가지며 초기 잡식에서 초식으로 진화했고 일부는 반수생 생활을 했을 가능성이 있다. - 1937년 기재된 화석 분류군 - 파르크소사우루스
파르크소사우루스는 6950만 년 전 호스슈 캐니언 지층에서 발견된 소형 초식 공룡으로, 튼튼한 뒷다리와 긴 두개골을 가진 조반류의 한 속이다. - 1937년 기재된 화석 분류군 - 엘리오츠미시아
엘리오츠미시아는 남아프리카 공화국 후기 미들페름기에 서식했던 바라노피드 시냅시드로, 길고 가느다란 주둥이와 톱니 모양의 이빨을 가지고 곤충과 작은 척추동물을 사냥했을 것으로 추정되며, 1937년 로버트 브룸 박사에 의해 명명되었고, 현재는 바라놉시드 쇄골로 분류된다. - 페름기 북아메리카의 단궁류 - 스페나코돈
스페나코돈은 석탄기에서 페름기로 전환되는 시기에 멸종한 초기 단궁류 파충류로, 북아메리카에서 화석이 발견되며 등 융기가 특징적이고, 육식성이었으며, 포유류 조상과 관련이 깊다. - 페름기 북아메리카의 단궁류 - 에다포사우루스
에다포사우루스는 석탄기와 페름기 초기에 살았던 단궁류의 한 속으로, 등에 돛을 가지고 있으며, 최초의 초식성 양막류 중 하나로 추정된다.
| 코틸로린쿠스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Cotylorhynchus |
| 명명자 | 스토발, 1937 |
| 모식종 | Cotylorhynchus romeri |
| 모식종 명명자 | 스토발, 1937 |
| 화석 발견 시기 | 쿵구르절 - 로디절, 약 2억 7950만 년 전 - 2억 7000만 년 전 |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 아문 | 척추동물아문 |
| 상강 | 사지동물상강 |
| 강 | 단궁강 |
| 목 | 반룡목 |
| 아목 | 카세아사우루스아목 |
| 과 | 카세아과 |
| 종 | |
| 하위 종 | C. romeri C. hancocki C. bransoni |
2. 발견 및 명명
속명 ''코틸로린쿠스''는 컵, 구멍을 의미하는 그리스어 ''kotyle''와 부리 또는 주둥이를 의미하는 ''rhynchos''에서 유래했다. 이 속은 함몰된 컵 모양의 뼈 표면으로 둘러싸인 콧구멍 때문에 그렇게 명명되었다.[1]
코틸로린쿠스는 술통형의 몸통과, 그것에 비해 불균형적으로 작은 머리를 특징으로 하는 단궁류이다. 타입 종인 ''C. romeri''는 전체 길이 4.5m, 체중 330kg[50], 최대 종인 ''C. hancocki''는 체중 500kg에 달했을 것으로 추정되며[51], 디메트로돈류 중에서도 최대급에 속한다.
속 ''코틸로린쿠스''는 크기와 비율이 다른 세 종, 즉 ''C. 로메리'' (기준종), ''C. 핸코키'', ''C. 브랜소니''를 포함한다. ''C. 로메리''에는 성적 이형태를 나타내는 것으로 추정되는 두 개의 크기 그룹이 있다. ''C. 로메리''와 ''C. 핸코키'' 성체 사이에는 크기 중복이 없지만, ''C. 브랜소니''의 더 큰 표본은 ''C. 로메리''의 더 작은 표본과 대략 같은 치수를 갖는다.[4] 2022년, 베르네부르크와 동료들은 종 ''C. 핸코키''와 ''C. 브랜소니''가 속 ''코틸로린쿠스''에 속하지 않을 수 있다고 제안했다. 이 저자들은 이 두 분류군의 지위를 명확히 하기 위해 자세한 수정이 필요하다고 생각한다.[2]
3. 특징
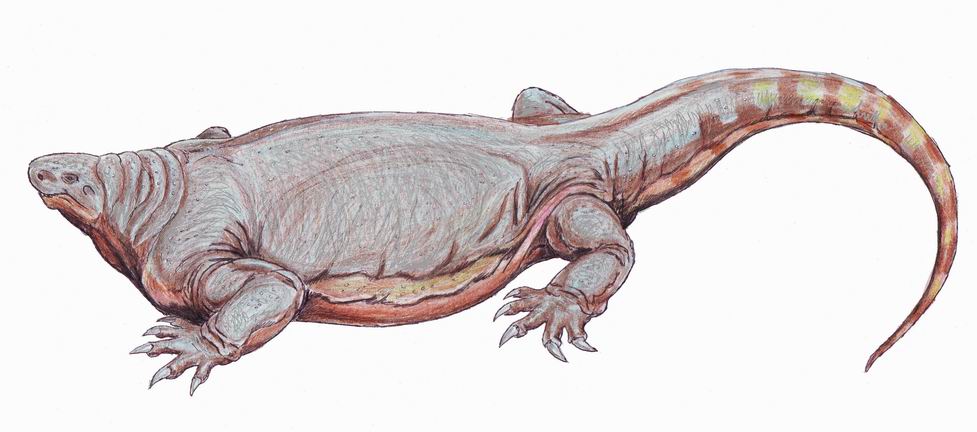
코틸로린쿠스는 크기와 비율이 다른 세 종, 즉 ''C. 로메리''(기준종), ''C. 핸코키'', ''C. 브랜소니''를 포함한다. ''C. 로메리''에는 성적 이형성을 나타내는 것으로 추정되는 두 개의 크기 그룹이 있다. ''C. 로메리''와 ''C. 핸코키'' 성체 사이에는 크기 중복이 없지만, ''C. 브랜소니''의 더 큰 표본은 ''C. 로메리''의 더 작은 표본과 대략 같은 치수를 갖는다.[4] 2022년, 베르네부르크와 동료들은 종 ''C. 핸코키''와 ''C. 브랜소니''가 속 ''코틸로린쿠스''에 속하지 않을 수 있다고 제안했다.[2]
