헤테로돈토사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
헤테로돈토사우루스는 쥐라기 전기 남아프리카에서 발견된 작은 조반류 공룡이다. 다양한 형태의 이빨을 가진 것이 특징이며, 이빨의 모양에 따라 잡식성, 초식성 또는 육식성 등 식성에 대한 논쟁이 있다. 1962년 처음 발견되었으며, 현재까지 가장 완전한 골격은 1.18m 길이로 추정된다.
더 읽어볼만한 페이지
- 쥐라기 전기 아프리카의 공룡 - 불카노돈
불카노돈은 1969년 짐바브웨에서 발견된 초기 용각류 공룡의 한 속으로, 완전한 네 발 보행을 했으며, 쥐라기 전기, 약 2억 년 전에 살았던 육상 생활에 적응한 용각류의 초기 형태를 보여주는 중요한 화석이다. - 쥐라기 전기 아프리카의 공룡 - 레소토사우루스
레소토사우루스는 초기 쥐라기 시대에 살았던 이족 보행 잡식 공룡으로, 남아프리카 레소토와 남아프리카에서 화석이 발견되었으며, 좁고 뾰족한 두개골과 날카로운 이빨을 가지고 있었다. - 1962년 기재된 화석 분류군 - 고르고돈
고르고돈은 페름기에 북아메리카에 살았던 단궁류로, 이형치아와 등 쪽에 돛을 가진 것이 특징이며, 텍사스에서 처음 발견되어 초식성 식습관 진화 연구에 중요한 사례이다. - 1962년 기재된 화석 분류군 - 녹소사우루스
녹소사우루스는 약 2억 7950만 년에서 2억 6800만 년 전에 북아메리카에서 서식하며 육식성 먹이를 섭취했던 멸종된 비맘류 시냅스 속이다. - 조반목 - 신조반류
신조반류는 1985년 쿠퍼에 의해 명명되었으며, 2021년 공식적으로 정의된 조반목 공룡의 분지군으로, 두꺼운 에나멜층을 가진 아래턱 치아와 장갑류와의 자매군 형성 등의 특징을 가진다. - 조반목 - 스파에로톨루스
스파에로톨루스는 백악기 후기 북미에 살았던 돔 형태 두개골을 가진 후두류 공룡 속으로, 두개골 형태와 뼈 장식 유무로 구별되는 다섯 종이 알려져 있으며, 일부 종의 유효성 논란과 함께 추가 연구가 필요한 분류군이다.
| 헤테로돈토사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Heterodontosaurus |
| 명명자 | Crompton & Charig, 1962년 |
| 종명 | tucki |
| 종 명명자 | Crompton & Charig, 1962년 |
| 이명 | Lycorhinus tucki Thulborn, 1970 |
| 형태 및 크기 | |
| 길이 | 약 1.18 m ~ 1.75 m |
| 무게 | 약 2 kg ~ 10 kg |
| 생물 분류 | |
| 속 | 헤테로돈토사우루스속 (Heterodontosaurus) |
| 과 | 헤테로돈토사우루스과 (Heterodontosauridae) |
| 아과 | 헤테로돈토사우루스아과 (Heterodontosaurinae) |
| 목 | †조반목 (Ornithischia) |
| 시대 | |
| 생존 시기 | 쥐라기 전기, 헤탕기절 |
| 발견 지층 | 상부 엘리엇층 |
2. 발견 및 명명

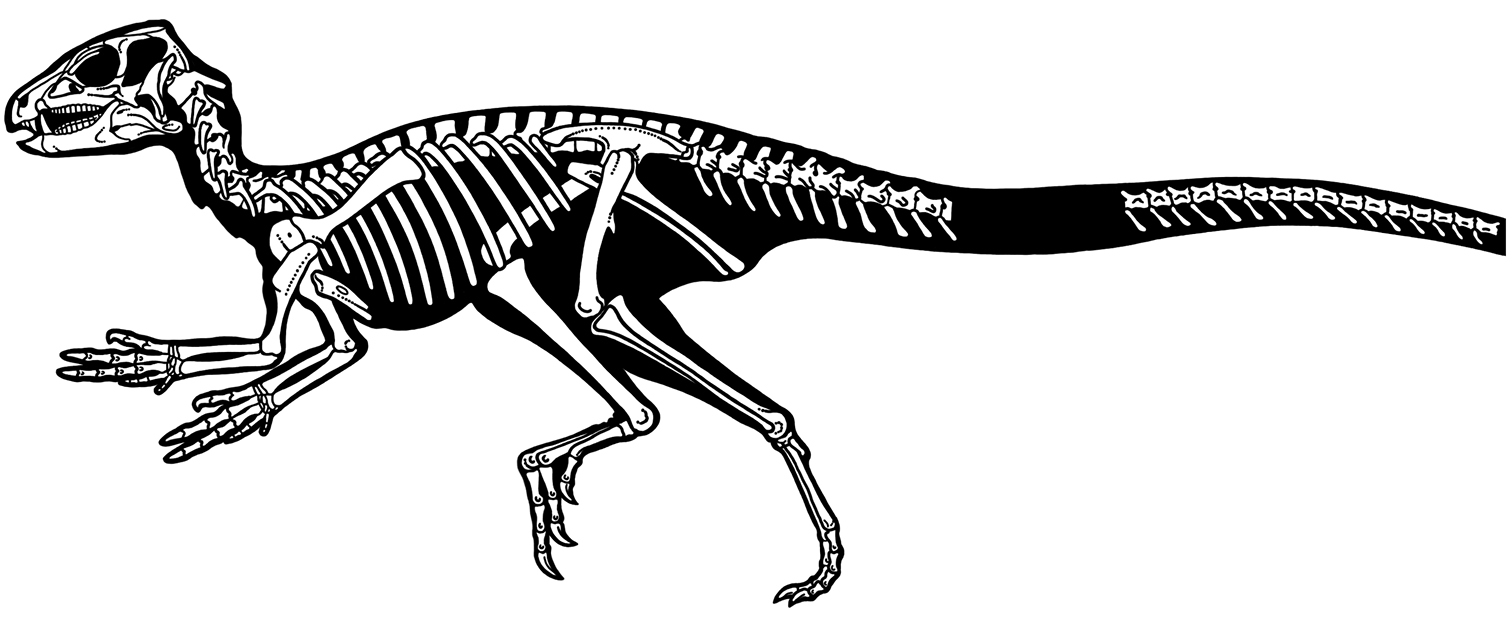
헤테로돈토사우루스는 1961~1962년 영국-남아프리카 원정대가 남아프리카 공화국과 바수톨란드(구 레소토의 옛 이름)에서 처음 발견했다. 모식 표본(SAM-PK-K337)은 이치코 남아프리카 박물관에 보관되어 있으며, 트란스케이(허셜이라고도 불림) 지구의 타이닌디니라는 지역에서 발견되었다. 이 표본은 으스러졌지만 거의 완전한 두개골 형태였다.[11][1] 1962년 고생물학자 알프레드 월터 크롬턴과 앨런 J. 채리그가 이 화석을 과학적으로 기술하고 명명했다. 속명은 서로 다른 모양의 이빨을 가리키며, 종명은 원정을 지원한 오스틴 모터 컴퍼니의 이사 조지 C. 턱을 기리는 것이다.[11][1]
1966년에는 스톰버그 군 암석 지층의 엘리엇 형성에 있는 보이자네 지역의 크롬메스프루이트 산에서 두 번째 표본(SAM-PK-K1332)이 발견되었다. 이 표본은 두개골과 골격이 함께 발견되어 보존 상태가 좋았다. SAM-PK-K1332는 현재까지 설명된 가장 완전한 헤테로돈토사우루스 골격이다.
이후 보이자네 지역에서 어린 개체의 두개골 앞부분(SAM-PK-K10487), 조각난 상악골(SAM-PK-K1326), 이빨과 인접한 뼈가 있는 왼쪽 상악골(SAM-PK-K1334) 등이 추가로 발견되었다.[1] 1975년에는 투쉴라와 농장에서 부분적인 주둥이(NM QR 1788)가 발견되었는데, 처음에는 마소스폰딜루스로 분류되었다가 헤테로돈토사우루스로 재분류되었다. 2005년에는 이스턴케이프 주 그레이엄스타운 근처에서 새로운 표본(AM 4766)이 발견되었으며, 이 표본은 유럽 싱크로트론 방사선 시설에서 스캔되어 연구에 활용되었다.[2][3][4]
한편, 1970년 고생물학자 리처드 A. 툴본은 ''헤테로돈토사우루스''가 리코리누스의 주니어 동의어라고 제안했지만, 1974년 채리그와 크롬턴은 두 속이 같은 과에 속하지만, 동종은 아니라고 반박했다. 1975년 제임스 A. 홉슨은 ''헤테로돈토사우루스''를 별개의 속으로 유지해야 한다고 주장했다.
2. 1. 초기 발견
''헤테로돈토사우루스 터키''(SAM-PK-K337)의 모식 표본은 1961~1962년 영국-남아프리카 원정대가 남아프리카 공화국과 바수톨란드(구 레소토의 옛 이름)에서 발견했다. 현재 이 표본은 이치코 남아프리카 박물관에 보관되어 있다. 이 표본은 트란스케이(때로는 허셜이라고도 불림) 지구의 케이프 주에 있는 타이닌디니라는 지역에서 발견되었다. 표본은 으스러졌지만 거의 완전한 두개골로 구성되어 있으며, 원래 설명에 언급된 연관된 두개골 이후 골격은 2011년에 발견되지 않았다.[11][1] 이 동물은 1962년 고생물학자 알프레드 월터 크롬턴과 앨런 J. 채리그에 의해 과학적으로 기술되고 명명되었다. 속명은 서로 다른 모양의 이빨을 가리키며, 종명은 원정을 지원한 오스틴 모터 컴퍼니의 이사 조지 C. 턱을 기리는 것이다.이 표본은 출판 당시 완전히 준비되지 않아 두개골과 아래턱의 앞부분만 묘사되었고, 저자들은 그들의 설명이 주로 동물을 명명하기 위한 예비적인 것이라고 인정했다. 당시 초기의 조반류 공룡이 거의 알려지지 않았기 때문에 중요한 발견으로 여겨졌다. 표본의 준비, 즉 뼈를 암석 매트릭스에서 분리하는 작업은 매우 시간이 많이 걸렸는데, 뼈가 적철광을 함유한 얇고 매우 단단한 철분이 많은 층으로 덮여 있었기 때문이다. 이것은 다이아몬드 톱으로만 제거할 수 있었고, 이는 표본을 손상시켰다.[11][1]
1966년, 두 번째 ''헤테로돈토사우루스'' 표본(SAM-PK-K1332)이 스톰버그 군 암석 지층의 엘리엇 형성에 있는 보이자네 지역의 크롬메스프루이트 산에서 발견되었다. 이 표본은 두개골과 골격을 포함하고 있었고, 관절 상태로 보존되었으며(즉, 뼈가 서로 자연적인 위치에 보존되어 있음), 뼈의 변위와 왜곡이 거의 없었다. 두개골 이후 골격은 1976년 고생물학자 앨버트 산타 루카, 크롬턴, 채리그에 의해 간략하게 설명되었다. 그 앞다리 뼈는 1974년 고생물학자 피터 갤턴과 로버트 T. 배커의 논문에서 이전에 논의되고 그림으로 묘사되었는데, 그 표본이 공룡이 단계통군 자연 집단임을 확립하는 데 중요하다고 여겨졌기 때문이다. 반면, ''헤테로돈토사우루스''를 설명한 과학자를 포함한 당시 대부분의 과학자들은 사우로포다와 조반류라는 두 주요 목이 직접적으로 관련이 없다고 생각했다. 이 골격은 1980년에 완전히 설명되었다. SAM-PK-K1332는 현재까지 설명된 가장 완전한 헤테로돈토사우루스 골격이다. ''헤테로돈토사우루스'' 두개골에 대한 보다 자세한 설명이 오랫동안 약속되었지만, 1997년 채리그가 사망할 때까지 출판되지 않았다. 2011년이 되어서야 고생물학자 데이비드 B. 노먼과 동료들이 두개골을 완전히 설명했다.
