가지돌기
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
가지돌기는 신경 세포의 세포체에서 뻗어 나오는 세포질 과정으로, 다른 뉴런으로부터 신호를 수신하는 역할을 한다. 가지돌기는 짧고 가늘며 복잡하게 분지되어 표면적을 넓혀 많은 신호를 효과적으로 받을 수 있도록 한다. 가지돌기 표면에는 시냅스가 존재하며, 흥분성 시냅스는 가지돌기 가시 위에 위치하고 억제성 시냅스는 가지돌기 축과 직접 접촉한다. 가지돌기는 뉴런의 입력 통합 방식에 영향을 미치며, 수동 케이블 이론을 통해 가지돌기 형태 변화가 세포체의 막 전압에 미치는 영향을 추적할 수 있다. 가지돌기 발달은 감각 입력, 환경 오염 물질 등 다양한 요인의 영향을 받으며, 시냅스 영양 가설이 가지돌기 발달 기작 중 하나로 제시되었다. 가지돌기는 전기적 특성을 가지며, 가소성을 통해 변화할 수 있다. "가지돌기"라는 용어는 빌헬름 히스에 의해 처음 사용되었으며, 오토 프리드리히 카를 다이터스가 축삭과 구별하여 발견한 것으로 알려져 있다.
더 읽어볼만한 페이지
- 신경조직학 - 미엘린
미엘린은 신경 세포 축삭을 둘러싸 절연 작용을 하여 전기적 신호의 빠른 전달을 돕는 지질 및 단백질 구조로, 중추신경계와 말초신경계에 존재하며 도약 전도를 통해 신호 전달 속도를 높이는 역할을 한다. - 신경조직학 - 세포체
- 신경계 - 자율신경계
자율신경계는 생명 유지에 필수적인 불수의적 기능을 조절하는 신경계로, 교감신경계와 부교감신경계로 나뉘어 길항적으로 작용하며, 장신경계와 신경전달물질을 통해 신체 기관에 영향을 미치고 기능 장애는 질환을 유발할 수 있다. - 신경계 - 교감신경계
교감신경계는 자율신경계의 일부로서 흉수와 요수에서 기원하여 아세틸콜린과 노르에피네프린을 신경전달물질로 사용하여 심박수 증가, 혈압 상승 등의 투쟁-도피 반응을 유발하며 다양한 질병과 관련되어 추가 연구가 필요한 시스템이다. - 조직학 - 광수용체
광수용체는 망막의 빛을 감지하는 세포로, 간상세포, 원추세포, ipRGCs로 구성되어 명암, 색깔, 생체 리듬 조절 등 시각 정보를 뇌로 전달하며, 이 과정의 이상은 시각 질환을 유발할 수 있다. - 조직학 - 근육 조직
근육 조직은 골격근, 민무늬근, 심장근으로 구분되며, 특히 골격근은 뼈, 인대, 힘줄과 함께 길항 작용을 통해 움직인다.
| 가지돌기 | |
|---|---|
| 개요 | |
| 정의 | 신경 세포에서 다른 신경 세포로부터 신호를 받는 짧은 돌기 |
| 기능 | 시냅스를 통해 다른 신경 세포에서 오는 신호를 수신하여 세포체로 전달 |
| 구조 | 세포체에서 뻗어 나온 가지 모양의 돌기. 표면에 시냅스가 존재하며, 수상돌기 가시를 가질 수 있음 |
| 상세 특징 | |