3. 1. 형태
속명 ''코틸로린쿠스''는 컵, 구멍을 의미하는 그리스어 ''kotyle''와 부리 또는 주둥이를 의미하는 ''rhynchos''에서 유래했다. 이 속은 함몰된 컵 모양의 뼈 표면으로 둘러싸인 콧구멍 때문에 그렇게 명명되었다.[1]

코틸로린쿠스의 두개골은 앞쪽으로 경사진 주둥이, 매우 큰 비강 개구부, 수많은 작은 함몰부가 있는 두개골 지붕, 그리고 매우 큰 송과선 구멍으로 전형적인 케이스과 형태를 보인다. 후자는 ''에나토사우루스''처럼 폭이 길이보다 넓어 유로미크테르의 아원형과는 다르다.[3] 위턱과 아래턱의 치아 수는 16개에서 20개 사이이다. 위턱에서 앞쪽 치아는 길고 가늘고, 뒤쪽 치아는 뒤로 갈수록 크기가 작아지고 약간 주걱 모양이다. 모든 가장자리 치아는 먼쪽 끝이 입 안쪽으로 약간 기울어져 있고, 치관 상단에는 각각 세 개의 작은 돌기가 세로로 배열되어 있다. 이러한 치아는 또한 치관 중앙 부분의 확대를 보여준다.[4] 아래턱에서 올슨에 따르면 치아가 없는 앞쪽 치아는 더 짧고 약간 앞으로 기울어져 있다. 다른 아래쪽 치아는 위턱의 치아와 유사하다.
몸통 골격은 튼튼하다. 갈비뼈는 매우 길고 무거우며 구부러져 둥근 몸체를 형성한다. 갈비뼈는 모든 전천추 척추와 첫 번째 꼬리 척추에 존재한다. 5개의 뒤쪽 천추 앞쪽 갈비뼈는 척추의 가로 돌기와 융합되어 있다. 천골은 3개의 척추를 포함한다. 더 큰 표본의 신경 가시는 특히 골반 부위에서 비례적으로 더 높아진다. 사지는 짧고 튼튼하다. 넙다리뼈는 넙다리뼈의 등쪽 표면을 약간 덮는 가장자리로 표시된 넓은 선반이 있는 근위 끝이 특징이다. 발과 손은 넓고 짧으며, 강력하고 날카롭고 구부러진 발톱 발가락뼈로 끝나며, 이는 강력한 발톱을 지탱했을 것이다. 근육과 힘줄의 흔적이 매우 발달했다.[4]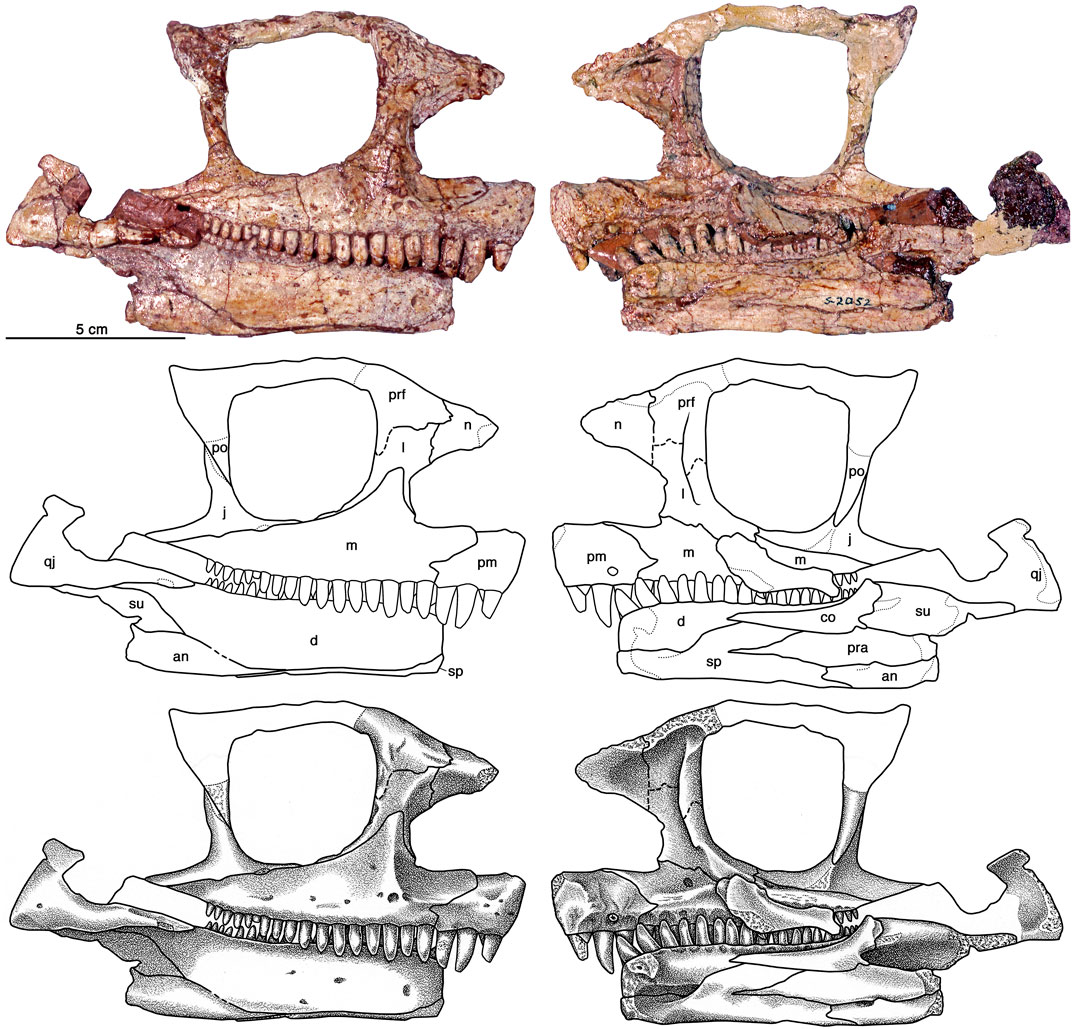
모식종인 ''Cotylorhynchus romeri''는 이 속에서 가장 잘 알려진 종이다. ''C. romeri''는 길이가 4.5m에 달하고, Romer와 Price에 따르면 무게가 330kg에 달할 수 있는 대형 종이다.[5][6]
다른 두 종의 ''Cotylorhynchus''와 마찬가지로, 치아는 삼첨 치아로 구성되어 있다(가장 앞쪽 치아 제외). 그러나 ''C. romeri''는 둔두가 가장 덜 발달한 종이다.[4]
척추는 25~26개의 전천추 척추, 3개의 천추 척추, 약 55개의 미추 척추로 구성되어 있다.[6]
''코틸로린쿠스 핸코키''는 1953년 에버렛 클레어 올슨과 제임스 R. 비어보어에 의해 명명되었다.[9]
길이가 6m에 달하고 무게가 500kg 이상인 ''C. 핸코키''는 속에서 가장 큰 종이며, 속 ''알리에라사우루스''와 함께 알려진 가장 큰 카세이드 중 하나이다.[11][12][13][14]
몸통 골격은 사지, 척추 및 골반의 형태와 비율로 구별된다. 견갑골은 관절와 구멍이 견갑골 날에 존재한다는 특징이 있다. 그러한 구멍은 다른 두 종의 ''코틸로린쿠스''와 일반적으로 카세이드에는 없지만, 속 ''라리에우도린쿠스''에는 존재한다.[2]
1962년, 에버렛 C. 올슨(Everett C. Olson)과 허버트 바르구센(Herbert Barghusen)은 ''Cotylorhynchus bransoni''을 명명했다.[15]
''C. bransoni''는 속 ''Cotylorhynchus''의 알려진 종 중 가장 작은 종으로, 가장 큰 개체의 크기가 ''C. romeri''의 가장 작은 개체와 비슷하다.[4] 두개골은 거의 알려져 있지 않으며, 위턱뼈 파편과 익상골의 두 개의 치아가 있는 뼈로만 대표된다.