''헤테로돈토사우루스''에 속하는 것으로 언급된 다른 표본으로는 어린 개체의 두개골 앞부분(SAM-PK-K10487), 조각난 상악골(SAM-PK-K1326), 이빨과 인접한 뼈가 있는 왼쪽 상악골(SAM-PK-K1334) 등이 있으며, 모두 1966~1967년 원정 기간 동안 보이자네 지역에서 수집되었지만, 처음 것은 2008년에야 이 속에 속하는 것으로 확인되었다. 보이자네 남쪽의 투쉴라와 농장에서 1975년에 발견된 부분적인 주둥이(NM QR 1788)는 2011년까지 ''마소스폰딜루스''에 속하는 것으로 생각되었지만, ''헤테로돈토사우루스''로 재분류되었다. 고생물학자 로버트 브룸은 부분적인 두개골을 발견했는데, 아마도 남아프리카 공화국의 클라렌스 형성에서 발견되었으며, 거의 전체가 시냅스류 화석으로 구성된 컬렉션의 일부로 1913년 미국 자연사 박물관에 판매되었다. 이 표본(AMNH 24000)은 세레노에 의해 처음으로 아성체 ''헤테로돈토사우루스''에 속하는 것으로 확인되었으며, 그는 헤테로돈토사우루스과에 관한 2012년 단행본에서 보고했다.[1] 이 검토에서는 보이자네에서 발견된 부분적인 두개골 이후 골격(SAM-PK-K1328)을 ''헤테로돈토사우루스''로 분류했다. 그러나 2014년, 갤턴은 대신 같은 지역에서 발견된 부분적인 두개골을 기반으로 세레노가 명명한 관련 속 ''페고마스탁스''에 속할 수 있다고 제안했다.
2005년, 새로운 ''헤테로돈토사우루스'' 표본(AM 4766)이 이스턴케이프 주 그레이엄스타운 근처의 하천 바닥에서 발견되었다. 표본은 매우 완전했지만, 그 주변의 암석이 완전히 제거하기에는 너무 단단했다. 따라서 이 표본은 2016년 유럽 싱크로트론 방사선 시설에서 스캔되어 골격을 드러내고, 해부학 및 생활 방식에 대한 연구를 돕기 위해 스캔되었으며, 그 중 일부는 2021년에 출판되었다.[2][3][4]
1970년, 고생물학자 리처드 A. 툴본은 ''헤테로돈토사우루스''가 1924년에 남아프리카 공화국에서 발견된 표본에서 명명된 종 ''L. angustidens''을 가진 속 ''리코리누스''의 주니어 동의어라고 제안했다. 그는 이빨의 약간의 차이와 지층학적 특징으로 인해 구별된다고 여겨지는 새로운 조합 ''리코리누스 터키''로 모식 종을 더 오래된 속의 구성원으로 재분류했다. 그는 1974년, 갤턴이 동의어에 대해 비판한 후 세 번째 ''리코리누스'' 종인 ''리코리누스 콘소르스''의 설명에서 이 주장을 반복했다. 1974년, 채리그와 크롬턴은 ''헤테로돈토사우루스''와 ''리코리누스''가 같은 과, 즉 헤테로돈토사우루스과에 속한다는 데 동의했지만, 동종으로 간주될 만큼 충분히 유사하지 않다는 데 동의하지 않았다. 그들은 또한 ''리코리누스 안구스티덴스'' 모식 표본의 조각난 특성과 열악한 보존 상태로 인해 이를 ''H. 터키''와 제대로 비교하는 것이 불가능하다고 지적했다. 논란에도 불구하고, 양측은 ''L. 안구스티덴스'' 모식 표본을 직접 검사하지 않았지만, 고생물학자 제임스 A. 홉슨은 1975년에 ''헤테로돈토사우루스''의 속별 분리를 옹호했고, ''L. 콘소르스''를 자신의 속인 ''아브릭토사우루스''로 옮겼다.
2. 2. 추가 발견
''헤테로돈토사우루스 터키''(SAM-PK-K337)의 모식 표본은 1961~1962년 영국-남아프리카 원정대가 남아프리카 공화국과 바수톨란드(구 레소토의 옛 이름)에서 발견했다. 현재 이 표본은 이치코 남아프리카 박물관에 보관되어 있다. 이 표본은 트란스케이(때로는 허셜이라고도 불림) 지구의 케이프 주에 있는 타이닌디니라는 지역에서 해발 1890m 높이의 산에서 발굴되었다. 표본은 으스러졌지만 거의 완전한 두개골로 구성되어 있으며, 원래 설명에 언급된 연관된 두개골 이후 골격은 2011년에 발견되지 않았다. 이 동물은 1962년 고생물학자 알프레드 월터 크롬턴과 앨런 J. 채리그에 의해 과학적으로 기술되고 명명되었다. 속명은 서로 다른 모양의 이빨을 가리키며, 종명은 원정을 지원한 오스틴 모터 컴퍼니의 이사 조지 C. 턱을 기리는 것이다.[11][1]
1966년, 두 번째 ''헤테로돈토사우루스'' 표본(SAM-PK-K1332)이 스톰버그 군 암석 지층의 엘리엇 형성에 있는 보이자네 지역 해발 1770m 높이의 크롬메스프루이트 산에서 발견되었다. 이 표본은 두개골과 골격을 포함하고 있었고, 관절 상태로 보존되었으며(즉, 뼈가 서로 자연적인 위치에 보존되어 있음), 뼈의 변위와 왜곡이 거의 없었다. 두개골 이후 골격은 1976년 고생물학자 앨버트 산타 루카, 크롬턴, 채리그에 의해 간략하게 설명되었다. SAM-PK-K1332는 현재까지 설명된 가장 완전한 헤테로돈토사우루스 골격이다.
''헤테로돈토사우루스''에 속하는 것으로 언급된 다른 표본으로는 어린 개체의 두개골 앞부분(SAM-PK-K10487), 조각난 상악골(SAM-PK-K1326), 이빨과 인접한 뼈가 있는 왼쪽 상악골(SAM-PK-K1334) 등이 있으며, 모두 1966~1967년 원정 기간 동안 보이자네 지역에서 수집되었지만, 처음 것은 2008년에야 이 속에 속하는 것으로 확인되었다. 보이자네 남쪽의 투쉴라와 농장에서 1975년에 발견된 부분적인 주둥이(NM QR 1788)는 2011년까지 ''마소스폰딜루스''에 속하는 것으로 생각되었지만, ''헤테로돈토사우루스''로 재분류되었다.[1]
2005년, 새로운 ''헤테로돈토사우루스'' 표본(AM 4766)이 이스턴케이프 주 그레이엄스타운 근처의 하천 바닥에서 발견되었다. 표본은 매우 완전했지만, 그 주변의 암석이 완전히 제거하기에는 너무 단단했다. 따라서 이 표본은 2016년 유럽 싱크로트론 방사선 시설에서 스캔되어 골격을 드러내고, 해부학 및 생활 방식에 대한 연구를 돕기 위해 스캔되었으며, 그 중 일부는 2021년에 출판되었다.[2][3][4]
1970년, 고생물학자 리처드 A. 툴본은 ''헤테로돈토사우루스''가 1924년에 남아프리카 공화국에서 발견된 표본에서 명명된 종 ''L. angustidens''을 가진 속 ''리코리누스''의 주니어 동의어라고 제안했다. 그는 이빨의 약간의 차이와 지층학적 특징으로 인해 구별된다고 여겨지는 새로운 조합 ''리코리누스 터키''로 모식 종을 더 오래된 속의 구성원으로 재분류했다. 1974년, 채리그와 크롬턴은 ''헤테로돈토사우루스''와 ''리코리누스''가 같은 과, 즉 헤테로돈토사우루스과에 속한다는 데 동의했지만, 동종으로 간주될 만큼 충분히 유사하지 않다는 데 동의하지 않았다. 그들은 또한 ''리코리누스 안구스티덴스'' 모식 표본의 조각난 특성과 열악한 보존 상태로 인해 이를 ''H. 터키''와 제대로 비교하는 것이 불가능하다고 지적했다. 논란에도 불구하고, 양측은 ''L. 안구스티덴스'' 모식 표본을 직접 검사하지 않았지만, 고생물학자 제임스 A. 홉슨은 1975년에 ''헤테로돈토사우루스''의 속별 분리를 옹호했고, ''L. 콘소르스''를 자신의 속인 ''아브릭토사우루스''로 옮겼다.
2. 3. 학명의 유래
헤테로돈토사우루스라는 속명(屬名)은 '서로 다른 모양의 이빨'을 뜻하며, 종명(種名)은 이 화석을 발견하기 위한 원정을 지원한 오스틴 모터 컴퍼니의 이사 조지 C. 턱을 기리기 위해 붙여졌다.[11][1]3. 특징
헤테로돈토사우루스는 이형치(Heterodonty)를 가진 공룡으로, 이는 속명의 유래이기도 하다. 대부분의 공룡이 턱에 한 가지 형태의 이빨을 가진 반면, 헤테로돈토사우루스는 세 가지 종류의 이빨을 가지고 있었다.[22] 턱 앞쪽에는 식물을 자르는 데 사용되는 작은 이빨이 있었고,[23] 그 뒤에는 한 쌍의 큰 송곳니가 있었다. 이 송곳니의 역할은 불분명하지만, 성적 과시나 고대 개미집을 파괴하는 데 사용되었을 가능성이 제기된다. 마지막으로 뺨 쪽에는 높고 사각형 모양의 이빨이 있어 음식을 씹는 데 적합했으며, 육질의 뺨은 음식이 입 안에 머물도록 도왔다.[23] 이러한 씹는 행위는 공룡에게는 일반적이지만, 다른 파충류 그룹에서는 흔하지 않다.
2009년에는 목에서 꼬리까지 수백 개의 길고 실 모양의 외피(때로는 강모와 비교됨)를 가진 관련 종인 ''티아니유롱''(Tianyulong)이 보고되면서, ''헤테로돈토사우루스''도 이러한 구조를 가졌을 것으로 추정되었다.[7] 폴 세레노는 헤테로돈토사우루스가 살아있을 때 "민첩한 두 다리의 호저"처럼 보였을 것이라고 언급했다.[7]
목은 9개의 경추로 구성되어 S자형 곡선을 이루었고, 몸통은 짧고 12개의 등뼈와 6개의 융합된 천추로 구성되었다. 꼬리는 몸에 비해 길고 34~37개의 미추로 구성되었을 것으로 추정된다. 등뼈는 골화된 힘줄로 인해 뻣뻣했지만, 꼬리에는 이러한 힘줄이 없어 유연했다.
