트리낙소돈
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
트리낙소돈은 약 50cm 크기의 털로 덮인 작은 수수류로, 육식에 적합한 이빨을 가지고 곤충, 작은 초식동물, 무척추동물을 주로 먹었을 것으로 추정된다. 턱뼈의 수염 흔적, 늑골 소실로 인한 효율적인 호흡, 몸을 둥글게 마는 습성 등 다양한 특징을 보이며, 두개골과 치열 연구를 통해 포유류로의 진화 과정을 보여주는 중요한 화석으로 평가받는다. 남아프리카와 남극에서 화석이 발견되었으며, 굴을 파고 생활하는 습성이 있었다.
더 읽어볼만한 페이지
- 1894년 기재된 화석 분류군 - 드리오사우루스
쥐라기 후기 북아메리카에 서식한 드리오사우루스는 긴 목과 다리, 각질 부리를 가진 최대 3m 크기의 조각류 공룡으로, 모리슨 층 등 다양한 지역에서 화석이 발견되었으며, 1894년에 'Dryosaurus altus'로 재분류되었다. - 1894년 기재된 화석 분류군 - 오르니토수쿠스
오르니토수쿠스는 후기 트라이아스기에 살았던 육식 파충류로, 악어와 관련이 있으며 공룡처럼 뒷다리로 걸을 수 있고, 수각류 공룡과 유사한 두개골, 다섯 개의 발가락, 등뼈를 따라 두 줄의 갑판을 가진 것이 특징이며, 오르니토수쿠스과의 모식 속이고, 공룡상목의 조상으로 여겨진다. - 상키노돈트류 - 게일사우루스과
게일사우루스과는 상키노돈트 중 가장 원시적인 키노돈트 무리이며, 2차 구개, 넓은 치아, 큰 측두창과 넓은 주둥이를 가진 것이 특징이고 다른 키노돈트처럼 직립보행을 했을 가능성이 높다. - 상키노돈트류 - 게일사우루스
게일사우루스는 약 2억 5천만 년 전 트라이아스기에 남아프리카에서 화석이 발견된 멸종된 키노돈트의 일종이며, 뻗은 자세를 가졌고 가냘픈 형태와 튼튼한 형태의 두 가지 변형이 존재하며, 두개골은 넓고 낮고 12cm까지 자랐다.
| 트리낙소돈 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| |
| 학명 | Thrinaxodon |
| 명명자 | Seeley, 1894 |
| 모식종 | Thrinaxodon liorhinus |
| 모식종 명명자 | Seeley, 1894 |
| 상위 분류군 권위자 | Watson & Romer, 1956 |
| 생물학적 분류 | |
| 계 | 동물계 Animalia |
| 문 | 척삭동물문 Chordata |
| 아문 | 척추동물아문 Vertebrata |
| 상강 | 사지동물상강 Tetrapoda |
| 강 | 단궁강 Synapsida |
| 목 | 수궁목 Therapsida |
| 미분류 | 수궁목 Therapsida 계급 없음 수치류 Theriodontia |
| 아목 | 키노돈아목 Cynodontia |
| 과 | †트리낙소돈과 Thrinaxodontidae |
| 지질 시대 | |
| 존속 시기 | 후기 페름기 - 전기 트라이아스기, |
| 존속 시기 | 트라이아스기 초기 - 중기 |
| 종 | |
| 하위 분류 | T. liorhinus T. brasiliensis |
2. 특징
트리낙소돈은 여우 정도 크기의 작은 동물로, 온몸이 털로 덮여 있었다. 치열을 보면 육식동물이었는데, 주로 곤충, 작은 초식동물, 무척추동물을 먹었을 것으로 보인다. 특히 2차 구개는 비강을 통해 호흡을 중단하지 않고 계속할 수 있게 하여 소화에 중요한 형질이었다.[3]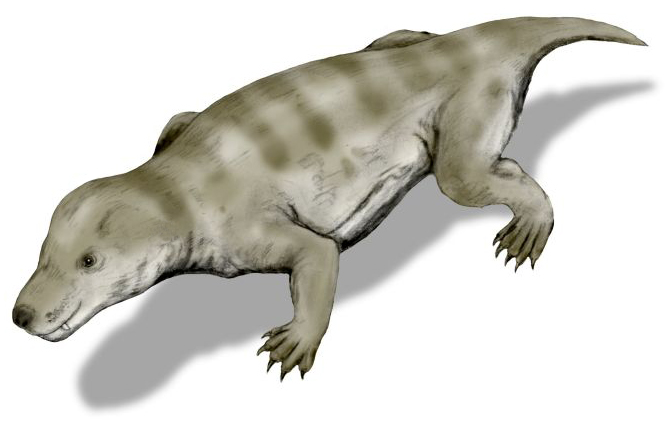
복부 늑골이 소실되어 횡격막에 의해 흉곽과 복곽이 나뉘어 효율적으로 호흡할 수 있었다. 이는 삼첩기 동안 지속된 저산소 상태에 대한 적응으로 생각된다. 몸통을 비틀 수 있게 되어 몸을 둥글게 말아 휴면한 모습 그대로 화석화된 것도 발견되었다. 이는 트리낙소돈을 포함한 키노돈류의 항온 동물화가 진행되어 잠자는 동안 체온 손실을 막기 위한 것으로 추정된다. 따라서 수염뿐만 아니라 털도 있었을 것으로 보인다. 다만, 머리 부분을 중심으로 몸을 둥글게 마는 것은 뱀, 도마뱀, 나방 애벌레 등 변온 동물에게도 흔히 볼 수 있는 휴식 자세이다.
