호메오박스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
호메오박스는 초파리에서 처음 발견된 유전자이며, DNA 결합 도메인을 암호화하는 180개의 염기쌍 염기서열을 포함한다. 호메오박스는 3개의 알파 나선으로 구성된 60개의 아미노산 길이의 호메오도메인을 형성하며, 이 도메인은 DNA와 직접 상호작용하여 유전자 발현을 조절하는 전사 인자로 기능한다. 호메오박스 유전자는 신체 부위의 위치 변화를 유발하며, 진화 과정에서 다양한 종에서 발견된다.
초파리에서 머리에 더듬이 대신 다리가 자라는 변형(Antennapedia)을 연구하는 과정에서 호메오박스 유전자가 처음 발견되었다.[68] 1984년, 스위스 바젤 대학교의 에른스트 하펜, 마이클 레빈, 윌리엄 맥기니스, 발터 게링과 미국 인디애나 대학교의 매튜 스콧과 에이미 와이너는 독립적으로 초파리에서 ''antennapedia'' 호메오박스 서열을 포함하는 추가적인 유전자들을 발견했다고 보고했다.[70][71]
호메오도메인은 호메오박스 유전자가 코딩하는 단백질의 특정 부위로, DNA에 결합하는 기능을 한다. 단백질 폴드의 특징을 보이며, 3개의 알파 나선으로 구성된 60개의 아미노산 길이의 도메인이다. 다음은 합의된 호메오도메인(~60 아미노산 사슬)을 보여준다:[15]
혹스 유전자와 이와 관련된 마이크로 RNA는 조직 특이적이고 시공간적인 제어가 엄격하게 이루어지는 고도로 보존된 발생 마스터 조절 인자이다. 이 유전자들은 여러 암에서 조절 이상이 나타나는 것으로 알려져 있으며, 종종 DNA 메틸화에 의해 제어된다.[18][19] 혹스 유전자의 조절은 매우 복잡하며 상호 작용, 주로 억제를 포함한다. 초파리는 유충 발달 동안 발생하는 쌍 규칙 유전자와 간극 유전자의 하향 조절 이후 혹스 유전자의 발현을 유지하기 위해 폴리콤 및 트리쏘락스 복합체를 사용하는 것으로 알려져 있다. 폴리콤 단백질군은 크로마틴 구조 조절을 통해 혹스 유전자를 침묵시킬 수 있다.[20]
호메오박스 유전자의 돌연변이는 초파리의 안테나포디아(Antennapedia)나 바이토락스(Bithorax) 돌연변이 표현형처럼 신체 부위의 위치 변화를 유발하는 등 쉽게 눈에 띄는 표현형 변화를 생성할 수 있다. 예를 들어, 초파리의 특정 유전자에 결손이 생기면 머리의 촉각 위치에서 다리가 자라나는 현상이 나타날 수 있다.[60] 호메오박스 유전자의 복제는 새로운 신체 분절을 생성할 수 있으며, 이러한 복제는 분절된 동물의 진화에서 중요한 역할을 했을 가능성이 있다. 척추동물에서 Hox 유전자의 변이는 보통 출생 전 사망을 초래한다.
호메오박스 유전자 서열과 호메오도메인 단백질 구조에 대한 계통 발생 분석 결과, 식물, 균류, 동물의 마지막 공통 조상에는 최소 두 개의 호메오박스 유전자가 있었을 것으로 추정된다.[79] 분자 생물학적 증거에 따르면 일부 제한적인 수의 Hox 유전자가 최초의 진정한 좌우대칭동물 이전부터 자포동물에 존재해 왔으며, 이는 이러한 유전자가 고생대 이전부터 존재했음을 나타낸다.[80] 세 개의 주요 동물 ANTP-클래스 유전자 클러스터인 Hox, ParaHox, NK (MetaHox)는 분절 중복의 결과로 여겨진다. 첫 번째 중복은 MetaHox와 ProtoHox를 만들었고, 후자는 나중에 Hox와 ParaHox로 중복되었다. 클러스터 자체는 단일 ANTP-클래스 호메오박스 유전자의 직렬 중복에 의해 생성되었다.[81] 유전자 중복 후의 신기능화는 진핵생물에서 발견되는 많은 호메오박스 유전자의 원인이다. 호메오박스 유전자와 유전자 클러스터의 비교는 후생동물의 게놈 구조와 체형 진화를 이해하는 데 사용되어 왔다.[82]
호메오도메인을 포함하는 단백질은 크게 두 가지로 분류할 수 있다.[55] 게놈 내에서 특징적인 클러스터를 형성하는 Hox 유전자군에서 유래한 '''Hox''' 단백질과, Hox 이외의 게놈 내에 산재하는 non-Hox 유전자에서 유래하는 non-Hox 호메오 단백질이다.
2. 발견

이후, 에드워드 드 로베르티스와 윌리엄 맥기니스가 상동 유전자를 분리하여 다양한 종에서 호메오박스 유전자가 발견되었다.[72][73] 이러한 유전자들은 모든 양측 대칭 동물에 존재한다는 것이 밝혀졌다.
