시클리드
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
시클리드는 전 세계에 분포하는 척추동물 과 중 하나로, 아프리카와 남아메리카에서 다양한 종이 서식한다. 육식, 초식, 잡식 등 다양한 먹이 섭취 방식을 보이며, 공격적인 행동과 독특한 번식 방식을 갖는다. 시클리드는 관상어로 인기가 높으며, 식용 및 낚시 대상어로도 활용된다. 일부 종은 잡종화가 용이하며, 품종 개량을 통해 다양한 관상어 품종이 개발되었다.
더 읽어볼만한 페이지
- 키클라목 - 폴리디크티스속
폴리디크티스속은 필리핀 남부와 솔로몬 제도 사이 열대 태평양 일부 지역에 서식하는 어류 속으로, 폴리디크티스 앵구이스와 폴리디크티스 루코테니아 두 종이 있으며, 폴리디크티스 루코테니아는 1856년에 명명되었고, 오발렌타리아류에 속하며 계통 분류 연구가 진행 중이다. - 동물 향명 - 맹금류
맹금류는 날카로운 발톱과 부리를 가진 사나운 새를 통칭하며, 생태계 최상위 포식자로서 뛰어난 시력과 비행 능력을 갖추고 있으며, 수리, 매, 독수리, 올빼미 등이 대표적이고, 서식지 파괴 등의 위협에 직면해 있다. - 동물 향명 - 돌고래
돌고래는 이빨고래류에 속하는 해양 포유류의 총칭으로, 다양한 종이 존재하며 높은 지능과 사회성을 가지지만, 서식지 파괴, 환경 오염 등으로 멸종 위기에 처해 있다. - 샤를 뤼시앵 보나파르트가 명명한 분류군 - 판새류
판새류는 상어, 가오리, 홍어 등을 포함하는 연골어류 아강으로, 아가미구멍과 여러 열의 이빨을 가지며 기름으로 가득 찬 간으로 부력을 조절하고, 턱 구조의 다양성을 보이는 특징을 지니며, 데본기에 출현하여 전 세계 해역과 일부 담수에 분포하지만 멸종 위기에 처해 보존 노력이 필요하다. - 샤를 뤼시앵 보나파르트가 명명한 분류군 - 공기호흡메기과
공기호흡메기과는 아프리카, 인도, 동남아시아 등지에 서식하며, 공기 호흡을 가능하게 하는 상아가미 기관을 통해 육지에서도 이동할 수 있는 메기목 조기어류의 한 과이다.
| 시클리드 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Cichlidae |
| 명명자 | Bonaparte, 1835 |
| 로마자 표기 | Sikeullideu |
| 분류 | |
| 상위 분류 | 신 기아강 극기상목 스즈키목 |
| 하위 분류 | 키클리다이아과 키클라소마티나아과 에트로플리아과 게오파기나아과 헤테로크로미디나아과 프세우도크레닐라브리나아과 프티코크로미나아과 |
| 아과별 분류 (대안적 분류) | |
| 키클리나아과 | 아스트로노티니족 카에토브란치니족 키클라소마티니족 키클리니족 게오파기니족 헤로이니족 레트로쿨리니족 |
| 에트로플리아과 | 바티바티니족 벤토크로미니족 볼렌게로크로미니족 크로미도틸라피니족 코엘로틸라피니족 콥토도니니족 시프리크로미니족 시포틸라피니족 엑토디니족 에레트모디니족 에티니족 에트로플리니족 고비오키클리니족 그린우도크로미니족 합플로크로미니족 헤미크로미니족 헤테로크로미디니족 헤테로틸라피니족 람프로로기니족 림노크로미니족 오레오크로미니족 펠마토크로미니족 펠마토라피니족 페리소디니족 프티코크로미니족 스테아토크라니니족 틸라피니족 트로페이니족 틸로크로미니족 |
| 생물학적 정보 | |
| 특징 | 인두골을 가짐 |
| 진화 및 분포 | |
| 화석 기록 | 초기 에오세 - 현재 |
| 최초 출현 | 4860만년 전 |
| 분포 | 아프리카 중앙아메리카 남아메리카 인도 |
| 기타 | |
| 참고 | 속은 아래에 있음 |
| 관련 항목 | |
| 문서 참고 | 카와스즈메과 |
2. 분포와 서식지
중앙아메리카에서 남아메리카에 걸쳐 분포하며, 마다가스카르를 포함한 아프리카, 중동, 남아시아까지 서식한다. 담수어와 기수어를 포함하여 최소 1,300종 이상이 확인되었다. 열대어 중에서는 비교적 사나운 편으로 알려져 있어 다른 종과 함께 기를 때는 주의가 필요하다.