트리낙소돈의 자세는 페름기 파충류의 기어가는 방식에서 현대 포유류와 멸종된 많은 포유류에서 발견되는 좀 더 직립적인 방식으로의 전환을 보여주기 때문에 흥미롭다. 트리낙소돈과 같은 키노돈트는 먼쪽 대퇴골 과가 다리의 나머지 부분에 대해 45도 각도로 나타나도록 관골구와 연결된다. 트리낙소돈은 굴 안에서 초기 포유류와 유사한 직립 자세를 취할 수 있었다.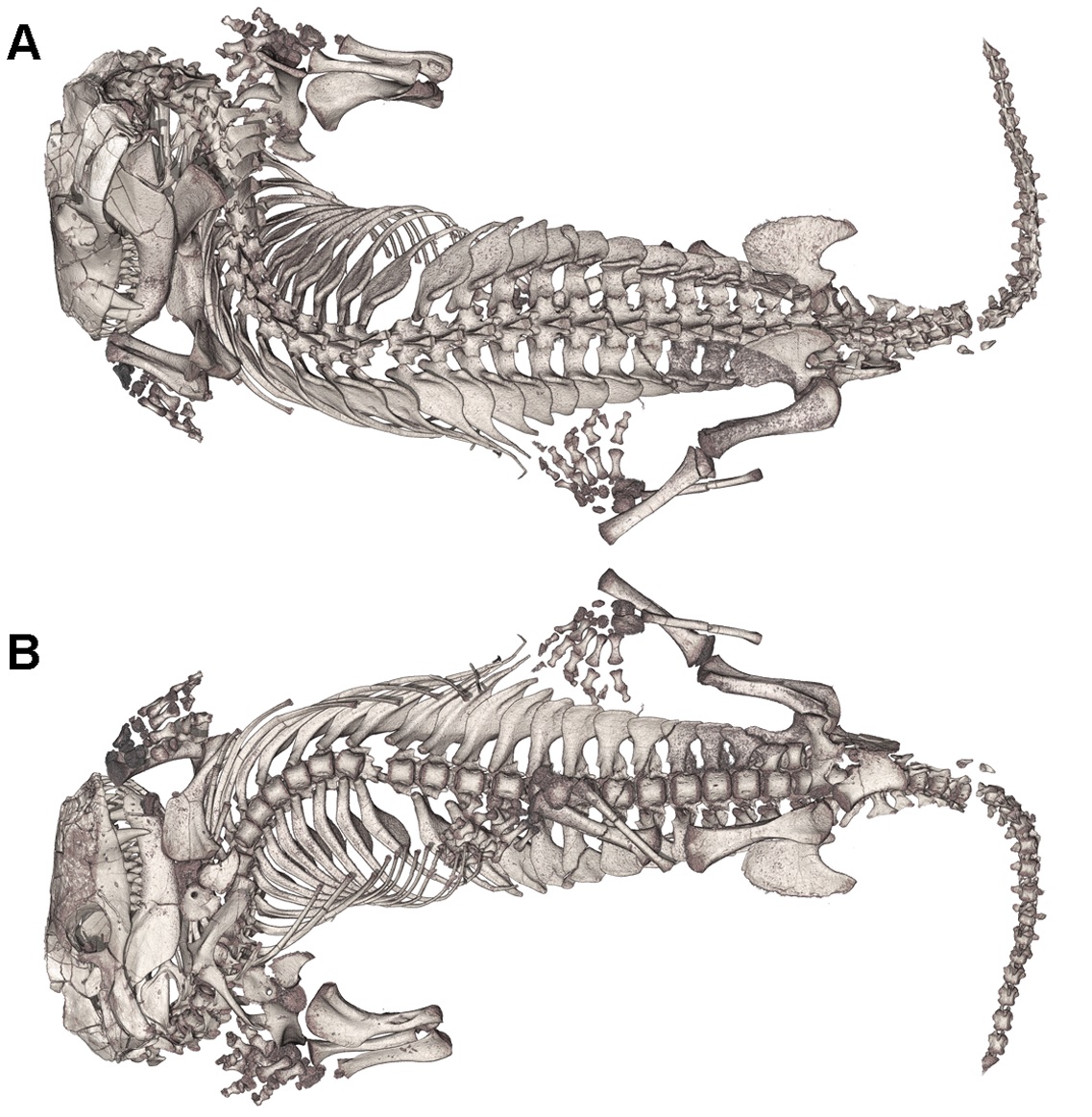
트리낙소돈은 굴속에서 발견된 수많은 증거를 통해 굴을 파고 사는 키노돈트류로 확인되었다. 이 굴은 다른 생물이 남긴 굴을 차지한 것이 아니라, 트리낙소돈이 직접 짓고 살았던 증거가 있다. 가슴, 허리, 천추로 분화된 분절 척추의 진화 덕분에 트리낙소돈은 작은 굴 안에서도 편안하게 쉴 수 있는 유연성을 갖게 되었고, 이는 여름잠이나 동면과 같은 습성으로 이어졌을 수 있다.[11]

2. 1. 두개골
트리낙소돈의 코뼈에는 많은 수의 구멍이 파여 있다. 코뼈는 앞쪽으로 좁아지고 확장되어 앞쪽에서 앞쪽과 직접 연결되며, 전두골, 전두개골, 누골과 연결되지만, 협골이나 안와골과는 상호 작용하지 않는다. 트리낙소돈의 상악골에도 구멍이 많이 파여 있다.[3] 트리낙소돈의 주둥이에 있는 구멍의 배열은 투피남비스와 같은 도마뱀과 유사하며, 하나의 큰 안와하 구멍을 가지고 있다.[3] 따라서 트리낙소돈은 포유류와 같은 움직이고 근육질의 입술이 아닌 도마뱀과 같은 비근육질의 입술을 가졌을 것이다.[3] 안와하 구멍과 관련된 안면 유연성이 없기 때문에 트리낙소돈은 수염을 가지지 못했을 것이다.트리낙소돈의 두개골 상부에서 전두-코뼈 봉합선은 더 원시적인 두개골 형태에서 볼 수 있는 일반적인 횡단 과정 대신 화살표 모양을 나타낸다. 전두골의 약간 앞쪽과 복부에 있는 전두개골은 크기가 매우 작으며, 후안와골, 전두골, 코뼈, 누골과 접촉한다. 두개골 뒤쪽에서 두정골에는 시상 능선이 없다. 두개골 천장은 두정골 구멍 바로 뒤쪽에서 가장 좁으며, 모양은 거의 원형이다. 측두 능선은 두개골 전체 길이에서 상당히 분리되어 있다. 측두 창은 골화된 근막과 함께 발견되어 측두근 부착의 증거를 제공한다.[3]
