스켈리도사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
스켈리도사우루스는 1850년대 영국에서 발견된 초기 조반류 공룡의 한 속이다. 1858년 리처드 오웬에 의해 명명되었으며, 4m까지 성장하는 갑옷 공룡으로, 골질의 갑옷과 스파이크가 특징이다. 1861년 스켈리도사우루스 해리소니가 기준종으로 명명되었으며, 현재까지 발견된 화석을 통해 두개골, 형태, 크기, 갑옷, 생태 등에 대한 연구가 이루어졌다. 스켈리도사우루스는 주로 초식성이었으며, 좁은 머리와 잎 모양의 이빨을 가지고 있었다. 분류학적으로는 안킬로사우루스류와 유사하며, 초기 쥐라기 시대 유럽 섬에서 서식했을 것으로 추정된다.
더 읽어볼만한 페이지
- 장순아목 - 안타륵토펠타
안타륵토펠타 올리베로이는 백악기 후기 남극에서 발견된 몸길이 약 4m의 파란킬로사우리아에 속하는 소형 안킬로사우루스류 공룡으로, 몸 전체가 골판으로 덮여 있고 꼬리에 무기가 있었을 것으로 추정되며, 온난하고 습한 환경에서 다른 공룡들과 함께 서식했다. - 장순아목 - 가르고일레오사우루스
가르고일레오사우루스는 쥐라기 후기에 살았던 안킬로사우루스류 초식 공룡으로, 노트르담 대성당의 가고일 조각상에서 유래된 이름처럼 독특한 두개골과 골격 특징을 지니며 초기 안킬로사우루스류 진화 연구에 중요한 화석이다. - 쥐라기 전기 유럽의 공룡 - 판티드라코
판티드라코는 트라이아스기 후기에 살았던 초기 용각류 공룡의 한 속으로, 테코돈토사우루스 속으로 분류되었던 화석 표본을 기반으로 2003년에 명명되었으며, 잡식성으로 추정되고 이족보행을 했을 가능성이 있다. - 쥐라기 전기 유럽의 공룡 - 드라코랍토르
드라코랍토르는 2014년 웨일스에서 발견된 초기 수각류 공룡으로, 2억 130만 년 전 초기 에탕주절에 살았으며, 2족 보행을 하고 육식을 했을 것으로 추정된다. - 1859년 기재된 화석 분류군 - 콤프소그나투스
콤프소그나투스는 후기 쥐라기에 살았던 작은 육식 공룡으로, 독일과 프랑스에서 발견된 완전한 골격 화석을 통해 알려져 있으며, 깃털 유무, 식성, 조류와의 관계로 논쟁이 있었고 쥬라기 공원에 등장하며 인기를 얻었다. - 1859년 기재된 화석 분류군 - 메갈라니아
메갈라니아는 플라이스토세 시대 오스트레일리아에 서식했던 몸길이 5.5m, 몸무게 575kg 이상으로 추정되는 거대한 왕도마뱀으로, 거대 유대류를 사냥했으며 독을 사용했을 가능성이 있고, 멸종 원인은 불분명하지만 환경 변화와 인류 활동의 영향으로 추정되며 원주민 전승에 영향을 주었을 수 있다.
| 스켈리도사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Scelidosaurus harrisonii |
| 명명자 | 오웬, 1861 |
| 속 명명자 | 오웬, 1859 |
| 어원 | 고대 그리스어 skelis (σκελίς, 갈비뼈) + sauros (σαυρος, 도마뱀) |
| 시대 | 쥐라기 전기 (시네무리아절-플린스바흐절) |