견갑골 위에는 파크소사우루스를 제외한 다른 공룡에서는 발견되지 않는 상견갑골이 있었다. 잘 발달된 한 쌍의 흉골은 흉골 늑골을 통해 늑골과 연결되었으며, 이는 다른 조반목 공룡과 달리 움직일 수 있어 호흡 시 몸통이 팽창할 수 있게 했다. 헤테로돈토사우루스는 복늑골을 가진 유일한 조반목 공룡이다.[4]
앞다리는 튼튼하고 길었으며, 뒷다리 길이의 70%에 달했다.[11] 손은 크고 5개의 손가락을 가졌으며, 그중 처음 세 손가락은 크고 강한 발톱으로 끝났다. 네 번째와 다섯 번째 손가락은 흔적기관으로 추정된다. 뒷다리는 길고 가늘었으며, 네 개의 발가락 중 첫 번째 발가락(무지)은 땅에 닿지 않았다. 경골과 비골은 거골, 종골과 융합되어 경비골을 형성했고, 하부 족근골은 중족골과 융합되어 족근중족골을 형성했다. 이는 현대 조류와 유사한 구성으로, 수렴 진화의 결과이다.[11]
3. 1. 크기 및 체형

''헤테로돈토사우루스''는 작은 공룡이었다. 가장 완벽한 골격인 SAM-PK-K1332는 길이가 약 1.18m인 동물에 속했다. 무게는 별도의 연구에서 1.8kg, 2.59kg, 3.4kg으로 다양하게 추정되었다. 골격의 척추 봉합이 닫힌 것은 표본이 성체였고 아마도 완전히 성장했음을 나타낸다. 불완전한 두개골로 구성된 두 번째 표본은 ''헤테로돈토사우루스''가 최대 1.75m의 길이와 거의 10kg의 체질량으로 상당히 더 커졌을 수 있음을 나타낸다. 두 표본 간의 크기 차이의 이유는 불분명하며 단일 종 내의 변동성, 성적 이형성, 또는 두 개의 별도 종의 존재를 반영할 수 있다. 이 공룡의 크기는 칠면조와 비교되었다.[5] ''헤테로돈토사우루스''는 헤테로돈토사우루스과(Heterodontosauridae)에서 가장 큰 구성원 중 하나였다.[6]

헤테로돈토사우루스는 소형 경량으로, 몸길이는 최대 90cm 정도의 조반류이며, 더 파생적인 조반류에서 보이는 것보다 좁은 골반과 치골을 가지고 있었다.[23] 앞발에는 5개의 발가락을 가지고 있으며, 그 중 2개는 명백히 서로 마주보고 있어 먹이를 쥐고 조작하는 것이 가능했다. 발과 발목의 뼈는 새와 같은 방식으로 유합되어 있었다.[23] 몇몇 특징으로 미루어 보아, 그들이 혹독한 계절에 하계 휴면이나 동면을 했을 가능성이 지적되던 시기도 있었다.[21]
3. 2. 두개골 및 치아
헤테로돈토사우루스의 두개골에서 상대적으로 큰 아래쪽 측두개창은 달걀 모양이고 뒤로 기울어져 있으며, 눈 구멍 뒤에 위치해 있었다. 타원형의 위쪽 측두개창은 두개골 상단을 볼 때만 보였다. 좌우 위쪽 측두개창은 시상 능선에 의해 분리되었으며, 이는 살아있는 동물에서 턱 근육의 측면 부착 표면을 제공했을 것이다.입천장은 좁았고 앞쪽으로 갈수록 좁아졌다. 외부 콧구멍은 작았고, 이 구멍의 위쪽 경계는 뼈에 의해 완전히 연결되지 않은 것으로 보인다. 파손되지 않은 경우 틈새는 뼈 대신 결합 조직에 의해 형성되었을 수 있다. 눈과 콧구멍 사이의 큰 함몰부인 전안와와는 두 개의 작은 구멍을 포함하고 있었다. 주둥이 위쪽의 함몰부를 "비강 와" 또는 "고랑"이라고 한다. 유사한 와는 ''티아니울롱'', ''아길리사우루스'', ''에오랩터''에서도 발견되지만 그 기능은 알려져 있지 않다.[8]
3. 2. 1. 이형치(異形齒)의 구조
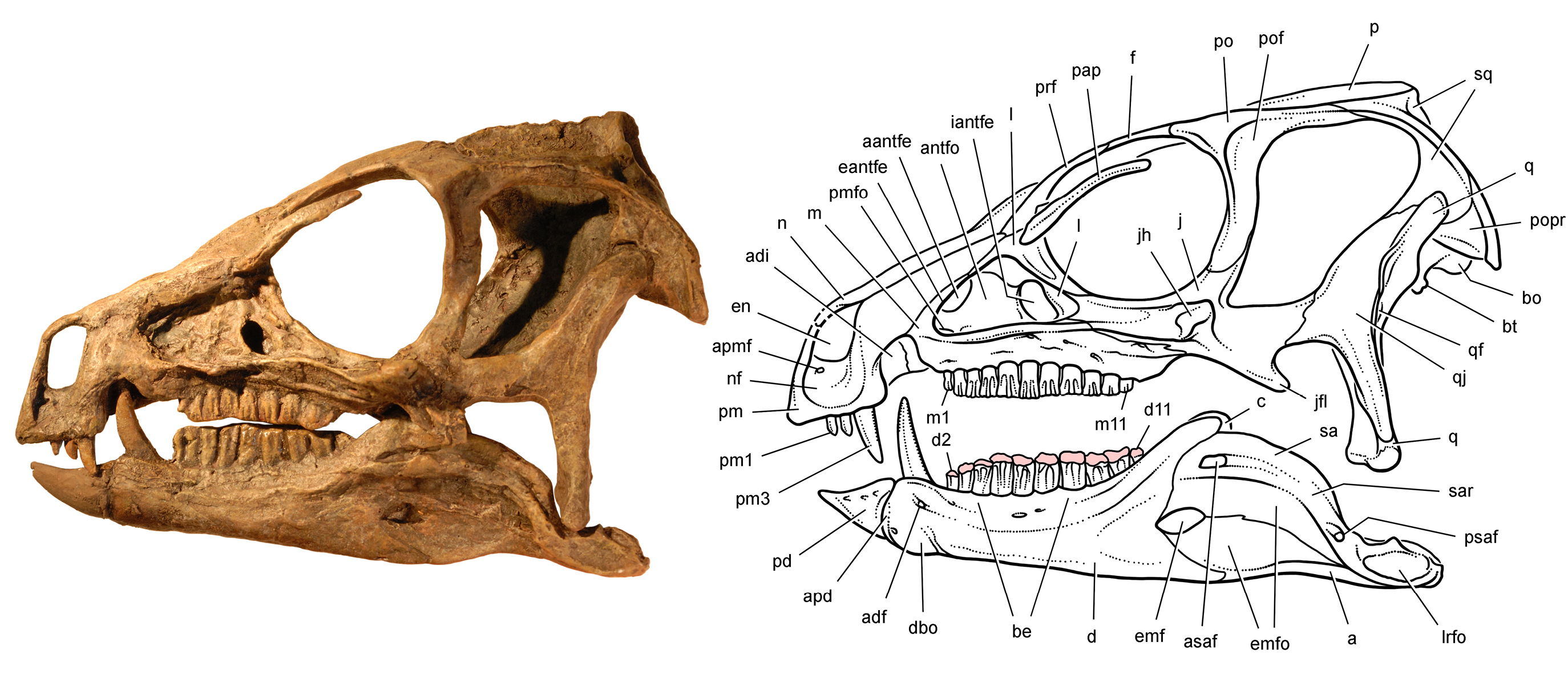
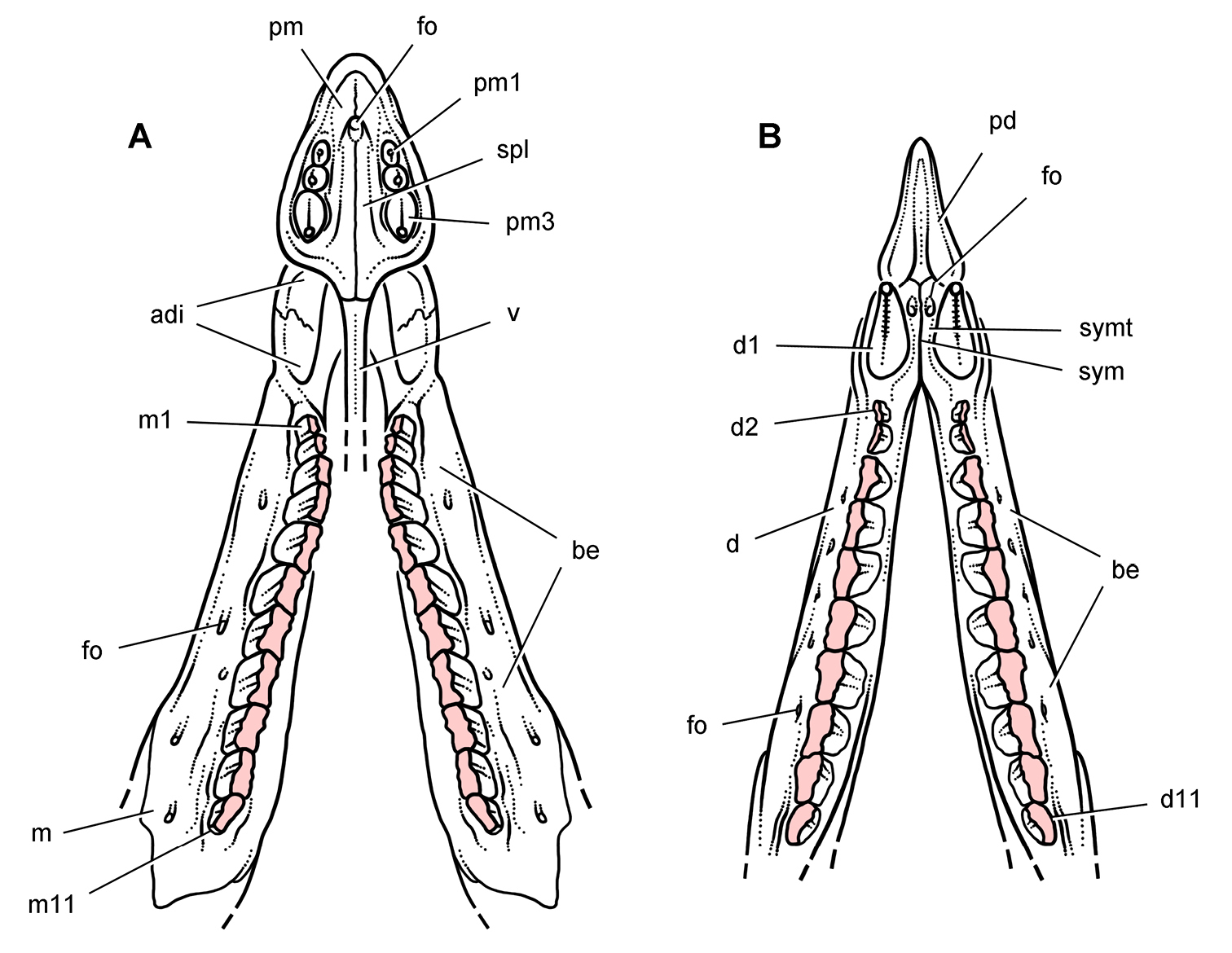
''헤테로돈토사우루스''의 두개골은 작지만 튼튼하게 만들어졌다. 가장 완전한 두개골 2개의 길이는 108mm(모식표본 SAM-PK-K337)와 121mm (표본 SAM-PK-K1332)로 측정되었다. 두개골은 옆에서 볼 때 길고 좁으며 삼각형 모양이며, 가장 높은 지점은 시상 능선으로, 이곳에서 두개골은 주둥이 끝을 향해 아래로 기울어져 있었다. 두개골 뒤쪽은 갈고리 모양으로 끝나 사두골에 닿았다. 안와 (눈 구멍)는 크고 둥글었으며, 큰 가시 모양의 뼈인 안검골이 구멍의 위쪽 부분으로 뒤쪽으로 돌출되었다. 눈구멍 아래에서 관골은 옆으로 돌출된 혹 또는 뿔 모양 구조를 형성했다. 관골은 또한 아래턱의 움직임을 안내하기 위해 익상골의 플랜지와 함께 슬롯을 만드는 "날"을 형성했다. 복부 쪽으로, 전안와는 동물의 살집 있는 뺨이 부착되었을 뚜렷한 뼈 능선에 의해 경계가 지어졌다. 또한 헤테로돈토사우루스와 다른 기저 (또는 "원시") 조반류는 도마뱀처럼 위턱과 아래턱 사이의 피부를 연결하는 대신 턱의 유사성을 바탕으로 입술과 같은 구조를 가졌을 가능성이 제기되었다.[9]
아래턱은 앞쪽으로 갈수록 좁아졌으며, 치골 (아래턱의 주요 부분)은 튼튼했다. 턱의 앞쪽은 이가 없는 케라틴 부리 (또는 주둥이)로 덮여 있었다. 위쪽 부리는 전상악골의 앞부분을 덮고, 아래쪽 부리는 전치골을 덮고 있는데, 이는 각각 조반류의 위턱과 아래턱에서 가장 앞쪽에 있는 뼈이다. 이는 이러한 구조의 거친 표면으로 입증된다.
두개골의 특이한 특징은 이 속의 이름이 붙여진 서로 다른 모양의 이빨 (이형치아)이었으며, 이는 포유류에서 주로 알려져 있다. 대부분의 공룡 (그리고 실제로 대부분의 파충류)은 턱에 단일 유형의 이빨을 가지고 있지만, ''헤테로돈토사우루스''는 세 가지 종류를 가지고 있었다. 주둥이의 부리 끝은 이가 없었지만, 위턱의 전상악골 뒤쪽 부분에는 각 면에 세 개의 이빨이 있었다. 처음 두 개의 위쪽 이빨은 작고 원뿔 모양(절치와 유사)이었으며, 각 면의 세 번째 이빨은 훨씬 커져서 눈에 띄는 송곳니와 같은 엄니를 형성했다. 이 처음 두 개의 이빨은 아마도 위쪽 부리에 의해 부분적으로 감싸여 있었을 것이다. 아래턱의 처음 두 개의 이빨도 송곳니를 형성했지만, 위쪽의 것보다 훨씬 컸다.