3. 호메오도메인
```
Helix 1 Helix 2 Helix 3/4
______________ __________ _________________
RRRKRTAYTRYQLLELEKEFHFNRYLTRRRRIELAHSLNLTERHIKIWFQNRRMKWKKEN
....|....|....|....|....|....|....|....|....|....|....|....|
10 20 30 40 50 60
```
호메오도메인 단백질은 진핵생물에서 발견된다.[4]
호메오도메인을 포함하는 단백질은 크게 두 가지로 분류된다.[55] Hox 유전자군에서 유래하여 게놈 내에서 특징적인 클러스터를 형성하는 '''Hox''' 단백질 (또는 Hox)과, Hox 이외의 게놈 내에 산재하는 non-Hox 유전자에서 유래하는 non-Hox 호메오 단백질이다. 포유류에서는 Hox 유전자가 서로 다른 염색체상에 4개의 클러스터를 형성하고 있으며, 염기 서열의 상동성으로부터 13개의 그룹으로 나뉘어 3' 측부터 번호가 매겨진다. 발생 과정에서는 이 순서에 대응하여 전후 축을 따라 발현하여, 그 위치에 특징적인 체절 구조를 유도하는데, 이는 초파리의 호메오틱 변이 해석을 통해 밝혀졌다.[56][57]
NK-2 패밀리나 MSX 패밀리와 같이 non-Hox 유전자 또한 다양한 발생 분화 과정과 관련된 진화적으로 보존된 패밀리를 형성하고 있다.[58][59] 출아 효모의 성 결정을 지배하는 MAT 유전자도 호메오박스를 가지고 있다.
컴퓨터 계산에 비유하면, 호메오박스 유전자는 서브루틴 호출과 유사하다고 생각할 수 있다. 이것은 DNA의 다른 위치에 이미 존재하고 있는 서브 시스템 전체의 생산을 전환한다.
3. 1. 구조
호메오도메인은 약 60개의 아미노산으로 구성되며, 3개의 알파 나선 구조를 가진다.[74] 이 중 2번과 3번 나선은 헬릭스-턴-헬릭스(HTH) 모티프를 형성하며, DNA의 주요 홈에 결합한다. N-말단 팔은 DNA의 작은 홈에 결합한다.[7]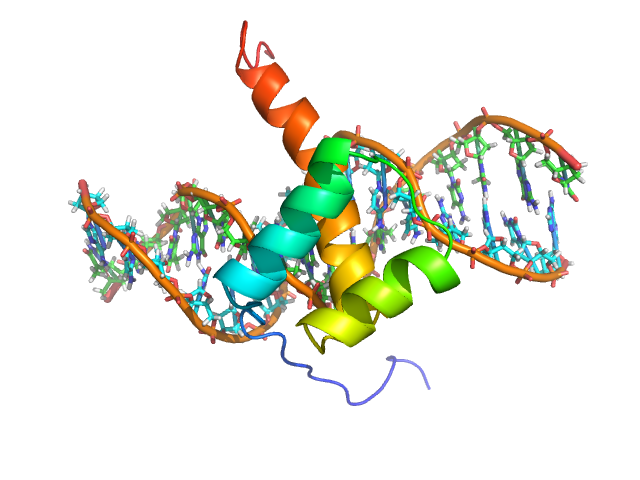
세 번째 나선은 DNA의 주요 홈 안에 있는 특정한 곁사슬과 노출된 염기 사이에 발생하는 물 분자를 통한 간접적인 상호작용뿐만 아니라 많은 수소 결합과 소수성 상호작용을 통해 DNA와 직접 상호작용한다.[7]
HTH 모티프를 통해, 호메오도메인 단백질은 원핵생물에서 유전자의 발현을 바꾸는 람다 파지 단백질과 같은 원핵생물 전사 인자와 제한적인 서열 유사성과 구조적 유사성을 공유한다.[16]
3. 2. 서열 특이성
호메오도메인은 5'-TAAT-3' 염기서열을 선호하여 DNA에 결합한다.[53][54] 아르기닌과 라이신 잔기가 풍부한 인식 나선과 나선간 루프는 DNA 골격에 수소 결합을 형성한다. 보존된 소수성 잔기는 인식 나선의 중심에서 나선 포장을 안정화하는 데 도움을 준다. 호메오도메인은 단독으로는 결합 특이성이 낮지만, 다른 전사 인자와 복합체를 형성하여 특정 유전자의 프로모터 영역을 인식하고 유전자 발현을 조절한다.[53][54]
3. 3. 생물학적 기능
호메오도메인 단백질은 보존된 HTH 모티프의 DNA 결합 특성으로 인해 전사 인자로 기능한다. 호메오도메인 단백질은 최종 제어 유전자로 간주되며, 이는 단일 단백질이 많은 표적 유전자의 발현을 조절할 수 있음을 의미한다.[75] 호메오도메인 단백질은 초기 배아 발달 동안 신체 축과 신체 구조의 형성을 지시한다. 많은 호메오도메인 단백질은 개별 조직과 기관을 생산하는데 필요한 공동 조절 유전자의 단계적 시작을 통해 세포 분화를 유도한다. NANOG와 같은 가족의 다른 단백질은 다능성을 유지하고 세포 분화를 방지하는 데 관여한다.