어깨뼈-부리뼈는 다른 두 종보다 상대적으로 좁은 어깨 날을 가지고 있다. 관절와는 다른 두 종보다 폭에 비해 약간 더 길고, 부리뼈 판의 앞부분은 앞쪽으로 덜 확장되어 있다.[4]
타입 종인 ''C. romeri''는 전체 길이 4.5m, 체중 330kg[50], 최대 종인 ''C. hancocki''는 체중 500kg에 달했을 것으로 추정되며[51], 디메트로돈류 중에서도 최대급이 된다. 술통형의 몸통과, 그것에 비해 불균형적으로 작은 머리를 특징으로 한다.
3. 2. 종별 특징
''C. romeri''는 길이가 4.5m 이상, 무게가 330kg에 달할 수 있는 대형 종이다.[5][6] 두개골은 주둥이가 앞쪽으로 경사져 있고, 매우 큰 비강 개구부와 수많은 작은 함몰부가 있는 두개골 지붕, 그리고 매우 큰 송과선 구멍을 가진다. 위턱과 아래턱의 치아 수는 16개에서 20개 사이이다. 모든 가장자리 치아는 먼쪽 끝이 입 안쪽으로 약간 기울어져 있고, 치관 상단에는 각각 세 개의 작은 돌기가 세로로 배열되어 있다.[4]
다른 두 종의 ''Cotylorhynchus''와 마찬가지로, 치아는 삼첨 치아로 구성되어 있다(가장 앞쪽 치아 제외). 그러나 ''C. romeri''는 둔두가 가장 덜 발달한 종이다.[4] 구개골에는 10개의 원추형 이빨이 있다. 익상골은 ''C. bransoni''의 익상골에 있는 이빨보다 더 많고, 더 작고, 가늘다.[4] 척추는 25~26개의 전천추 척추, 3개의 천추 척추, 약 55개의 미추 척추로 구성되어 있다.[6] ''C. romeri''는 등쪽 척추의 관절돌기가 넓게 떨어져 있는 특징을 가지는 반면, ''C. hancocki''와 ''C. bransoni''에서는 일반적으로 정중선을 따라 접촉한다.[7] 손과 발은 2-2-3-3-2의 지골수를 보인다.[6][4]
길이가 6m에 달하고 무게가 500kg 이상인[4] ''C. 핸코키''는 속에서 가장 큰 종이며, 속 ''알리에라사우루스''와 함께 알려진 가장 큰 카세이드 중 하나이다.[11][12][13][14] 또한 그 크기는 그것을 가장 큰 비 포유류 단궁류 중 하나로 만든다. 여러 개의 고립된 하악골은 하악 치열이 최대 18개의 약간 주걱 모양의 삼첨 치아를 가지고 있음을 보여준다. 상악 치아의 치관은 하악 치아보다 약하다. 또한 ''C. 핸코키''의 치관은 ''C. 로메리''보다 더 두드러지지만 ''C. 브랜소니''의 치관보다는 덜 발달되어 있다.[10][4]
몸통 골격은 사지, 척추 및 골반의 형태와 비율로 구별된다. 견갑골은 관절와 구멍이 견갑골 날에 존재한다는 특징이 있다. 그러한 구멍은 다른 두 종의 ''코틸로린쿠스''와 일반적으로 카세이드에는 없지만, 속 ''라리에우도린쿠스''에는 존재한다.[2] 척추는 표본 FMNH UR 581에서 17개의 천추 전 척추와 39개의 꼬리 척추가 관절을 이루고 있다.[10] 이 종의 매우 큰 크기와 관련된 특징은 등 척추의 후관절돌기에 두드러진 ''저위돌기-반구조 연결''이 존재한다는 것이다.[7] 이는 ''라리에우도린쿠스''와 공유되는 특징이다.[2] 사지 뼈는 매우 강하다. 특히 대퇴골은 상대적으로 짧은 뼈몸통과 매우 발달된 내측 전자를 가진 매우 튼튼한 뼈이다.[10][8] 이는 ''라리에우도린쿠스''와 공유되는 또 다른 특징이다.[2]
1962년, 에버렛 C. 올슨(Everett C. Olson)과 허버트 바르구센(Herbert Barghusen)은 오클라호마주 킹피셔 카운티의 오메가 채석장에서 발견된 다수의 뼈를 바탕으로 ''Cotylorhynchus bransoni''을 명명했다. 종명은 칼 C. 브랜슨(Carl C. Branson) 박사를 기리기 위해 붙여졌다.[15]