송곳니는 뒷면에 미세한 톱니가 있었지만, 아래쪽 송곳니만 앞쪽에 톱니가 있었다. 열한 개의 높고 끌 모양의 협측 치아가 위턱의 뒤쪽 부분의 각 측면을 따라 정렬되어 있었고, 이는 큰 이개 (틈새)로 송곳니와 분리되어 있었다. 협측 치아는 크기가 점차 증가하여 가운데 치아가 가장 컸으며, 이 지점 이후에는 크기가 감소했다. 이 이빨은 안쪽 면에 두꺼운 법랑질 층을 가지고 있었고, 마모에 적응되었으며 (고치성), 긴 뿌리를 가지고 있어 소켓에 단단히 박혀 있었다. 아래턱의 송곳니는 위턱의 이개 내의 함몰부에 맞았다. 아래턱의 협측 치아는 일반적으로 위턱의 협측 치아와 일치했지만, 이의 법랑질 표면은 바깥쪽에 있었다. 위쪽과 아래쪽 치아 열은 안쪽으로 들어가 다른 조반류에서도 볼 수 있는 "뺨 뒤쪽"을 만들었다. 이빨의 종류가 다름에도 불구하고, 그들의 조직학과 법랑질 미세 구조는 복잡하지 않았다. 그러나 법랑질이 이빨의 바깥쪽 표면을 향해 얇아지는 동안, 얇아지는 법랑질과 동시에 두꺼운 마모 방지 상아질 띠가 생겨 교합면의 절단 능선을 형성했으며, 이는 일반적으로 법랑질이 채우는 역할이었다.[10]
이빨의 특수화는 이 동물의 이름의 유래가 되기도 한다. 턱의 앞쪽에는 부리에 따라 잎이나 줄기를 자르는 작은 이빨, 턱 부분에는 큰 한 쌍의 송곳니가 이어지는데, 이 역할은 불분명하지만 성적 과시에 사용했을 가능성이 있거나, 고대의 개미집을 파괴하는 데 사용했을 것이라고 추측된다. 마지막 유형의 이빨은 높고 사각형이며 씹기에 적합하다. 씹는 동안 육질의 뺨이 음식을 입 안에 유지하는 것을 도왔다.[23] 씹는 행위는 공룡에게는 일반적이지만, 다른 파충류 그룹에서는 일반적이지 않다.
이 기묘한 이빨은 헤테로돈토사우루스의 식성에 대한 논쟁을 불러일으켰다. 헤테로돈토사우루스는 잡식성이며, 식물과 작은 동물을 모두 먹기 위해 모양이 다른 이빨을 사용했다고 생각하는 과학자도 있다.[24]
3. 3. 골격
헤테로돈토사우루스는 작은 공룡이었다. 가장 완벽한 골격(SAM-PK-K1332)은 길이가 약 1.18m이고, 무게는 1.8kg, 2.59kg, 3.4kg 등으로 다양하게 추정되었다. 골격의 척추 봉합이 닫혀 있는 것은 이 표본이 성체이며 완전히 성장했음을 나타낸다. 불완전한 두개골로 구성된 두 번째 표본은 헤테로돈토사우루스가 최대 1.75m의 길이와 거의 10kg의 체질량을 가질 수 있음을 보여준다. 두 표본 간 크기 차이의 원인은 불분명하며, 단일 종 내 변동성, 성적 이형성, 또는 두 개의 별도 종 존재를 반영할 수 있다. 이 공룡의 크기는 칠면조와 비교되기도 한다.[5]
목은 9개의 경추로 구성되었고, 척추 몸통 모양으로 보아 S자형 곡선을 이루었을 것이다. 몸통은 짧았고, 12개의 등뼈와 6개의 융합된 천추로 구성되었다. 꼬리는 몸에 비해 길었고, 불완전하게 알려져 있지만 아마 34~37개의 미추로 구성되었을 것이다. 등뼈는 네 번째 등뼈부터 골화된 힘줄에 의해 뻣뻣해졌다. 이는 다른 많은 조반목 공룡에서도 나타나는 특징으로, 이족 보행 시 척추에 가해지는 굴곡력에 의한 스트레스를 상쇄했을 것으로 보인다. 다른 많은 조반목 공룡과 달리, 헤테로돈토사우루스의 꼬리는 골화된 힘줄이 없어 유연했을 것이다.
견갑골은 파크소사우루스를 제외한 공룡에서는 알려지지 않은 상견갑골로 덮여 있었다. 가슴 부위에서 헤테로돈토사우루스는 수각류와 유사하지만, 다른 조반목 공룡의 흉골판과는 다른 잘 발달된 한 쌍의 흉골을 가지고 있었다. 흉골판은 흉골 늑골에 의해 늑골에 연결되었다. 다른 조반목 공룡과 달리, 이 연결은 움직일 수 있어 호흡 시 몸이 팽창할 수 있었다. 헤테로돈토사우루스는 복늑골(흉골판과 골반의 치골 사이 뼈)을 가진 유일한 조반목 공룡이다. 복늑골은 두 개의 길이 방향 열로 배열되었으며, 각각 약 9개의 요소를 포함하고 있다.[4] 골반은 길고 좁았으며, 더 진화된 조반목 공룡이 가진 것과 유사한 치골을 가지고 있었다.

앞다리는 튼튼하게 만들어졌고,[11] 뒷다리 길이의 70%를 차지할 정도로 길었다. 팔뚝의 요골은 상완골(팔뚝 뼈) 길이의 70%였다. 손은 컸으며, 길이에서 상완골에 접근했고, 잡기에 적합한 5개의 손가락을 가지고 있었다.[11] 두 번째 손가락이 가장 길었고, 세 번째와 첫 번째 손가락(엄지손가락)이 그 뒤를 이었다. 처음 세 손가락은 크고 강한 발톱으로 끝났다. 네 번째와 다섯 번째 손가락은 강하게 감소했으며, 아마도 흔적기관이었을 것이다. 첫 번째 손가락부터 시작하여 각 손가락의 뼈 수를 나타내는 지골 공식은 2-3-4-3-2였다.
뒷다리는 길고 가늘었으며, 네 개의 발가락으로 끝났고, 그중 첫 번째 발가락(무지)은 땅에 닿지 않았다. 조반목 공룡에게는 독특하게도 다리와 발의 여러 뼈가 융합되었다. 경골과 비골은 거골과 종골과 융합되어 경비골을 형성했고, 하부 족근골은 중족골과 융합되어 족근중족골을 형성했다. 이 구성은 현대 조류에서도 발견되며, 수렴 진화의 결과이다.[11] 경비골은 대퇴골보다 약 30% 더 길었다. 발가락의 발톱뼈는 발굽 모양이 아니라 발톱 모양이었으며, 이는 더 진화된 조반목 공룡에서 나타나는 특징이다.
헤테로돈토사우루스는 최대 몸길이가 90cm 정도인 조반류 공룡이며, 더 파생적인 조반류보다 좁은 골반과 치골을 가지고 있었다.[23] 앞발에는 5개의 발가락이 있었고, 그 중 2개는 서로 마주보는 듯한 형태를 하고 있어 먹이를 쥐고 조작하는 것이 가능했다. 발과 발목 뼈는 새와 같은 방식으로 유합되어 있었다.[23]
4. 분류
배커와 갈턴은 ''헤테로돈토사우루스''가 조반목 공룡의 진화에 중요하다고 보았다. 그 이유는 헤테로돈토사우루스의 손 패턴이 원시 수각류와 공유되었기 때문에 두 그룹 모두에게 원시적이거나 기저적이었기 때문이다. 그러나 일부 과학자들은 두 그룹이 "테코돈트" 시조류 조상으로부터 독립적으로 진화했고, 그들의 유사성은 수렴 진화 때문이라고 주장하며 이의를 제기했다. 헤테로돈토사우루스과와 파브로사우루스과 사이의 관계(예: 원시적인 조반류인 후손/조상 관계) 및 원시 각룡류인 ''프시타코사우루스''와의 관계도 제시되었지만, 이러한 관계의 본질에 대해서는 논쟁이 있었다.
4. 1. 조반목 내에서의 위치
1962년에 처음 기술되었을 때, ''헤테로돈토사우루스''는 공룡강(Dinosauria)의 두 주요 목 중 하나인 조반목의 원시적인 구성원으로 분류되었다. 연구자들은 이 동물이 ''게라노사우루스''와 ''리코리누스''와 가장 유사하다고 판단했는데, 후자는 이빨 때문에 그 당시까지 수궁류의 줄기 포유류로 여겨졌다. 그들은 조각류와 유사점을 보였고, 임시적으로 이 새로운 속을 그 그룹에 배치했다. 1966년에 알프레드 로머와 오스카 쿤은 헤테로돈토사우루스과(Heterodontosauridae)를 독립적으로 명명했다.[12][13]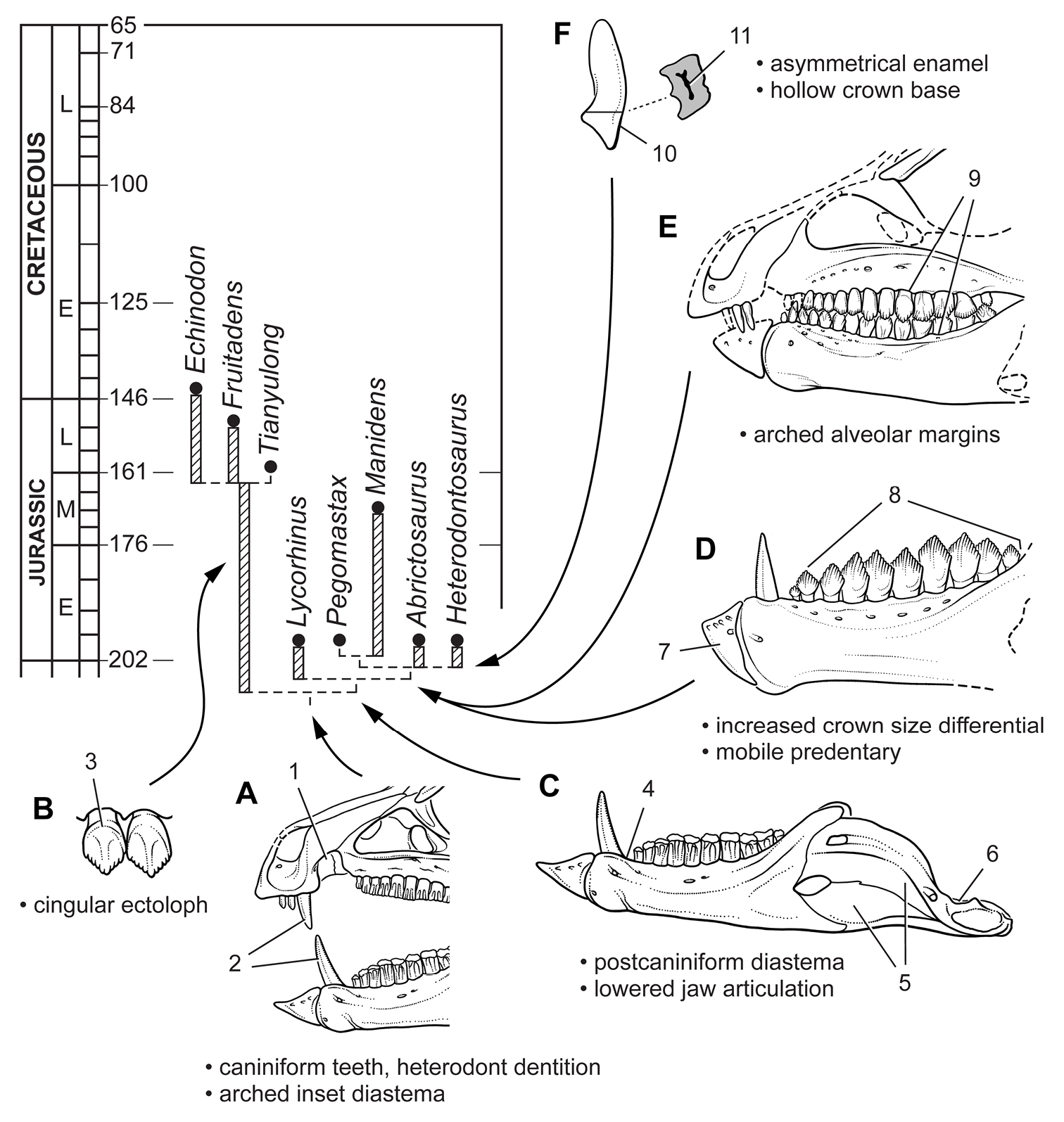
1980년대에 이르러 대부분의 연구자들은 헤테로돈토사우루스과를 조반류 공룡의 별개의 과로 간주했지만, 목 내의 다른 그룹들과의 관계는 불확실했다. 21세기 초까지, 지배적인 이론은 이 과가 경계두류 (후두류와 각룡류를 포함) 또는 케라포다 (전술한 그룹과 조각류)의 자매군이거나, 또는 겐나사우리아 (파생된 조반류를 포함)의 분할 이전의 가장 기저적인 진화적 방사 중 하나라는 것이었다. 2006년, 고생물학자 쉬싱과 동료들은 헤테로돈토사우루스과와 경계두류를 포함하는 헤테로돈토사우루스류 분기도를 명명했다.[14]