| 생존 시기 | 약 1억 9650만년 전 ~ 1억 8300만년 전 |
| 크기 | 약 4m |
| 분류 | |
| 상목 | 공룡 |
| 목 | 조반목 |
| 목 계통군 | 협룡류 |
| 아목 | 장순아목 |
| 과 | 스켈리도사우루스과 |
| 속 | 스켈리도사우루스속 |
| 종 | |
| 하위 분류 | S. harrisonii (오웬, 1861) (모식종) |
2. 형태
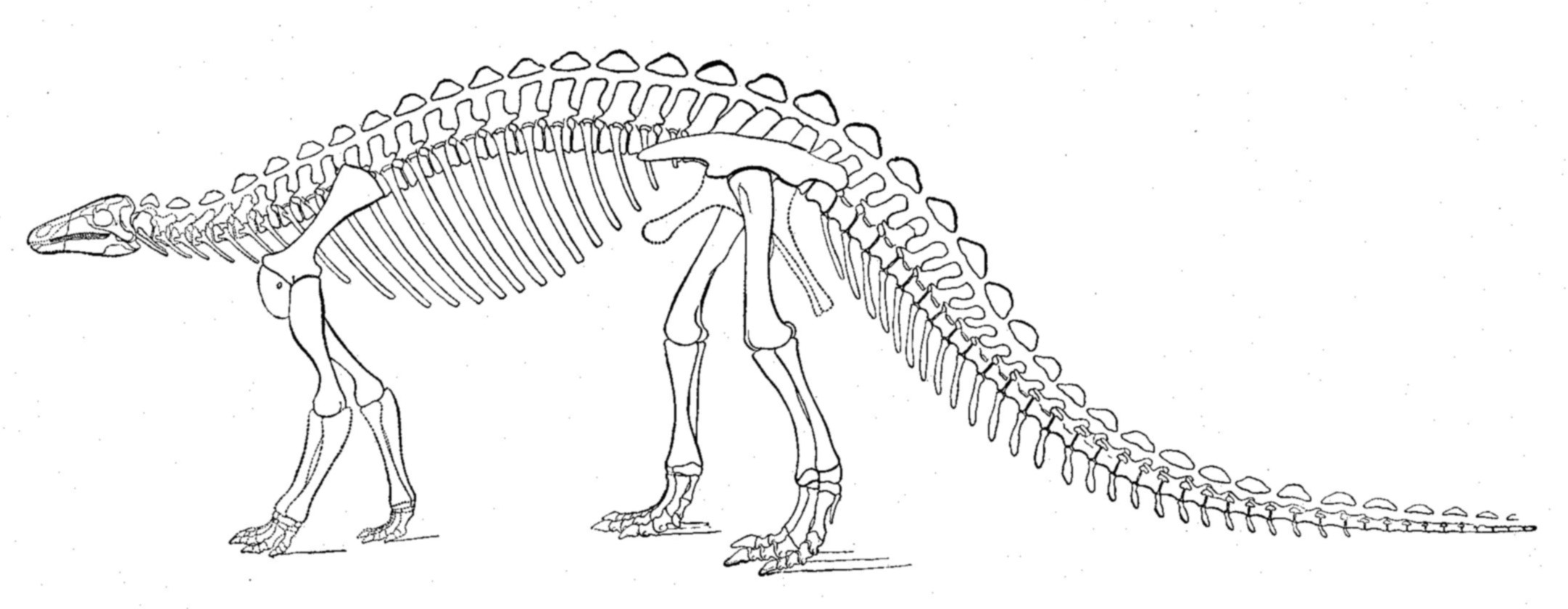
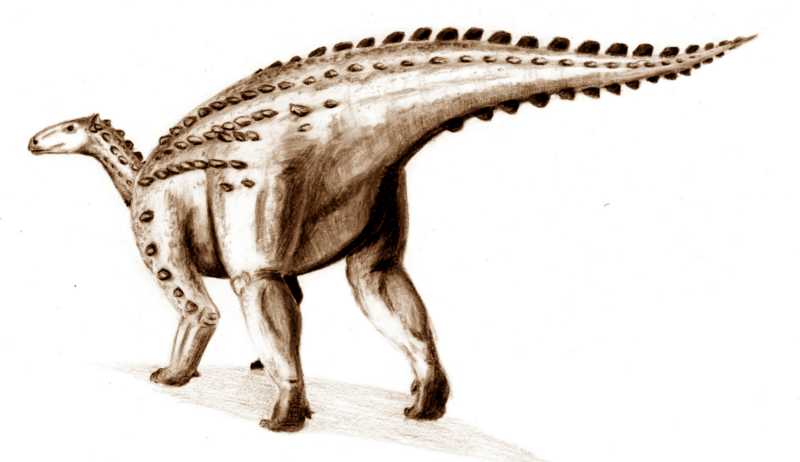
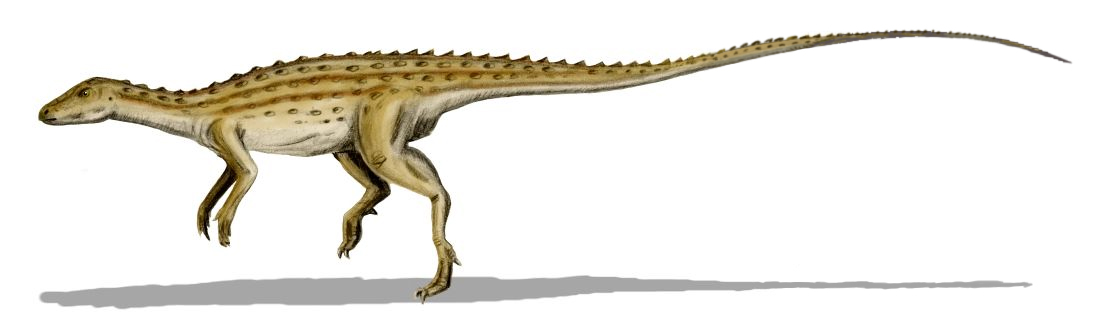
''스켈리도사우루스''는 초기 쥐라기에 살았던 중간 크기의 공룡으로, 다 자란 개체는 몸길이가 약 4m에 달했을 것으로 추정된다.[9][3] 주로 사족 보행을 했으며, 뒷다리가 앞다리보다 길었지만 앞다리도 비교적 길고 튼튼하여 네 발로 걷는 데 적합했다.[41] 머리는 몸에 비해 작고 길쭉했으며[41], 이빨은 식물을 자르기에 적합한 단순한 잎 모양이었다.[3][47] 가장 큰 특징은 몸 전체를 덮는 뼈 골편(피골)으로 이루어진 갑옷이다.[9]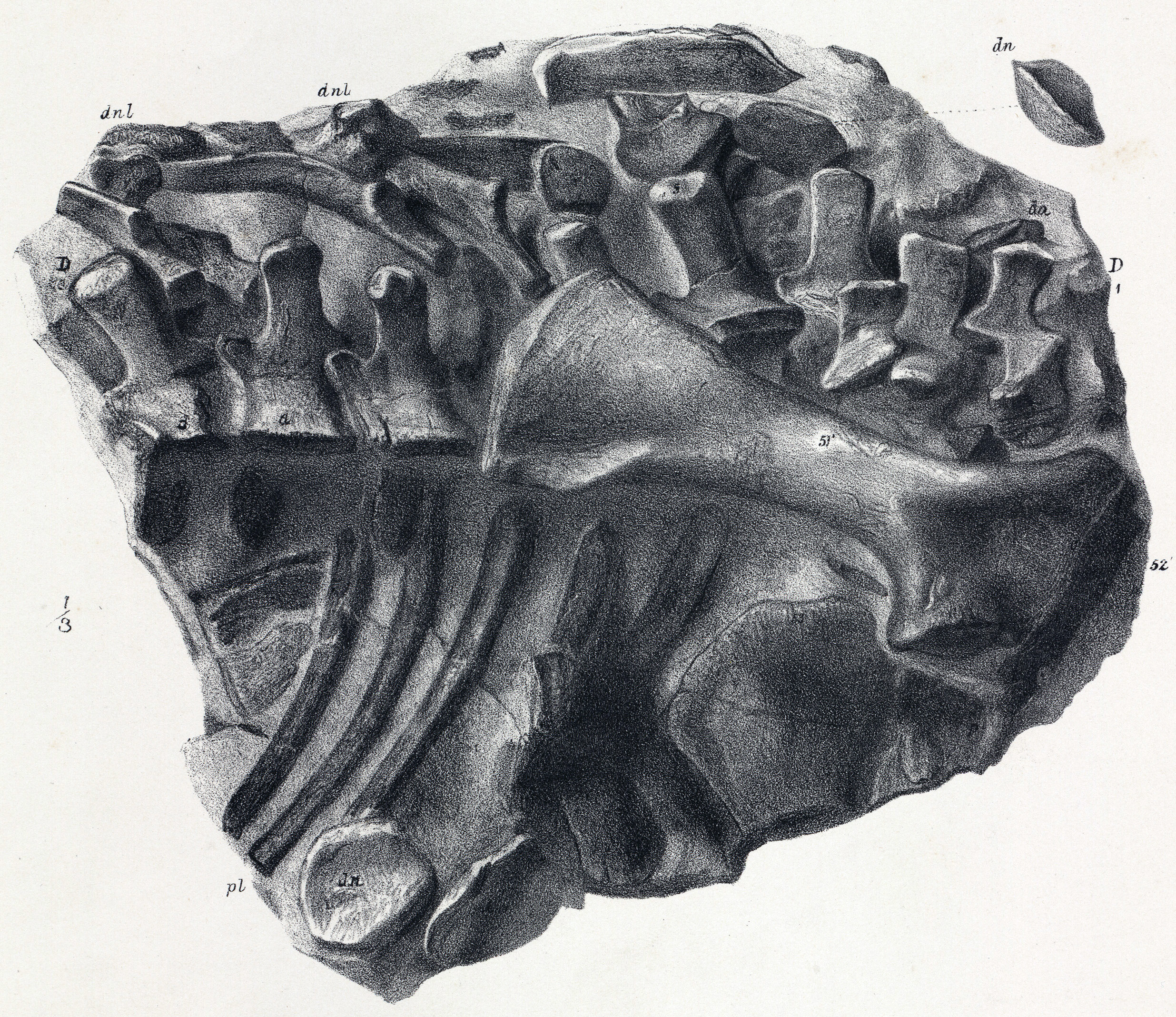
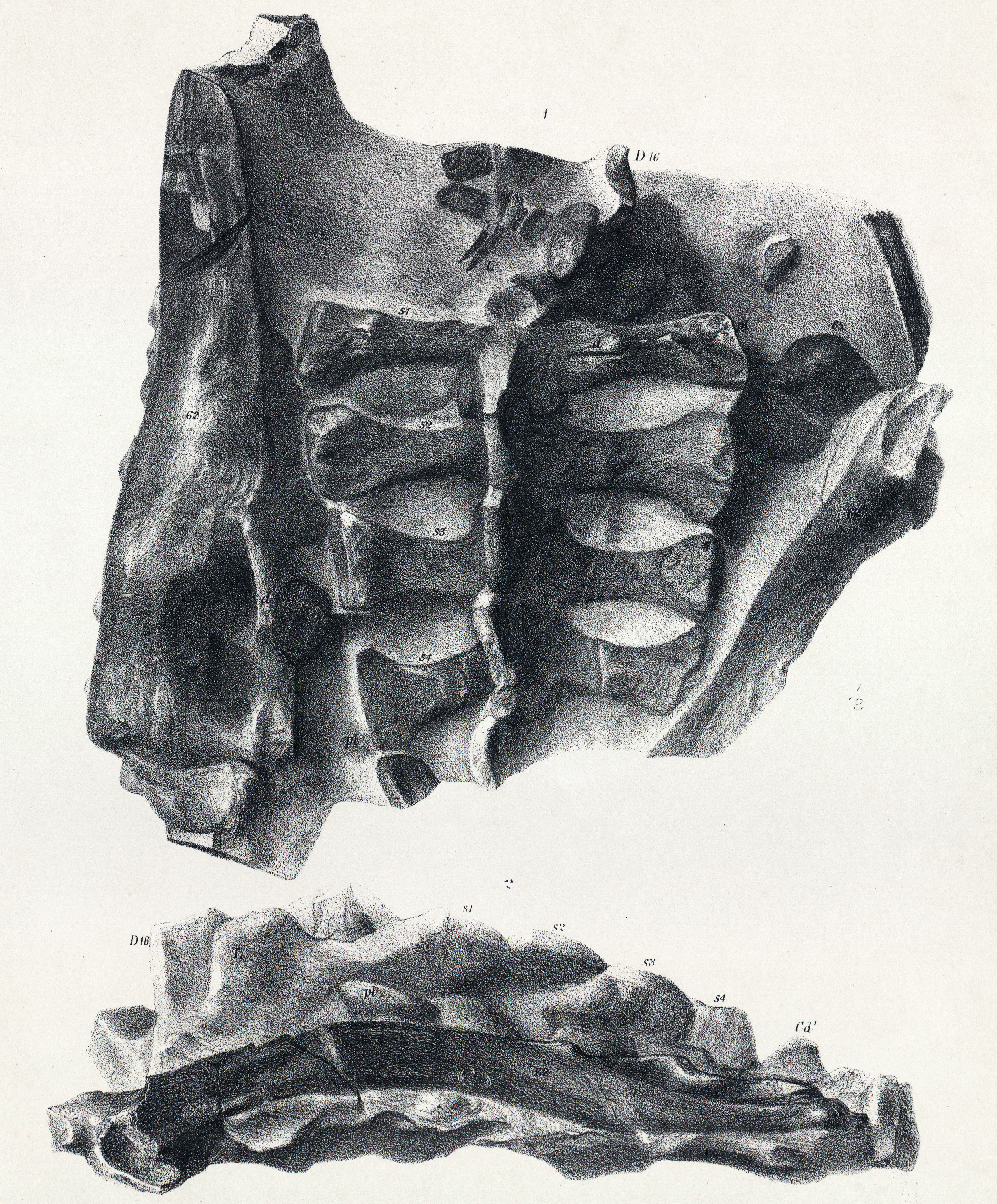
''스켈리도사우루스''의 척주는 최소 6개의 목뼈, 17개의 등뼈, 4개의 엉치뼈, 그리고 최소 35개의 꼬리뼈로 구성되었다.[41][6] 목은 그다지 길지 않았으며, 몸통은 옆에서 보았을 때 비교적 평평했지만 배가 넓어 높이보다 넓지는 않았다. 마지막 세 개의 등뼈에는 갈비뼈가 없었다. 엉치뼈의 가시는 서로 맞닿아 있었으나 하나로 합쳐지지는 않았다.[6] 꼬리는 비교적 짧고 뒤로 갈수록 빠르게 가늘어졌으며, 엉덩이 부분과 꼬리 밑부분은 다수의 골화된 힘줄로 뻣뻣하게 지지되었다.[41]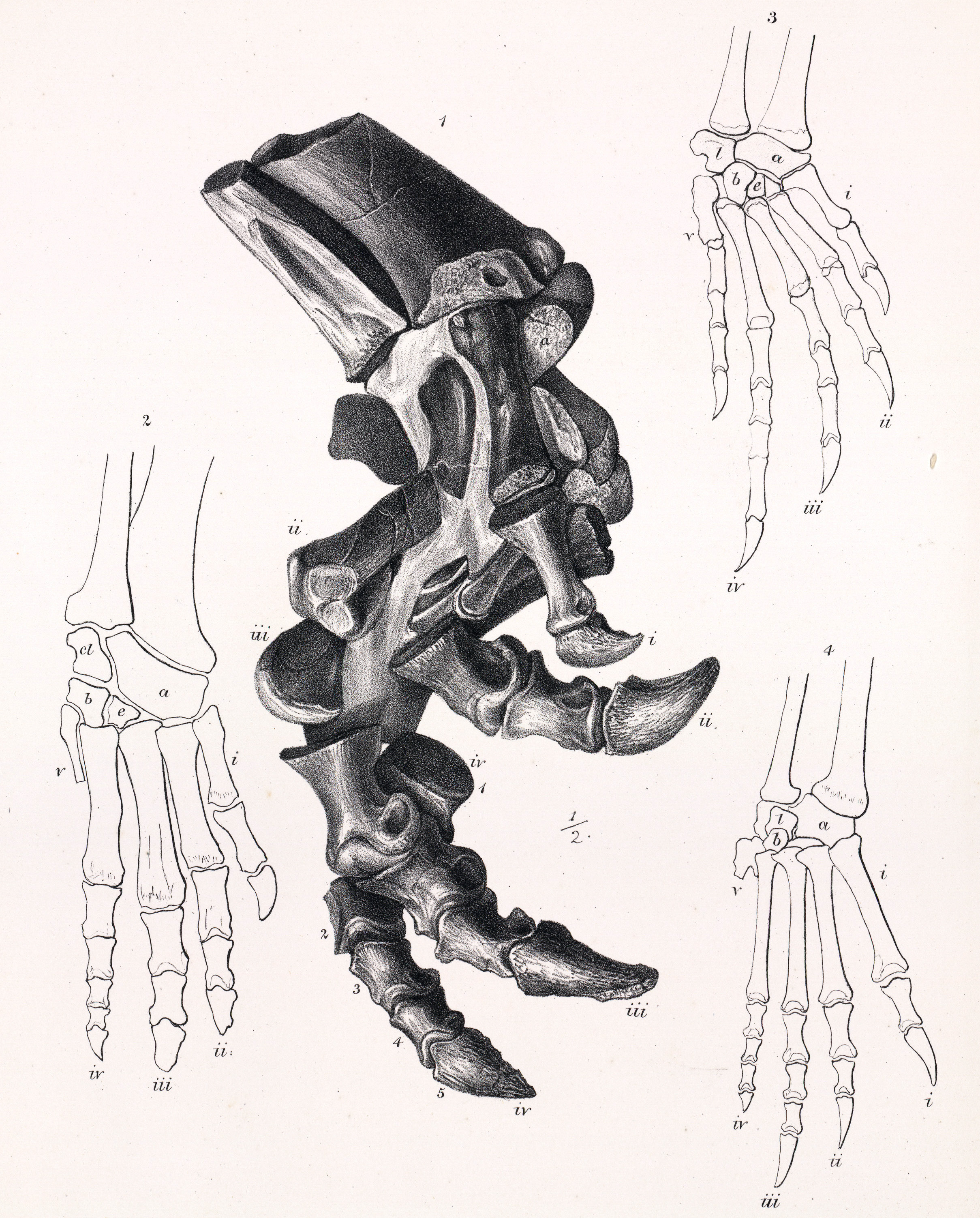
어깨뼈는 짧았고 위쪽 끝이 적당히 넓었으며, 가슴뼈는 옆에서 볼 때 둥근 모양이었다. 앞다리는 적당히 길고 곧으며 튼튼했다. 골반은 상당히 넓었고, 엉덩이뼈는 옆에서 볼 때 곧은 막대 모양의 앞쪽 날개를 가졌으며 배를 위한 공간을 만들기 위해 바깥쪽으로 약간 벌어져 있었다.[6] 두덩뼈는 짧은 앞쪽 돌기를 가졌고, 그 줄기는 아래쪽 끝이 납작한 곧은 궁둥뼈 줄기와 평행하게 뻗어 있었다. 넙다리뼈(허벅지뼈)는 옆에서 보면 곧고 앞에서 보면 약간 바깥쪽으로 휘어 있었다. 넙다리뼈 머리는 뚜렷한 목 없이 몸통과 연결되었고, 뒤쪽에는 꼬리 근육(꼬리넙다리근)이 붙는 네 번째 돌기(전자)가 잘 발달해 있었다. 종아리는 넙다리뼈보다 약간 짧았으며[6], 정강이뼈는 위아래 끝이 튼튼하고 위쪽 앞부분에는 경골 능선이 튀어나와 있었다. 정강이뼈 아래쪽 끝은 위쪽에 비해 약 70° 틀어져 있어 발이 바깥쪽을 향하게 했다. 발은 매우 크고 넓었으며, 네 개의 튼튼한 발가락을 가졌다(다섯 번째 중족골은 흔적만 남음). 발톱은 납작하고 말굽 모양이었으며 안쪽으로 휘어 있었다.[41][6]