상악에는 비강을 입의 나머지 부분과 분리하는 이차 구개가 있어 트리낙소돈은 입에 음식을 보관하더라도 방해받지 않고 숨을 쉴 수 있었다. 이러한 적응을 통해 트리낙소돈은 음식을 더 많이 으깰 수 있어 소화에 필요한 시간을 줄일 수 있었다. 상악골과 구개골은 중간에서 만나 상악에 중간 봉합선을 형성한다. 상악구개 봉합선에는 후부 구개 구멍도 포함되어 있다. 트리낙소돈의 서골의 큰 구개 지붕 구성 요소는 코안 또는 내부 비강 바로 등쪽에 있다. 익상골은 상악에서 확장되어 부비골의 칼 모양 돌기 각 측면에 존재하는 작은 익상골 간극을 둘러싼다. 부비골과 기저접형골은 융합되어 있지만, 융합된 뼈의 가장 앞쪽/등쪽 끝에서는 기저접형골의 섬유주 부착에 약간의 분리가 있다.[3]

이골 부위는 측두 창을 둘러싼 부위로 정의된다. 가장 주목할 만한 것은 난원창 바로 앞의 깊은 함몰의 증거로, 두개골과의 평활근 상호 작용의 증거가 포함되어 있다. 이러한 평활근 상호 작용은 고막을 나타내는 것으로 해석되며, 이 함몰부가 난원창과 함께 트리낙소돈의 귀의 기원을 나타낸다는 것을 암시한다. 이것은 트리낙소돈에서 발생하여 후기 키노돈트를 통해 보존된 새로운 공유파생형질이다. 등자뼈에는 두꺼운 연골 플러그가 들어 있었고, 이는 난원창의 측면에 맞았지만, 등자뼈의 관절 끝의 절반만 난원창을 덮을 수 있었다. 이 구멍의 나머지 부분은 "비골화된" 영역으로 열려 있으며, 달팽이관 함몰에 어느 정도 접근하여 내이 관절이 이 영역 내에서 직접적으로 발생했다고 추측하게 한다.[3]
트리낙소돈의 두개골은 시냅시드 두개골의 시간이 지남에 따른 단순화를 뒷받침하는 중요한 과도기적 화석이다. 뼈 수의 가장 두드러진 감소는 트리낙소돈과 프로바이노그나투스 사이에서 발생했으며, 이러한 변화는 너무 극적이어서 이 특정 전환에 대한 화석 기록이 불완전할 가능성이 높다. 트리낙소돈은 페리코사우루스 조상보다 두개골에 뼈가 적다.[4]
턱뼈에는 다수의 작은 구멍이 나 있으며, 아마도 수염이 나 있던 흔적이라고 추정된다. 아래턱의 이를 수납하는 치골은 테로케팔루스류 등보다 더욱 확대되었으며, 각골과 관절골은 축소되었다.