2012년 세레노의 분석에 따르면, 헤테로돈토사우루스과 내의 상호 관계는 다음과 같다:[15]
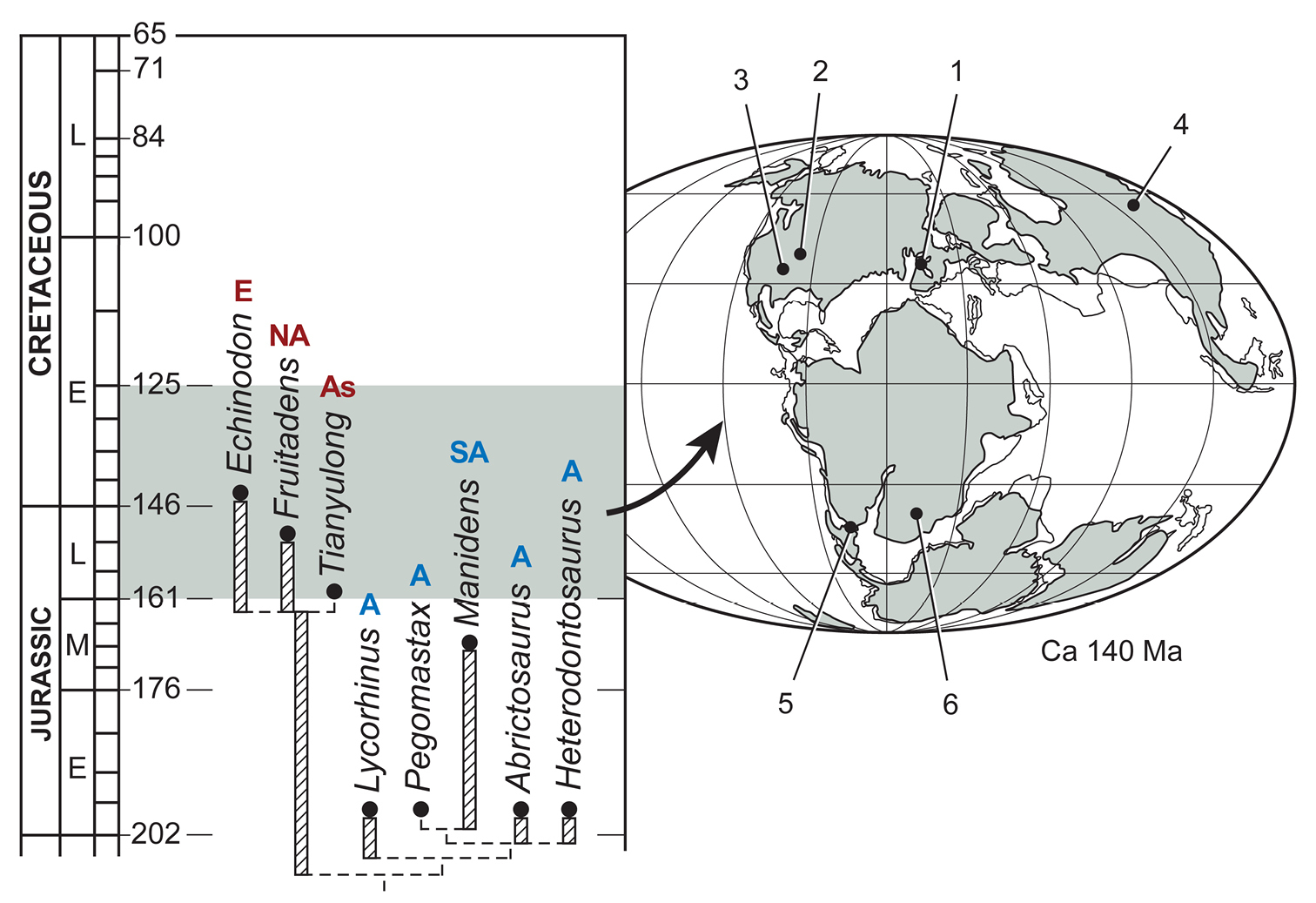
헤테로돈토사우루스과는 후기 트라이아스기부터 초기 백악기까지 지속되었으며, 적어도 1억 년 동안 존재했다. 이들은 아프리카, 유라시아, 아메리카 대륙에서 발견되었지만, 대다수는 남아프리카에서 발견되었다. 헤테로돈토사우루스과는 초기 쥐라기까지 두 개의 주요 계통으로 분화된 것으로 보이며, 하나는 낮은 치관 이빨을 가지고, 다른 하나는 높은 치관 이빨을 가지고 있다(''헤테로돈토사우루스'' 포함). 이 그룹의 구성원들은 생물지리적으로 나뉘는데, 낮은 치관 그룹은 로라시아 (북부 대륙)의 일부였던 지역에서 발견되었고, 높은 치관 그룹은 곤드와나 (남부 대륙)의 일부였던 지역에서 발견되었다.
4. 2. 헤테로돈토사우루스과
1966년, 고생물학자 알프레드 로머와 오스카 쿤은 조반류 공룡의 과로서 헤테로돈토사우루스과(Heterodontosauridae)를 독립적으로 명명했다. 여기에는 헤테로돈토사우루스와 리코리누스가 포함되었다.[12][13] 그러나 썰번은 이들을 별개의 과가 아닌 힙실로포돈과로 간주했다.21세기 초, 헤테로돈토사우루스과는 경계두류(후두류와 각룡류 포함) 또는 케라포다(전술한 그룹과 조각류)의 자매군이거나, 겐나사우리아(파생된 조반류를 포함)의 분할 이전의 가장 기저적인 진화적 방사 중 하나로 여겨졌다. 1998년과 2005년 세레노는 헤테로돈토사우루스과를 분기도로 정의했으며, 이 그룹은 각 앞턱뼈에 3개 이하의 이빨, 송곳니 모양의 이빨과 그 뒤에 이어진 이개, 눈 아래의 협골 뿔과 같은 두개골 특징을 공유한다. 2006년, 고생물학자 쉬싱과 동료들은 헤테로돈토사우루스과와 경계두류를 포함하는 헤테로돈토사우루스류 분기도를 명명했다.[14]
헤테로돈토사우루스과는 후기 트라이아스기부터 초기 백악기까지 적어도 1억 년 동안 지속되었다. 이들은 아프리카, 유라시아, 아메리카 대륙에서 발견되었지만, 대다수는 남아프리카에서 발견되었다. 헤테로돈토사우루스과는 초기 쥐라기까지 두 개의 주요 계통으로 분화된 것으로 보인다. 하나는 낮은 치관 이빨을 가지고, 다른 하나는 높은 치관 이빨을 가졌다(''헤테로돈토사우루스'' 포함). 이 그룹의 구성원들은 생물지리적으로 나뉘는데, 낮은 치관 그룹은 로라시아(북부 대륙)에서, 높은 치관 그룹은 곤드와나(남부 대륙)에서 발견되었다.[15]
2012년, 세레노는 높은 치관을 가진 그룹을 별개의 아과인 헤테로돈토사우루스아과로 명명했다. 헤테로돈토사우루스는 매우 얇은 법랑질과 비대칭적인 패턴으로 배열된 이빨과 같은 세부 사항 때문에 가장 파생된 헤테로돈토사우루스아과로 보인다. 헤테로돈토사우루스아과의 독특한 이빨과 턱 특징은 식물 재료를 효과적으로 처리하기 위한 전문화인 것으로 보이며, 그 정교함의 수준은 후기 조반류와 유사하다.[15]
아래의 분지학은 헤테로돈토사우루스과 내의 상호 관계를 보여주며, 2012년 세레노의 분석을 따른다:[15]
4. 3. 최근의 분류 논쟁
1962년에 처음 기술되었을 때, ''헤테로돈토사우루스''는 조반목의 원시적인 구성원으로 분류되었다. 연구자들은 이 동물이 ''게라노사우루스''와 ''리코리누스''와 가장 유사하다고 판단했으며, 조각류와 유사점을 보였고, 임시적으로 이 새로운 속을 그 그룹에 배치했다. 1966년에 알프레드 로머와 오스카 쿤은 ''헤테로돈토사우루스''와 ''리코리누스''를 포함하는 헤테로돈토사우루스과(Heterodontosauridae)를 독립적으로 명명했다.[12][13]1980년대에 이르러 대부분의 연구자들은 헤테로돈토사우루스과를 조반류 공룡의 별개의 과로 간주했지만, 목 내의 다른 그룹들과의 관계는 불확실했다. 21세기 초까지, 지배적인 이론은 이 과가 경계두류 (후두류와 각룡류를 포함) 또는 케라포다 (전술한 그룹과 조각류)의 자매군이거나, 또는 겐나사우리아 (파생된 조반류를 포함)의 분할 이전의 가장 기저적인 진화적 방사 중 하나라는 것이었다. 2006년, 고생물학자 쉬싱과 동료들은 헤테로돈토사우루스과와 경계두류를 포함하는 헤테로돈토사우루스류 분기도를 명명했는데, 그 이유는 헤테로돈토사우루스에서만 알려졌던 일부 특징이 기저 각룡류 속인 ''인롱''에서도 발견되었기 때문이다.[14]
헤테로돈토사우루스과는 후기 트라이아스기부터 초기 백악기까지 지속되었으며, 적어도 1억 년 동안 존재했다. 이들은 아프리카, 유라시아, 아메리카 대륙에서 발견되었지만, 대다수는 남아프리카에서 발견되었다. 헤테로돈토사우루스과는 초기 쥐라기까지 두 개의 주요 계통으로 분화된 것으로 보이며, 하나는 낮은 치관 이빨을 가지고, 다른 하나는 높은 치관 이빨을 가지고 있다(''헤테로돈토사우루스'' 포함). 이 그룹의 구성원들은 생물지리적으로 나뉘는데, 낮은 치관 그룹은 한때 로라시아 (북부 대륙)의 일부였던 지역에서 발견되었고, 높은 치관 그룹은 곤드와나 (남부 대륙)의 일부였던 지역에서 발견되었다.[15]
2017년, ''헤테로돈토사우루스''와 초기 수각류 ''에오랩터''의 골격 사이의 유사성은 고생물학자 매튜 G. 배런과 동료들이 조반류를 수각류와 함께 조반두각류라는 그룹으로 묶을 것을 제안하는 데 사용되었다.[16] 2020년, 고생물학자 폴-에밀 디외도네와 동료들은 헤테로돈토사우루스과의 구성원이 자체 자연 그룹을 형성하지 않고 대신 점진적으로 후두류로 이어지는 기저 경계두류이며, 따라서 그 그룹의 기저 구성원이라고 제안했다. 이 가설은 후두류의 유령 계통을 줄이고, 조각류의 기원을 초기 쥐라기까지 되돌릴 것이다.[17]
5. 생태
헤테로돈토사우루스는 칠면조와 비교될 정도로 작은 공룡으로, 헤테로돈토사우루스과에서 가장 큰 구성원 중 하나였다.[5][6] 가장 완벽한 골격(SAM-PK-K1332)은 길이 1.18m, 무게 1.8kg~3.4kg 정도였으나, 불완전한 두개골 표본(최대 길이 1.75m, 체질량 10kg)은 더 컸을 가능성도 시사한다. 이 과에는 프루이다덴스처럼 매우 작은(길이 65cm) 조반류 공룡도 포함된다.
2009년, 목에서 꼬리까지 강모와 비슷한 외피가 보존된 티아니유롱이 발견된 후, 헤테로돈토사우루스도 이러한 구조를 가졌을 것으로 추정되었으며, 폴 세레노는 "민첩한 두 다리의 호저"처럼 보였을 것이라고 언급했다.[7]
헤테로돈토사우루스는 짧은 몸통과 긴 꼬리를 가졌다. 목은 9개의 경추로 S자형 곡선을 이루었고, 몸통은 12개의 등뼈와 6개의 융합된 천추로 구성되었다. 꼬리는 34~37개의 미추로 추정되며, 다른 조반목 공룡과 달리 골화된 힘줄이 없어 유연했다.
견갑골 위에는 파크소사우루스를 제외한 다른 공룡에게는 알려지지 않은 상견갑골이 있었다. 가슴 부위에는 늑골과 연결된 한 쌍의 흉골이 있어 호흡 시 몸통이 팽창할 수 있었다. 복늑골을 가진 유일한 조반목 공룡으로, 복늑골은 두 개의 길이 방향 열로 배열되었다.[4] 골반은 길고 좁았으며, 더 진화된 조반목 공룡과 유사한 치골을 가졌다.