2. 1. 크기 및 자세

다 자란 ''스켈리도사우루스''는 초기 쥐라기에는 중간 크기의 공룡이었지만, 대부분의 후기 비조류 공룡에 비해서는 상당히 작았다. 일부 과학자들은 길이를 4m로 추정했다.[9] 2010년, 그레고리 S. 폴은 몸길이 3.8m에 무게는 270kg이라고 추정했다.[3] 이는 스켈리도사우루스과에서는 가장 큰 편에 속한다. 두개골은 비교적 작았다.
''스켈리도사우루스''는 뒷다리가 앞다리보다 긴 사족 보행 동물이었다.[41] 묵직한 몸통과 튼튼한 앞다리 구조 때문에 네 발로 걸었을 것으로 여겨진다. 팔이 상대적으로 길다는 점도 주로 사족 보행 자세를 취했음을 나타낸다.[41] 하지만 나무에서 잎을 뜯어먹기 위해 뒷다리로 일어섰을 수도 있다.[41] 폴란드의 성 십자가 산맥에서 발견된 발자국은 스켈리도사우루스와 유사한 동물이 이족 보행을 하는 것을 보여주며, ''스켈리도사우루스''가 이전에 생각했던 것보다 이족 보행에 더 능숙했을 수 있음을 시사한다.[4]
몸통에는 골질의 갑옷이 발달했으며, 등에는 판 모양의 골판 및 2열의 스파이크가 규칙적으로 배열되어 있었다. 또한 몸 측면과 꼬리 아래쪽에는 다수의 스파이크 열이 있었고, 목에는 3개의 스파이크를 가진 갑옷판 한 쌍이 존재했다. 이빨은 매우 빈약했으며, 주둥이가 폭이 좁은 부리 모양을 하고 있는 것이 특징적이다. 아마도 지표면 근처의 부드러운 식물을 선택적으로 섭취했을 것으로 생각된다.
2. 2. 두개골

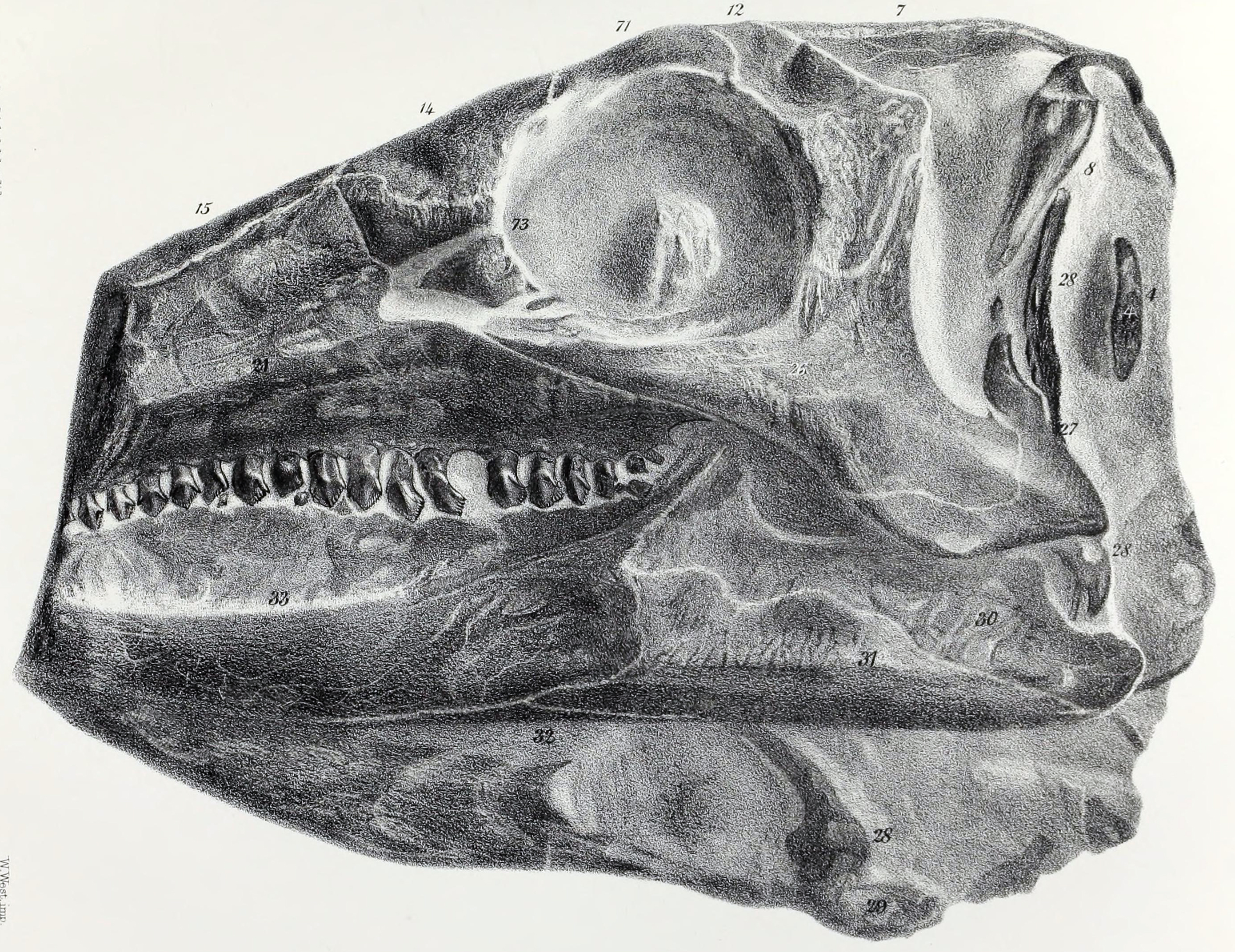
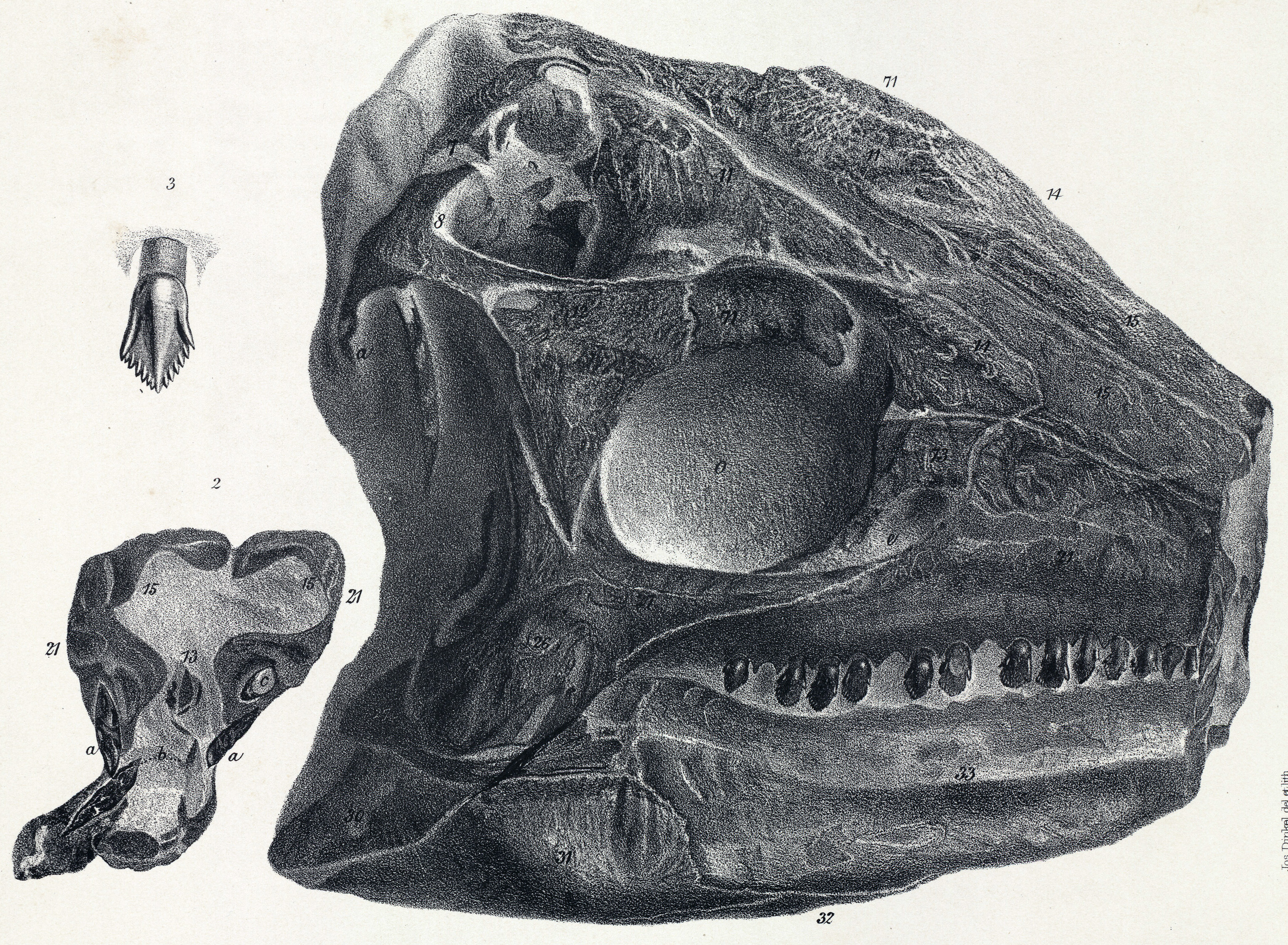
''스켈리도사우루스''의 머리는 작았으며, 길이는 약 20cm 정도로 길쭉한 형태였다. 두개골은 옆에서 보면 낮고 위에서 보면 삼각형 모양으로, 너비보다 길이가 더 길어 초기 조반류와 유사한 특징을 보인다. 주둥이 부분은 주로 비골로 이루어져 있으며 위쪽이 평평하다. ''스켈리도사우루스''는 기저 조반류에서 발견되는 5쌍의 두개골창(skull openings)을 그대로 가지고 있었다. 콧구멍과 안와(눈구멍) 외에도, 많은 후기 장갑류 공룡들과는 달리 전안와창 및 위쪽과 아래쪽의 측두창이 닫히거나 뼈로 덮여 있지 않았다. 특히 위쪽 측두창은 매우 커서 두개골 뒤쪽 상단에 뚜렷한 둥근 구멍을 형성했는데, 이는 아래턱을 닫는 강력한 근육이 부착되는 자리였을 것으로 추정된다. 눈구멍 앞부분은 눈썹 뼈처럼 보이는 융기에 의해 약간 가려져 있었는데, 처음에는 전두정골로 여겨졌으나 2020년 데이비드 노먼은 이것이 융합된 눈꺼풀뼈라고 결론지었다.[22] 눈구멍의 위쪽 가장자리는 상안와골에 의해 형성되었다. 서새나 메이드먼트 등의 연구에 따르면, 어린 개체의 표본에서는 이 뼈가 앞쪽, 뒤쪽, 안쪽에 있는 세 개의 작은 뼈 요소가 융합되어 만들어진 것임을 보여준다.[5]