''C. bransoni''는 속 ''Cotylorhynchus''의 알려진 종 중 가장 작은 종으로, 가장 큰 개체의 크기가 ''C. romeri''의 가장 작은 개체와 비슷하다.[4] 두개골은 거의 알려져 있지 않으며, 위턱뼈 파편과 익상골의 두 개의 치아가 있는 뼈로만 대표된다. 위턱뼈에 보존된 두 개의 삼첨 치아는 ''C. romeri''와 ''C. hancocki''에서 관찰되는 것보다 더 발달된 작은 융기를 보여준다. 익상골은 ''C. romeri''의 익상골에 존재하는 것보다 더 적고, 크고, 더 튼튼한 이빨을 가지고 있다.[10][4]
관절와는 다른 두 종보다 폭에 비해 약간 더 길고, 부리뼈 판의 앞부분은 앞쪽으로 덜 확장되어 있다.[4] 목말뼈는 다른 두 종에는 없는 매우 큰 구멍의 존재로 특징지어진다.[10][4] 올슨과 바르구센은 ''C. bransoni'' 발의 지골 공식이 2-2-2-3-2라고 생각했지만,[15][4] 2015년 로마노와 니코시아는 셋째 발가락이 세 개의 발가락뼈를 가지고 있으며 두 개가 아니라는 것을 보여주었다. 따라서 ''C. bransoni'' 발의 지골 공식은 ''C. romeri''와 마찬가지로 2-2-3-3-2이며, 아마도 ''C. hancocki''에서도 마찬가지일 것이다.[7]
4. 고생물학
'''코틸로린쿠스 로메리'''. 위: 두개골 재구성, 왼쪽 측면도, 턱뼈의 내측 및 외측 보기; 아래: 두개골 재구성, 등쪽, 배쪽, 뒤쪽 및 앞쪽 보기.
코틸로린쿠스는 초식동물이었고, 그 크기로 인해 포식자의 위협을 적게 받았을 것으로 보인다. 거대한 몸통은 열적 관성을 높여 체온 유지에 도움을 주었을 것으로 추정된다.[53]
4. 1. 식성
코틸로린쿠스는 잘 발달된 둥근 형태의 늑골을 가지고 있었는데, 이는 많은 양의 영양분이 적은 식물을 섭취하기에 적합한 거대한 소화 기관이 있었음을 나타낸다. 코틸로린쿠스의 치열 또한 초식성이었음을 보여준다. 앞니는 더 길고 약간 구부러져 입 안의 식물을 모으는 역할을 했을 것이다. 삼첨두(세 개의 뾰족한 돌기) 형태의 가장자리 이빨은 식물을 자르고 절단하는 데 매우 적합했다.[4][3]일부 카세아사우루스과 동물(''유로미크테르'' 및 ''에나토사우루스'')에서 보존된 설골은 비교적 움직일 수 있는 거대한 혀가 있었음을 나타낸다. 이는 삼키기 과정에서 구개(입천장) 치아와 함께 작동했을 것이다. 혀는 큰 구개 치아로 음식을 뚫기 위해 식물 조각을 구개에 눌렀을 것이고, 이 행위는 장내에서 음식의 섬유소 분해 발효를 강화하는 역할을 했을 수 있다.[4][3]
코틸로린쿠스의 이빨에 있는 적은 수의 작은 돌기(3개)는 이 속이 다른 초식성 카세아사우루스과 동물(각각 5개, 5~8개, 5~7개의 작은 돌기를 가진 ''안젤로사우루스'', ''유로미크테르'' 및 ''에나토사우루스'')과 다른 종류의 먹이, 혹은 다른 범위의 먹이에 적응했음을 나타낸다.[3]
초식동물이었으며[52], 몸통에 많은 식물을 삼켜 시간을 들여 발효하고 소화했을 것으로 보인다. 이빨은 가늘고 길었으며, 이것으로 양치류 등의 식물을 잘게 찢어 먹었을 것으로 보인다. 머리가 작고 목이 짧아서 땅에 입을 가까이 댈 수 없었다.[52]
4. 2. 생활 방식

에버렛 C. 올슨은 골격의 골화 정도, 상대적으로 짧은 발과 손, 거대한 발톱, 매우 강력한 신전근을 가진 사지, 그리고 튼튼한 천골을 근거로 ''코틸로린쿠스''를 포함한 카세이드가 주로 육상 동물이었을 것이라고 보았다. 올슨은 카세이드가 물속에서 시간을 보냈을 가능성을 배제하지 않았지만, 육상에서의 이동을 그들의 생활 방식의 중요한 측면으로 간주했다.[4]