앞다리는 튼튼하고 길었으며(뒷다리 길이의 70%),[11] 손은 크고 5개의 손가락을 가져 잡기에 적합했다. 처음 세 손가락은 크고 강한 발톱으로 끝났고, 네 번째와 다섯 번째 손가락은 흔적기관이었을 것이다. 뒷다리는 길고 가늘었으며, 네 개의 발가락 중 첫 번째 발가락(무지)은 땅에 닿지 않았다. 경골과 비골은 상부 족근골과 융합되어 경비골을, 하부 족근골은 중족골과 융합되어 족근중족골을 형성했는데, 이는 현대 조류와 수렴 진화한 것이다.[11] 발톱은 발굽 모양이 아닌 발톱 모양이었다.
5. 1. 식성 논쟁
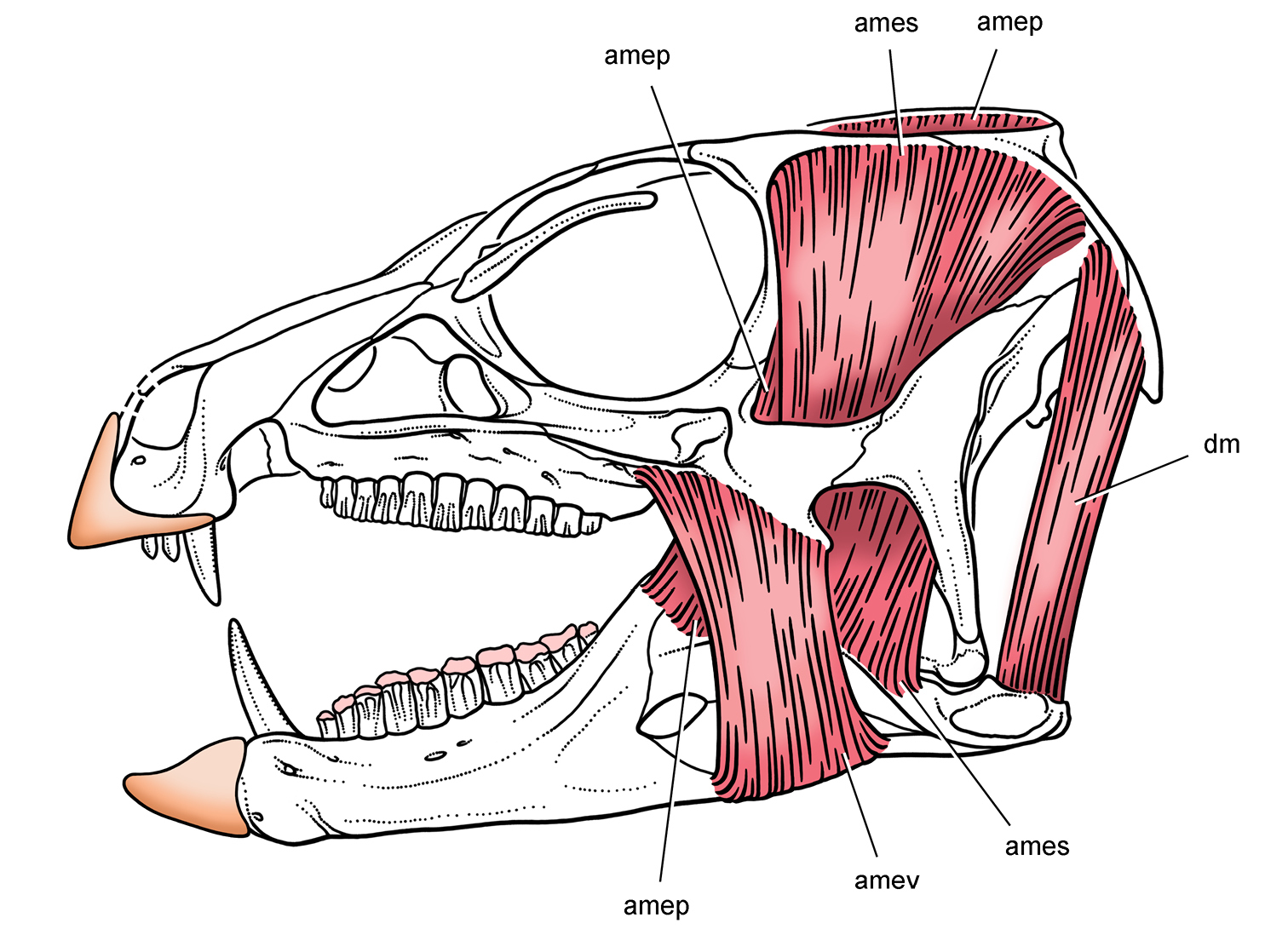
''헤테로돈토사우루스''는 일반적으로 초식 공룡으로 여겨진다.[18] 1974년 툴본은 이 공룡의 엄니가 먹이 섭취에 중요한 역할을 하지 않고, 동종과의 싸움, 과시, 시각적 위협 또는 능동적 방어에 사용되었을 것이라고 제안했다. 이는 현대의 고라니와 말사슴의 확대된 엄니와 유사하지만, 혹멧돼지의 굽은 엄니(땅 파는 데 사용)와는 다르다.
최근 연구에서는 ''헤테로돈토사우루스''가 잡식일 가능성이 있으며, 때때로 사냥감을 죽이는 데 엄니를 사용했을 수도 있다고 제기했다. 2000년 폴 배럿은 앞턱 이빨의 모양과 엄니의 미세한 톱니가 육식 동물과 유사하여 육식을 암시한다고 제안했다. 반면, 고라니는 엄니에 톱니가 없다. 2008년 버틀러와 동료들은 확대된 엄니가 개체의 발달 초기에 형성되므로 성적 이형성을 나타낼 수 없다고 주장했다. 엄니가 전투 도구라면 수컷에게만 나타나야 하므로 동종 간의 싸움은 적절한 기능이 아니다. 대신 먹이 섭취 또는 방어 기능이 더 타당하다. ''헤테로돈토사우루스''가 턱뼈로 타격을 가하고 눈꺼풀 뼈가 공격으로부터 눈을 보호했을 것이라는 제안도 있었다.[19] 2011년 노먼과 동료들은 비교적 길고 크며 굽은 발톱이 달린 팔과 손에 주목했다. 이러한 특징은 빠른 달리기를 가능하게 하는 긴 뒷다리와 결합하여 작은 먹이를 잡을 수 있게 했을 것이다. 잡식 동물로서 ''헤테로돈토사우루스''는 식물이 부족한 건기에 상당한 자연 선택적 이점을 가졌을 것이다.
2012년 세레노는 순수하거나 적어도 압도적으로 초식성 식단을 시사하는 몇 가지 두개골 및 치열 특징을 지적했다. 여기에는 뿔 부리와 특화된 어금니(식물을 잘라내기에 적합), 그리고 씹기 동안 입안에 음식을 유지하는 데 도움이 되는 살이 많은 뺨이 포함된다. 턱 근육은 확대되었고, 턱 관절은 치아 높이 아래에 위치하여 육식 공룡의 가위 모양 물기와 달리 치열을 따라 균등한 물기를 허용했을 것이다. 엄니의 크기와 위치는 헤테로돈토사우루스과 개체마다 매우 다르므로 먹이 섭취에 특정한 기능을 갖는 것은 타당하지 않다. 세레노는 헤테로돈토사우루스과가 오늘날의 페커리와 유사하며, 유사한 엄니를 가지고 뿌리, 덩이줄기, 과일, 씨앗, 풀 등 다양한 식물 재료를 먹는다고 추측했다.[18] 버틀러와 동료들은 ''헤테로돈토사우루스''의 먹이 섭취 장치가 질긴 식물 재료를 처리하도록 특화되었으며, 이 과의 후기 생존 구성원(''프루이타덴스'', ''티아니울롱'', ''에키노돈'')은 식물과 무척추동물을 모두 포함하는 더 일반화된 식단을 보였을 것이라고 제안했다. ''헤테로돈토사우루스''는 작은 입 벌림 각도에서 강한 물기로 특징지어졌지만, 후기 구성원은 더 빠른 물기와 더 넓은 입 벌림에 적응했다. 2016년 조반류 턱 역학 연구에서는 ''헤테로돈토사우루스''의 상대적 물기 힘이 더 분화된 스켈리도사우루스와 유사하다고 밝혔다. 이 연구는 엄니가 식물을 뜯어 먹는 동안 아래 부리에 대고 뜯어 먹음으로써 먹이 섭취에 역할을 했을 수 있다고 제안했다.[20]
헤테로돈토사우루스는 이빨의 특수화로 인해 그 이름이 붙여졌다. 대부분의 공룡은 턱에 늘어선 모든 이빨이 같은 형태이지만, 헤테로돈토사우루스는 3종류의 이빨을 가지고 있다.[22] 턱 앞쪽에는 부리에 따라 잎이나 줄기를 자르는 작은 이빨, 턱 부분에는 한 쌍의 큰 송곳니, 그리고 씹기에 적합한 높고 사각형의 이빨이 있다. 씹는 동안 육질의 뺨이 음식을 입 안에 유지하는 것을 도왔다.[23] 송곳니의 역할은 불분명하지만, 성적 과시에 사용되었거나 고대 개미집을 파괴하는 데 사용되었을 것으로 추측된다.
이러한 독특한 이빨은 헤테로돈토사우루스의 식성에 대한 논쟁을 불러일으켰다. 일부 과학자들은 헤테로돈토사우루스가 잡식성이며, 식물과 작은 동물을 모두 먹기 위해 모양이 다른 이빨을 사용했다고 생각한다.[24]
5. 2. 치아 교체 및 하계 휴면
'헤테로돈토사우루스'는 다른 공룡과 파충류에게 전형적인 지속적인 치아 교체를 보였는지, 그리고 어느 정도까지 보였는지에 대해 많은 논란이 있었다. 1974년과 1978년에 툴본(Thulborn)은 당시 알려진 두개골에서 지속적인 치아 교체의 징후를 발견하지 못했다. 알려진 두개골의 볼 치아는 균일하게 마모되어 동시에 형성되었음을 시사했으며, 새로 치아 맹출된 치아는 없었다. 치아의 마모면은 아래쪽 치아와 위쪽 치아의 접촉에 의해 형성되었고, 서로 융합되어 치열 전체를 따라 연속적인 표면을 형성했다. 이 표면은 음식물 처리가 '파브로사우루스'와 같은 관련 공룡에서 볼 수 있는 단순한 수직 운동이 아닌 턱의 앞뒤 움직임에 의해 이루어졌음을 나타낸다. 앞뒤 움직임은 치아가 균일하게 마모된 경우에만 가능하므로, 이는 지속적인 치아 교체가 없다는 주장을 강화했다. 툴본은 질긴 식물성 물질로 구성된 것으로 추정되는 식단은 치아의 빠른 마모를 유발했을 것이기 때문에 정기적인 치아 교체가 필수적이라고 강조했다. 이러한 관찰을 통해 툴본은 '헤테로돈토사우루스'가 정기적으로 모든 치아 세트를 한 번에 교체했음에 틀림없다고 결론 내렸다. 이러한 완전한 교체는 동물이 먹이를 먹지 않는 하계 휴면 단계에서만 가능했을 것이며, 하계 휴면은 이 동물의 서식지로 추정되는, 음식물 공급이 부족한 뜨겁고 건조한 계절을 포함하는 사막과 같은 환경과 일치한다.1980년 홉슨(Hopson)은 툴본의 생각에 의문을 제기했다. 홉슨은 치아의 마모면 패턴이 앞뒤가 아닌 수직 및 측면 턱 움직임을 나타낸다고 제시했다. 또한, 치아 마모 정도의 가변성을 보여 지속적인 치아 교체를 나타냈다. 그는 가장 완벽한 표본의 X-ray 이미지를 통해 이 개체에서 맹출되지 않은 교체 치아가 없다는 것을 인정했지만, 이는 유체만이 지속적으로 치아를 교체하며 성체가 되면 이 과정이 중단됨을 나타낸다고 보았다. 툴본의 하계 휴면 가설은 증거 부족으로 인해 홉슨에 의해 거부되었다.