호메오박스는 대략 180개의 염기쌍을 가지며, DNA에 결합할 수 있는 단백질 부위(호메오도메인)를 코딩한다. 호메오박스 유전자는 예를 들어 다리를 만드는 데 필요한 모든 유전자 등, 전형적으로 다른 유전자의 캐스케이드를 스위치하는 전사 인자를 코딩한다.
Hox 유전자는 체축의 패턴 형성에 기능한다. 따라서 특이적인 몸의 부분의 동일성을 부여하며, Hox 유전자는 발생 중인 태아나 유생에서 사지나 다른 체절의 성장을 결정한다. 이들 유전자의 변이는 여분의 성장을 일으킬 수 있으며, 전형적으로 무척추동물에서 기능적이지 않은 몸의 부분, 예를 들어 초파리의 한 유전자의 결손으로 인해 aristapaedia 복합체가 머리의 촉각 위치에서 다리를 자라게 한다. 척추동물에서의 Hox 유전자의 변이는 통상, 출생 전에 사망한다.
호메오박스 유전자는 처음에 초파리에서 발견되었고, 이어서 곤충에서 파충류, 포유류에 이르기까지 많은 다른 종에서 동정되었다.[60] 호메오박스는 단세포 효모와 같은 균류나 식물에도 발견된다. 이 사실은 이 유전자 패밀리가 매우 일찍 진화하여, 형태 형성의 기본 메커니즘이 많은 생물에서 동일하다는 것을 보여준다.
호메오박스 유전자의 변이는 쉽게 눈에 보이는 표현형의 변화를 만들어낸다. 위에서 언급한 촉각 위치의 다리나, 두 번째 날개와 같은 초파리의 예가 그것이다. 호메오박스 유전자의 중복은 새로운 몸의 부분을 만들 수 있으며, 그러한 중복이 체절이 있는 동물의 진화에는 중요할 수 있다.
4. 조절
5. 돌연변이
6. 진화
7. 호메오박스 유전자의 종류
non-Hox 유전자에는 NK-2 패밀리나 MSX 패밀리 등이 있으며, 이들은 다양한 발생 분화 과정과 관련된 진화적으로 보존된 패밀리를 형성한다.[58][59] 출아 효모의 성 결정을 지배하는 MAT 유전자도 호메오박스를 가지고 있다.
Hox 유전자는 체축 패턴 형성에 기능하며, 발생 중인 태아나 유생에서 사지나 다른 체절의 성장을 결정한다.
호메오박스 유전자는 초파리에서 처음 발견되었고, 이후 곤충, 파충류, 포유류 등 다양한 종에서 동정되었다.[60] 쥐의 Pit-1 호메오박스 함유 단백질(보라색)이 DNA에 결합하는 구조 모델에서 볼 수 있듯이, Pit-1은 성장 호르몬 유전자 전사 조절 인자이며, POU 도메인과 호메오도메인 두 가지를 사용하여 DNA에 결합하는 POU DNA 결합 도메인 패밀리 중 하나이다. 호메오박스는 단세포 효모와 같은 균류나 식물에도 발견된다. 이는 이 유전자 패밀리가 매우 일찍 진화하여, 형태 형성의 기본 메커니즘이 많은 생물에서 동일하다는 것을 보여준다.
호메오박스 유전자의 변이는 쉽게 눈에 보이는 표현형 변화를 일으킨다.
컴퓨터 계산에 비유하면, 호메오박스 유전자는 서브루틴 호출과 유사하다고 생각할 수 있다. 이는 DNA의 다른 위치에 이미 존재하는 서브 시스템 전체의 생산을 전환한다.
인간의 Hox 유전자는 4개의 염색체 클러스터(HOXA, HOXB, HOXC, HOXD)로 구성되어 있다.이름 염색체 유전자 HOXA (또는 HOX1) 7번 염색체 HOXA1, HOXA2, HOXA3, HOXA4, HOXA5, HOXA6, HOXA7, HOXA9, HOXA10, HOXA11, HOXA13 HOXB 17번 염색체 HOXB1, HOXB2, HOXB3, HOXB4, HOXB5, HOXB6, HOXB7, HOXB8, HOXB9, HOXB13 HOXC 12번 염색체 HOXC4, HOXC5, HOXC6, HOXC8, HOXC9, HOXC10, HOXC11, HOXC12, HOXC13 HOXD 2번 염색체 HOXD1, HOXD3, HOXD4, HOXD8, HOXD9, HOXD10, HOXD11, HOXD12, HOXD13
ParaHox 유전자 (CDX1, CDX2, CDX4, GSX1, GSX2, PDX1)와 Hox 유사 유전자 (EVX1, EVX2, GBX1, GBX2, MEOX1, MEOX2, MNX1)도 유사하게 네 가지 영역에서 발견된다. 일부 "MetaHox"로 간주되는 NK-유사(NKL) 유전자는 Hox-유사 유전자와 함께 큰 ANTP-유사 그룹으로 묶인다.[106][107]
인간에게는 말단 없는(distal-less) 호메오박스 (DLX1, DLX2, DLX3, DLX4, DLX5, DLX6)가 있으며, 신경계와 팔다리 발달에 관여한다.[108] 이들은 NK-유사 유전자의 부분집합으로 간주된다.[106]
비정형 호메오도메인을 가진 인간 TALE(3개의 아미노산 루프 확장) 호메오박스 유전자는 63개의 아미노산으로 구성된다. 여기에는 IRX1, IRX2, IRX3, IRX4, IRX5, IRX6, MEIS1, MEIS2, MEIS3, MKX, PBX1, PBX2, PBX3, PBX4, PKNOX1, PKNOX2, TGIF1, TGIF2, TGIF2LX, TGIF2LY가 있다.[106]
인간은 다음과 같은 호메오박스 유전자와 단백질을 가지고 있다.[106]7. 1. Hox 유전자
Hox 유전자는 호메오박스 유전자의 가장 잘 알려진 하위 집합으로, 전후 축을 따라 배아 영역의 정체성을 결정하는 필수적인 다세포생물 유전자이다.[27] 1984년 에드워드 데 로베르티스와 그의 동료들이 제노푸스에서 최초의 척추동물 Hox 유전자를 분리했다.[28] 이 유전자 집합이 주목받는 이유는 게놈 내에서의 독특한 행동과 배열 때문이다. Hox 유전자는 일반적으로 조직화된 클러스터로 발견되며, 클러스터 내 Hox 유전자의 선형 순서는 발생 과정에서 시간적, 공간적 발현 순서와 직접적으로 연관된다. 이 현상을 공선성(colinearity)이라고 한다.