매우 강력하고 힘줄이 많은 신전근을 가진 앞다리와 매우 거대한 발톱은 뿌리나 덩이줄기를 파는 데 사용되었을 수 있다는 가설이 제시되었다.[4] 그러나 매우 짧은 목은 머리의 수직 운동 진폭이 작아 대형 종이 지면에서 먹이를 먹는 것을 불가능하게 만들었다.[20] 다른 가설은 카세이드가 강력한 앞다리를 사용하여 큰 식물을 자신 쪽으로 접어 강력한 발톱으로 찢어냈을 수 있다는 것이다.[4]
일부 학자들은 ''코틸로린쿠스''와 같은 카세이드가 강력한 발톱을 가진 사지를 사용하여 포식자나 생식과 관련된 종내 활동으로부터 자신을 방어했을 수 있다고 주장한다. 올슨은 이와 관련하여 ''코틸로린쿠스 한코키'' 종의 알려진 표본 거의 모두가 생존 기간 동안 1개에서 10개의 갈비뼈가 부러지고 치유되었다는 점이 흥미롭다고 언급했다.[10][4] 또한, 일부 학자들은 대형 파생 카세이드가 큰 발톱을 가진 손을 노처럼 사용하여 먹이를 먹는 식물을 조작할 수도 있는 반수생 동물이었을 것이라고 보았다.[20]
2016년 람베르츠와 동료들은 ''코틸로린쿠스''와 같은 대형 카세이드의 육상 생활 방식에 의문을 제기했다. 이들은 성체 및 미성숙 ''코틸로린쿠스'' 표본의 뼈 미세 구조가 육상 동물보다는 수생 동물과 유사하며, 매우 해면질 뼈 구조, 극도로 얇은 피질, 뚜렷한 골수강의 부재를 보인다는 것을 보여주었다. 이러한 낮은 골밀도는 수백 킬로그램의 무게를 가진 육상 동물에게 불리했을 것이다. 또한 람베르츠 등은 척추뼈와 등쪽 갈비뼈 사이의 관절이 갈비뼈의 움직임을 제한하여 환기를 제한한다는 것을 발견했다. 이를 극복하기 위해 그들은 특히 수생 환경에서 호흡을 돕기 위해 원시 횡격막이 존재했을 것이라고 제안했다. 이들 저자는 또한 카세이드 서식지가 해당하는 건조한 고기후가 이러한 동물의 반수생 생활 방식과 양립할 수 없지 않다고 주장했다. 이러한 고생태는 상당한 수의 수생 서식지(강, 호수 및 석호)를 포함했다. 건조한 조건은 동물들이 때때로 모여 결국 죽게 된 이유일 수 있으며, 식물의 밀도가 낮아 먹이를 찾기 위해 더 많은 이동 노력이 필요했을 것이다. 람베르츠 등에 따르면 ''코틸로린쿠스''와 같은 대형 카세이드는 주로 수생 동물이었으며, 번식이나 체온 조절의 목적으로만 육지에 나왔다.[20]
그러나 이러한 가설은 고생물학적 및 매몰학적 데이터를 바탕으로 하고, 대형 카세이드에게 수생 생활 방식에 대한 형태학적 적응이 없다는 점을 고려하여 케네스 앤젤치크와 크리스티안 카머러, 그리고 로버트 라이스와 동료들에 의해 논쟁이 제기되었다. 앤젤치크와 카머러에 따르면, 람베르츠 등이 확인한 카세이드의 낮은 골밀도는 반수생 동물의 골밀도와 유사하지 않으며, 반수생 동물은 수동적인 부력 조절과 조류 및 파도에 대한 안정성을 제공하기 위해 더 강력하게 골화된 골격을 갖는 경향이 있다. ''코틸로린쿠스''의 뼈 미세 구조는 원양에 사는 고래와 기각류와 같은 동물에서 보이는 것과 더 유사하며, 높은 기동성, 빠른 가속 및 부력에 대한 유체 역학적 제어를 강조한다. 그러나 카세이드의 형태는 원양 생활 방식과 완전히 양립할 수 없었다. 따라서 이러한 특이한 데이터로 인해 앤젤치크와 카머러는 현재 증거가 카세이드의 육상 생활 방식에 의문을 제기하기에는 여전히 불충분하다고 생각한다.[21] 라이스와 동료들에 따르면, 양서류 ''브라키덱테스''의 많은 골격이 하계 휴면 상태로 보존되어 있고, 폐어 ''그나토리지자''가 하계 휴면을 하는 다른 잘 알려진 동물이라는 점, 그리고 의무적인 수생 척추 동물이 없다는 사실을 통해 헤네시 동물군은 주기적으로 몬순의 영향을 받는 건조한 서식지에서 살았음을 알 수 있다. ''코틸로린쿠스''가 수생 생활 방식에 대한 형태학적 적응을 보이지 않는다는 사실과 결합하여, 이들은 이 동물을 몬순 비를 견뎌야 했던 육상 동물로 간주하며, 일부 개체는 가끔 대규모 홍수에 굴복했다.[3]