2006년 버틀러(Butler)와 동료들은 유체 두개골 SAM-PK-K10487의 컴퓨터 단층 촬영 스캔을 실시했다. 놀랍게도 이 초기 발생 단계에서도 아직 맹출되지 않은 교체 치아가 존재했다. 그럼에도 불구하고 저자들은 유체가 성체와 동일한 치아 형태를 보였기 때문에 치아 교체가 발생했음에 틀림없다고 주장했다. 만약 치아가 단순히 지속적으로 자랐다면 이러한 형태가 바뀌었을 것이다. 결론적으로, 버틀러와 동료들은 '헤테로돈토사우루스'의 치아 교체가 관련 공룡보다 더 산발적으로 발생했을 것이라고 제안했다. '헤테로돈토사우루스'의 맹출되지 않은 교체 치아는 2011년 노먼(Norman)과 동료들이 표본 SAM-PK-K1334의 위턱을 묘사할 때까지 발견되지 않았다. 2012년 세레노(Sereno)가 묘사한 또 다른 유체 두개골(AMNH 24000)에서도 맹출되지 않은 교체 치아가 발견되었다. 이러한 발견을 통해 볼 때, '헤테로돈토사우루스'의 치아 교체는 다른 헤테로돈토사우루스과와 달리 간헐적이었으며 지속적이지 않았다. 맹출되지 않은 치아는 측면에서 삼각형 모양이며, 이는 기저 조반류에서 전형적인 치아 형태이다. 따라서 완전히 맹출된 치아의 특징적인 끌 모양은 위턱과 아래턱의 치아 사이의 치아 대 치아 접촉의 결과였다.[11][18]
5. 3. 이동 방식 및 신진대사
헤테로돈토사우루스는 대부분의 연구자들에게 두 발 보행 동물로 여겨지지만, 초기 연구에서는 부분적으로 또는 완전히 네 발 보행을 하는 것으로 제시되기도 했다. 1980년 산타 루카는 앞다리의 여러 특징들을 묘사했는데, 이는 네 발 동물에게서도 나타나며 강력한 팔 근육을 가졌음을 암시한다. 여기에는 팔뚝의 지렛대를 확대하는 팔꿈치 머리의 크기가 크다는 점, 팔뚝의 강력한 굴근 부착 부위를 제공하는 상완골의 내상과가 크다는 점 등이 포함된다. 산타 루카에 따르면, 헤테로돈토사우루스는 천천히 움직일 때는 네 발로 걷고, 더 빠른 속도로는 두 발로 달릴 수 있었다.테레사 마리안스카와 할슈카 오스몰스카는 1985년에 산타 루카의 가설을 지지하며, 등쪽 척추가 아래로 강하게 굽혀져 있다는 점에 주목했다. 1987년, 그레고리 S. 폴은 헤테로돈토사우루스가 의무적으로 네 발로 걸었을 것이며, 빠른 이동을 위해 갤럽했을 것이라고 제안했다. 그러나 1990년 데이비드 와이샴펠과 로렌스 위트머, 2004년 노먼 등은 발톱과 어깨 거들의 형태를 근거로 전적으로 두 발 보행을 지지했다. 산타 루카가 제시한 해부학적 증거는 먹이를 찾는 데 적응한 것으로, 튼튼하고 강한 팔은 뿌리를 파거나 곤충 둥지를 부수는 데 사용되었을 수 있다.
대부분의 연구에서는 공룡을 오늘날의 포유류와 조류와 유사한 수준의 높은 신진대사를 가진 내온성(온혈) 동물로 간주한다. 2009년 연구에서 헤르만 폰처와 동료들은 다양한 공룡의 유산소 지구력을 계산했는데, 보통의 달리기 속도에서도 헤테로돈토사우루스는 외온성(냉혈) 동물에게 가능한 최대 유산소 능력을 초과하여, 이 속의 내온성을 나타냈다.
공룡은 현대 조류처럼 기낭 시스템을 가지고 있어 움직이지 않는 폐를 환기시켰을 가능성이 높다. 공기 흐름은 가동 가능한 새 가슴 갈비뼈와 복부 갈비뼈의 존재로 흉부 수축에 의해 생성되었다. 헤테로돈토사우루스를 제외한 조반류는 가동 가능한 새 가슴 갈비뼈와 복부 갈비뼈가 없었고, 모든 조반류(헤테로돈토사우루스 포함)는 두정골 골격 공기화가 없었다. 대신 조반류는 다른 공룡에게는 없었던 치골의 전방 확장인 전방 치골 돌기(APP)를 가지고 있었다. 2021년, 잘 보존된 헤테로돈토사우루스 표본(AM 4766)의 싱크로트론 데이터를 기반으로, 빅토르 라더마허와 동료들은 조반류의 호흡 시스템이 다른 공룡과 크게 다르며, 헤테로돈토사우루스는 중간 단계를 나타낸다고 주장했다. 이들에 따르면, 조반류는 호흡을 위해 흉부를 수축하는 능력을 잃었고, 대신 폐를 직접 환기시키는 근육, 즉 ''치골 복막 근육''에 의존했다. 골반의 APP는 이 근육의 부착 부위를 제공했을 것이다. 헤테로돈토사우루스는 초기 APP를 가지고 있었고, 복부 갈비뼈는 비조반류 공룡에 비해 감소했는데, 이는 골반이 이미 호흡에 관여하는 반면 흉부 수축의 중요성은 줄어들었음을 시사한다.[4]
5. 4. 호흡 방식
헤테로돈토사우루스는 다른 조반목 공룡과 달리 가슴 부위에 흉골 늑골이라는 요소에 의해 늑골과 연결된 한 쌍의 흉골을 가지고 있었다. 이 연결은 움직일 수 있어서 호흡할 때 몸통이 팽창할 수 있었다. 또한, 복늑골(흉골판과 골반의 치골 사이 피부 속 뼈)을 가진 유일한 조반목 공룡으로 알려져 있다. 복늑골은 두 개의 길이 방향 열로 배열되었으며, 각각 약 9개의 요소를 포함하고 있었다.[4]5. 5. 성장 및 성적 이형성
헤테로돈토사우루스의 개체 발생, 즉 어린 개체에서 성체로의 발달 과정은 어린 표본이 드물어 잘 알려져 있지 않다. 어린 개체의 두개골 SAM-PK-K10487에서 볼 수 있듯이, 동물이 성장함에 따라 안와(눈구멍)가 비례적으로 작아지고 주둥이는 더 길어져 추가적인 이빨이 생겨났다. 이러한 변화는 다른 여러 공룡에서도 보고되었다. 그러나 이빨의 형태는 나이에 따라 변하지 않아 어린 개체의 식단이 성체의 식단과 동일했음을 나타낸다. 어린 개체의 두개골 길이는 45mm로 추정되었다. 성체와 유사한 신체 비율을 가정하면 이 어린 개체의 신체 길이는 450mm였을 것이다. 실제로 어린 동물은 일반적으로 머리가 비례적으로 더 크기 때문에 개체가 더 작았을 것이다.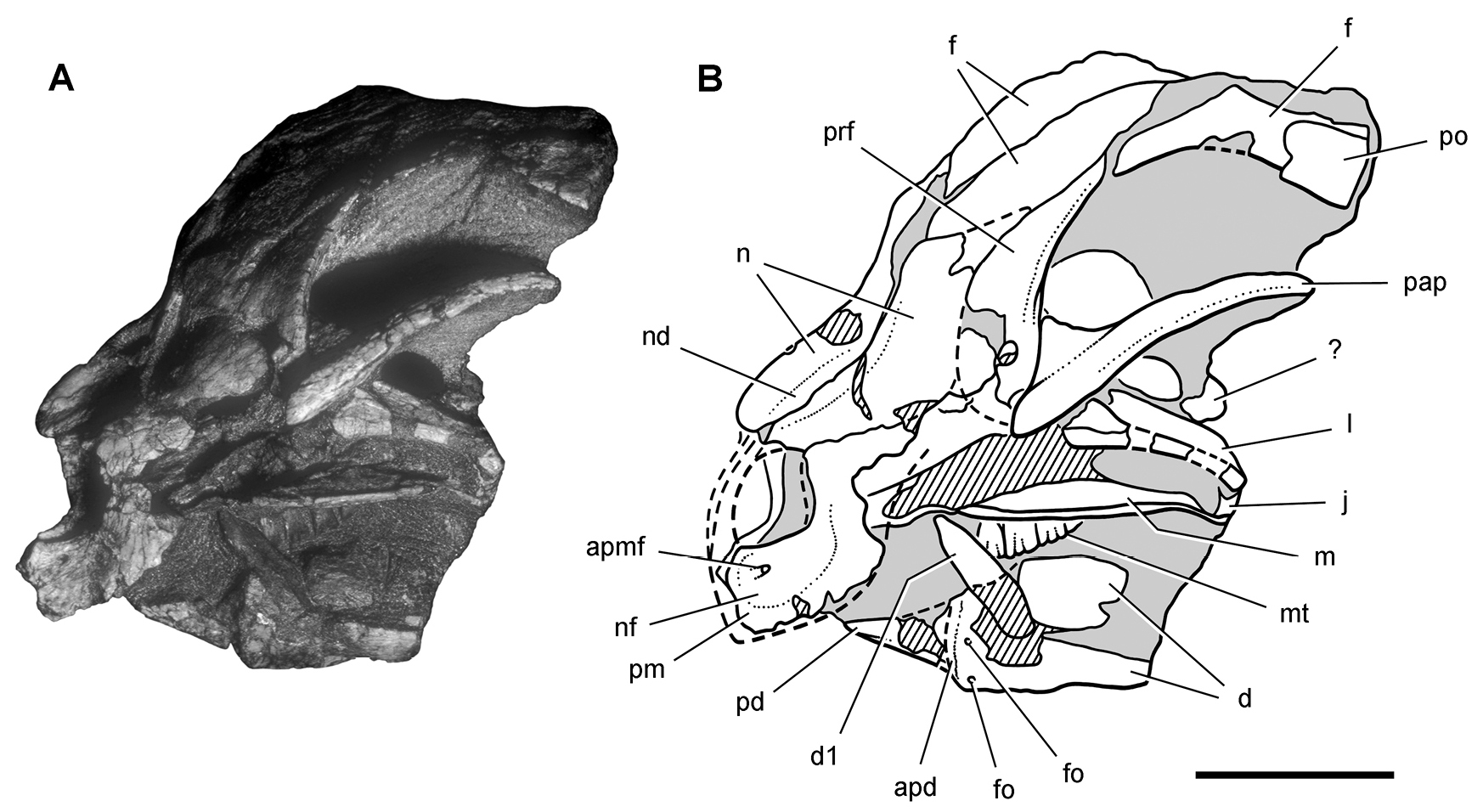
1974년 툴본(Thulborn)은 헤테로돈토사우루스과의 큰 송곳니가 2차 성징을 나타낸다고 제안했다. 이 이론에 따르면 성체 수컷만 완전히 발달된 송곳니를 가지고 있었을 것이며, 송곳니가 전혀 없는 관련 종인 아브릭토사우루스의 모식 표본은 암컷을 나타낼 것이다. 이 가설은 2006년 고생물학자 리처드 버틀러(Richard Butler)와 그의 동료들에 의해 의문을 제기받았는데, 그들은 어린 두개골 SAM-PK-K10487이 초기 발달 단계에도 불구하고 송곳니를 가지고 있다고 주장했다. 이 단계에서는 2차 성징이 나타날 것으로 예상되지 않는다. 또한, 송곳니는 알려진 거의 모든 헤테로돈토사우루스 두개골에서 나타난다. 그러나 성적 이형성이 존재한다면 송곳니를 가진 개체와 송곳니가 없는 개체의 비율이 50:50이 되어야 한다. 유일한 예외는 아브릭토사우루스의 모식 표본이다. 이 개체에서 송곳니가 없는 것은 이 특정 속의 전문화로 해석된다.
6. 고환경
헤테로돈토사우루스는 카루 초집단의 상부 엘리엇 층과 클라렌스 층에서 발견되었는데, 이 지층들은 쥐라기 전기의 헤탕기아절과 시네무리아절에 해당하며 약 2억~1억 9천만 년 전으로 거슬러 올라간다.[1] 원래 헤테로돈토사우루스는 트라이아스기 후기에 살았던 것으로 여겨졌다.
상부 엘리엇 층은 붉은색/자주색 이암과 붉은색/흰색 사암으로 구성되어 있으며, 이보다 약간 젊은 클라렌스 층은 흰색/크림색 사암으로 구성되어 있다. 클라렌스 층은 상부 엘리엇 층보다 화석이 적고, 퇴적물이 종종 절벽을 이루어 화석 사냥꾼의 접근을 제한한다.[1] 상부 엘리엇 층에서는 템노스폰딜리 양서류, 거북, 렙토사우루스, 에토사우루스, 크로커다일모르프, 비포유류 키노돈트를 포함한 풍부한 척추동물 화석이 발견된다.
이 지층에서는 레소토사우루스, 마소스폰딜루스, 메가프노사우루스와 같은 공룡들도 발견되었다. 상부 엘리엇 층은 헤테로돈토사우루스과 공룡들이 다양하게 발견되는 곳으로, 헤테로돈토사우루스 외에도 리코리누스, 아브릭토사우루스, 페고마스탁스 등이 발견되었다. 게라노사우루스는 클라렌스 층에서 발견되는 헤테로돈토사우루스과의 공룡이다.