주둥이 끝을 이루는 전상악골은 짧았다. 아래턱 끝에서 아래쪽 부리의 뼈대를 이루는 전치골은 발견되지 않아, 모든 조반류에 있었을 것으로 추정되는 각질 부리는 매우 짧았을 가능성이 높다. 이빨은 후기의 장갑 공룡들에 비해 옆에서 보았을 때 더 길고 삼각형 모양에 가까웠다.[39] 각 전상악골에는 최소 5개의 이빨이 있었고, 상악골에는 최소 19개, 아래턱의 치골에는 16개의 이빨이 있었다.[41] 하지만 상악골과 치골의 이빨 수는 초기에 발견된 불완전한 두개골 표본 하나를 기준으로 센 것이므로, 실제 이빨 수는 상악골의 경우 최대 약 20개, 아래턱의 경우 아마 26개까지 다양했을 수 있다. 전상악골의 이빨은 다소 길고 약간 휘어 있었다. 뒤쪽으로 갈수록 상악골 이빨의 형태와 점차 비슷해지면서 톱니 구조가 나타나기 시작했다. 상악골과 치골의 이빨은 머리 부분 가장자리에 톱니가 있으며, 이빨 뿌리 부분은 부풀어 오른 형태를 띤다.[41]
한 쌍으로 이루어진 전상악골의 위로 뻗은 가지는 좌우가 합쳐진 비골 사이를 파고드는 형태였는데, 이는 다른 조반류 공룡들과는 반대되는 특징이다. 전두골 표면에는 미세한 능선들이 덮여 있었는데, 이는 현생 거북처럼 케라틴으로 된 판이 덮고 있었음을 시사한다. 뇌를 감싸는 뇌함 앞쪽에는 한 쌍의 도끼 모양으로 골화된 안와접형골이 있어 뇌의 후각엽 바닥 부분을 형성했다. 모식 표본의 두개골은 과거 연구 과정에서 손상되어 입천장 부분의 삼각형 판들이 분리되었다. 이 조각들은 손상되기 전인 1970년대에 노먼이 스케치하여 익상골의 일부로 해석했으나, 2020년 그는 이 뼈들이 비강의 천장을 덮는 특수한 뼈이며 "에피보머(epivomer)"라고 명명해야 한다고 결론 내렸다. 이러한 구조는 다른 동물에서는 알려지지 않은 특징이다.[22]
2. 3. 치아
대부분의 다른 갑옷룡류와 마찬가지로, ''스켈리도사우루스''는 초식동물이었던 것으로 알려져 있다.[3] 그러나 일부 후기 조반류와는 달리 식물 재료를 갈 수 있는 이빨을 가지고 있지 않았다. 대신 초목을 자르는 데 적합한 작고 덜 복잡한 잎 모양의 이빨을 가지고 있었으며, 이빨 자체는 매우 빈약했다.[3] 짧은 턱 관절 때문에 턱은 수직 운동만 가능했다.[3]폴 바렛은 ''스켈리도사우루스''가 이빨끼리 직접 닿지 않고 찌르고 으깨는 방식으로 먹이를 먹었을 것이라고 분석했다. 정밀하지만 단순한 위아래 턱 움직임으로, 음식은 위쪽 이빨의 안쪽 면과 아래쪽 이빨의 바깥쪽 면 사이에서 으깨졌다. 아래쪽 이빨에만 매우 긴 수직 마모 흔적이 발견되는 것은 이빨이 서로 직접 맞닿지 않았음을 시사한다.[47] 이러한 원시적인 이빨과 단순한 턱 구조는 검룡류와 유사한 특징이다.[48]
''스켈리도사우루스''의 식단은 주로 양치류나 구과식물로 구성되었을 것으로 추정된다. 풀은 백악기 후기에 이르러서야 진화했기 때문에 ''스켈리도사우루스''가 살았던 시기에는 존재하지 않았다. 주둥이가 폭이 좁은 부리 모양을 하고 있는 점으로 미루어, 지표면 근처의 부드러운 식물을 선택적으로 섭취했을 가능성이 있다.