이러한 호메오 유전자에 돌연변이가 발생하면 배아 발달 과정에서 신체 분절의 위치가 변하는 이소성이 나타난다. 예를 들어, 유전자 하나가 손실되면 해당 분절이 더 앞쪽으로 발달하고, 기능 획득 돌연변이는 분절이 더 뒤쪽으로 발달하게 한다. 대표적인 예로 안테나피디아와 바이토락스가 있는데, 이는 각각 더듬이 대신 다리가 발달하거나 흉부가 중복 발달하는 현상을 유발할 수 있다.[29]
척추동물에는 4개의 상동 클러스터(HoxA, HoxB, HoxC, HoxD)가 존재하며, 이들은 기능적으로 부분적인 중복을 보이지만, 여러 파생된 기능도 획득했다. 예를 들어, HoxA와 HoxD는 사지 축을 따라 분절 정체성을 지정한다.[30][31] Hox 계열의 특정 구성원들은 기질 분해, 인테그린 및 ECM 구성 요소의 변화를 조율하여 혈관 재형성, 혈관 신생 및 질병에 관여한다.[32] HoxA5는 죽상경화증과 관련이 있다.[33][34] HoxD3 및 HoxB3는 EC에서 각각 b3 및 a5 인테그린과 Efna1을 상향 조절하는 침윤성 촉진 및 혈관 신생 유전자이다.[35][36][37][38] HoxA3는 MMP14 및 uPAR을 상향 조절하여 내피 세포(EC) 이동을 유도한다. 반대로, HoxD10 및 HoxA5는 TIMP1을 상향 조절/uPAR 및 MMP14를 하향 조절하고, Tsp2를 상향 조절/VEGFR2, Efna1, Hif1alpha 및 COX-2를 하향 조절함으로써 EC 이동과 혈관 신생을 억제하고 부착 연접을 안정화시키는 정반대 효과를 갖는다.[39][40] HoxA5는 또한 PTEN을 하향 조절하여 종양 억제 유전자 p53 및 Akt1을 상향 조절한다.[41] HoxA5의 억제는 혈관종 성장을 약화시키는 것으로 나타났다.[42] HoxA5는 유전자 발현에 광범위한 영향을 미쳐, 유방암 세포주에서 유도되면 약 300개의 유전자가 상향 조절된다.[42] HoxA5 단백질 전달 도메인 과발현은 HUVEC에 대한 TNFalpha 유도 단핵구 결합의 억제를 통해 나타나는 염증을 예방한다.[43][44]
포유류에서 Hox 유전자는 서로 다른 염색체 상에 4개의 클러스터를 형성하며, 염기 서열의 상동성에 따라 13개의 그룹으로 나뉘고 3' 말단부터 번호가 매겨진다. 발생 과정에서 이 순서에 따라 전후 축을 따라 발현하여 해당 위치에 특징적인 체절 구조를 유도한다. 이러한 사실은 초파리의 호메오틱 변이 해석을 통해 밝혀졌다.[56][57]
7. 2. LIM 유전자
LIM 유전자는 특징 도메인이 처음 확인된 3개의 단백질 이름 첫 글자를 따서 명명되었다. LIM 유전자는 2개의 아미노산, 시스테인, 히스티딘이 풍부한 LIM 도메인과 호메오도메인을 암호화한다.[100] LIM 도메인은 단백질-단백질 상호작용에서 기능하고 아연 분자에 결합할 수 있다. LIM 도메인 단백질은 세포질과 핵 모두에서 발견된다. 이들은 세포골격 재형성, 국소 접착 부위, 단백질 복합체의 지지체, 전사 인자로 기능한다.[100]
7. 3. Pax 유전자
대부분의 Pax 유전자는 호메오박스와 결합 특이성을 증가시키기 위해 DNA에 결합하는 짝을 이루는 도메인을 포함하지만, 일부 Pax 유전자는 호메오박스 서열의 전부 또는 일부를 잃었다.[101] Pax 유전자는 배아 분할, 신경계 발달, 전두엽 생성, 골격 발달, 얼굴 구조 형성에 기능한다. Pax 6은 안구 발달의 최종조절자이므로 유전자가 시신경 소포, 후속 안구 구조의 발달에 필요하다.[102]
7. 4. POU 유전자
POU 영역을 포함하는 단백질은 호메오도메인과 POU 도메인으로 구성되는데, POU 도메인은 두 개의 헬릭스-턴-헬릭스 모티프를 포함하며 DNA에 결합한다. 두 도메인은 DNA 나선을 감쌀 수 있을 만큼 충분히 긴 유연한 루프로 연결되어 있다. 이를 통해 두 도메인이 표적 DNA의 반대편에 결합하여 5'-ATGCAAAT-3' 서열을 가진 8개의 염기쌍 부분을 덮는다. POU 단백질의 개별 도메인은 DNA에 약하게 결합하지만, 연결되면 강한 서열 특이적 친화성을 가진다. POU 도메인 자체는 람다 파지와 같은 세균 파지에서 발현되는 억제인자와 구조적으로 유사하다.[53][54]
7. 5. 식물 호메오박스 유전자
동물에서와 같이 식물 호메오박스 유전자는 전형적인 60개 아미노산 길이의 DNA 결합 호메오도메인을 암호화하거나, TALE(3개의 아미노산 루프 확장) 호메오박스 유전자의 경우 63개 아미노산으로 구성된 비정형 호메오도메인을 암호화한다. 보존된 인트론-엑손 구조와 고유한 코도메인 아키텍처에 따라 HD-ZIP I ~ IV, BEL, KNOX, PLINC, WOX, PHD, DDT, NDX, LD, SAWADEE, PINTOX의 14가지 고유한 클래스로 그룹화되었다.[103] 코도메인의 보존은 TALE[104] / 비 TALE[105] 호메오도메인 단백질에 대한 일반적인 진핵생물 조상을 시사한다.