시클리드과는 세계에서 가장 큰 척추동물 과 중 하나로, 특히 아프리카와 남아메리카에서 매우 다양한 종들이 서식한다. 아프리카 대륙에만 약 1,600종 이상이 살고 있는 것으로 추정된다.[19] 중앙아메리카와 멕시코에는 약 120종이 분포하며, 북쪽으로는 미국 남부 텍사스의 리오그란데강까지 서식지가 이어진다. 마다가스카르에는 아프리카 본토의 시클리드와는 유전적으로 거리가 있는 고유한 속들(''Katria'', ''Oxylapia'', ''Paratilapia'', ''Paretroplus'', ''Ptychochromis'', ''Ptychochromoides'')이 서식한다.[16][24]
아시아에는 시클리드과 어류가 상대적으로 적다. 이스라엘, 레바논, 시리아 등 중동 지역에 9종(''Astatotilapia flaviijosephi'', ''Oreochromis aureus'', ''O. niloticus'', ''Sarotherodon galilaeus'', ''Coptodon zillii'', 그리고 ''Tristramella'' 속의 종들), 이란에 2종(''Iranocichla''), 그리고 인도와 스리랑카에 3종(''Etroplus''와 ''Pseudetroplus'')만이 자연적으로 분포한다.[19] 카리브해의 앤틸리스 제도에서는 쿠바와 히스파니올라에 서식하는 Nandopsis 속의 3종만이 유일한 시클리드이다(트리니다드 토바고의 토착 시클리드는 남아메리카 본토에 널리 분포하는 속에 속하므로 제외).[19]
유럽, 오스트레일리아, 남극, 그리고 리오그란데강 유역 북쪽의 북아메리카에는 토착 시클리드가 없다. 하지만 미국의 플로리다, 하와이, 그리고 일본, 오스트레일리아 북부 등 여러 지역에서는 외래종으로 유입된 시클리드가 야생에서 성공적으로 정착하여 개체군을 형성하기도 했다.[23][25][26][27][28][29][30]
대부분의 시클리드는 비교적 얕은 물에서 서식하지만, 일부 예외적인 종들도 존재한다. 가장 깊은 곳에서 발견된 시클리드는 Trematocara 속의 종들로, 탕가니카 호수의 수심 300m 이상 깊은 곳에서도 발견되었다.[31] 말라위 호수에서는 Alticorpus macrocleithrum와 Pallidochromis tokolosh 같은 종들이 수면 아래 150m 깊이까지 내려가 서식하며,[32][33] 콩고 강에서는 색소가 없어 몸이 희고 눈이 보이지 않는 Lamprologus lethops가 수면 아래 160m 깊이까지 사는 것으로 알려져 있다.[34]
시클리드는 주로 담수 환경에 서식하지만, 많은 종들이 기수 환경에서도 상당 기간 생존할 수 있다. 예를 들어, 멕시코 시클리드(Mayaheros urophthalmus)는 담수 습지나 맹그로브 늪뿐만 아니라, 기수 환경인 석호나 해안가의 맹그로브 지대에서도 서식하며 번식한다.[14] 틸라피아(Tilapia), Sarotherodon, Oreochromis 속의 여러 종들은 염분에 대한 내성(광염성)이 강해 강 하구 사이의 기수 해안선을 따라 이동할 수 있다.[19] 하지만 주로 기수나 해수 환경에서 서식하는 시클리드는 소수에 불과하며, 대표적인 예로는 인도와 스리랑카의 오렌지 크로마이드(Etroplus maculatus)와 그린 크로마이드(Etroplus suratensis), 그리고 아프리카의 검은턱 틸라피아(Sarotherodon melanotheron) 등이 있다.[35][40][41] 시클리드가 서식하는 가장 극한 환경 중 하나는 아프리카의 따뜻하고 염도가 매우 높은 호수들로, Alcolapia 속과 Danakilia 속의 종들이 이러한 환경에 적응하여 살아가고 있다. 예를 들어, 에리트레아의 아바에데드 호수(Lake Abaeded)는 Danakilia dinicolai의 유일한 서식지로, 수온이 29°C에서 45°C까지 오르내리는 환경이다.[36]
쿠바, 히스파니올라, 마다가스카르의 종들을 제외하면, 시클리드는 대양 섬에서는 발견되지 않는다. 이러한 분포 패턴은 곤드와나 대륙의 분열과 관련이 있는 것으로 해석되기도 한다. 즉, 아프리카-남아메리카, 인도-마다가스카르의 시클리드들이 지리적으로 가까운 관계를 보이는 것은 대륙 이동의 결과라는 것이다.[37] 반면, 일부 학자들은 시클리드가 바다를 건너 분산했을 가능성을 제기하지만, 이 경우 왜 다른 섬에는 정착하지 못했는지 설명하기 어렵다는 문제점이 있다. 마다가스카르의 시클리드 대부분은 담수에만 서식하지만, ''Ptychochromis grandidieri''와 ''Paretroplus polyactis''는 해안 기수역에서도 흔히 발견되어 염분에 대한 내성이 있는 것으로 보인다.[38][39]
3. 생태
틸라피아와 같은 일부 종은 수산업에서 중요한 위치를 차지하며, 엔젤피시나 디스커스처럼 아름다운 색채를 가진 종들은 관상용 열대어로 인기가 높다.
시클리드는 특징적인 번식 형태와 잘 발달된 행동 양식을 가지고 있다. 번식 형태는 크게 알을 입 속에서 부화시키는 유형, 부모가 산란된 알을 지키는 유형, 그리고 부화한 치어를 입 속에서 기르는 유형 등으로 나눌 수 있다. 특히 알이나 치어를 입 속에서 기르는 행동을 구강보육(mouth brooding)이라고 하며, 이러한 습성을 가진 물고기를 구강보육어(mouth brooder)라고 부른다.
알려진 종의 대부분은 동아프리카(900종 이상), 중앙아메리카(약 100종), 남아메리카(약 300종)에 집중적으로 분포한다. 이 지역들에서는 과거에 폭발적인 종 분화가 일어났을 것으로 추정된다.
틸라피아 등 일부 종은 일본 등 다른 지역에 유입되어 양식되거나 외래종으로 정착하기도 했다.
3. 1. 먹이
시클리드과는 동물계에서 가능한 거의 모든 먹이 섭취 방식을 포괄하며, 육식성, 초식성, 잡식성, 플랑크톤식성, 그리고 부식성 어종이 모두 존재한다. 여러 종들은 특정 먹이원에 대한 형태학적 적응을 보이지만,[42] 대부분의 시클리드는 이용 가능성에 따라 더 다양한 종류의 먹이를 섭취한다.
육식성 시클리드는 먹이의 종류와 사냥 방식에 따라 어식성과 연체동물식성으로 나눌 수 있다. 어식성 시클리드는 다른 물고기, 치어, 유생 및 알을 먹는다. 일부 종은 입으로 새끼를 품는 종의 어미를 머리로 들이받아 새끼를 빼앗아 먹기도 한다.[43] 연체동물식성 시클리드는 다양한 사냥 전략을 사용한다. 예를 들어, 말라위 호에 서식하는 일부 시클리드는 바닥의 퇴적물을 입에 머금고 아가미갈퀴를 이용해 걸러내어 그 안에 사는 연체동물을 먹는다. 아가미갈퀴는 물고기의 아가미에 줄지어 있는 빗살 모양의 구조물로, 물과 함께 들어온 먹이가 아가미 밖으로 빠져나가지 않도록 걸러내는 역할을 한다.[44]
많은 시클리드는 주로 초식동물이며, 조류 (예: ''Petrochromis'' 속)나 식물 (예: ''Etroplus suratensis'')을 먹는다. 작은 동물, 특히 무척추동물은 식단에서 아주 작은 부분만을 차지한다.
다른 시클리드는 부식성으로, 'Aufwuchs'(아우프북스)라고 불리는 바위나 식물 표면에 붙어 자라는 조류, 작은 생물, 유기 찌꺼기 등을 먹는다. 이러한 종에는 ''Oreochromis'', ''Sarotherodon'', 그리고 ''Tilapia'' 속의 틸라피아아과 어류들이 포함된다.