스테고사우루스처럼 머리가 좁다는 점은 고품질의 먹이를 선별하여 먹는 식습관을 가졌을 가능성을 시사한다. 그러나 바렛은 ''스켈리도사우루스'' 정도 크기의 동물은 효율적인 소화 발효를 위한 큰 내장을 가지고 있었을 것이므로, 에너지 가치가 높은 소화하기 쉬운 음식 섭취가 작은 동물만큼 결정적으로 중요하지는 않았을 수 있다고 지적했다.[47] 노먼은 ''스켈리도사우루스''가 주로 높이 1m 정도까지 자라는 낮은 관목 식물을 먹었을 것으로 보았다. 뒷다리로 몸을 일으켜 더 높은 곳의 식물을 먹는 것이 해부학적으로 불가능하지는 않았겠지만, 이것이 일반적인 섭식 행동이었을 가능성은 낮다고 보았다.[41]
2. 4. 갑옷

''스켈리도사우루스''의 가장 눈에 띄는 특징은 피부에 박힌 뼈 골편으로 이루어진 갑옷이다. 이 피골들은 몸을 따라 수평으로 평행하게 배열되어 있었다.[9] 오늘날 이런 피골은 악어, 아르마딜로, 일부 도마뱀의 피부에서도 볼 수 있다. ''스켈리도사우루스''의 피골은 크기와 모양이 다양했다. 대부분은 바깥쪽으로 솟아오른 융기가 있는 작거나 큰 타원형 판이었고, 융기의 가장 높은 지점은 뒤쪽에 있었다. 어떤 골편은 작고 평평하며 안쪽이 움푹 파인 형태였다. 더 큰 융기 골편들은 규칙적인 수평 열을 이루며 정렬되었다. 몸통 양쪽에는 이런 골편이 세 줄씩 있었다. 가장 아래쪽 측면 열의 골편은 ''스쿠텔로사우루스''의 칼날 모양 피골보다 더 원뿔 형태에 가까웠다.[7] 이 주요 골편 열 사이에는 작고 타원형이며 융기된 골편이 한두 줄 더 있었다. 꼬리에는 총 네 줄의 큰 골편이 배열되었다. 위쪽 중앙선에 한 줄, 아래쪽 중앙선에 한 줄, 그리고 꼬리 양 측면에 각각 한 줄씩 있었다. 중앙선의 꼬리 골편이 몸통과 목까지 이어졌는지는 확실하지 않으며, 특히 목 부분에서는 그럴 가능성이 낮지만, 종종 ''스켈리도사우루스''는 이런 모습으로 복원되곤 한다.[8]

두개골 표면의 많은 부분은 뼈 돌출물로 덮여 있었고, 눈구멍 위쪽을 감싸며 두개골 뒤쪽에서 위와 뒤로 뻗어 나가는 한 쌍의 뿔을 형성하는 별도의 뼈판도 있었다.[8] 목의 양쪽에는 두 줄의 큰 골편이 있었다. 목 아랫줄의 피골은 매우 크고 평평하며 판 모양이었다. 이 부위의 피부 갑옷은 표면의 뼈 요소 아래에 깊은 기초 판 구조를 포함하고 있었다. 이 기초 판은 성장 구역 역할을 하며, 위에 있는 골편들이 자라면서 두꺼워지고 퍼질 수 있도록 지지하는 역할을 했다. 목 윗줄의 첫 번째 피골은 머리 바로 뒤에서 독특한 세 개의 뾰족한 골편 한 쌍을 형성했다. 이 뾰족한 부분들은 축추의 후관절돌기와 힘줄로 연결되었을 것으로 추정된다.[41][8] 이 골편들의 가장자리는 홈이 파여 결국 서로 맞물리면서, 목의 위쪽과 옆면을 보호하는 뼈 돌출 장식이 있는 연결된 구조(마치 깃과 같은)를 형성했다. 이 구조의 각 부분이 중앙선을 따라 합쳐져 연속적인 목 고리를 만들었는지 여부는 불확실하다. 목의 뼈판은 작고 뾰족한 것부터 평평하고 모자 모양인 것, 높고 능선이 있는 것, 넓고 칼날 모양인 것까지 다양한 형태를 보였다.[8]
목 뒤쪽의 몸통 갑옷에는 기초 판이 없었으며, 몸을 따라 길이 방향으로 세 개의 주요 열로 배열되었다. 가장 큰 골편들은 몸의 측면을 따라 위치했고, 균일하게 타원형이며 능선이 있고 속이 비어 있었다. 작은 골편들은 이 주요 열 사이의 공간을 채웠고, 몸의 나머지 부분은 촘촘하게 배열된 작고 미세한 뼈 구조로 덮여 있었는데, 이 중 다수는 각질화된 비늘을 지지했을 것으로 보인다.[8] 일반적으로 골편은 몸통 앞쪽에서 더 컸고, 뒤쪽으로 갈수록, 특히 허벅지 표면에서 크기가 줄어들었다. 가장 작은 평평하고 둥근 골편들은 더 큰 피골 열 사이의 공간을 채웠을 것이다.[8] 상완에도 수직적인 피골 열이 있었을 가능성이 있다. 후기 곡룡류와 비교했을 때, ''스켈리도사우루스''는 연속적인 판, 가시 또는 골반 방패가 없는 비교적 가벼운 갑옷을 가지고 있었다. 두개골과 아래턱의 거친 표면은 피부가 뼈처럼 단단해진(골화) 것을 나타낸다.[8] 사지는 작은 뼈 결절들이 모자이크처럼 배열되어 있었고, 능선이 있는 좁고 길쭉한 구조의 열로 덮여 있었다. 꼬리는 위, 아래, 옆면에 높고 속이 비어 있으며 능선이 있는 뼈 요소가 네 줄로 배열된 뚜렷한 특징을 보였다. 꼬리 밑부분 근처의 아래쪽에는 간격이 좁고 깊은 뼈 융기가 있었다. 특히 목과 옆면에서 나타나는 이 동물의 뼈 갑옷의 다양성은 더 큰 돌출부가 각질로 덮여 있었고, 더 작고 속이 빈 형태는 다른 기능적 또는 적응적 역할을 했을 수 있음을 시사한다.[8]
발견된 최신 표본 중 일부는 융기가 가시 모양이거나 가시와 유사한 골편을 포함하여 부분적으로 다른 형태의 피골을 보여준다. 이 표본들은 또한 머리 뒤쪽 모서리의 비늘뼈에 작은 뿔이 있었던 것으로 보인다.[26] 화석화된 피부 흔적도 발견되었다. 뼈 골편 사이에서 ''스켈리도사우루스''는 오늘날의 애리조나악어붙이도마뱀(Arizona alligator lizard)처럼 서로 겹치지 않는 둥근 비늘을 가지고 있었다.[9] 큰 골편 사이에는 매우 작은(5mm~10mm) 평평한 뼈 "과립"들이 피부 내에 분포했을 것으로 추정된다. 후기 곡룡류에서는 이러한 작은 골편들이 더 큰 골편으로 발달하여, ''안킬로사우루스''와 같은 속에서 볼 수 있는 여러 피골 판이 융합된 갑옷으로 진화했을 가능성이 있다.[7][8]
등에는 판 모양의 골판 및 두 줄의 스파이크(가시)가 규칙적으로 배열되어 있었다. 또한 몸 측면과 꼬리 아래쪽에는 여러 줄의 스파이크가 있었고, 목 등에는 세 개의 스파이크를 가진 갑옷판 한 쌍이 존재했다.
3. 발견
1850년대 영국 웨스트도싯의 Charmouth에 있는 채석장 주인이었던 제임스 해리슨은 Charmouth와 라임 레지스(Lyme Regis) 사이의 Black Ven 절벽에서 채석된 화석들을 발견했다. 이 화석들은 아마도 시멘트 제조의 원료로 사용되었을 것이다. 해리슨은 이 중 일부를 수집가이자 은퇴한 외과 의사 헨리 노리스에게 주었다. 1858년, 노리스와 해리슨은 일부 조각난 사지 뼈를 런던 영국 박물관(당시 자연사 부서, 현재는 런던 자연사 박물관)의 리처드 오웬 교수에게 보냈다. 이 중에는 왼쪽 대퇴골(표본 GSM 109560)도 포함되어 있었다. 1859년, 오웬은 ''Encyclopædia Britannica''의 고생물학 항목에서 속명 ''스켈리도사우루스''(Scelidosaurus)를 명명했다.[10] 이 항목에는 간단한 설명이 포함되어 있어, 정의가 모호하고 특정 표본이 명시되지 않았음에도 불구하고 이 속명이 유효하게 명명되었음을 시사한다.[11] 오웬은 이 공룡의 이름을 "뒷다리 도마뱀"이라는 의미로 지으려 했으나, 그리스어 단어 'skelos'("뒷다리")를 'skelis'("소갈비")와 혼동했다.[12][13] 이름은 뒷다리가 튼튼하게 발달한 것에서 유래했다. 이후 해리슨은 무릎 관절, 발톱(GSM 109561), 어린 개체의 표본, 그리고 두개골을 오웬에게 보냈고, 오웬은 이를 1861년에 기술했다. 이때 기준종인 ''스켈리도사우루스 해리소니''(Scelidosaurus harrisonii)가 명명되었는데, 종명 '해리소니'는 발견자 해리슨을 기리기 위한 것이었다.[12] 이 두개골은 나중에 오웬이 1863년에 기술한 거의 완전한 골격의 일부임이 밝혀졌다.[14]