| 수신 역할 | 다른 신경 세포의 축삭에서 전달된 신호를 수신 |
| 신호 전달 | 수신된 신호를 세포체로 전달하여 통합 및 처리 |
| 시냅스 가소성 | 경험과 학습에 따라 시냅스의 강도가 변화하는 가소성에 중요한 역할 |
| 가지 형태의 다양성 | 신경 세포 종류에 따라 다양한 형태와 구조를 가지며, 이는 신경 회로의 복잡성을 증가시킴 |
| 관련 용어 | |
| 관련 용어 | 신경 세포 축삭 시냅스 세포체 신경 전달 물질 활동 전위 수상돌기 가시 |
2. 구조 및 기능
가지돌기(Dendrite)는 신경 세포의 세포체에서 뻗어 나오는 두 가지 유형의 세포질 과정 중 하나이며, 다른 하나는 축삭이다. 축삭은 모양, 길이, 기능 등에서 가지돌기와 구별된다. 가지돌기는 보통 가늘고 짧지만, 축삭은 일정한 굵기를 유지하며 매우 길 수 있다. 일반적으로 축삭은 전기화학적 신호를 전달하고 가지돌기는 신호를 받지만, 특정 신경 세포는 축삭 없이 가지돌기로 신호를 전달하기도 한다. 가지돌기는 넓은 표면적을 통해 다른 신경 세포의 축삭 종말로부터 신호를 받는다.[2] 예를 들어 큰 피라미드 세포의 가지돌기는 약 30,000개의 시냅스 이전 뉴런으로부터 신호를 받는다.[3]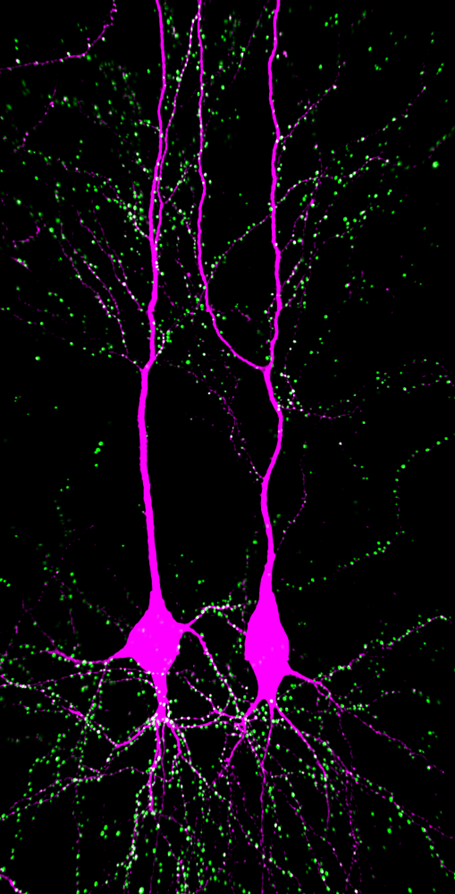
흥분성 시냅스는 가지돌기 가시에 연결되는데, 이는 신경 전달 물질 수용체가 많이 분포하는 가지돌기의 작은 돌출부이다. 대부분의 억제성 시냅스는 가지돌기 축과 직접 연결된다. 시냅스 활동은 가지돌기 세포막 전위의 국소적 변화를 일으킨다. 이 변화는 가지돌기를 따라 수동적으로 퍼지지만, 활동 전위 없이는 약해진다. 활동 전위를 생성하려면 많은 흥분성 시냅스가 동시에 활성화되어 가지돌기와 세포체(체)를 강하게 탈분극시켜야 한다. 축삭 언덕에서 시작되는 활동 전위는 축삭을 따라 축삭 종말로 전달되어 신경 전달 물질을 방출하지만, 가지돌기 안으로 역행 전파되어 스파이크 타이밍 의존 가소성 (STDP)에 중요한 신호를 제공한다.[2]
대부분의 시냅스는 축삭이 가지돌기에 신호를 보내는 축삭-가지돌기 시냅스이다. 가지돌기끼리 신호를 주고받는 가지돌기-가지돌기 시냅스도 있고,[4] 자기 시냅스는 한 뉴런의 축삭이 자신의 가지돌기에 신호를 보내는 시냅스이다.
가지돌기는 광범위하게 분지될 수 있으며, 단일 뉴런이 최대 100,000개의 입력을 받기도 한다.[2] 다른 신경 세포와의 시냅스가 많은 가지돌기는 시냅스에서 받은 정보를 세포체 내에서 활동 전위로 변환하여 전기 신호로 만든다. 이 신호가 축삭 말단에 도달하면 글루탐산 등의 신경 전달 물질이 방출되고, 다음 신경 세포가 이를 수용하여 정보가 전달된다.
소뇌의 푸르키녜 세포처럼 특징적인 형태의 가지돌기를 가진 신경 세포도 있다.