또 다른 시클리드들은 포식성으로, 식물성 먹이는 거의 또는 전혀 먹지 않는다. 여기에는 다른 물고기나 곤충 유충 등 다양한 작은 동물을 잡아먹는 잡식성 종 (예: ''Pterophyllum'')과 특정 먹이에 특화된 종들이 포함된다. ''Trematocranus'' 속은 달팽이를 전문적으로 먹고, ''Pungu maclareni''는 해면동물을 먹는다. 여러 시클리드 종은 다른 물고기를 통째로 또는 부분적으로 먹는다. ''Crenicichla'' 속은 은신처에 숨어 있다가 지나가는 작은 물고기를 덮치는 매복형 포식자이고, ''Rhamphochromis'' 속은 넓은 물에서 먹이를 쫓는 추격형 포식자이다.[46] ''Caprichromis'' 속과 같은 포식 시클리드는 다른 종의 알이나 새끼를 먹으며, 때로는 입으로 새끼를 품는 어미의 머리를 들이받아 새끼를 토하게 만들어 잡아먹기도 한다.[47][48][49][50] 더욱 특이한 먹이 섭취 전략으로는 다른 물고기의 비늘이나 지느러미를 먹는 종들이 있는데, 이를 비늘식(lepidophagy)이라고 한다. ''Corematodus'', ''Docimodus evelynae'', ''Plecodus'', ''Perissodus'', ''Genyochromis'' 속 등이 여기에 해당한다.[51][52][53] ''Nimbochromis''와 ''Parachromis'' 속의 일부 종은 죽은 척하는 행동으로 작은 물고기를 유인한 후 갑자기 덮쳐 잡아먹는다.[54][55]
이처럼 다양한 먹이 섭취 방식은 시클리드가 매우 다양한 서식 환경에 적응하여 살아가는 데 중요한 역할을 했다. 특히 시클리드의 인두치(목구멍에 있는 이빨)는 매우 다양한 먹이를 처리할 수 있게 해주는 중요한 구조이다. 턱으로 먹이를 잡고 고정하는 동안 인두치를 이용해 먹이를 부수거나 찢는 방식으로 다양한 종류의 먹이를 효율적으로 섭취할 수 있다.
3. 2. 행동
시클리드의 공격적 행동은 상당히 의례화되어 있으며, 경쟁자를 평가하고 대립하는 과정에서 여러 과시 행동을 보인다.[56] 이런 과시 행동은 주로 짝짓기 시기가 다가올 때 나타난다. 의례화된 공격성의 예시로는 빠른 색깔 변화가 있는데, 싸움에서 이겨 우위를 점한[56] 수컷은 더욱 선명하고 밝은 색을 띠는 반면, 경쟁에서 밀려난 열성 수컷은 칙칙하고 탁한 색을 띤다. 색깔 변화 외에도, 시클리드는 측선을 이용해 상대방 주변 물의 움직임을 감지하여 경쟁 수컷의 신체적 능력이나 건강 상태를 평가하기도 한다.[58]
수컷 시클리드는 번식기에 매우 강한 영역성을 보이는데,[59] 자신의 영역과 사회적 지위를 지키기 위해 도전자[60]를 물리적으로 몰아낸다.[59] 이때 사용하는 방식으로는 서로 나란히 서서 몸 크기를 비교하거나 아가미를 부풀리는 측면 과시,[61] 직접 물어뜯기, 또는 입을 크게 벌리고 정면으로 부딪혀 서로의 턱 크기를 재고 턱을 무는 입싸움[61] 등이 있다. 시클리드 사회는 보통 하나의 우점 개체와 여러 마리의 열성 개체로 나뉘는 이분법적 구조를 가지며, 수컷의 신체적 공격성은 짝, 영역, 먹이와 같은 자원을 차지하기 위한 경쟁의 수단이 된다.[59] 암컷 시클리드는 일반적으로 선명한 색을 띠고 먹이가 풍부한 영역을 확보한, 성공적인 알파 수컷과 짝짓기를 선호한다.
시클리드는 열대어 중에서는 사나운 편으로 알려져 있어, 다른 종과 함께 기를 때는 주의가 필요하다.
번식 행동 역시 특징적이며 잘 발달된 양상을 보인다. 시클리드의 번식 형태는 크게 세 가지로 나눌 수 있다.
이처럼 입 속에서 알이나 치어를 기르는 행동을 구강보육(mouth brooding)이라고 하며, 이런 습성을 가진 물고기를 구강보육어(mouth brooder)라고 부른다.
3. 3. 번식
시클리드는 매우 조직적인 번식 활동을 보이며,[19] 모든 종은 알과 유생 모두에 대해 어느 정도의 부모 양육을 보여준다. 자유롭게 헤엄치는 새끼를 몇 주 또는 몇 달 동안 보살피는 경우도 많다.
짝짓기 시기가 가까워지면 시클리드의 의례화된 공격 행동이 나타난다. 이는 경쟁자를 평가하고 우위를 점하기 위한 여러 과시 행동으로 이루어진다.[56] 특히 색깔이 빠르게 변하는데, 우점하는[56] 수컷은 더욱 선명하고 밝은 색을 띠는 반면, 열성 수컷은 칙칙하고 탁한 색을 띤다. 또한 측선을 이용해 상대 주변 물의 움직임을 감지하여 경쟁 수컷의 신체 능력을 평가하기도 한다.[58] 수컷 시클리드는 번식기에 매우 영역성이 강해지며, 측면 과시(평행 배열, 아가미 드러내기),[61] 물어뜯기, 또는 입싸움(입을 벌린 채 정면으로 부딪혀 턱 크기를 재고 서로 물어뜯기)[61]을 통해 다른 수컷[60]을 물리적으로 몰아내고[59] 자신의 영역과 사회적 지위를 확립한다.[59] 이러한 경쟁은 짝, 영역, 먹이와 같은 자원을 얻기 위함이다.[59] 암컷 시클리드는 선명한 색깔을 띠고 먹이가 풍부한 영역을 가진 성공적인 알파 수컷과 짝짓기를 선호한다.