2. 2. 치열
''트리낙소돈''의 치열에 대한 데이터는 30mm에서 96mm 사이의 다양한 ''트리낙소돈'' 두개골 표본을 마이크로 CT 스캐너를 사용하여 수집되었다.[5] 이러한 치열 패턴은 모르가누코돈과 유사하여 트리낙소돈과에서 기원하여 초기 포유류의 기록으로 확장되었다고 추정할 수 있다.성체 ''T. liorhinus''는 위턱의 각 측면에 4개의 앞니, 1개의 송곳니, 6개의 뒷송곳니를 가지며, 아래턱에는 각 측면에 3개의 앞니, 1개의 송곳니, 7~8개의 뒷송곳니를 갖는다. 따라서 성체 ''트리낙소돈''은 총 44~46개의 치아를 가지고 있었다.[5]
''T. liorhinus''의 위턱 앞니는 뒤쪽으로 향하는 교두(cusp)를 가지며, 가장 먼 지점에서 굽고 뾰족하다. 전상악골(premaxilla)에 삽입되는 근위 지점에 도달하면서 더 넓고 둥글게 된다. 네 번째 위턱 앞니는 형태적으로 작은 송곳니와 비슷하지만, 기능적인 송곳니가 되기에는 너무 앞쪽에 위치하여 수렴 진화의 예로 간주될 수 없다. 아래턱 앞니는 매우 넓은 기저부를 가지며, 치아의 끝으로 갈수록 점차 좁아진다. 아래턱 앞니의 설면(혀쪽 면)은 대부분 오목하고, 순면(볼쪽 면)은 종종 볼록하며, 앞쪽으로 향한다. 단, 세 번째 아래턱 앞니는 경우에 따라 더 배측-복측(등쪽-배쪽) 방향을 가질 수 있다. 앞니는 대부분 넓고 원뿔 모양의 단일 기능 치아이다.
''T. liorhinus''의 송곳니는 표면에 작은 배측-복측 방향의 면을 가지고 있는데, 이는 교합(위턱과 아래턱이 닫힐 때의 치열 정렬)과 관련이 있는 것으로 보인다. 각 송곳니는 기존 송곳니의 뒤쪽에 위치한 교체 송곳니를 턱 안에 가지고 있으며, 교체 송곳니와 기능성 송곳니는 모두 톱니 모양의 가장자리가 없고 작은 면만 있다. 아래턱 송곳니는 거의 수직(배측-복측)으로 향하는 반면, 위턱 송곳니는 약간 앞쪽으로 향한다.[5]

''T. liorhinus''의 위턱과 아래턱의 뒷송곳니는 몇 가지 공통적인 특징을 공유하지만 서로 상당히 다르다. 첫 번째 뒷송곳니(송곳니 바로 뒤)는 다른 뒷송곳니보다 작고 이첨두(bicuspid, 교두가 2개)인 경우가 많다. 첫 번째 뒷송곳니를 포함하여 다른 뒷송곳니가 이첨두라면, 뒤쪽의 부속 교두가 존재하고 해당 치아에는 어떤 치경(cingular) 또는 순측(labial) 교두가 없을 것으로 추정할 수 있다. 그러나 치아가 삼첨두(tricuspid, 교두가 3개)인 경우 치경 교두가 발달할 가능성이 있으며, 이러한 경우 앞쪽 교두가 가장 먼저 나타나고 가장 두드러진 교두가 된다. 위턱 뒷송곳니에는 3개 이상의 교두를 가진 치아가 없어야 하며, 위턱 뒷송곳니에는 순측 교두가 나타나지 않는다.
어린 ''트리낙소돈''의 위턱 뒷송곳니 대부분은 이첨두이며, 이 중 하나만 삼첨두이다. 중간(어린 개체와 성체 사이) ''트리낙소돈''의 위턱 뒷송곳니는 모두 삼첨두이며 순측 또는 치경 교두가 없다. 성체 위턱 뒷송곳니는 중간 시기의 특징을 유지하며 삼첨두 치아만 가지고 있다. 그러나 이러한 성체 치아에서 치경 교두가 발달할 수 있다. 가장 마지막(뒤쪽) 위턱 송곳니는 전체 턱 시스템에서 모든 송곳니 중 가장 작은 경우가 많다. ''트리낙소돈''의 어린 개체와 중간 형태의 아래턱 뒷송곳니에 대해서는 알려진 바가 거의 없지만, 성체 아래턱 뒷송곳니는 모두 여러 개(3개 이상)의 교두와 순측 교두가 유일하게 나타난다. 일부 오래된 표본에서는 여러 개의 교두가 없는 아래턱 송곳니가 발견되었는데, 이는 노화 또는 치아 교체의 반응일 수 있다.[5]
''트리낙소돈''은 키노돈트에서 치아 교체의 첫 번째 사례 중 하나를 보여준다. 이것은 앞니와 뒷송곳니에서 기능 치아의 설쪽(혀쪽)에 위치한 교체 구덩이의 존재로 구별되었다. 교체 송곳니가 존재하지만, 대부분의 경우 맹출되지 않고 원래의 기능적 송곳니가 남아있다.[5]
턱뼈에는 다수의 작은 구멍이 나 있으며, 아마도 수염이 나 있던 흔적이라고 한다. 아래턱의 이를 수납하는 치골은 테로케팔루스류 등보다 더욱 확대되었으며, 각골과 관절골은 축소되었다.