2022년 베르네부르크와 동료들은 약간 다른 반수생 생활 방식을 제안했는데, ''랄리에우도린쿠스''(뼈 조직이 ''코틸로린쿠스''보다 더 골다공증을 띤다)와 같은 대형 카세이드는 현대의 하마와 생태적 동등물을 이루며, 물에서 시간을 보내지만(수영 동물보다는 물속 보행 동물), 먹이를 구하기 위해 육지에 나온다는 것이다.[2]
''코틸로린쿠스''는 초식동물이었으며,[52] 몸통에 많은 식물을 삼켜 시간을 들여 발효하고 소화했을 것으로 보인다. 이빨은 가늘고 길었으며, 이것으로 양치류 등의 식물을 잘게 찢어 먹었을 것으로 보인다. 머리가 작고 목이 짧아서 땅에 입을 가까이 댈 수 없었다.[52] 매우 둔한 생물이었지만, 그 크기 때문에 포식자를 끌어들일 일은 없었을 것으로 보인다. 또한, 그 거대한 몸통은 열적 관성을 높여 체온을 유지하기 위한 적응이었을 것이다.[53]
2016년 연구에서는 호흡기의 구조 등으로 코틸로린쿠스가 반수생일 가능성이 있다고 발표되었지만,[54] 그 특징이 수생 적응에 의한 것인지 의심스러워 보이며, 홍수에 견딜 수 있었지만 주로 육상 동물이었을 것이라고 여겨진다.[50]
5. 계통 발생
계통 발생학 연구에 따르면 케이스과(Caseidae)에 대한 모든 연구는 ''코틸로린쿠스''(Cotylorhynchus)를 ''에나토사우루스(Ennatosaurus)''와 ''안젤로사우루스(Angelosaurus)'' 속과 가까운 분류군으로 간주한다.[17] 2008년에 발표된 케이스과에 대한 최초의 계통 발생학적 분석에서, 종 ''코틸로린쿠스 로메리''(Cotylorhynchus romeri)는 ''안젤로사우루스 돌라니(Angelosaurus dolani)''의 자매군으로 밝혀졌다.[17]
2012년 Benson이 수행한 또 다른 계통 발생학적 분석에서는 ''코틸로린쿠스 로메리''를 두 종인 ''C. Hancocki''와 ''C. bransoni''의 자매군으로 확인했다.[18]
2015년, Romano와 Nicosia는 매우 단편적인 분류군인 ''알리에라사우루스 론키''와 ''안젤로사우루스 그린니''를 제외하고 거의 모든 케이스과를 포함하는 최초의 분류학적 연구를 발표했다. 이 분석에서 ''코틸로린쿠스''의 세 종은 ''루테노사우루스(Ruthenosaurus)'' 속과 함께 한 분지를 형성하며, 이 분지는 ''안젤로사우루스''와 ''에나토사우루스'' 속을 포함하는 분지의 자매군이다.[7]
2020년, Berman과 동료들이 발표한 두 개의 분류도 역시 ''코틸로린쿠스''를 가장 파생된 케이스과 중 하나로 분류했다. 첫 번째 분류도에서 ''코틸로린쿠스''의 세 종은 ''안젤로사우루스''와 ''알리에라사우루스''와 함께 해결되지 않은 분기도를 형성한다. 두 번째 분류도에서, ''코틸로린쿠스 헨코키''와 ''C. 브랜소니''는 자매 분류군이며, ''코틸로린쿠스 로메리''와 ''알리에라사우루스''와 함께 분기도를 형성한다.[19]
2022년 Werneburg와 동료들이 발표한 계통 발생학적 분석에 따르면, ''코틸로린쿠스'' 속은 측계통군일 것으로 보인다. 이 연구자들에 따르면, 종 ''코틸로린쿠스 헨코키''와 ''C. 브랜소니''는 이 속에 속하지 않으며, 1960년대 이후로 이 분류군이 연구되지 않았기 때문에 그들의 지위를 명확히 하기 위해 상세한 재검토가 필요할 것이다. 이 분석에서, 모식종 ''C. 로메리''는 ''안젤로사우루스'' 속 바로 위에 위치하며, ''루테노사우루스''와 ''케이스옵시스''를 포함하는 분지 및 ''알리에라사우루스'', 다른 두 종의 ''코틸로린쿠스'', 그리고 ''라리에우도린쿠스''를 포함하는 또 다른 분지와 함께 분기도를 형성한다. 후자의 분지 내에서, ''알리에라사우루스''는 ''“코틸로린쿠스” 브랜소니''의 자매군이며, ''라리에우도린쿠스''와 ''“코틸로린쿠스” 헨코키''를 포함하는 더욱 파생된 분지이다.[2]