영국의 고생물학자 데이비드 B. 노먼은 오웬이 이전에 공룡이 활동적인 사족보행 동물이라고 주장했음에도, ''스켈리도사우루스''가 그 주장의 좋은 예시가 될 수 있었고 당시 발견된 가장 완전한 공룡 중 하나였음에도 불구하고 거의 주목하지 않았다고 지적했다. 노먼은 오웬이 당시 여러 행정 업무, 동료 과학자들과의 논쟁, 그리고 ''시조새''(Archaeopteryx)와 같이 더 흥미로운 새로운 발견들에 대한 연구 등으로 인해 과도한 업무량 때문이었을 것이라고 설명했다.[15] 노먼은 또한 오웬이 1861년에 ''스켈리도사우루스''가 물고기를 먹고 부분적으로 바다에 서식했을 것이라고 추정했는데, 이는 현재 학설과는 매우 다른 생각이었다고 지적했다.[2][12]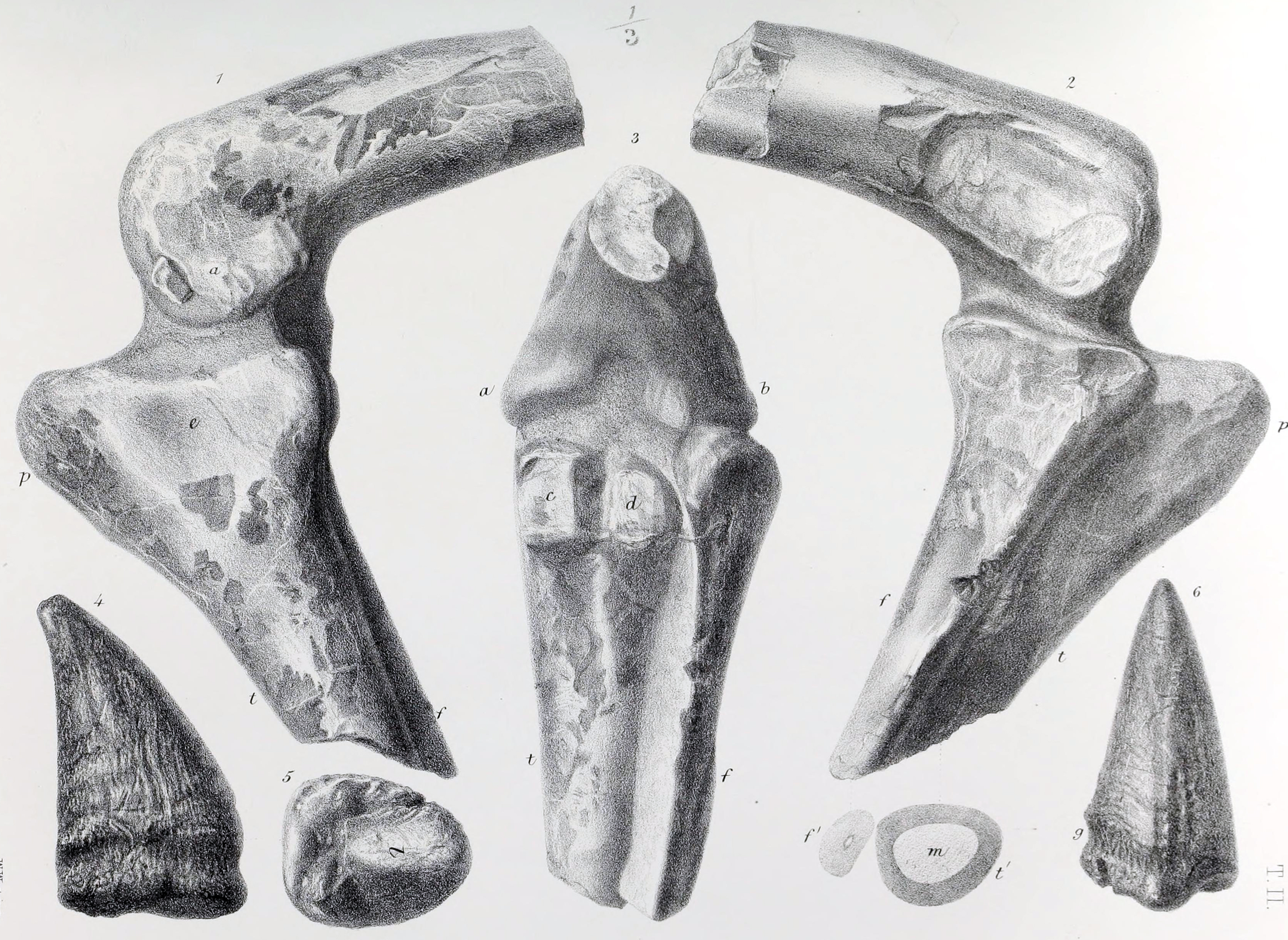
오웬은 완모식표본을 지정하지 않았다. 1888년, 리처드 리데커는 영국 자연사 박물관(NHMUK)의 화석 목록을 작성하면서 1861년에 기술된 사지 파편 중 일부인 대퇴골 하단부와 경골 및 비골 상단부로 구성된 무릎 관절 표본(NHMUK PV 39496)을 ''스켈리도사우루스''의 선정기준표본으로 암묵적으로 선택했다. 리데커가 이 표본을 선택한 이유는 명확하지 않지만, 아마도 크기가 컸기 때문일 수 있다.[16] 그러나 불행하게도, ''스켈리도사우루스'' 화석과 섞여 있던 다른 공룡의 부분적인 유해가 있었고, 이 대퇴골과 경골은 수각류 공룡의 것이었다. 이 사실은 1968년 버나드 뉴먼에 의해 밝혀졌다.[17] 같은 해 뉴먼은 이 무릎 관절이 ''메갈로사우루스''(Megalosaurus)와 관련된 종의 것이라고 생각하며, 리데커의 렉토타입 지정을 국제 동물 명명 위원회(ICZN)가 공식적으로 취소하도록 제안했다. 뉴먼이 사망한 후, 앨런 잭 차리그가 1992년에 이 요청서를 실제로 제출했다.[11] 1994년 ICZN은 이 요청을 받아들여, 의견 1788호를 통해 두개골과 골격으로 이루어진 표본 NHMUK PV R.1111을 ''스켈리도사우루스''의 새로운 렉토타입으로 지정했다.[18] 문제의 무릎 관절 표본은 1995년 사무엘 폴 웰스 등이 비공식적으로 "메로사우루스"(Merosaurus)라고 명명했지만, 이 이름은 아직 정식으로 발표되지 않았다.[19] 이 표본은 아마도 Coelophysoidea 또는 Neoceratosauria에 속하는 수각류의 것일 가능성이 높다.[20] 또한, 뉴먼과 로저 벤슨은 오웬에게 처음 보내졌던 왼쪽 대퇴골(GSM 109560) 역시 수각류의 것이라고 확인했다.[20]
새로운 렉토타입 골격(NHMUK PV R.1111)은 약 1억 9100만 년 전 시네뮤리안 단계에 해당하는 Charmouth Mudstone Formation의 해양 퇴적층인 Black Ven Marl 또는 Woodstone Nodule Bed에서 발견되었다.[21] 이 표본은 두개골과 아래턱을 포함한 상당히 완전한 골격으로 구성되어 있으며, 코 끝, 목 아랫부분, 앞다리, 꼬리 끝부분만 빠져 있다. 수백 개의 골편이 골격과 함께 발견되었고, 대부분 원래 위치에 있었다. 1960년대부터 이 화석은 로널드 크라우처에 의해 산성 용액을 사용하여 주변 암석에서 뼈를 분리하는 방식으로 추가적으로 처리되었다. 1992년 차리그는 단 하나의 암석 블록만 아직 처리되지 않았다고 보고했지만,[11] 그 결과를 발표하기 전에 사망했다. 이 작업을 이어받은 노먼은 2004년에 몇 가지 새로운 해부학적 세부 사항을 공개했으며,[41] 이 외에는 현대적인 상세 기술이 거의 없었다.[21] 2020년 노먼은 두개골과 골반뼈에 대한 논문을 발표했는데, 이 과정에서 산성 용액 누출과 부주의한 취급으로 인해 뼈의 상태가 심하게 악화되고 파손 및 붕괴가 발생했음이 밝혀졌다.[22]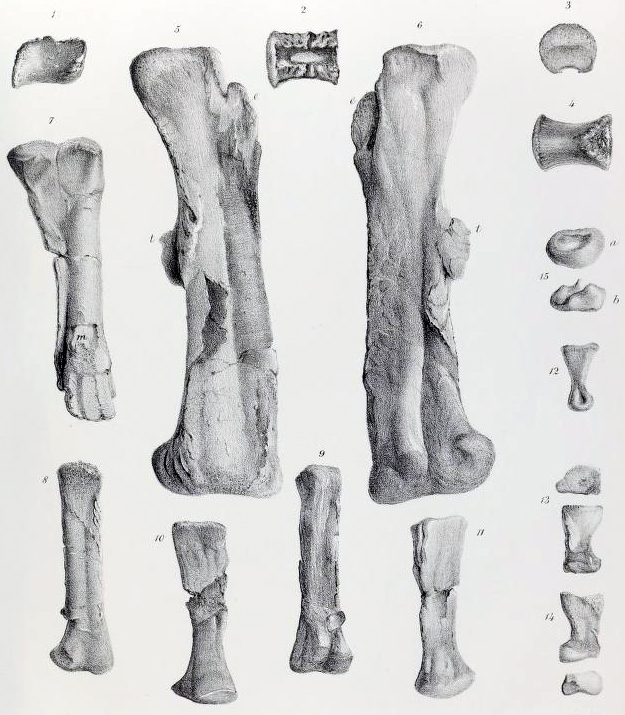
렉토타입 외에도 다른 ''스켈리도사우루스'' 화석들이 알려져 있다. 1888년 리데커는 노리스 컬렉션에서 NHMUK가 입수한 다수의 단일 뼈, 주로 사지 골격과 골판들을 목록화했다.[16] 오웬은 1861년에 어린 개체의 부분적인 두 번째 골격을 기술했는데, 이 표본은 나중에 엘리자베스 필포트의 컬렉션에 추가되었고 현재는 라임 레지스 박물관에 표본 LYMPH 1997.37.4-10으로 소장되어 있다. 오웬은 이 어린 개체가 비교적 크다는 점과 자신이 추정한 해양 생활 방식을 근거로 ''스켈리도사우루스''가 난태생이었을 수도 있다고 추측했다.[12] 이 표본의 짧은 전치골(prepubic process)은 과학자들이 이 구조가 주요 치골체와 동일하지 않다는 것을 확신하는 데 도움을 주었다. 당시 일부 학자들은 조반목 공룡의 뒤쪽을 향하는 얇은 치골이 대부분의 파충류 그룹에서 앞쪽을 향하는 훨씬 더 큰 치골과 상동적이라고 믿기 어려워했다.
더 최근에는 상업적 채석보다는 아마추어 고생물학자들의 노력 덕분에 Charmouth에서 새로운 발견들이 이루어졌다. 1968년에는 두 번째 부분적인 어린 개체 골격(표본 NHMUK PV R6704)이 기술되었는데,[23] 이는 이미 1959년에 보고된 바 있었다.[24] Charmouth의 지질학자 제임스 프레데릭 잭슨(1894–1966)이 발견한 이 표본은 약간 더 젊은 지층인 플린스바키안 초기(약 1억 9천만 년 전)의 Stonebarrow Marl Member에서 나왔다.[21] 1985년에는 사이먼 반슬리, 데이비드 코스타인, 피터 랭햄이 매우 완전한 두개골과 피부 흔적을 포함한 부분적인 골격을 발굴했다.[25] 이 표본은 브리스톨 시립 박물관 및 미술관에 판매되어 표본 BRSMG CE12785로 등록되었다. 표본 CAMSMX.39256은 케임브리지의 세지윅 박물관 컬렉션의 일부이다.[21] 여러 표본은 개인 소장품에 보관되어 있어 아직 기술되지 않았다. 여기에는 2000년 데이비드 솔이 발견한 길이 3.1m의 골격이 포함되는데, 이는 아마도 영국 제도에서 발견된 가장 완전한 비조류 공룡 표본일 것이다. 골격의 모든 부위가 현재 알려져 있다.[21] 솔의 발견은 갑옷의 세부 사항에서 렉토타입과 차이가 있어, 별도의 분류군을 나타내거나 성적 이형성을 반영할 가능성이 제기되었으나,[26] 2020년 노먼은 이를 부인했다.[22]