7. 6. 인간 호메오박스 유전자
인간의 Hox 유전자는 4개의 염색체 클러스터(HOXA, HOXB, HOXC, HOXD)로 구성되어 있다.
| 이름 | 염색체 | 유전자 |
|---|---|---|
| HOXA (또는 HOX1) | 7번 염색체 | HOXA1, HOXA2, HOXA3, HOXA4, HOXA5, HOXA6, HOXA7, HOXA9, HOXA10, HOXA11, HOXA13 |
| HOXB | 17번 염색체 | HOXB1, HOXB2, HOXB3, HOXB4, HOXB5, HOXB6, HOXB7, HOXB8, HOXB9, HOXB13 |
| HOXC | 12번 염색체 | HOXC4, HOXC5, HOXC6, HOXC8, HOXC9, HOXC10, HOXC11, HOXC12, HOXC13 |
| HOXD | 2번 염색체 | HOXD1, HOXD3, HOXD4, HOXD8, HOXD9, HOXD10, HOXD11, HOXD12, HOXD13 |
ParaHox 유전자도 유사하게 네 가지 영역에서 발견된다. 여기에는 CDX1, CDX2, CDX4, GSX1, GSX2, PDX1이 포함된다. Hox와 유사한 것으로 간주되는 다른 유전자에는 EVX1, EVX2, GBX1, GBX2, MEOX1, MEOX2, MNX1이 있다. 일부 "MetaHox"로 간주되는 NK-유사(NKL) 유전자는 Hox-유사 유전자와 함께 큰 ANTP-유사 그룹으로 묶인다.[106][107]
인간에게는 말단 없는(distal-less) 호메오박스 (DLX1, DLX2, DLX3, DLX4, DLX5, DLX6)가 있다. Dlx 유전자는 신경계와 팔다리의 발달에 관여한다.[108] 이들은 NK-유사 유전자의 부분집합으로 간주된다.[106]
비정형 호메오도메인을 가진 인간 TALE(3개의 아미노산 루프 확장) 호메오박스 유전자는 60개가 아닌 63개의 아미노산으로 구성된다. 여기에는 IRX1, IRX2, IRX3, IRX4, IRX5, IRX6, MEIS1, MEIS2, MEIS3, MKX, PBX1, PBX2, PBX3, PBX4, PKNOX1, PKNOX2, TGIF1, TGIF2, TGIF2LX, TGIF2LY가 있다.[106]
또한 인간은 다음과 같은 호메오박스 유전자와 단백질을 가지고 있다.[106]
- LIM-class: ISL1, ISL2, LHX1, LHX2, LHX3, LHX4, LHX5, LHX6, LHX8, LHX9, LMX1A, LMX1B
- POU-class: HDX, POU1F1, POU2F1, POU2F2, POU2F3, POU3F1, POU3F2, POU3F3, POU3F4, POU4F1, POU4F2, POU4F3, POU5F1, POU5F1P1, POU5F1P4, POU5F2, POU6F1, POU6F2
- CERS-class: LASS2, LASS3, LASS4, LASS5, LASS6
- HNF-class: HMBOX1, HNF1A, HNF1B
- SINE-class: SIX1, SIX2, SIX3, SIX4, SIX5, SIX6
- CUT-class: ONECUT1, ONECUT2, ONECUT3, CUX1, CUX2, SATB1, SATB2
- ZF-class: ADNP, ADNP2, TSHZ1, TSHZ2, TSHZ3, ZEB1, ZEB2, ZFHX2, ZFHX3, ZFHX4, ZHX1, HOMEZ
- PRD-class: ALX1 (CART1), ALX3, ALX4, ARGFX, ARX, DMBX1, DPRX, DRGX, DUXA, DUXB, DUX (1, 2, 3, 4, 4c, 5), ESX1, GSC, GSC2, HESX1, HOPX, ISX, LEUTX, MIXL1, NOBOX, OTP, OTX1, OTX2, CRX, PAX2, PAX3, PAX4, PAX5, PAX6, PAX7, PAX8, PHOX2A, PHOX2B, PITX1, PITX2, PITX3, PROP1, PRRX1, PRRX2, RAX, RAX2, RHOXF1, RHOXF2/2B, SEBOX, SHOX, SHOX2, TPRX1, UNCX, VSX1, VSX2
- NKL-class: BARHL1, BARHL2, BARX1, BARX2, BSX, DBX1, DBX2, EMX1, EMX2, EN1, EN2, HHEX, HLX1, LBX1, LBX2, MSX1, MSX2, NANOG, NOTO, TLX1, TLX2, TLX3, TSHZ1, TSHZ2, TSHZ3, VAX1, VAX2, VENTX