6. 분포 및 지질 시대
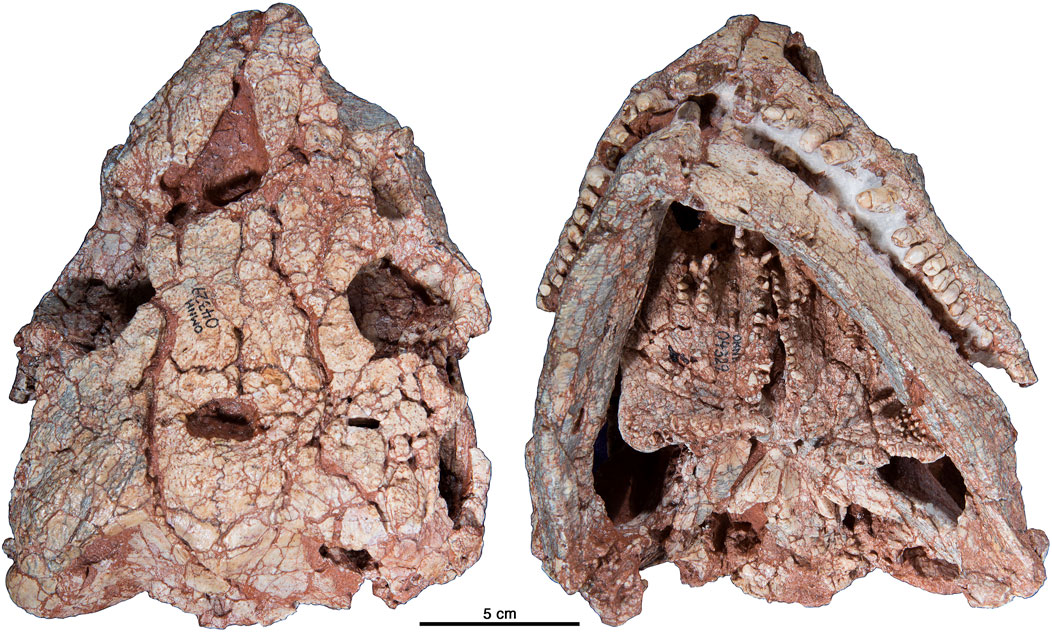
모식종인 ''코틸로린쿠스 로메리''(Cotylorhynchus romeri)는 오클라호마 로건 군 나비나 인근 헤네시 층 하부의 붉은 이암에서 발견된 OMNH 00637표본(두개골의 오른쪽 부분, 불완전한 쇄골간, 오른쪽 및 왼쪽 손)을 바탕으로 명명되었다.[1][4] 모식 표본 발견 직후, 노먼 마을 주변, 클리블랜드 군의 헤네시 층에서 약 20곳의 추가 표본이 발견되었다. 이들은 몇몇 비교적 완전한 골격과 더 많은 단편들을 포함하여 총 40개체 정도이다.[4] 두 지역 표본은 거의 동시대에 살았으며, 두께의 층서 간격 내에서만 알려져 있다. 나비나의 모식 표본은 헤네시 층 기저부에서 약 위에 있는 층에서, 노먼 지역 표본들은 층 기저부에서 사이에 위치한 여러 층에서 발견되었다.[4]
''코틸로린쿠스 핸코키''(C. hancocki)는 텍사스주 하디먼 군 피스 강 근처 샌 안젤로 층 상부에서 발견된 FMNH UR 154 표본(오른쪽 상완골과 경골의 근위 단부)에서 유래되었다.[9] 이후, 텍사스주 녹스 군의 여러 지역, 특히 칸 채석장에서 60개 이상의 표본(고립된 뼈부터 거의 완전한 골격까지)이 발견되었다.[10][4]
''코틸로린쿠스 브란소니''(Cotylorhynchus bransoni)는 오클라호마주 킹피셔 카운티 오메가 채석장에서 발견된 다수의 뼈를 바탕으로 명명되었다. 이 화석은 원래 플라워포트 층 중부에서 발견된 것으로 묘사되었으나,[15] 이후 치카샤 층(엘 레노 군 군)의 일부이며, 플라워포트, 블레인, 도그 크릭 형성의 퇴적물과 부분적으로 겹쳐진다고 수정되었다.[16][4]
''코틸로린쿠스'' 화석은 북아메리카 남부, 텍사스 주와 오클라호마 주에서 발견된다.
참조
[1]
논문
"''Cotylorhynchus romeri'', a new genus and species of pelycosaurian reptile from Oklahoma"
[2]
논문
A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France)
https://hal.science/[...]
[3]
논문
Cranial Anatomy of the Caseid Synapsid ''Cotylorhynchus romeri'', a Large Terrestrial Herbivore From the Lower Permian of Oklahoma, U.S.A
[4]
논문
The family Caseidae
[5]
논문
Review of the pelycosauria
[6]
논문
The Postcranial Skeleton of the Giant Permian Pelycosaur ''Cotylorhynchus romeri''
[7]
논문
Cladistic analysis of Caseidae (Caseasauria, Synapsida): using the gap-weighting method to include taxa based on incomplete specimens
[8]
논문
Cladistic analysis of Caseidae (Caseasauria, Synapsida): Using the gap-weighting method to include taxa based on incomplete specimens
2015
[9]
논문
The San Angelo formation, Permian of Texas and its vertebrates
[10]
논문
Late Permian terrestrial vertebrates, U.S.A and U.S.S.R.