1980년부터 2000년 사이에 고생물학자 로저 번은 북아일랜드 더 고빈스 근처 해변에서 세 개의 화석을 발견했다. 정확한 지질학적 출처는 기록되지 않았지만, 표본의 매우 어두운 색상은 리아스 군 암석, 구체적으로는 워털루 머드스톤 형성의 Planorbis Zone 또는 Pre-planorbis Zone에서 유래했음을 시사한다. 표본에는 1980년 1월 발견된 대퇴골 근위부 조각(BELUM K3998), 1981년 4월 발견된 경골 몸통 조각(BELUM K12493), 2000년 발견된 작은 오각형 물체(BELUM K2015.1.54)가 포함된다. 조직학자 로빈 리드는 뼈 조직과 구조를 통해 첫 번째 표본을 공룡으로 인식하고 1989년에 보고했으며, ''스켈리도사우루스'' 또는 유사한 동물에 속할 것으로 의심했다. 번은 이후 유사한 근거로 경골 표본도 공룡으로 인식했다. 두 표본은 같은 동물에서 유래했을 것으로 추정되었고, 오각형 물체는 같은 논리에 따라 스켈리도사우루스 골판으로 추정되었다.[27]
이 아일랜드 표본들은 19세기 말 또는 20세기 초 화석 수집가 윌리엄 그레이가 발견한 또 다른 표본과 함께 마이클 J. 심스 등에 의해 정식으로 연구되었고, 2021년 12월 ''지질학자 협회 회보''에 발표되었다. 대퇴골 조각은 명확한 조반목 공룡의 특징을 가지며 크기와 형태가 ''스켈리도사우루스''와 매우 유사하고 가까운 친척인 ''스쿠텔로사우루스''와는 다르다는 점에서 ''스켈리도사우루스''로 잠정적으로 분류되었다. 그러나 경골은 불확실한 수각류의 것으로 재해석되었고, 오각형 물체는 화석과 유사한 현무암 조각으로 밝혀졌다. 그레이의 표본은 어룡의 것으로 확인되었다. 스켈리도사우루스 대퇴골과 수각류 경골은 아일랜드에서 발견된 유일한 공룡 유해이며, 아일랜드는 중생대 화석 기록이 빈약한 해양 지역으로 구성되어 있다. 스켈리도사우루스 표본은 섬에서 보고된 최초의 공룡 화석이었다.[27] 그러나 2024년 새첼은 이 대퇴골 조각(BELUM K3998)을 ''스켈리도사우루스''나 조반목 공룡에 속하지 않는 불확실한 공룡 유해로 재식별했다.[28]
2000년, 데이비드 마르틸 등은 cf. ''스켈리도사우루스'' sp.(즉, ''스켈리도사우루스'' 속에 잠정적으로 분류되지만 특정 종으로 지정되지 않은 표본)에 해당하는 표본에서 연조직 보존 사례를 발표했다. 표본 번호 BRSMG CF2781인 이 화석은 1990년대 초에 이미 준비된 상태로 고 존 챌리너 교수의 유품에서 발견되었으며, 그는 이를 강의에 사용했다. 그 출처는 알려지지 않았다. 이 화석은 탄산염 이암 판에 보존된 8개의 꼬리 척추로 구성되어 있으며, 헤탕기안 후기에서 시네뮤리안 동물상 단계에 해당할 것으로 추정된다. 화석의 일부는 척추 주위에 연조직의 흔적이 보이는 방식으로 보존되었으며, 골편 위에 표피층이 존재했음을 보여준다.[7] 연구자들은 이를 근거로 모든 기저 갑옷 공룡의 골판이 단단하고 아마도 케라틴질의 피부층으로 덮여 있었을 것이라고 결론지었다.[7]
오언이 명명하고 기술한 ''스켈리도사우루스 해리소니''(''S. harrisonii'')는 여러 거의 완전한 골격을 바탕으로 현재 유일하게 인정되는 종이다. 중국 하부 루펑 지층의 시네무리아 시대 지층에서 발견된 것으로 추정되는 두 번째 종 ''스켈리도사우루스 욀레리''(''S. oehleri'')는 1965년 데이비드 제이 시몬스에 의해 별도의 속인 ''타티사우루스''로 기술되었다. 1996년 스펜서 G. 루카스는 이를 ''스켈리도사우루스'' 속으로 옮겼다.[29] 그러나 화석이 단편적이기 때문에 이러한 재평가는 널리 받아들여지지 않았고, 오늘날 ''S. 욀레리''는 다시 ''타티사우루스''로 인정받고 있다.[41][30]
1989년, 미국 북부 애리조나의 카이엔타 지층(글렌 캐년 그룹)에서 발견된 골판들이 케빈 패디언에 의해 ''스켈리도사우루스'' sp.로 언급되었다. 당시에는 ''S. 해리소니''가 초기 시네무리아 시대에 해당한다고 여겨졌기 때문에, 이 발견은 해당 지층의 연대를 추정하는 데 사용되었다.[31] 이 골판들은 이전에 ''스켈리도사우루스'' 화석이 발견된 유럽과 애리조나의 글렌 캐년 지역 사이의 지리적 연관성을 시사하는 것으로 여겨졌다.[31] 그러나 이후 과학자들은 골판의 형태가 다르다는 이유로 이 표본들을 ''스켈리도사우루스''로 분류하는 것을 거부했다.[41][21] 2014년 로만 울란스키는 이 표본들을 바탕으로 새로운 종인 ''S. 아리조넨시스''(''S. arizonensis'')를 명명했으나,[32] 2016년 피터 말콤 갈턴과 케네스 카펜터는 이를 명칭 불확실 종(nomen dubium)으로 간주하고, 이 표본들을 미확정 갑옷을 지닌 공룡(Thyreophora indet.)으로 분류했다.[33]
4. 분류
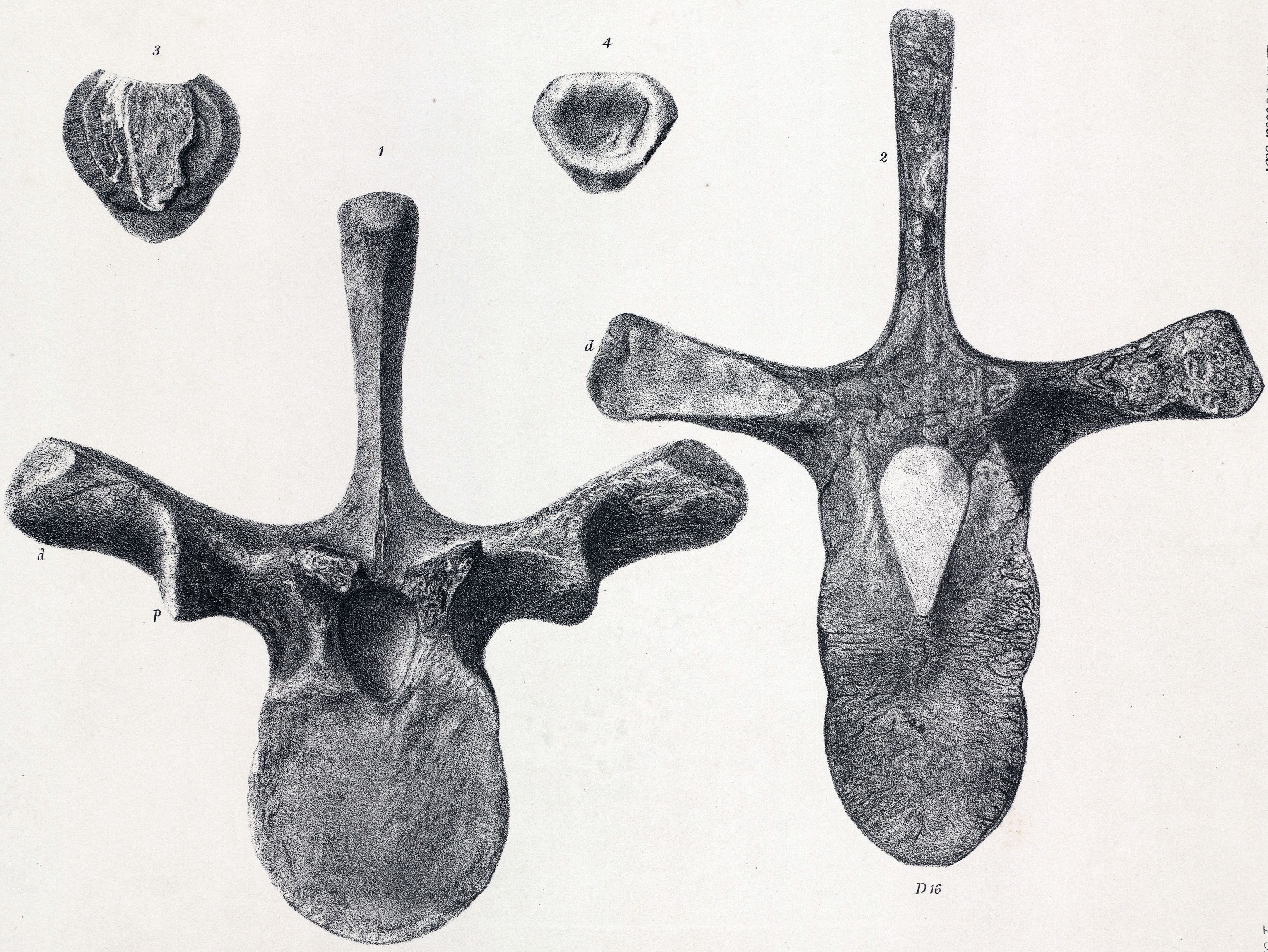
''스켈리도사우루스''(''Scelidosaurus'')는 1861년 오언에 의해 공룡(Dinosauria)으로 처음 분류되었다. 1868년 또는 1869년에 에드워드 드링커 코프는 '''스켈리도사우루스과'''(Scelidosauridae)라는 분류군을 제안했으나, 이는 1871년 12월에야 공식적으로 출판되었다.[34] 따라서 1869년에 스켈리도사우루스과를 정식으로 명명한 사람은 토머스 헨리 헉슬리로 인정된다.[11][35] 19세기에는 당시 알려진 거의 모든 갑옷 공룡이 스켈리도사우루스과에 포함되는 것으로 여겨졌다. 20세기 후반에는 이 용어가 안킬로사우리아(Ankylosauria)와 스테고사우리아(Stegosauria)의 조상에 가까운 원시적인 조반류 공룡들을 묶는 데 사용되기도 했다. 예를 들어 ''스쿠텔로사우루스''(Scutellosaurus), ''에마우사우루스''(Emausaurus), ''루시타노사우루스''(Lusitanosaurus), ''타티사우루스''(Tatisaurus) 등이 여기에 해당되었다.[9] 오늘날 대부분의 고생물학자들은 스켈리도사우루스과가 실제 자연적인 그룹(즉, 계통군, clade)을 형성하지 않는 다계통군(polyphyletic group)이라고 생각한다. 하지만 벤턴(2004)과 같이 이 그룹을 단일계통군(monophyletic group)으로 보는 학자도 있다.[36] 스켈리도사우루스과는 2001년 중국의 고생물학자 둥즈밍이 ''스켈리도사우루스''와 가까운 관계인 티레오포라 공룡 ''비에노사우루스''(Bienosaurus)를 기술하면서 다시 주목받기도 했다.[37]
''스켈리도사우루스''는 조반류 공룡으로, 1911년 ''게라노사우루스''(Geranosaurus)가 기술되기 전까지 가장 오래된 조반류 중 하나로 여겨졌다.[41] 20세기 동안 ''스켈리도사우루스''의 분류학적 위치는 여러 차례 바뀌었다. 알프레트 폰 치텔(1902), 윌리엄 엘긴 스윈턴(1934), 로버트 애플비 등(1967)은 이 공룡을 스테고사우루스류로 분류했다.[38] 하지만 당시 스테고사우루스류는 모든 종류의 갑옷 공룡을 포괄하는 넓은 의미로 사용되었다. 1968년 로머는 ''스켈리도사우루스''를 안킬로사우루스류로 분류했다.[38] 1977년에는 퀸즐랜드 대학교의 리처드 썰본이 ''스켈리도사우루스''를 ''테논토사우루스''(Tenontosaurus)나 ''이구아노돈''(Iguanodon)과 유사한 조각류 공룡으로 재분류하려는 시도를 했다.[38] 썰본은 ''스켈리도사우루스''가 달리기에 적합한 가볍고 이족보행을 하는 공룡이라고 주장했지만, 그의 이론은 이후 받아들여지지 않았다.