2. 1. 가지돌기의 유형
신경 세포는 가지돌기의 형태에 따라 다극 뉴런, 쌍극 뉴런, 단극 뉴런 등으로 분류될 수 있다.[4][5][6] 다극 뉴런은 하나의 축삭과 많은 가지돌기 가지로 구성된다. 피라미드 세포는 피라미드 모양의 세포체와 피질 표면을 향해 뻗어 있는 큰 가지돌기(정단 가지돌기)를 가진 다극 피질 뉴런이다. 쌍극 뉴런은 세포체의 반대쪽 끝에 두 개의 주요 가지돌기를 가지고 있다. 많은 억제성 뉴런이 이러한 형태를 가지고 있다. 곤충에서 전형적인 단극 뉴런은 세포체에서 뻗어 나와 두 개의 가지로 분리되는 줄기를 가지고 있으며, 하나는 가지돌기를 포함하고 다른 하나는 종말 단추를 포함한다. 척추동물에서 촉각 또는 온도를 감지하는 감각 뉴런은 단극성이다.가지돌기 분지는 '무수지(adendritic)', '방추형', '구형', '층상', '원통형', '원추형', '부채형' 등 다양한 패턴을 보일 수 있다. 각 패턴에 대한 설명은 다음과 같다.
| 형태 | 설명 |
|---|---|
| 무수지 | 분지 구조가 없거나 나무와 같지 않음. |
| 방추형 | 두 개의 가지돌기가 세포체의 반대쪽 극에서 몇 개의 가지와 함께 방사되는 경우 (양극성 뉴런 참조). |
| 구형 | 가지돌기가 부분적으로 또는 모든 방향으로 세포체에서 방사되는 경우 (소뇌 과립 세포 참조). |
| 층상 | 가지돌기가 평면적으로 방사되거나 하나 이상의 줄기에 의해 세포체에서 오프셋되거나 다중 평면적으로 방사될 수 있음 ( 망막 수평 세포, 망막 신경절 세포, 망막 아마크린 세포 참조). |
| 원통형 | 가지돌기가 원통형, 디스크와 같은 방식으로 모든 방향으로 방사되는 경우 (담창구 뉴런 참조). |
| 원추형 | 가지돌기가 세포체에서 원뿔처럼 방사되는 경우 (피질 층상 세포 참조). |
| 부채형 | 가지돌기가 푸르키네 세포처럼 평평한 부채 모양으로 방사되는 경우. |
뉴런의 가지돌기 구조, 분지 패턴, 전압 개폐 이온 통로 분포는 뉴런이 다른 뉴런으로부터 입력을 통합하는 방식에 큰 영향을 미친다.[14] 가지돌기는 한때 전기 신호를 수동적으로 전달하는 것으로만 여겨졌으나,[15][16] 최근 연구에 따르면 가지돌기에도 전압 개폐 이온 통로가 존재하며, 활동 전위의 역행 전파와 같은 능동적인 신호 전달 과정에도 관여한다.[17][18] 가지돌기 역행 전파는 스파이크 타이밍 의존 가소성에 대한 중요한 신호를 제공하며, 시냅스 조절 및 장기 강화에 중요한 역할을 한다.[2] 일부 뉴런에서는 가지돌기 개시 구역에서 칼슘 활동 전위 (가지돌기 스파이크)를 유발할 수도 있다.[17][18]
가지돌기 자체는 무척추동물을 포함한 동물의 성체 생애 동안 가소적 변화가 가능한 것으로 보인다.[19] 신경 가지돌기는 들어오는 자극을 계산할 수 있는 기능적 단위로 알려진 다양한 구획을 가지고 있다. 이러한 기능적 단위는 입력 처리에 관여하며, 가시, 가지 또는 가지의 그룹과 같은 가지돌기의 하위 도메인으로 구성된다. 따라서 가지돌기 구조의 변화를 유발하는 가소성은 세포의 통신 및 처리에 영향을 미친다. 발달 과정에서 가지돌기 형태는 세포의 유전체 내 고유 프로그램과 다른 세포의 신호와 같은 외적 요인에 의해 형성된다. 그러나 성체 생애에서는 외적 신호가 더 큰 영향을 미치며, 발달 과정의 내적 신호에 비해 가지돌기 구조에 더 큰 변화를 유발한다. 암컷의 경우, 가지돌기 구조는 임신, 수유, 발정 주기와 같은 기간 동안 호르몬에 의해 유도된 생리적 조건의 결과로 변경될 수 있다. 이는 해마의 CA1 영역의 피라미드 세포에서 특히 두드러지는데, 여기서 가지돌기의 밀도는 최대 30%까지 변화할 수 있다.