시클리드는 일부일처제 또는 다부일처제로 짝짓기를 한다.[14] 특정 종의 짝짓기 방식이 육아 방식과 반드시 일치하는 것은 아니다. 예를 들어, 대부분의 일부일처제 시클리드는 구강 포란(입 속에서 알이나 새끼를 키우는 방식)을 하지 않지만, ''Chromidotilapia'', ''Gymnogeophagus'', ''Spathodus'', ''Tanganicodus'' 속에는 일부일처제이면서 구강 포란을 하는 종들이 포함되어 있다. 반대로, 개방된 곳이나 동굴에 알을 낳는 시클리드 중에는 다부일처제인 경우가 많다. 예를 들어 ''Apistogramma'', ''Lamprologus'', ''Nannacara'', ''Pelvicachromis'' 속의 많은 종들이 그렇다.[14][62]
대부분의 성체 수컷 시클리드, 특히 Haplochromini 부족의 수컷은 뒷지느러미에 독특한 타원형의 색 점 무늬를 가지는데, 이를 알 반점이라고 부른다. 이 알 반점은 시클리드의 구강 포란 과정에서 중요한 역할을 한다. 알 반점은 카로티노이드 색소 세포로 이루어져 있는데, 물고기는 스스로 카로티노이드를 합성할 수 없으므로 이는 생존에 상당한 비용을 요구하는 특징이다.[63] 수컷은 이 알 반점을 이용하여 수정 과정을 돕는다. 구강 포란을 하는 암컷은 알을 낳자마자 즉시 입으로 물어 보호한다. 이때 수컷은 뒷지느러미를 암컷 앞에 보여주며 알 반점을 드러낸다. 암컷은 이것을 자신이 낳은 알로 착각하고 수컷의 뒷지느러미 근처(특히 생식 유두)에 입을 가져다 대는데, 바로 이 순간 수컷이 정자를 방출하여 암컷의 입 안에 있는 알들을 수정시킨다.[63][64] 진정한 알 반점은 노란색, 빨간색 또는 주황색의 내부 원과 이를 둘러싼 무색 고리로 이루어져 있다. 미토콘드리아 ''ND2'' 유전자 분석 결과, 진정한 알 반점은 ''Astatoreochromis'' 계통과 현대 ''Haplochromini'' 종의 공통 조상에서 진화한 것으로 추정된다. 이 조상은 강과 같은 하천 환경에 살았을 가능성이 높다.[65] 탁한 하천 환경에서는 이러한 시각적 신호가 종 내부 의사소통에 특히 유용했을 것으로 보인다.[65] ''fhl2-a''와 ''fhl2-b''라는 두 상동 유전자가 알 반점 패턴 및 색상 형성과 관련이 있는 것으로 밝혀졌다.[64]
시클리드는 기질(바닥이나 물체 표면)에 알을 낳는 것과 관련하여 다양한 행동을 보인다. 여기에는 구애, 둥지 짓기, 알과 새끼 돌보기 등이 포함된다. 번식기가 아닐 때 시클리드는 주로 영역을 확보하고 방어하는 데 집중한다. 수컷끼리 또는 암컷끼리의 만남은 공격적이지만, 수컷과 암컷의 만남은 구애로 이어질 수 있다.[79] 수컷의 구애는 영역 확보로 시작되며, 때로는 짝을 유인하기 위해 둥지를 짓기도 한다.[68][75][79] 이후 수컷은 다양한 과시 행동(렉킹)을 통해 암컷을 자신의 영역으로 유인하거나 직접 암컷을 찾아 나선다.[68] 산란 시기에는 행동이 변하여 외부 자극에 덜 반응하게 되며,[79] 이는 종종 외형의 생리적 변화를 동반한다.[67][70][79]
시클리드는 어미, 아비 또는 부모 모두가 새끼를 돌보는 다양한 양육 방식을 보인다. 구강 포란 종에서는 어미가 주로 돌보지만, 시클리드의 공통 조상은 아비만 새끼를 돌봤을 것으로 여겨진다.[76] 부모 외 다른 개체가 새끼 양육을 돕기도 한다. 예를 들어, 부모 모두 새끼를 돌보는 수선화 시클리드(Neolamprologus pulcher)의 경우, 주된 번식 쌍 외에 주변 영역에 머무는 가까운 혈연관계의 위성 수컷들이 새끼 양육을 돕는다.[80]
새끼에게 먹이를 공급하는 것도 흔한 돌봄 행동이다. 예를 들어, 긴꼬리 시클리드(Neolamprologus modabu) 암컷은 새끼가 있을 때 영양분이 풍부한 유기물과 동물성 플랑크톤을 물속에 흩뿌리기 위해 모래 바닥을 더 많이 파헤친다.[73][81] 이러한 행동은 컨빅트 시클리드(Cichlasoma nigrofasciatum)에서도 관찰된다.[72][81] 어떤 시클리드는 피부에서 점액을 분비하여 새끼에게 먹이기도 하고, 잡은 먹이를 씹어서 나눠주기도 한다. 다만, 동굴에 알을 낳는 종에서는 이러한 먹이 공급 방식이 덜 흔하다.[81]
여러 쌍이 새끼 무리를 함께 돌보는 공동 양육은 Amphilophus citrinellus, Etroplus suratensis, Tilapia rendalli 등 여러 종에서 관찰되었다.[82][83][84] 또한, 큰 무리를 이루어 사는 Neolamprologus brichardi의 치어는 성체뿐만 아니라 이전에 태어난 형제자매 유어들의 보호도 받는다.[85] 디스커스(Symphysodon spp.), 일부 Amphilophus 종, Etroplus 종, Uaru 종 등은 피부 분비물(점액)을 먹여 새끼를 키운다.[14][108] Neolamprologus pulcher 종은 우점하는 번식 쌍에게 복종하는 여러 마리의 조력자가 있는 협동 번식 시스템을 사용한다.