[11]
논문
A huge caseid pelycosaur from north-western Sardinia and its bearing on European Permian stratigraphy and palaeobiogeography
[12]
논문
"''Alierasaurus ronchii'', gen. et sp. nov., a caseid from the Permian of Sardinia, Italy"
[13]
논문
New material of ''Alierasaurus ronchii'' (Synapsida, Caseidae) from the Permian of Sardinia (Italy), and its phylogenetic affinities
[14]
논문
New basal synapsid discovery at the Permian outcrop of Torre del Porticciolo (Alghero, Italy)
[15]
논문
Vertebrates from the Flowerpot formation, Permian of Oklahoma, Part I of Permian Vertebrates from Oklahoma and Texas
http://ogs.ou.edu/do[...]
[16]
논문
New Permian Vertebrates from the Chickasha formation in Oklahoma
http://ogs.ou.edu/do[...]
[17]
논문
Cranial anatomy of ''Ennatosaurus tecton'' (Synapsida: Caseidae) from the Middle Permian of Russia and the evolutionary relationships of Caseidae
[18]
논문
Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies
[19]
논문
New primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany
[20]
논문
A caseian point for the evolution of a diaphragm homologue among the earliest synapsids
[21]
서적
Handbook of Zoology : Mammalian Evolution, Diversity and Systematics
de Gruyter
[22]
논문
Early Permian Vertebrates of Oklahoma
http://www.ogs.ou.ed[...]
[23]
서적
Non-Marine Permian Biostratigraphy and Biochronology
Geological Society, Special Publication 265
[24]
논문
Late Paleozoic–early Mesozoic continental biostratigraphy — Links to the Standard Global Chronostratigraphic Scale
[25]
논문
The age of North America's youngest Paleozoic continental vertebrates : a review of data from the Middle Permian Pease River (Texas) and El Reno (Oklahoma) Groups
[26]
논문
Age and duration of Olson's Gap, a global hiatus in the Permian tetrapod fossil record
http://permian.strat[...]
[27]
논문
The reptile ''Macroleter'': First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia
[28]
논문
A new varanopid synapsid from the early Permian of oklahoma and the evolutionary stasis in this clade
[29]
논문
Extending the footprint record of Pareiasauromorpha to the Cisuralian : earlier appearance and wider palaeobiogeography of the group
[30]
논문
The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism
2001
[31]
논문
Tectonics of the Tethysides: orogenic collage development in a collisional setting
1987
[32]
논문
Depositional Systems, San Angelo Formation (Permian), North Central Texas – Facies Control of Red-Bed Copper Mineralization
1974
[33]
서적
Mammal-like reptiles and the origin of Mammals
Academic Press
[34]
서적
Tectonics, Sedimentary Basins, and Provenance: A Celebration of William R. Dickinson's Career
Geological Society of America Special Paper 540
[35]
논문
New and little known genera and species of vertebrates from the lower Permian of Oklahoma
http://archive.org/d[...]
1970
[36]
논문
A New Genus of Captorhinid Reptile (Amniota: Eureptilia) from the Lower Permian Hennessey Formation of Central Oklahoma, and a Consideration of Homoplasy in the Family Captorhinidae
2021
[37]
논문
Taxonomic revision of the Lysorophia, Permo-Carboniferous lepospondyl amphibians
http://digitallibrar[...]
1991
[38]
논문
Cranial Morphology of the Carboniferous-Permian Tetrapod ''Brachydectes newberryi'' (Lepospondyli, Lysorophia) : New Data from µCT
2016
[39]
논문
Micro-CT Study of ''Rhynchonkos stovalli'' (Lepospondyli, Recumbirostra) with Description of Two New Genera
2016
[40]
논문
Amphibian and reptile tracks from the Hennessey Formation (Leonardian, Permian), Oklahoma County, Oklahoma
http://ogs.ou.edu/do[...]
2002
[41]
논문
The Artinskian Warming Event : an Euramerican change in climate and the terrestrial biota during the early Permian
2022
[42]
논문
Tetrapod footprints from the uppermost level of the Permian Ikakern Formation (Argana basin, western High Atlas, Morocco)
2010
[43]
논문
''Hyloidichnus'' trackways with digit and tail drag traces from the Permian of Gonfaron (Var, France): New insights on the locomotion of captorhinomorph eureptiles
2021
[44]
논문
Climatic and paleogeographic significance of eolian sediment in the Middle Permian Dog Creek Shale (Midcontinent U.S.)
2014
[45]
논문
An Early Permian flora with Late Permian and Mesozoic affinities from north-central Texas
2001
[46]
서적
Wetlands through time
The Geological Society of America Special Paper 399
[47]
논문
Reappraisal of the Permian dissorophid ''Fayella chickashaensis''
[48]
논문
Discussion and reply : The reptile ''Macroleter'': First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia – Reply
[49]
논문
A reevaluation of the enigmatic Permian synapsid ''Watongia'' an dits stratigraphic significance
[50]
논문
Cranial Anatomy of the Caseid Synapsid Cotylorhynchus romeri, a Large Terrestrial Herbivore From the Lower Permian of Oklahoma, U.S.A
https://www.frontier[...]
2022
[51]
논문
The Oldest Caseid Synapsid from the Late Pennsylvanian of Kansas, and the Evolution of Herbivory in Terrestrial Vertebrates
https://www.ncbi.nlm[...]
2014-04-16
[52]
서적
リアルサイズ古生物図鑑 古生代編
[53]
서적
哺乳類型爬虫類 : ヒトの知られざる祖先
[54]
논문
A caseian point for the evolution of a diaphragm homologue among the earliest synapsids
https://pubmed.ncbi.[...]
2016-12
[55]
논문
[56]
웹인용
http://palaeos.com/vertebrates/synapsida/caseasauria.html
http://palaeos.com/v[...]
2012-08-27
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com