2. 3. 골격 조직학
''트리낙소돈''의 뼈 조직은 섬유판상골로 구성되어 있으며, 모든 사지에서 정도의 차이가 있지만 대부분 주변부로 갈수록 평행 섬유 뼈 조직으로 발달한다. 각 뼈에는 다수의 가지가 있는 소관을 방사하는 풍부한 구형 골세포 소강이 포함되어 있다. 발생학적으로 초기에 형성된 뼈는 대부분 섬유판상 조직으로 구성되어 있으며 많은 수의 혈관관을 가지고 있었다. 이 관들은 방사형 문합을 포함하는 일차 골원 내에서 종 방향으로 정렬되어 있다. 대부분 평행 섬유 뼈 조직으로 구성된 영역은 인접한 섬유판상 조직에 비해 단순 혈관관이 거의 없다. 평행 섬유 주변 뼈 조직은 뼈 성장이 둔화되기 시작했음을 나타내며, 이러한 성장의 변화가 해당 표본의 나이 때문이라는 가정을 가져온다. 이것을 성체 ''T. liorhinus''의 주변부에서 골세포 소강의 더 큰 조직화와 결합하면, 이 생물이 성체에 빠르게 도달하기 위해 매우 빠르게 성장했다는 가설에 접근하게 된다.[10]''트리낙소돈''의 대퇴골 내에는 평행 섬유 조직으로 이루어진 주요 영역이 없지만, 중간 피질 내에 작은 평행 섬유 뼈 고리가 있다. 대퇴골의 나머지 부분은 섬유판상 조직으로 구성되어 있지만, 구형 골세포 소강은 훨씬 더 조직화되고 일차 골원은 주변 골막 표면에 가까워질수록 다른 많은 뼈보다 혈관이 적어진다. 대퇴골에는 골 소주가 거의 없다. 상완골은 대퇴골과 여러 면에서 다르며, 그 중 하나는 뼈의 수질강 근처에 더 광범위한 골 소주 네트워크가 있다는 것이다. 구형 골세포 소강은 상완골의 중간부에 가까워질수록 더 평평해진다. 혈관이 존재하지만 상완골에는 이차 골원이 없다. ''트리낙소돈''의 요골과 척골은 대략 동일한 조직학적 패턴을 나타낸다. 상완골 및 대퇴골과 달리, 평행 섬유 영역은 앞다리의 원위 골에서 훨씬 더 뚜렷하다. 수질강은 여러 층의 혈관화가 매우 불량한 골내판 조직과, 골간단 부위의 수질강 근처에 매우 큰 공동으로 둘러싸여 있다.[10]
트리낙소돈의 뼈 조직학적 분석 결과, 어린 개체 발달 동안 매우 빠른 뼈 성장을 보였고, 성체 기간 동안 훨씬 느린 발달을 보여 트리낙소돈이 생애 초기에 최대 크기에 도달했을 가능성이 높다는 것을 나타낸다.[10]
3. 생태
트리낙소돈은 여우만한 크기의 작은 동물로, 온몸이 털로 덮여 있었다. 치열을 보면 육식동물이었는데, 이들은 주로 곤충, 작은 초식동물, 무척추동물을 먹었을 것으로 보인다. 특히 2차 구개는 비강을 트리낙소돈이 호흡을 중단하지 않고 계속 할 수 있도록 했으며, 이는 소화에 중요한 형질이다.[2]
트리낙소돈은 언덕이나 강의 범람원 등에 얕은 구멍을 파서 둥지를 만들어 새끼를 키웠던 것으로 보인다. 또한 후각을 이용한 의사소통이 발달하여 마킹 행동으로 각자의 영역을 주장했던 것으로 여겨진다. 굴을 파고 사는 키노돈트류로 확인되었으며, 이들이 직접 굴을 짓고 살았던 증거가 있다. 분절 척추의 진화 덕분에 트리낙소돈은 작은 굴 안에서도 편안하게 쉴 수 있었고, 이는 여름잠이나 동면과 같은 습성으로 이어졌을 수 있다.