부모 양육 방식은 크게 네 가지 범주로 나눌 수 있다.[108]
# 기질 산란 (개방형 산란): 알을 바위, 잎, 통나무 등 개방된 곳에 낳는다. 예로는 ''디스커스속'' 및 ''엔젤피시속'' 종, ''아노말로크로미스 토마시'' 등이 있다. 수컷과 암컷 부모는 보통 역할을 분담한다. 수컷은 영역을 순찰하며 침입자를 쫓아내고, 암컷은 알에 물을 부채질해주거나 수정되지 않은 알을 골라내고, 치어를 이끌고 먹이를 찾는다. 하지만 두 성별 모두 모든 양육 행동을 할 수 있다.[87]
# 동굴 산란 (은밀한 동굴 산란): 동굴, 틈새, 구멍 또는 버려진 조개껍데기 안에 알을 낳으며, 종종 천장에 알을 붙인다. 예를 들어 ''펠비카크로미스'' 속, ''아르코센트루스'' 속, ''Apistogramma'' 속 등이 있다.[108] 자유롭게 헤엄치는 치어와 부모는 몸짓(흔들기, 배지느러미 흔들기 등)으로 소통한다. 또한, 부모는 잎을 뒤집거나 지느러미로 바닥을 파헤쳐 치어가 먹이를 찾도록 돕기도 한다.[87]
# 구강 포란 (Mouth brooding): 알이나 부화한 유생을 입 안에 넣고 기르는 방식으로, '마우스브루딩'이라고도 불린다. 구강 포란은 다시 두 가지 유형으로 나뉜다.[87]
구강 포란은 대부분 암컷이 담당하지만, 예외적으로 수컷도 참여하는 경우가 있다. 에레트모딘 시클리드(속: 스파토두스, 에레트모두스, 탕가니코두스), 일부 사로테로돈 종(예: 사로테로돈 멜라노테론)[88], 크로미도틸라피아 귄테리, 그리고 일부 아에퀴덴스 종에서는 수컷이 구강 포란에 참여한다.[14][87][89] 구강 포란 방식은 아프리카 시클리드의 여러 그룹에서 독립적으로 진화한 것으로 보인다.[19]
4. 형태
시클리드과는 몸 크기의 범위가 매우 넓어, 암컷 ''네올람프롤로구스 멀티파스키아투스''처럼 몸길이가 2.5cm에 불과한 종부터 ''불렌게로크로미스''와 ''시클라''처럼 몸길이가 1m에 가까운 종까지 다양하다. 시클리드과 어류는 전반적으로 몸 형태의 다양성도 높아, ''알톨람프롤로구스'', ''프테로필룸'', ''심피소돈''처럼 옆으로 강하게 납작한 종부터 ''줄리도크로미스'', ''텔레오그라마'', ''텔레오시클라'', ''크레니키클라'', ''고비오시클라''처럼 원통형이고 길쭉한 종까지 다양하다.[14] 하지만 일반적으로 시클리드과 어류는 중간 크기에, 타원형이고 약간 옆으로 납작하며, 북아메리카산 태양어과 어류와 형태, 행동, 생태적으로 비슷하다.[15]
시클리드과 어류는 아래 인두골이 단일 치아 구조로 융합된다는 하나의 중요한 특징을 공유한다. 복잡한 근육 세트는 위아래 인두골을 음식물을 처리하는 두 번째 턱 세트로 사용할 수 있게 하여, "진짜 턱" (하악골)과 "인두 턱" 사이에 노동 분담을 가능하게 한다. 시클리드과 어류는 효율적이고 종종 고도로 특수화된 먹이 섭취자로, 매우 다양한 먹이를 포획하고 처리한다. 이것이 시클리드과가 그렇게 다양한 이유 중 하나로 여겨진다.[14]
농어목 다른 과와 구별되는 특징은 다음과 같다.[16]
5. 분류
쿨란더(Sven O. Kullander)는 1998년 시클리드과를 8개의 아과로 분류하였다. 여기에는 아스트로노투스아과(Astronotinae), 키클라소마아과(Cichlasomatinae), 키클라아과(Cichlinae), 에트로플루스아과(Etroplinae), 게오파구스아과(Geophaginae), 헤테로크로미스아과(Heterochromidinae), 프세우도크레닐라브루스아과(Pseudocrenilabrinae), 레트로쿨루스아과(Retroculinae)가 포함된다.[17] 이후 스파크스와 스미스는 9번째 아과인 프티코크로미스아과(Ptychochrominae)를 추가하였다.[18]
그러나 시클리드과의 분류는 여전히 논쟁 중이며, 속(genus) 수준의 분류는 아직 명확하게 정립되지 않았다. 단계통군(monophyly)을 이루는 속에 종들을 배정하는 포괄적인 시스템이 부족하며, 어떤 속을 인정해야 하는지에 대한 완전한 합의도 이루어지지 않은 상태이다.[19]
분류 문제의 예로, 쿨란더[20]는 아프리카 속인 ''Heterochromis''를 계통발생학적으로 신열대(Neotropical) 시클리드 내에 위치시켰지만, 후속 연구에서는 다른 결론을 내렸다. 다른 문제는 빅토리아호 초대집단(단일 서식지를 공유하는 많은 밀접하게 관련된 종)의 가상의 공통 조상의 정체와 탕가니카호 시클리드의 조상 계통에 집중되어 있다.
형태적 특징에서 유래한 계통 발생은 유전자좌(Locus (genetics))를 기반으로 한 계통 발생과 속 수준에서 차이를 보인다.[21] 시클리드과(Cichlidae)는 단계통군이라는 데에는 여전히 합의가 있다.[22]
시클리드 분류에서 치열(dentition)은 이전에는 분류 특징으로 사용되었지만, 많은 시클리드에서 마모로 인해 나이에 따라 치아 모양이 변하기 때문에 신뢰할 수 없다는 점이 복잡한 문제였다. DNA 서열 분석(DNA sequencing) 및 기타 기술은 시클리드 분류를 변화시켰다.
또는, 신세계(New World)에 서식하는 모든 시클리드 종은 키클라아과(Cichlinae)로 분류하고, 구세계(Old World)에 서식하는 모든 시클리드 종은 에트로플루스아과(Etroplinae)로 분류하는 대안도 있다.