"가지돌기"라는 용어는 1889년 빌헬름 히스가 처음 사용했으며, 이는 신경 세포에 부착된 작은 "원형질 돌기"의 수를 설명하기 위해 사용되었다.[7] 독일 해부학자 오토 프리드리히 카를 다이터스는 일반적으로 축삭을 가지돌기와 구별하여 발견한 것으로 알려져 있다.
[1]
논문
Freeze-frame imaging of synaptic activity using SynTagMA
2020-05-18
3. 가지돌기 발달
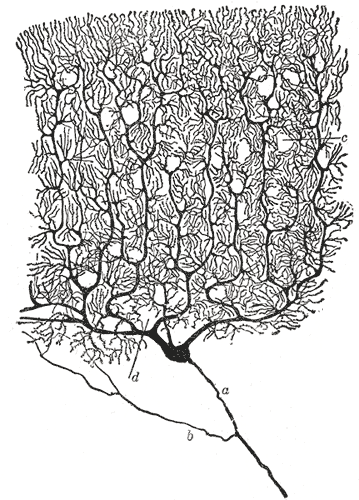
푸르키녜 세포는 부채꼴 모양의 가지돌기를 갖는다
가지돌기 발달은 감각 입력, 환경 오염 물질, 체온, 약물 사용 등 여러 요인의 영향을 받는다.[10] 어두운 곳에서 자란 쥐는 시각 피질의 피라미드 세포에서 스파인(spine) 수가 줄고, 가지돌기 분지 분포가 변하는 것이 관찰되었다.[11]
가지돌기 수지상 돌기 발달 기작 중 하나로 시냅스 영양 가설이 있다. 이 가설은 시냅스 전 뉴런에서 시냅스 후 뉴런으로의 입력, 특히 흥분성 시냅스 입력의 성숙이 가지돌기 및 축삭 수지상 돌기에서의 시냅스 형성을 조절한다고 설명한다.[12]
가지돌기 발달에는 복잡한 내/외부 신호가 관여하는데, 여기에는 전사 인자, 수용체-리간드 상호 작용, 신호 전달 경로, 국소 번역 기계, 세포 골격 요소 등이 포함된다. β-액틴 집코드 결합 단백질 1(ZBP1)은 가지돌기 분지에 기여하며, CUT, Abrupt, Collier, Spineless, ACJ6/drifter, CREST, NEUROD1, CREB, NEUROG2 등도 중요한 전사 인자이다. 분비 단백질 및 세포 표면 수용체에는 신경 영양 인자 및 티로신 키나아제 수용체, BMP7, Wnt/dishevelled, EPHB 1–3, Semaphorin/plexin-neuropilin, slit-robo, netrin-frazzled, reelin이 포함된다. Rac, CDC42 및 RhoA는 세포 골격 조절자 역할을 하며, 운동 단백질에는 KIF5, 다이네인, LIS1이 포함된다. 물질 P는 소뇌 푸르키녜 세포에서 가지돌기 수지상 돌기 형성을 유도한다.[13]
최근 연구에 따르면, 소뇌 푸르키녜 세포의 가지돌기 발달에는 과립 세포에서 발현되는 이노시톨 삼인산 (IP3) 수용체가 중요한 역할을 한다. 과립 세포의 IP3 수용체가 활성화되면 칼슘 방출 및 BDNF (뇌 유래 신경 영양 인자) 생성이 촉진되고, BDNF는 과립 세포의 축삭 말단에 작용하여 글루탐산 방출을 유도하며, 이 신호를 받은 푸르키녜 세포가 가지돌기를 형성한다. 푸르키녜 세포에도 IP3 수용체가 있지만, 자신의 수용체가 아닌 과립 세포의 IP3 수용체에 의해 제어된다.[23]
4. 전기적 특성
5. 가소성
최근의 실험적 관찰에 따르면 적응은 신경 가지돌기 트리에서 수행되며, 적응 시간 척도는 몇 초까지 짧게 관찰되었다.[20][21]
6. 역사
신경계에서 이루어진 최초의 세포 내 기록 중 일부는 1930년대 후반 케네스 S. 콜과 하워드 J. 커티스에 의해 이루어졌다. 스위스 출신 뤼돌프 알베르트 폰 쾰리커와 독일 출신 로베르트 레마크는 축삭 초기 분절을 처음으로 식별하고 특징을 규명했다. 앨런 호지킨과 앤드루 헉슬리는 오징어 거대 축삭을 사용했으며(1939년), 1952년까지 활동 전위의 이온적 기초에 대한 완전한 정량적 설명을 얻어 호지킨-헉슬리 모델을 공식화하게 되었다. 호지킨과 헉슬리는 이 연구로 1963년 공동으로 노벨상을 수상했다. 축삭 전도도를 상세히 설명하는 공식은 프랑켄호이저-헉슬리 방정식에서 척추동물로 확장되었다. 루이-앙투안 랑비에는 축삭에서 발견되는 간극 또는 마디를 처음으로 설명했으며, 이 기여로 인해 이러한 축삭 특징은 현재 랑비에 결절이라고 불린다. 스페인 해부학자 산티아고 라몬 이 카할은 축삭이 뉴런의 출력 구성 요소라고 제안했다.[8] 그는 또한 뉴런이 세포 간의 특수 접합부 또는 공간(현재는 시냅스라고 함)을 통해 서로 통신하는 개별 세포라고 제안했다. 라몬 이 카할은 그의 경쟁자였던 카밀로 골지가 개발한 골지 염색법으로 알려진 은 염색 과정을 개선했다.[9]
참조
[2]
서적
Essential Cell Biology
Garland Science
2009
[3]
논문