- * Nkx: NKX2-1, NKX2-4, NKX2-2, NKX2-8, NKX3-1, NKX3-2, NKX2-3, NKX2-5, NKX2-6, HMX1, HMX2, HMX3, NKX6-1, NKX6-2, NKX6-3
참조
[1]
논문
Determination of the nuclear magnetic resonance solution structure of an Antennapedia homeodomain-DNA complex
1993-12
[2]
논문
The homeobox in perspective
1992-08
[3]
논문
Exploring the homeobox
1993-12
[4]
논문
Homeodomain proteins: an update
2016-06
[5]
웹사이트
Homeoboxes
https://ghr.nlm.nih.[...]
U.S. National Library of Medicine
2019-11-20
[6]
서적
Materials for the study of variation, treated with especial regard to discontinuity in the origin of species
Macmillan
1894
[7]
논문
Patterns, puzzles and paradigms - The riddle of the homeobox
[8]
논문
The structure and function of the homeodomain
1989-07
[9]
논문
Genomic and cDNA clones of the homeotic locus Antennapedia in Drosophila
1983
[10]
웹사이트
Walter Jakob Gehring (1939-2014) {{!}} The Embryo Project Encyclopedia
https://embryo.asu.e[...]
2019-12-09
[11]
논문
A conserved DNA sequence in homoeotic genes of the Drosophila Antennapedia and bithorax complexes
1984
[12]
논문
Structural relationships among genes that control development: sequence homology between the Antennapedia, Ultrabithorax, and fushi tarazu loci of Drosophila
1984-07
[13]
논문
Cloning of an X. laevis gene expressed during early embryogenesis coding for a peptide region homologous to Drosophila homeotic genes
1984-06
[14]
논문
A homologous protein-coding sequence in Drosophila homeotic genes and its conservation in other metazoans
https://www.cell.com[...]
1984-06
[15]
웹사이트
The homeobox page
http://www.csb.ki.se[...]
Karolinksa Institute
2010-01-30
[16]
웹사이트
CATH Superfamily 1.10.10.60
http://www.cathdb.in[...]
2018-03-27
[17]
논문
Differential DNA binding properties of three human homeodomain proteins
1992-09
[18]
논문
Flow-Dependent Epigenetic DNA Methylation in Endothelial Gene Expression and Atherosclerosis
2015-07
[19]
논문
HOX genes and their role in the development of human cancers
2014-08
[20]
서적
RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity
http://www.horizonpr[...]
Caister Academic Press
2008-02-27
[21]
논문
Did homeodomain proteins duplicate before the origin of angiosperms, fungi, and metazoa?