아래는 FishBase 기준 속 목록이다. 특히 아프리카 동아프리카 지구대 호수의 압상어과(Haplochromine) 어류에 대한 연구는 계속 진행 중이다.[16]
| 속명 | 명명자 | 연도 |
|---|---|---|
| Abactochromis | 올리버 & 아르네가르드 | 2010 |
| Acarichthys | 아이겐만 | 1912 |
| Acaronia | 마이어스 | 1940 |
| Alcolapia | 티스 반 덴 아우데나르데 | 1969 |
| Alticorpus | 스토퍼 & 맥케이 | 1988 |
| Altolamprologus | 폴 | 1986 |
| Amatitlania | 슈미터-소토 | 2007 |
| Amphilophus | 아가시 | 1859 |
| Andinoacara | 무실로바, 리칸 & 노박 | 2009 |
| Anomalochromis | 그린우드 | 1985 |
| Apistogramma | 리건 | 1913 |
| Apistogrammoides | 마이켄 | 1965 |
| Aristochromis | 트레바스 | 1935 |
| Astatoreochromis | 펠레그랭 | 1904 |
| Astatotilapia | 펠레그랭 | 1904 |
| Astronotus | 스웨인슨 | 1839 |
| Aulonocara | 리건 | 1922 |
| Aulonocranus | 리건 | 1920 |
| Australoheros | 리칸 & 쿨란더 | 2006 |
| Baileychromis | 폴 | 1986 |
| Bathybates | 불렌저 | 1898 |
| Benitochromis | 람보이 | 2001 |
| Benthochromis | 폴 | 1986 |
| Biotodoma | 아이겐만 & 케네디 | 1903 |
| Biotoecus | 아이겐만 & 케네디 | 1903 |
| Boulengerochromis | 펠레그랭 | 1904 |
| Buccochromis | 에클스 & 트레바스 | 1989 |
| Bujurquina | 쿨란더 | 1986 |
| Callochromis | 리건 | 1920 |
| Caprichromis | 에클스 & 트레바스 | 1989 |
| Caquetaia | 파울러 | 1945 |
| Cardiopharynx | 폴 | 1942 |
| Chaetobranchopsis | 슈타인다흐너 | 1875 |
| Chaetobranchus | 헤켈 | 1840 |
| Chalinochromis | 폴 | 1974 |
| Champsochromis | 불렌저 | 1915 |
| Cheilochromis | 에클스 & 트레바스 | 1989 |
| Chetia | 트레바스 | 1961 |
| Chilochromis | 불렌저 | 1902 |
| Chilotilapia | 불렌저 | 1908 |
| Chromidotilapia | 불렌저 | 1898 |
| Cichla | 블로흐 & 슈나이더 | 1801 |
| Cichlasoma | 스웨인슨 | 1839 |
| Cleithracara | 쿨란더 & 니센 | 1989 |
| Coelotilapia[127] | ||
| Congochromis | 스티아스니 & 슐리벤 | 2007 |
| Congolapia | ||
| Copadichromis | 에클스 & 트레바스 | 1989 |
| Coptodon[127] | ||
| Corematodus | 불렌저 | 1897 |
| Crenicara | 슈타인다흐너 | 1875 |
| Crenicichla | 헤켈 | 1840 |
| Cryptoheros | 알가이어 | 2001 |
| Ctenochromis | 페퍼 | 1893 |
| Ctenopharynx | 에클스 & 트레바스 | 1989 |
| Cunningtonia | 불렌저 | 1906 |
| Cyathochromis | 트레바스 | 1935 |
| Cyathopharynx | 리건 | 1920 |
| Cyclopharynx | 폴 | 1948 |
| Cynotilapia | 리건 | 1922 |
| Cyphotilapia | 리건 | 1920 |
| Cyprichromis | 쉐어만 | 1977 |
| Cyrtocara | 불렌저 | 1902 |
| Danakilia | 티스 반 덴 아우데나르데 | 1969 |
| Dicrossus | 슈타인다흐너 | 1875 |
| Dimidiochromis | 에클스 & 트레바스 | 1989 |
| Diplotaxodon | 트레바스 | 1935 |
| Divandu | 람보이 & 스녹스 | 2000 |
| Docimodus | 불렌저 | 1897 |
| Eclectochromis | 에클스 & 트레바스 | 1989 |
| Ectodus | 불렌저 | 1898 |
| Enigmatochromis | 람보이 | 2009 |
| Eretmodus | 불렌저 | 1898 |
| Etia | 슐리벤 & 스티아스니 | 2003 |
| Etroplus | 퀴비에 | 1830 |
| Exochochromis | 에클스 & 트레바스 | 1989 |
| Fossorochromis | 에클스 & 트레바스 | 1989 |
| Genyochromis | 트레바스 | 1935 |
| Geophagus | 헤켈 | 1840 |
| Gephyrochromis | 불렌저 | 1901 |
| Gnathochromis | 폴 | 1981 |
| Gobiocichla | 카나자와 | 1951 |
| Grammatotria | 불렌저 | 1899 |
| Greenwoodochromis | 폴 | 1983 |
| Guianacara | 쿨란더 & 니센 | 1989 |
| Gymnogeophagus | 미란다 리베이로 | 1918 |
| Haplochromis | 힐겐도르프 | 1888 |
| Haplotaxodon | 불렌저 | 1906 |
| Hemibates | 리건 | 1920 |
| Hemichromis | 페터스 | 1857 |
| Hemitaeniochromis | 에클스 & 트레바스 | 1989 |
| Hemitilapia | 불렌저 | 1902 |
| Herichthys | 베어드 & 지라드 | 1854 |
| Heroina | 쿨란더 | 1996 |
| Heros | 헤켈 | 1840 |
| Heterochromis | 리건 | 1922 |
| Heterotilapia[127] | ||
| Hoplarchus | 카우프 | 1860 |
| Hypselecara | 쿨란더 | 1986 |
| Hypsophrys | 아가시 | 1859 |
| Interochromis | 야마오카, 호리 & 쿠와무라 | 1988 |
| Iodotropheus | 올리버 & 루아젤 | 1972 |
| Iranocichla | 코드 | 1982 |
| Julidochromis | 불렌저 | 1898 |
| Katria | 스티아스니 & 스파크스 | 2006 |
| Konia | 트레바스 | 1972 |
| Krobia | 쿨란더 & 니센 | 1989 |
| Labeotropheus | 알 | 1926 |
| Labidochromis | 트레바스 | 1935 |
| Laetacara | 쿨란더 | 1986 |
| Lamprologus | 쉴트후이스 | 1891 |