3. 1. 자세
''트리낙소돈''의 자세는 도마뱀과 유사한 페름기 파충류의 기어가는 방식에서 현대 포유류와 멸종된 많은 포유류에서 발견되는 좀 더 직립적인 방식으로의 전환을 나타내기 때문에 흥미로운 주제이다. ''트리낙소돈''과 같은 키노돈트에서, 먼쪽 대퇴골 과는 다리의 나머지 부분에 대해 45도 각도로 나타나도록 하는 방식으로 관골구와 연결된다. 이것은 페름기 파충류의 먼쪽 대퇴골 과와 비교했을 때 큰 차이인데, 페름기 파충류는 대퇴골이 지면과 평행하도록 하여 기어가는 듯한 자세를 취하게 한다.[2] 트리낙소돈과에서만 관찰된 적응 현상은 굴 안에서 초기 포유류와 유사한 직립 자세를 취할 수 있게 해준다. 이러한 자세의 변화는 ''트리낙소돈''의 몸통의 생리적 변화에 의해 뒷받침된다. 그러한 변화로는 분절된 늑골 구획이 처음으로 나타나는 것으로, ''트리낙소돈''은 흉추와 요추를 모두 나타낸다. 척추의 흉추 부위에는 큰 늑간판이 있는 늑골이 포함되어 있는데, 이는 보호하거나 등의 주요 프레임을 지지하는 데 도움이 되었을 가능성이 높다. 새롭게 개발된 이 배열은 횡격막을 위한 적절한 공간을 제공했지만, 적절한 연조직 기록이 없기 때문에 횡격막의 존재는 순전히 추측에 불과하다.[11]또한, 몸통을 비틀 수 있게 되었으며, 몸을 둥글게 말아 휴면한 모습 그대로 화석화된 것도 발견되었다. 이것은 트리낙소돈을 포함한 키노돈류의 항온 동물화가 진행되었기 때문에, 잠자는 동안 체온이 손실되지 않도록 하기 위해서라고 추정된다. 이로 인해, 이미 수염뿐만 아니라 털까지 생겨났을 것이라고 한다. 다만, 중요한 기관인 머리 부분을 중심으로 몸을 둥글게 마는 것은 뱀, 도마뱀이나 나방의 애벌레 등 변온 동물에서도 흔히 볼 수 있는 휴식 자세이다.
3. 2. 굴 파기 습성
''트리낙소돈''은 굴속에서 발견된 수많은 증거를 통해 굴을 파고 사는 키노돈트류로 확인되었다. 이 굴들은 다른 생물이 남긴 굴을 단순히 차지한 것이 아니라, 실제로 ''트리낙소돈''이 직접 짓고 살았던 증거가 있다. 가슴, 허리, 천추로 분화된 분절 척추의 진화 덕분에 ''트리낙소돈''은 작은 굴 안에서도 편안하게 쉴 수 있는 유연성을 갖게 되었고, 이는 여름잠이나 동면과 같은 습성으로 이어졌을 수 있다. 이러한 분절 늑골의 진화는 시냅스과 화석 기록에서 횡격막이 처음 나타난 사례일 수 있음을 시사하지만, 적절한 연조직 흔적이 없어 추정에 불과하다.[11]
굴을 파고 사는 ''트리낙소돈''의 가장 초기 발견 사례는 약 2억 5100만 년 전, 즉 페름기-트라이아스기 대멸종 시기에 해당하는 표본이다. 이 화석들은 대부분 남아프리카 공화국의 카루 분지 지역의 범람원에서 발견되었다. 이러한 행동은 선신생대에서 비교적 낮은 빈도로 나타났으며, 테라프시드, 초기 트라이아스기 키노돈트류, 일부 초기 포유류가 주를 이루었다. ''트리낙소돈''은 실제로 발견된 최초의 굴을 파는 키노돈트였으며, 트리라코돈과 유사한 행동 양식을 보였다. 화석 기록상 최초의 굴을 파는 척추동물은 디키노돈트 시냅스과인 ''디익토돈''이었으며, 대멸종 기간 동안 굴을 파는 것이 적응적 이점을 가지면서 이러한 굴을 파는 패턴이 후대 키노돈트류에게 전해졌을 가능성이 있다. ''트리낙소돈''의 굴은 두 개의 옆으로 기울어진 반쪽으로 구성되어 있는데, 이는 굴을 파는 비포유류 키노돈트에서만 관찰되는 패턴이다. ''트리낙소돈''에서 나타난 척추/늑골 해부학적 변화는 동물에게 더 넓은 유연성을 부여했고, 따뜻함을 유지하고 여름잠을 자기 위해 좁은 공간에 적응하여 주둥이를 뒷다리 아래에 놓을 수 있게 했다.
''트리낙소돈''의 굴에서 부상당한 템노스폰딜인 ''Broomistega''가 발견되었다. 이 굴은 싱크로트론을 사용하여 스캔했는데, 이 도구는 손상되지 않은 표본을 손상시키지 않으면서 굴의 내용을 관찰하는 데 사용되었다. 싱크로트론은 부상당한 리네수키드인 ''브룸스테가 putterilli''를 보여주었는데, 이는 부러지거나 손상된 사지와 두개골의 천공 흔적을 보였으며, 이는 다른 육식동물의 송곳니에 의해 생긴 것으로 보인다. 천공 사이의 거리를 문제의 ''트리낙소돈'' 송곳니 사이의 거리와 비교하여 측정했지만, 그러한 관련성은 발견되지 않았다. 따라서 템노스폰딜은 외상 후 굴로 피신했고, ''T. liorhinus''는 템노스폰딜이 결국 죽을 때까지 굴에 머물도록 허용했다고 추정할 수 있다. 종간 쉼터 공유는 화석 기록에서 드문 이례적인 현상이며, 이 ''T. liorhinus''는 화석 기록에서 이러한 유형의 행동이 처음 발생한 사례 중 하나를 보여주지만, 템노스폰딜이 둥지를 틀고 있던 ''트리낙소돈''이 죽기 전인지 후인지 현재 알 수 없다.[13]
언덕이나 강의 범람원 등에 얕은 구멍을 파서 둥지를 만들어 새끼를 키웠던 것으로 보인다. 또한 후각을 이용한 의사소통이 발달하여 마킹 행동으로 각자의 영역을 주장했던 것으로 여겨진다.