1997-12
[22]
논문
Pre-bilaterian origins of the Hox cluster and the Hox code: evidence from the sea anemone, Nematostella vectensis
2007-01
[23]
논문
The genesis and evolution of homeobox gene clusters
2005-12
[24]
논문
A comprehensive classification and evolutionary analysis of plant homeobox genes
2009-12
[25]
논문
Evolution of homeobox genes
2013
[26]
논문
Evolution of Homeobox Gene Clusters in Animals: The Giga-Cluster and Primary vs. Secondary Clustering
2016
[27]
논문
Hox proteins: sculpting body parts by activating localized cell death
2002-11
[28]
논문
Cloning of an X. laevis gene expressed during early embryogenesis coding for a peptide region homologous to Drosophila homeotic genes
1984-06
[29]
논문
Redesigning the body plan of Drosophila by ectopic expression of the homoeotic gene Antennapedia
1987
[30]
논문
Hoxa-13 and Hoxd-13 play a crucial role in the patterning of the limb autopod
1996-10
[31]
논문
Hox genes in digit development and evolution
1999-04
[32]
논문
The role of homeobox genes in vascular remodeling and angiogenesis
2000-11
[33]
논문
Flow-Dependent Epigenetic DNA Methylation in Endothelial Gene Expression and Atherosclerosis
2015-07
[34]
논문
The role of epigenetics in the endothelial cell shear stress response and atherosclerosis
2015-10
[35]
논문
Induction of the angiogenic phenotype by Hox D3
1997-10
[36]
논문
The homeobox transcription factor Hox D3 promotes integrin alpha5beta1 expression and function during angiogenesis
2004-02
[37]
논문
Homeobox B3 promotes capillary morphogenesis and angiogenesis
2000-01
[38]
논문
Retroviral delivery of homeobox D3 gene induces cerebral angiogenesis in mice
2004-11
[39]
논문
Sustained expression of homeobox D10 inhibits angiogenesis
2002-12
[40]
논문
HOXA3 induces cell migration in endothelial and epithelial cells promoting angiogenesis and wound repair
2005-06
[41]
논문
A role for Hox A5 in regulating angiogenesis and vascular patterning
[42]
논문
HoxA5 stabilizes adherens junctions via increased Akt1
[43]
논문
Restoring transcription factor HoxA5 expression inhibits the growth of experimental hemangiomas in the brain
2009-06
[44]
논문
Identification of transcriptional targets of HOXA5
2005-05
[45]
논문
The LIM domain: from the cytoskeleton to the nucleus
2004-11
[46]
논문
Pax in development
https://www.cell.com[...]
2019-12-11
[47]
논문
Pax-6, a murine paired box gene, is expressed in the developing CNS
1991-12
[48]
논문
Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals
1997-11
[49]
논문
Homeodomain proteins belong to the ancestral molecular toolkit of eukaryotes
[50]
논문
MetaHox gene clusters
2000-12-15
[51]
논문
Dlx homeobox gene control of mammalian limb and craniofacial development
2006-07
[52]
논문
Classification and nomenclature of all human homeobox genes
2007-10
[53]
논문
The homeobox in perspective
https://www.ncbi.nlm[...]
1992-08
[54]
논문
Homeodomain proteins: an update
https://www.ncbi.nlm[...]
2016-06
[55]
논문
Hox proteins: sculpting body parts by activating localized cell death
https://www.ncbi.nlm[...]
2002-11-19
[56]
논문
Structural relationships among genes that control development: sequence homology between the Antennapedia, Ultrabithorax, and fushi tarazu loci of Drosophila
https://www.ncbi.nlm[...]
1984-07
[57]
논문
A conserved DNA sequence in homoeotic genes of the Drosophila Antennapedia and bithorax complexes
https://www.ncbi.nlm[...]
1984-03-29
[58]
논문
NKX2-1 activation by SMAD2 signaling after definitive endoderm differentiation in human embryonic stem cell
https://www.ncbi.nlm[...]
2013-05-01
[59]
논문
Isolation of a diverged homeobox gene, MOX1, from the BRCA1 region on 17q21 by solution hybrid capture
https://www.ncbi.nlm[...]
1994-08
[60]
논문
Pre-bilaterian origins of the Hox cluster and the Hox code: evidence from the sea anemone, Nematostella vectensis
https://www.ncbi.nlm[...]
2007-01-24
[61]
논문
Homeodomain proteins: an update
http://dx.doi.org/10[...]
2015-10-13
[62]
논문
The homeobox in perspective
http://dx.doi.org/10[...]
1992-08
[63]
논문
Exploring the homeobox
http://dx.doi.org/10[...]
1993-12
[64]
논문
Genetics Home Reference2004147Genetics Home Reference. Rockville, MD: US National Library of Medicine 2003, frequent updates. Last visited December 2003. Gratis URL: http://ghr.nlm.nih.gov
http://dx.doi.org/10[...]
2004-04
[65]
서적
Materials for the study of variation treated with especial regard to discontinuity in the origin of species. By William Bateson.
http://dx.doi.org/10[...]
Macmillan,
1894
[66]
논문
Patterns, puzzles and paradigms: the riddle of the homeobox
http://dx.doi.org/10[...]
1987-01
[67]
논문
The structure and function of the homeodomain
http://dx.doi.org/10[...]
1989-07-28
[68]
논문
Genomic and cDNA clones of the homeotic locus Antennapedia in Drosophila.
http://dx.doi.org/10[...]
1983-11
[69]
논문
Walter Jakob Gehring (1939–2014)
http://dx.doi.org/10[...]
2014-07
[70]
논문
A conserved DNA sequence in homoeotic genes of the Drosophila Antennapedia and bithorax complexes
http://dx.doi.org/10[...]
1984-03
[71]
저널
Structural relationships among genes that control development: sequence homology between the Antennapedia, Ultrabithorax, and fushi tarazu loci of Drosophila.
http://dx.doi.org/10[...]
1984-07-01
[72]
저널
Cloning of an X. laevis gene expressed during early embryogenesis coding for a peptide region homologous to Drosophila homeotic genes
http://dx.doi.org/10[...]