| Lepidiolamprologus | 펠레그랭 | 1904 |
| Lestradea | 폴 | 1943 |
| Lethrinops | 리건 | 1922 |
| Lichnochromis | 트레바스 | 1935 |
| Limbochromis | 그린우드 | 1987 |
| Limnochromis | 리건 | 1920 |
| Limnotilapia | 리건 | 1920 |
| Lobochilotes | 불렌저 | 1915 |
| †Mahengechromis | 머레이 | 2001[128] |
| Maylandia | 마이어 & 포스터 | 1984 |
| Mazarunia | 쿨란더 | 1990 |
| Mchenga | 스토퍼 & 코닝스 | 2006 |
| Melanochromis | 트레바스 | 1935 |
| Mesonauta | 귄터 | 1862 |
| Microchromis | 존슨 | 1975 |
| Mikrogeophagus | 뮬렌그라흐트-매드슨 | 1968 |
| Myaka | 트레바스 | 1972 |
| Mylochromis | 리건 | 1920 |
| Naevochromis | 에클스 & 트레바스 | 1989 |
| Nandopsis | 길 | 1862 |
| Nannacara | 리건 | 1905 |
| Nanochromis | 펠레그랭 | 1904 |
| Neolamprologus | 콜롬브 & 알가이어 | 1985 |
| Nimbochromis | 에클스 & 트레바스 | 1989 |
| Nosferatu | 드 라 마자-베니뇨스, 오르넬라스-가르시아, 로자노-빌라노, 가르시아 라미레즈 & 도아드리오 | 2015[129] |
| Nyassachromis | 에클스 & 트레바스 | 1989 |
| Ophthalmotilapia | 펠레그랭 | 1904 |
| Oreochromis | 귄터 | 1889 |
| Orthochromis | 그린우드 | 1954 |
| Otopharynx | 리건 | 1920 |
| Oxylapia | 키에너 & 모주 | 1966 |
| Pallidochromis | 터너 | 1994 |
| Parachromis | 아가시 | 1859 |
| Paracyprichromis | 폴 | 1986 |
| Parananochromis | 그린우드 | 1987 |
| Paraneetroplus | 리건 | 1905 |
| Paratilapia | 블리커 | 1868 |
| Paretroplus | 블리커 | 1868 |
| Pelmatochromis | 슈타인다흐너 | 1894 |
| Pelmatolapia[127] | ||
| Pelvicachromis | 티스 반 덴 아우데나르데 | 1968 |
| Perissodus | 불렌저 | 1898 |
| Petenia | 귄터 | 1862 |
| Petrochromis | 불렌저 | 1898 |
| Petrotilapia | 트레바스 | 1935 |
| Pharyngochromis | 그린우드 | 1979 |
| Placidochromis | 에클스 & 트레바스 | 1989 |
| Plecodus | 불렌저 | 1898 |
| †Plesioheros | 말라바르바, 줄레타 & 델 파파 | 2006[3] |
| †Proterocara | 페레스, 말라바르바 & 델 파파 | 2010[3] |
| Protomelas | 에클스 & 트레바스 | 1989 |
| Pseudocrenilabrus | 파울러 | 1934 |
| Pseudosimochromis | 넬리센 | 1977 |
| Pseudotropheus | 리건 | 1922 |
| Pterochromis | 트레바스 | 1973 |
| Pterophyllum | 헤켈 | 1840 |
| Ptychochromis | 슈타인다흐너 | 1880 |
| Ptychochromoides | 키에너 & 모주 | 1966 |
| Pungu | 트레바스 | 1972 |
| Reganochromis | 휘틀리 | 1929 |
| Retroculus | 아이겐만 & 브레이 | 1894 |
| Rhamphochromis | 리건 | 1922 |
| Rocio | 슈미터-소토 | 2007 |
| Sargochromis | 리건 | 1920 |
| Sarotherodon | 뤼펠 | 1852 |
| Satanoperca | 귄터 | 1862 |
| Schwetzochromis | 폴 | 1948 |
| Sciaenochromis | 에클스 & 트레바스 | 1989 |
| Serranochromis | 리건 | 1920 |
| Simochromis | 불렌저 | 1898 |
| Spathodus | 불렌저 | 1900 |
| Steatocranus | 불렌저 | 1899 |
| Stigmatochromis | 에클스 & 트레바스 | 1989 |
| Stomatepia | 트레바스 | 1962 |
| Symphysodon | 헤켈 | 1840 |
| Taeniacara | 마이어스 | 1935 |
| Taeniochromis | 에클스 & 트레바스 | 1989 |
| Taeniolethrinops | 에클스 & 트레바스 | 1989 |
| Tahuantinsuyoa | 쿨란더 | 1991 |
| Tangachromis | 폴 | 1981 |
| Tanganicodus | 폴 | 1950 |
| Teleocichla | 쿨란더 | 1988 |
| Teleogramma | 불렌저 | 1899 |
| Telmatochromis | 불렌저 | 1898 |
| Theraps | 귄터 | 1862 |
| Thoracochromis | 그린우드 | 1979 |
| Thorichthys | 미크 | 1904 |
| Thysochromis | 다제 | 1988 |
| Tilapia | 스미스 | 1840 (틸라피아과 참조) |
| Tomocichla | 리건 | 1908 |
| Tramitichromis | 에클스 & 트레바스 | 1989 |
| Trematocara | 불렌저 | 1899 |
| Trematocranus | 트레바스 | 1935 |
| †Tremembichthys | 말라바르바 & 말라바르바 | 2008[3] |
| Triglachromis | 폴 & 티스 반 덴 아우데나르데 | 1974 |
| Tristramella | 트레바스 | 1942 |
| Tropheops | 트레바스 | 1984 |
| Tropheus | 불렌저 | 1898 |
| Tylochromis | 리건 | 1920 |
| Tyrannochromis | 에클스 & 트레바스 | 1989 |
| Uaru | 헤켈 | 1840 |
| Variabilichromis | 콜롬브 & 알가이어 | 1985 |
| Xenochromis | 불렌저 | 1899 |
| Xenotilapia | 불렌저 | 1899 |
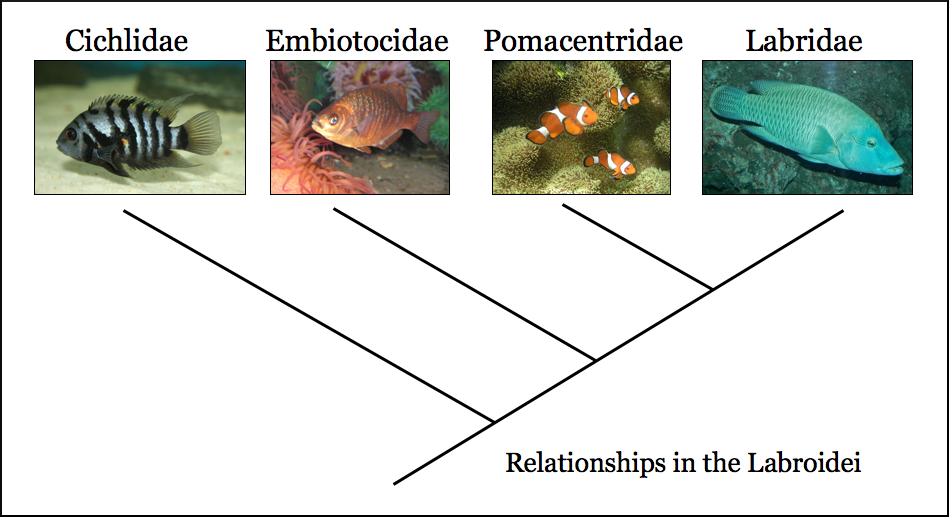
6. 계통 분류
시클리드는 전체 분류군과 서로 다른 서식지의 여러 군집 내에서 모두 최근에 일어난 빠른 진화 방산으로 유명하다.[69][71][75][78][76][77] 계통발생에서 여러 계통이 동일한 형질로 진화하는 평행 진화 현상과 조상 형질로 되돌아가는 복귀 사례가 여러 차례 관찰된다.