Human Cortical Pyramidal Neurons: From Spines to Spikes via Models
2018-06-29
[4]
서적
Physiology of Behavior
Pearson
2013
[5]
서적
Biopsychology
Allyn & Bacon
2011
[6]
논문
Branching out: mechanisms of dendritic arborization
2010-05
[7]
서적
Origins of neuroscience : a history of explorations into brain function
Oxford University Press
[8]
논문
Axon physiology
https://hal-amu.arch[...]
2011-04
[9]
논문
Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal
2006-10
[10]
논문
Stress, sex, and neural adaptation to a changing environment: mechanisms of neuronal remodeling
2010-09
[11]
논문
The effects of dark rearing on the development of the visual cortex of the rat
1978-07
[12]
논문
The regulation of dendritic arbor development and plasticity by glutamatergic synaptic input: a review of the synaptotrophic hypothesis
2008-03
[13]
논문
Intraventricular Administration of Substance P Increases the Dendritic Arborisation and the Synaptic Surfaces of Purkinje Cells in Rat's Cerebellum
https://www.tandfonl[...]
1999
[14]
서적
Principles of neural science.
https://archive.org/[...]
McGraw Hill
2003
[15]
서적
Biophysics of computation : information processing in single neurons
Oxford Univ. Press
1999
[16]
서적
Dendrites
Oxford University Press
2008
[17]
논문
Dendritic action potentials and computation in human layer 2/3 cortical neurons
2020-01-03
[18]
논문
The Guide to Dendritic Spikes of the Mammalian Cortex In Vitro and In Vivo
2022
[19]
논문
Synaptic Plasticity: A Unified Model to Address Some Persisting Questions
https://www.tandfonl[...]
2011
[20]
논문
Efficient dendritic learning as an alternative to synaptic plasticity hypothesis
2022-04
[21]
논문
Adaptive nodes enrich nonlinear cooperative learning beyond traditional adaptation by links
2018-03
[22]
논문
Learning on tree architectures outperforms a convolutional feedforward network
2023-01
[23]
논문
Inositol 1,4,5-Trisphosphate Receptor Type 1 in Granule Cells, Not in Purkinje Cells, Regulates the Dendritic Morphology of Purkinje Cells through Brain-Derived Neurotrophic Factor Production
2006
[24]
서적
LIFE
Mc Graw Hill
2010-10-04
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com