4. 분류
트리낙소돈은 키노돈티아(Cynodontia)의 하위 분류군인 에피키노돈티아(Epicynodontia)에 속한다. 키노돈티아는 결국 모르가누코돈(Morganucodon)과 모든 다른 포유류의 진화를 이끌었다. 키노돈티아는 테라프시다(Therapsida)에 속하며, 테라프시다는 시냅시다(Synapsida) 계통에서 최초의 주요 분류군이었다. 시냅시다는 양막류(Amniota) 분류군 아래에서 두 개의 주요 분기점 중 하나를 나타내며, 양막류는 오늘날의 파충류, 조류, 그리고 악어(Crocodilia)를 포함하는 더 큰 분류군인 사우롭시다(Sauropsida)로도 갈라졌다. 트리낙소돈은 인간과 다른 현존하는 포유류로 가는 과정에서 형태학적으로 화석 과도기를 나타낸다.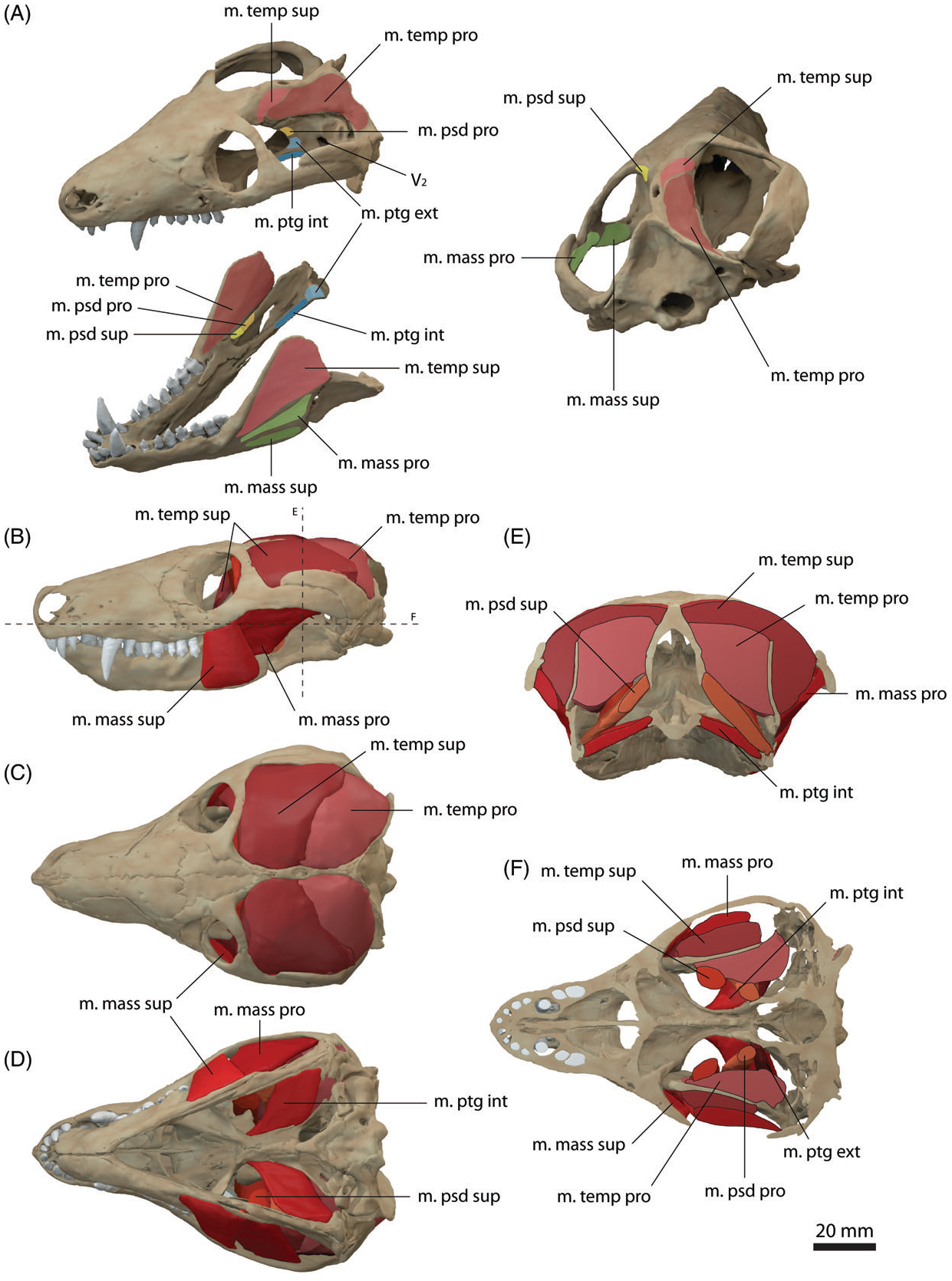
{| class="wikitable"
|-
| 키노돈티아
|
{| class="wikitable"
|-
|
|-
! 에피키노돈티아
|-
|
{| class="wikitable"
|-
| ''키노사우루스(Cynosaurus)''
|-
|
| 게일사우루스(Galesaurus) |
| 프로게일사우루스(Progalesaurus) |
|-
|
{| class="wikitable"
|-
| '''''트리낙소돈'''''
|-
|
{| class="wikitable"
|-
| ''플라티크라니엘루스(Platycraniellus)''
|-
! 유키노돈티아
|-
|
| 키노그나티아(Cynognathia) |
| 프로베이노그나티아(Probainognathia) |
|}
|}
|}
|}
|}
5. 분포

''트리낙소돈''은 원래 남아프리카 보퍼트 그룹의 ''리스트로사우루스 집합소''(Lystrosaurus Assemblage Zone)에서 발견되었다. 처음에는 그 지역에만 고립된 것으로 여겨졌으나, 남아프리카의 노르만디엔 지층(Normandien Formation)과 카트베르그 지층(Katberg Formation)에서 다른 화석들이 발견되었다.[7] 1977년 남극의 프레모우 지층(Fremouw Formation)에서 ''트리낙소돈''의 추가 화석이 발견되기 전까지는 남아프리카 지역에만 분포하는 것으로 알려져 있었다.
남극에서 화석이 발견된 후, 새로운 종인지 아니면 ''T. 리오리누스''의 또 다른 서식지인지 확인하기 위해 여러 실험이 진행되었다. 첫 번째 실험은 남극과 아프리카 ''트리낙소돈''의 평균 척추뼈(추골) 수를 비교하는 것이었다. 데이터에 따르면 아프리카 ''T. 리오리누스''는 26개, 남극 ''트리낙소돈''은 27개의 척추뼈를 가지고 있어 약간의 차이를 보였다. 다른 키노돈트와 비교했을 때 27개의 척추뼈는 화석 기록상 이 하위 구역 전체에서 표준으로 나타났다. 다음으로 두 그룹의 두개골 크기를 비교했으나 차이점이 발견되지 않아, 동일한 종일 수 있다는 첫 번째 증거가 되었다. 마지막으로 늑골을 비교했을 때 늑골 확장에는 약간의 차이가 있었지만, 가장 중요한 공유 파생 형질은 두 그룹 모두에서 늑간판이 서로 겹친다는 점으로 일관성을 보였다.