6. 1. 발견 지역의 지층
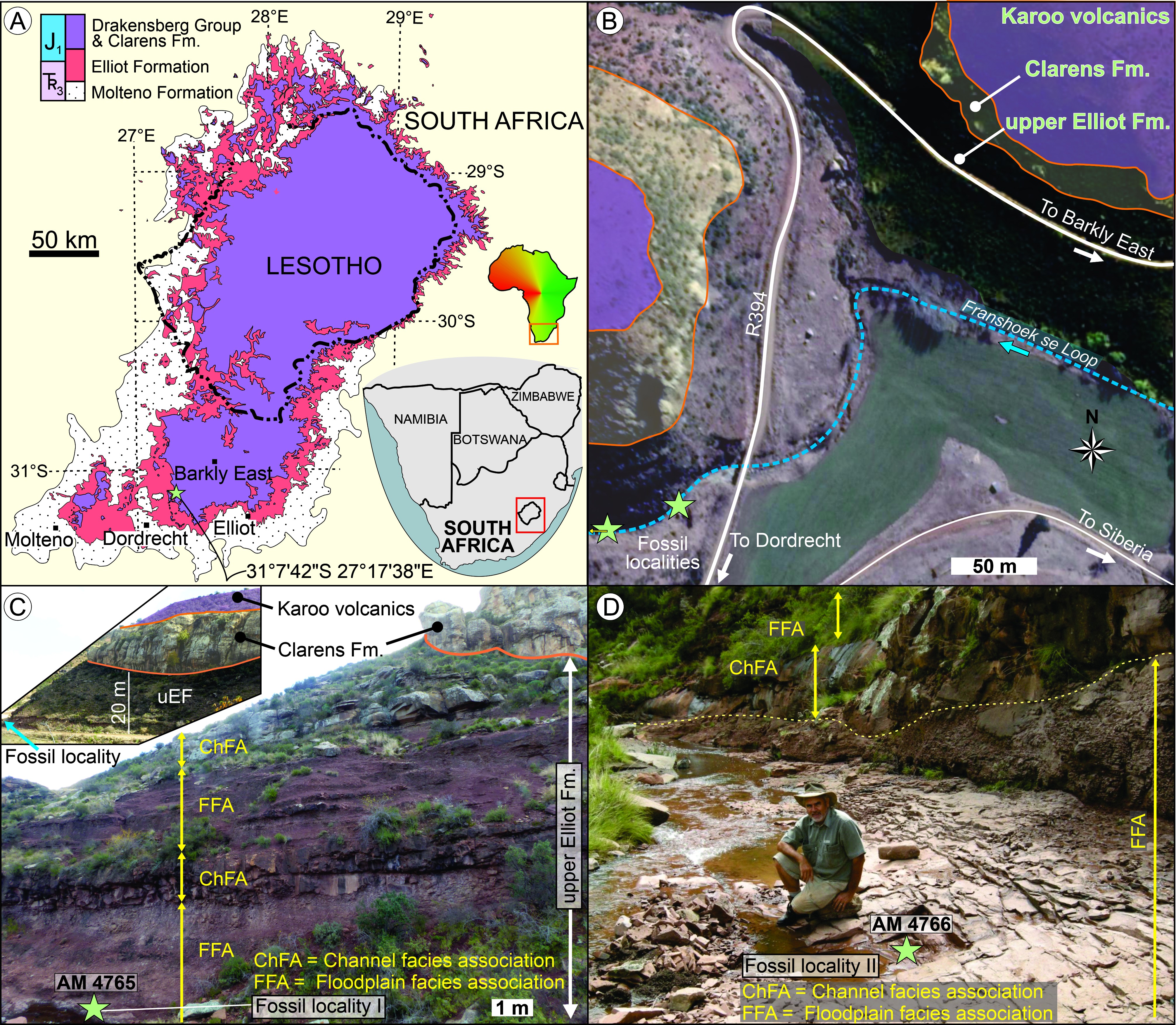
''헤테로돈토사우루스''는 카루 초집단의 상부 엘리엇 층과 클라렌스 층에서 발견되었다. 이 지층들은 쥐라기 전기의 헤탕기아절과 시네무리아절에 해당하며, 약 2억~1억 9천만 년 전으로 거슬러 올라간다.[1] 원래 ''헤테로돈토사우루스''는 트라이아스기 후기에 살았던 것으로 여겨졌다. 상부 엘리엇 층은 붉은색/자주색 이암과 붉은색/흰색 사암으로 구성되어 있으며, 이보다 약간 젊은 클라렌스 층은 흰색/크림색 사암으로 구성되어 있다. 클라렌스 층은 상부 엘리엇 층보다 화석이 적고, 퇴적물이 종종 절벽을 이루어 화석 사냥꾼의 접근을 제한한다.[1] 상부 엘리엇 층은 남부 아프리카의 이 시기 건조한 기후에 적응한 것으로 보이는, 하부 엘리엇 층보다 가볍게 구성된 동물들이 특징이다. 두 지층 모두 템노스폰딜리 양서류, 거북, 렙토사우루스, 에토사우루스, 크로커다일모르프, 비포유류 키노돈트를 포함한 풍부한 척추동물 화석으로 유명하다.
이 지층에서 발견된 다른 공룡으로는 게나사우르 ''레소토사우루스'', 기초 용각류 ''마소스폰딜루스'', 수각류 ''메가프노사우루스''가 있다. 상부 엘리엇 층은 모든 암석 단위에서 가장 큰 헤테로돈토사우루스과 다양성을 보여준다. ''헤테로돈토사우루스'' 외에도 ''리코리누스'', ''아브릭토사우루스'', ''페고마스탁스''가 발견되었다. 헤테로돈토사우루스과의 또 다른 구성원인 ''게라노사우루스''는 클라렌스 층에서 알려져 있다. 높은 헤테로돈토사우루스과 다양성은 연구자들이 서로 다른 종들이 경쟁을 피하기 위해 별도의 식량원을 먹었을 것이라고 결론짓게 했다 (생태적 지위 분할). 특화된 치열을 가진 ''헤테로돈토사우루스''는 질긴 식물 재료에 특화되었을 수 있으며, 덜 특화된 ''아브리크토사우루스''는 주로 부드러운 식물을 소비했을 수 있다. 암석 계열 내 개별 헤테로돈토사우루스과 표본의 위치는 잘 알려져 있지 않아, 이 종들 중 얼마나 많은 종이 실제로 동시대에 살았는지, 어떤 종이 별도의 시기에 존재했는지 결정하기 어렵다.
6. 2. 당시의 환경
''헤테로돈토사우루스''는 카루 초집단의 지층에서 발견된 화석으로 알려져 있다. 여기에는 쥐라기 전기의 헤탕기아절과 시네무리아절에 해당하는 상부 엘리엇 층과 클라렌스 층이 포함되며, 약 2억~1억 9천만 년 전으로 거슬러 올라간다.[1] 원래 ''헤테로돈토사우루스''는 트라이아스기 후기에 살았던 것으로 여겨졌다. 상부 엘리엇 층은 붉은색/자주색 이암과 붉은색/흰색 사암으로 구성되어 있으며, 이보다 약간 젊은 클라렌스 층은 흰색/크림색 사암으로 구성되어 있다. 클라렌스 층은 상부 엘리엇 층보다 화석이 적고, 퇴적물이 종종 절벽을 이루어 화석 사냥꾼의 접근을 제한한다.[1]
상부 엘리엇 층은 남부 아프리카의 이 시기 건조한 기후에 적응한 것으로 보이는 하부 엘리엇 층보다 가볍게 구성된 동물들이 특징이다. 두 지층 모두 템노스폰딜리 양서류, 거북, 렙토사우루스, 에토사우루스, 크로커다일모르프, 비포유류 키노돈트를 포함한 풍부한 척추동물 화석으로 유명하다.
이 지층에서 발견된 다른 공룡으로는 게나사우르 ''레소토사우루스'', 기초 용각류 ''마소스폰딜루스'', 수각류 ''메가프노사우루스''가 있다. 상부 엘리엇 층은 모든 암석 단위에서 가장 큰 헤테로돈토사우루스과 다양성을 보여준다. ''헤테로돈토사우루스'' 외에도 ''리코리누스'', ''아브릭토사우루스'', ''페고마스탁스''가 발견되었다. 헤테로돈토사우루스과의 또 다른 구성원인 ''게라노사우루스''는 클라렌스 층에서 알려져 있다.
높은 헤테로돈토사우루스과 다양성은 연구자들이 서로 다른 종들이 경쟁을 피하기 위해 별도의 식량원을 먹었을 것이라고 결론짓게 했다 (생태적 지위 분할). 특화된 치열을 가진 ''헤테로돈토사우루스''는 질긴 식물 재료에 특화되었을 수 있으며, 덜 특화된 ''아브리크토사우루스''는 주로 부드러운 식물을 소비했을 수 있다. 암석 계열 내 개별 헤테로돈토사우루스과 표본의 위치는 잘 알려져 있지 않아, 이 종들 중 얼마나 많은 종이 실제로 동시대에 살았는지, 어떤 종이 별도의 시기에 존재했는지 결정하기 어렵다.
6. 3. 함께 발견되는 동물들
''헤테로돈토사우루스''(Heterodontosaurus)는 카루 초집단의 상부 엘리엇 층과 클라렌스 층에서 발견되었다. 이 지층들은 쥐라기 전기의 헤탕기아절과 시네무리아절에 해당하며, 약 2억~1억 9천만 년 전으로 거슬러 올라간다.[1] 상부 엘리엇 층은 붉은색/자주색 이암과 붉은색/흰색 사암으로 구성되어 있으며, 이보다 약간 젊은 클라렌스 층은 흰색/크림색 사암으로 구성되어 있다. 클라렌스 층은 상부 엘리엇 층보다 화석이 적고, 퇴적물이 종종 절벽을 이루어 화석 사냥꾼의 접근을 제한한다.[1]
이 지층에서 발견된 다른 공룡으로는 게나사우르 ''레소토사우루스'', 기초 용각류 ''마소스폰딜루스'', 수각류 ''메가프노사우루스''가 있다. 상부 엘리엇 층은 모든 암석 단위에서 가장 큰 헤테로돈토사우루스과 다양성을 보여준다. ''헤테로돈토사우루스'' 외에도 ''리코리누스'', ''아브릭토사우루스'', ''페고마스탁스''가 발견되었다. 헤테로돈토사우루스과의 또 다른 구성원인 ''게라노사우루스''는 클라렌스 층에서 알려져 있다.
참조
[1]
서적
undefined
[2]
웹사이트
Dinosaur fossil found in SA finally gives up its secrets
http://www.news24.co[...]
2016-07-27
[3]
뉴스
ESRF scans most complete ''Heterodontosaurus'' skeleton ever found
http://www.esrf.eu/h[...]
2016
[4]
논문
A new ''Heterodontosaurus'' specimen elucidates the unique ventilatory macroevolution of ornithischian dinosaurs
2021
[5]
서적
The Ultimate Dinosaur Book
https://archive.org/[...]
Dorling Kindersley
[6]
서적
undefined
[7]
웹사이트
New fanged dwarf dinosaur from southern Africa ate plants
http://www.eurekaler[...]
2016-01-28
[8]
서적
undefined
[9]
논문
New reconstruction of cranial musculature in ornithischian dinosaurs: implications for feeding mechanisms and buccal anatomy
2018
[10]
논문
''Heterodontosaurus tucki'' enamel microstructure and dental histology
2023
[11]
서적
undefined
[12]
서적
undefined
[13]
서적
Die Reptilien
Verlag Oeben
[14]
논문
A basal ceratopsian with transitional features from the Late Jurassic of northwestern China
2006
[15]
서적
undefined
[16]
논문
A new hypothesis of dinosaur relationships and early dinosaur evolution
http://eprints.esc.c[...]
2017
[17]
논문
A new phylogeny of cerapodan dinosaurs
2020
[18]
서적
undefined
[19]
서적
Dinosaurs, the encyclopedia
https://archive.org/[...]
McFarland & Company, Inc. Publishers
[20]
논문
Evolutionary Trends in the Jaw Adductor Mechanics of Ornithischian Dinosaurs
2016
[21]
논문
A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of LesothoLesotho
[22]
문서
これは基本的にはほ乳類の歯に見られる特徴である。
[23]
서적
The Age of Dinosaurs
Publications International, LTD.
[24]
뉴스
Tiny Juvenile Dinosaur Fossil Sheds Light on Evolution of Plant Eaters
http://newswise.com/[...]
2008-10-23
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com