이 논쟁은 여전히 진행 중이다. 현재, ''스켈리도사우루스''는 스테고사우루스류보다 안킬로사우루스류와 더 밀접한 관련이 있거나, 확장하여 진정한 안킬로사우루스이거나,[39][40] 안킬로사우루스-스테고사우루스 분할의 기저로 간주된다.[41] 스테고사우루스 분류는 인기를 잃었지만, 오래된 공룡 책에서 볼 수 있다.[42] 분지학 분석은 예외 없이 ''스켈리도사우루스''의 기저 위치를 유리포다 외부로 복원했다.[21] 조반류 계통 발생에 대한 포괄적인 검토에서, 안드레 폰세카와 동료들은 ''스켈리도사우루스''를 유리포다 외부의 기저 티레오포란으로 회복시켰고, ''피로코드''(PhyloCode)에서 '''스켈리도사우루스과'''를 "''스켈리도사우루스 해리소니''(Scelidosaurus harrisonii)를 포함하지만, ''안킬로사우루스 마그니벤트리스''(Ankylosaurus magniventris)와 ''스테고사우루스 스테놉스''(Stegosaurus stenops)는 포함하지 않는 가장 큰 계통군"으로 정의했다.[43]
2011년의 분지학적 연구에서는 ''스켈리도사우루스''가 ''스쿠텔로사우루스''나 ''에마우사우루스''보다는 분화되었지만, 유리포다(스테고사우리아와 안킬로사우리아를 포함하는 그룹)보다는 더 원시적인 위치에 있는 것으로 나타났다.[44]
2022년 모노그래프에서 데이비드 노먼은 티레오포란 간의 다른 관계를 제안했는데, 스테고사우리아가 가장 기저 그룹이고 ''스켈리도사우루스''는 안킬로사우리아와 가장 밀접한 관련이 있다는 것이다.[45] 이 연구에서 제시된 관계는 다음과 같다:

''스켈리도사우루스''보다 기저에 있는 티레오포란의 화석 기록은 희소하다. 또한 애리조나에서 발견된 더 "원시적인" ''스쿠텔로사우루스''(Scutellosaurus)는 선택적 이족 보행을 했던 초기 속이었다. 약 1억 9천 5백만 년 전 프랑스에서 초기 갑옷 공룡의 트랙웨이가 발견되었다.[46] 이 기저 티레오포란의 조상은 후기 트라이아스기 동안 ''레소토사우루스''(Lesothosaurus)와 유사한 초기 조반류에서 진화했다.[41]
5. 생태
대부분의 다른 갑옷룡류와 마찬가지로, ''스켈리도사우루스''는 초식동물이었던 것으로 알려져 있다. 그러나 일부 후기 조반류는 식물 재료를 갈 수 있는 이빨을 가지고 있었지만, ''스켈리도사우루스''는 초목을 자르는 데 적합한 작고 덜 복잡한 잎 모양의 이빨을 가지고 있었고, 짧은 턱 관절로 인해 수직 운동만 가능한 턱을 가지고 있었다.[3] 폴 바렛은 ''스켈리도사우루스''가 이빨 대 이빨 작용으로 찌르고 으깨는 방식으로 먹이를 먹었으며, 정밀하지만 단순한 위아래 턱 움직임으로, 음식은 위쪽 이빨의 안쪽과 아래쪽 이빨의 바깥쪽 사이에서 으깨졌으며, 아래쪽 이빨에만 매우 긴 수직 마모면이 있는 것으로 보아 실제로 이빨이 서로 닿지 않았다고 보았다.[47]
이러한 측면에서 그것은 또한 원시적인 이빨과 단순한 턱을 가진 검룡류와 유사했다.[48] 그것의 식단은 양치류나 구과식물로 구성되었을 것으로 추정된다. 백악기 후기에 이르러서야 풀이 진화했기 때문에 ''스켈리도사우루스'' 시대에는 풀이 없었다.
스테고사우루스와의 또 다른 유사점은 좁은 머리인데, 이는 고품질 사료로 구성된 선택적 식단을 나타낼 수 있다. 그러나 바렛은 ''스켈리도사우루스'' 크기의 동물은 효율적인 발효를 가능하게 하는 큰 내장을 가지고 있어, 작고 종종 그것에 결정적으로 의존하는 작은 동물보다 에너지 가치가 높은 소화하기 쉬운 음식의 섭취가 덜 중요하다고 지적했다.[47] 노먼은 ''스켈리도사우루스''가 높이가 1m까지 되는 낮은 관목 식물을 먹었다고 결론지었다. 뒷다리로만 몸을 일으켜서 섭식 범위를 수직으로 증가시킬 수 있었고 아마도 해부학적으로 가능했을 것이지만, 노먼은 그것이 행동의 관련 부분이라고 의심했다.[41]
초기 쥐라기 동안, 유럽은 섬의 군도를 형성했다.[49] Charmouth 노두는 당시 섬을 형성했던 여러 구조적 고지에 인접해 있었으며, 북쪽으로는 웨일스 고지 (현재 웨일스의 상당 부분을 포함), 서쪽으로는 코누비안 매시프 (현재 데번과 콘월을 포함)가 있었다.[50] 퇴적물에서 ''Classopolis'' 꽃가루가 발견된 것은 멸종된 Cheirolepidiaceae과의 침엽수가 섬에서 자랐음을 시사한다.[51]
참조
[1]
서적
Greek-English Lexicon, Abridged Edition
Oxford University Press, Oxford, UK
1980
[2]
서적
"Scelidosaurus, the earliest complete dinosaur"
Indiana University Press, Bloomington
2001
[3]
서적
The Princeton Field Guide to Dinosaurs
Princeton University Press
2010
[4]
논문
Tracks of a large thyreophoran from the Early Jurassic of Poland
https://www.app.pan.[...]
2016-07-27
[5]
논문
Homology of the palpebral and origin of the supraorbital ossifications in ornithischian dinosaurs
2010
[6]
논문
Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: postcranial skeleton
http://dx.doi.org/10[...]
2019-12-17
[7]
논문
A New Specimen of the Thyreophoran Dinosaur cf. Scelidosaurus with Soft Tissue Preservation
2000
[8]
논문
Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: the dermal skeleton
https://academic.oup[...]
2020-08-20
[9]
서적
The Ultimate Dinosaur Book
Dorling Kindersley, New York
1993
[10]
간행물
Palaeontology
1859
[11]
논문
"Scelidosaurus harrisonii Owen, 1861 (Reptilia, Ornithischia): proposed replacement in inappropriate lectotype"
1992
[12]
간행물
A monograph of a fossil dinosaur (Scelidosaurus harrisonii, Owen) of the Lower Lias, part I. Monographs on the British fossil Reptilia from the Oolitic Formations 1
1861
[13]
논문
On the grammar of names formed with -scelus, -sceles, -scelis, etc.
1970
[14]
간행물
A monograph of the fossil Reptilia of the Liassic Formations. Part 2. A monograph of a fossil dinosaur (Scelidosaurus harrisonii Owen) of the Lower Lias
1863
[15]
논문
Professor Richard Owen and the important but neglected dinosaur Scelidosaurus harrisonii
2000
[16]
간행물
Catalogue of the Fossil Reptilia and Amphibia in the British Museum. Part 1. Containing the Orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia and Proterosauria
British Museum (Natural History)
1888
[17]
논문
The Jurassic dinosaur Scelidosaurus harrisoni, Owen
1968
[18]
간행물
"Scelidosaurus harrisonii Owen, 1861 (Reptilia, Ornithischia): lectotype replaced"
1994
[19]
서적
Jurassic Park: Unauthorized Jewish Fractals in Philopatry. A Fractal Scaling in Dinosaurology Project
Capitola, California
1995
[20]
논문
The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans
2010
[21]
논문
Dinosaurs of Dorset: Part III, the ornithischian dinosaurs (Dinosauria, Ornithischia) with additional comments on the sauropods
2011
[22]
논문
Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: cranial anatomy
2020
[23]
논문
The development of the remains of a small Scelidosaurus from a Lias nodule
1968
[24]
논문
The Mesozoic reptiles of Dorset: Part Two
1959
[25]
논문
New scelidosaur remains from the Lower Lias of Dorset
1989
[26]
논문
Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia
2007
[27]
논문
First dinosaur remains from Ireland
https://www.scienced[...]
2021
[28]
논문
A re-evaluation of Scelidosaurus remains from Ireland and the importance of apomorphy-based identifications
2024
[29]
간행물
The Thyreophoran Dinosaur Scelidosaurus from the Lower Jurassic Lufeng Formation, Yunnan, China
Museum of Northern Arizona Bulletin
1996
[30]
논문
Reconsidering the status and affinities of the ornithischian dinosaur Tatisaurus oehleri Simmons, 1965
2007
[31]
논문
Presence of the dinosaur Scelidosaurus indicates Jurassic age for the Kayenta Formation (Glen Canyon Group, northern Arizona)
1989-05
[32]
웹사이트
Evolution of the stegosaurs (Dinosauria; Ornithischia). Dinologia
http://dinoweb.narod[...]
2014
[33]
논문
The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen.
2016
[34]
간행물
Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society, New Series 14
1871
[35]
논문
On the Dinosauria of the Trias, with observations on the classification of the Dinosauria
1869
[36]
서적
Vertebrate Palaeontology
Blackwell Publishing
[37]
서적
Mesozoic Vertebrate Life
Indiana University Press
[38]
간행물
Relationships of the lower Jurassic dinosaur ''Scelidosaurus harrisonii''
1977-07
[39]
웹사이트
Ornithischia: Ankylosauromorpha
http://www.palaeos.c[...]
2007-02-11
[40]
서적
The Armored Dinosaurs
Indiana University Press
[41]
서적
The Dinosauria, 2nd Edition
University of Californian Press
[42]
서적
The Age of Reptiles
W. W. Norton & Company, Inc.
[43]
간행물
A comprehensive phylogenetic analysis on early ornithischian evolution
[44]
간행물
Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora)
[45]
간행물
Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships
https://academic.oup[...]
2021-01-01
[46]
간행물
Discovery of a thyreophoran trackway in the Hettangian of central France
[47]
서적
The Armored Dinosaurs
Indiana University Press
[48]
서적
The Dinosauria
University of California Press
[49]
간행물
The Oldest Jurassic Dinosaur: A Basal Neotheropod from the Hettangian of Great Britain
2016-04-20
[50]
간행물
Million-year-scale alternation of warm–humid and semi-arid periods as a mid-latitude climate mode in the Early Jurassic (late Sinemurian, Laurasian Seaway)
https://cp.copernicu[...]
2021-07-21
[51]
간행물
Use of palynofacies analysis to define Lower Jurassic (Sinemurian to Pliensbachian) genetic stratigraphic sequences in the Wessex Basin, England
1998
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com