이러한 평가를 통해 새로운 종이 아니라, ''트리낙소돈''이 오늘날 넓은 바다로 분리된 두 개의 다른 지리적 지역(아프리카 및 남극)에 서식했다는 결론이 내려졌다. 이 발견은 초기 트라이아스기 동안 아프리카와 남극이 연결되어 있었다는 육괴설을 뒷받침하는 여러 발견 중 하나이다.[8] ''트리낙소돈''은 아프리카 및 남극에 서식했으며, 특히 남아프리카 공화국의 카루 분지에서 다수의 화석이 발견되었다.
6. 기타
BBC/군마 현립 자연사 박물관의 '부활하는 공룡·고생물'에서는 트리낙소돈의 두개골이 사진으로 게재되었지만, 실제로는 프로가레사우루스의 두개골이다.[14]
참조
[1]
논문
Earliest evidence of cynodont burrowing
[2]
간행물
Evolution of hindlimb posture in non-mammalian therapsids: biomechanical tests of paleontological hypotheses.
[3]
간행물
Cranial anatomy of the cynodont ''Thrinaxodon'' ''liorhinus''.
Museum of comparative Zoology, Harvard University
[4]
논문
Simplification as a trend in synapsid cranial evolution
[5]
논문
Ontogeny of the Early Cynodont ''Thrinaxodon'' ''liorhinus'' (Therapsida): Dental Morphology and Replacement
[6]
논문
On the Skull and Dentition of a Triassic Saurian (''Galesaurus planiceps'', Ow.)
https://jgs.lyellcol[...]
1887-02
[7]
웹사이트
Thrinaxodon liorhinus
https://paleobiodb.o[...]
2015-03-05
[8]
논문
Triassic Cynodont Reptiles from Antarctica
http://digitallibrar[...]
[9]
논문
Ontogeny of the Early Triassic Cynodont ''Thrinaxodon'' ''liorhinus'' (Therapsida): Cranial Morphology
[10]
간행물
Growth patterns of ''Thrinaxodon'' ''liorhinus'', a non-mammalian cynodont from the lower Triassic of South Africa.
[11]
문서
Note on a new skeleton of ''Thrinaxodon liorhinus''. Abstract.
[12]
논문
Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow
[13]
논문
Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow
[14]
문서
Padrões de diversidade e distribuição de cinodontes não-mamaliaformes do Triássico da América do Sul e África.
[15]
저널
https://royalsociety[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com