1984-06
[73]
저널
A homologous protein-coding sequence in drosophila homeotic genes and its conservation in other metazoans
http://dx.doi.org/10[...]
1984-06
[74]
서적
Homeobox
http://dx.doi.org/10[...]
Elsevier
2001
[75]
저널
Differential DNA binding properties of three human homeodomain proteins
http://dx.doi.org/10[...]
1992
[76]
저널
Flow-Dependent Epigenetic DNA Methylation in Endothelial Gene Expression and Atherosclerosis
http://dx.doi.org/10[...]
2015-07
[77]
저널
HOX genes and their role in the development of human cancers
http://dx.doi.org/10[...]
2014-07-05
[78]
서적
RNA and the regulation of gene expression : a hidden layer of complexity
https://www.worldcat[...]
Caister Academic Press
2008
[79]
저널
Did homeodomain proteins duplicate before the origin of angiosperms, fungi, and metazoa?
http://dx.doi.org/10[...]
1997-12-09
[80]
저널
Pre-Bilaterian Origins of the Hox Cluster and the Hox Code: Evidence from the Sea Anemone, Nematostella vectensis
http://dx.doi.org/10[...]
2007-01-24
[81]
저널
The genesis and evolution of homeobox gene clusters
http://dx.doi.org/10[...]
2005-11-10
[82]
간행물
Evolution of Homeobox Gene Clusters in Animals: The Giga-Cluster and Primary vs. Secondary Clustering
Frontiers in Ecology and Evolution
2016
[83]
저널
Hox Proteins: Sculpting Body Parts by Activating Localized Cell Death
http://dx.doi.org/10[...]
2002-11
[84]
저널
Cloning of an X. laevis gene expressed during early embryogenesis coding for a peptide region homologous to Drosophila homeotic genes
http://dx.doi.org/10[...]
1984-06
[85]
저널
Hoxa-13 and Hoxd-13 play a crucial role in the patterning of the limb autopod
http://dx.doi.org/10[...]
1996-10-01
[86]
저널
Hox genes in digit development and evolution
http://dx.doi.org/10[...]
1999-03-29
[87]
저널
The Role of Homeobox Genes in Vascular Remodeling and Angiogenesis
http://dx.doi.org/10[...]
2000-11-10
[88]
저널
Flow-Dependent Epigenetic DNA Methylation in Endothelial Gene Expression and Atherosclerosis
http://dx.doi.org/10[...]
2015-07
[89]
저널
The role of epigenetics in the endothelial cell shear stress response and atherosclerosis
http://dx.doi.org/10[...]
2015-10
[90]
저널
Induction of the Angiogenic Phenotype by Hox D3
http://dx.doi.org/10[...]
1997-10-06
[91]
저널
The Homeobox Transcription Factor Hox D3 Promotes Integrin α5β1 Expression and Function during Angiogenesis
http://dx.doi.org/10[...]
2004-02
[92]
저널
Homeobox B3 Promotes Capillary Morphogenesis and Angiogenesis
http://dx.doi.org/10[...]
2000-01-24
[93]
저널
Retroviral Delivery of Homeobox D3 Gene Induces Cerebral Angiogenesis in Mice
http://dx.doi.org/10[...]
2004-11
[94]
저널
Sustained Expression of Homeobox D10 Inhibits Angiogenesis
http://dx.doi.org/10[...]
2002-12
[95]
저널
HOXA3 induces cell migration in endothelial and epithelial cells promoting angiogenesis and wound repair
http://dx.doi.org/10[...]
2005-06-15
[96]
저널
A Role for Hox A5 in Regulating Angiogenesis and Vascular Patterning
http://dx.doi.org/10[...]
2005-12
[97]
저널
HoxA5 Stabilizes Adherens Junctions Via Increased Akt1
http://dx.doi.org/10[...]
2007-10
[98]
저널
Restoring Transcription Factor HoxA5 Expression Inhibits the Growth of Experimental Hemangiomas in the Brain
http://dx.doi.org/10[...]
2009-06
[99]
저널
Identification of Transcriptional Targets of HOXA5
http://dx.doi.org/10[...]
2005-05
[100]
저널
The LIM domain: from the cytoskeleton to the nucleus
http://dx.doi.org/10[...]
2004-11
[101]
저널
Pax in development
http://dx.doi.org/10[...]
1992-05
[102]
저널
Pax-6, a murine paired box gene, is expressed in the developing CNS
http://dx.doi.org/10[...]
1991-12-01
[103]
저널
A Comprehensive Classification and Evolutionary Analysis of Plant Homeobox Genes
http://dx.doi.org/10[...]
2009-09-04
[104]
저널
Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals
http://dx.doi.org/10[...]
1997-11-01
[105]
저널
Homeodomain proteins belong to the ancestral molecular toolkit of eukaryotes
http://dx.doi.org/10[...]
2007-05-09
[106]
저널
Classification and nomenclature of all human homeobox genes
http://dx.doi.org/10[...]
2007-10-26
[107]
저널
MetaHoxgene clusters
http://dx.doi.org/10[...]
2000-12-15
[108]
저널
Dlx homeobox gene control of mammalian limb and craniofacial development
http://dx.doi.org/10[...]
2006
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com