시클리드과는 농어목(Perciformes) 내에서 약 8천만 년에서 1억 년 전 사이에 처음 나타난 것으로 추정된다.[78] 시클리드과는 지리적 위치에 따라 크게 마다가스카르, 인도, 아프리카, 신열대(남미)의 몇 가지 그룹으로 나눌 수 있다. 이 중 가장 잘 알려져 있고 다양한 종을 포함하는 그룹은 아프리카 시클리드인데, 이는 다시 동부와 서부 종류로 나뉘거나, 종이 유래한 말라위호, 빅토리아호, 탕가니카호 등의 호수에 따라 분류하기도 한다.[78][76] 이러한 아종 중 마다가스카르와 인도 시클리드가 계통적으로 가장 먼저 분기되었으며, 종 다양성은 가장 낮다.
아프리카 시클리드 중에서는 서아프리카 또는 탕가니카호 시클리드가 가장 먼저 분기된 기저 그룹에 속한다.[71][78] 시클리드의 공통 조상은 알을 바닥이나 돌 같은 기질에 낳는 종이었을 것으로 여겨진다.[76] 마다가스카르와 인도 시클리드는 모두 이러한 기질 산란 특징을 유지하고 있다. 그러나 아프리카 시클리드 중 현존하는 기질 산란 종은 모두 탕가니카호에서만 유래한다.[69][76] 말라위호와 빅토리아호 시클리드의 조상은 입 속에서 알을 품는 종이었다. 마찬가지로, 남미 시클리드의 약 30%만이 조상의 기질 산란 형질을 유지하는 것으로 생각된다. 입 속에서 알을 품는 습성은 아프리카와 신열대 종 모두에서 최대 14번 개별적으로 진화했고, 기질 산란으로의 복귀는 최대 3번 일어난 것으로 생각된다.[76]
7. 주요 종
- '''아프리칸 시클리드''': 아프리카산 시클리드의 총칭이다.
- * 멈부나: 말라위호산 소형종 그룹의 현지명이다.
- * 틸라피아: 아시아, 아프리카에 분포하며, 일부 종은 일본의 온난한 지역에서 외래종으로 정착하기도 했다. (좁은 의미의 납자루 포함)
- * 펠비카크로미스 풀케르: 나이지리아 원산의 소형종으로, '펠마토'라는 별명으로도 불린다.
- '''아메리칸 시클리드''': 중남미산 시클리드의 총칭이다. 엔젤피시나 디스커스처럼 단독으로 유명한 그룹은 제외하는 경우도 있다.
- * 오스카(아스트로노투스) (''Astronotus ocellatus'')
- * 아이스팟 시클리드 (''Cichla ocellaris'') - 현지명 투쿠나레, 별명 피콕크바스
- * 컨빅트 시클리드 (''Amatitlania nigrofasciata'')
- * 플라밍고 시클리드 (''Amphilophus citrinellus'')
- * 디스커스 (''Symphysodon'' 속)의 2종류
- * 엔젤피시 (''Pterophyllum'' 속)의 4종류
8. 인간과의 관계
시클리드는 중앙아메리카에서 남아메리카, 마다가스카르를 포함한 아프리카, 중동, 남아시아에 이르기까지 넓은 지역에 분포하며 인간과 다양한 관계를 맺고 있다. 최소 1,300종 이상이 확인된 이들은 담수와 기수 환경에 서식한다.
인간과의 주요 관계로는 식용, 관상용, 낚시 대상 등이 있다. 틸라피아와 같은 종은 중요한 수산 자원으로 전 세계적으로 양식된다. 또한 엔젤피시나 디스커스처럼 화려한 색상을 가진 종들은 관상용 열대어로 인기가 높다. 일부 대형 종은 낚시 대상어로도 주목받는다.
독특하고 잘 발달된 번식 행동 양식은 시클리드의 중요한 특징 중 하나로, 알이나 치어를 입안에서 기르는 구강보육(mouth brooding) 등이 관찰된다.[69] 이러한 특성은 진화생물학 및 동물행동학 연구에서 중요한 대상이 되기도 한다.[70][71]
한편, 틸라피아처럼 일부 종은 본래 서식지가 아닌 다른 지역에 도입되어 양식되거나 외래종으로 정착하기도 했는데, 이 과정에서 토착 생태계에 영향을 미치는 문제가 발생하기도 한다.
8. 1. 관상어

1945년 이후로 시클리드는 수족관 어류로서 인기가 높아졌다.[14][108][87][109][110][111][112]
수족관에서 가장 흔하게 볼 수 있는 종은 열대 남아메리카의 아마존 강 유역이 원산지인 ''Pterophyllum scalare''로, 흔히 "엔젤피쉬"라는 이름으로 알려져 있다. 그 외에도 오스카 (''Astronotus ocellatus''), 컨빅트 시클리드 (''Archocentrus nigrofasciatus''), 디스커스 (''Symphysodon'') 등이 인기 있거나 쉽게 구할 수 있는 종이다.[14] 특히 엔젤피쉬나 디스커스처럼 색채가 아름다운 종들이 관상용 열대어로 많이 유통된다.
다만, 시클리드는 열대어 중에서는 비교적 사나운 종으로 알려져 있어 다른 종류의 물고기와 함께 기를 때(혼영) 주의가 필요하다.
8. 2. 식용 및 낚시 대상어
시클리드는 대부분 소형에서 중형 크기이지만, 많은 종이 식용 및 낚시 대상 어종으로 주목받는다. 살이 맛있고 뼈가 굵지 않아 중앙아메리카와 남아메리카, 그리고 아프리카 동아프리카 열곡대 주변 지역에서는 소규모 어업의 대상이 되기도 한다.[105]가장 중요한 식용 시클리드는 북아프리카 원산의 틸라피아이다. 틸라피아는 성장이 빠르고 높은 밀도의 사육 환경에도 잘 견디며 적응력이 뛰어나, 아시아 여러 지역에 도입되어 널리 양식되고 있으며 다른 지역에서도 양식 대상으로 인기를 얻고 있다. 틸라피아 양식 생산량은 연간 약 150만ton에 달하며, 그 가치는 18억달러[106]로 추정되어 연어나 송어 양식 규모와 비슷하다.
다른 육식성 어류와 달리 틸라피아는 조류나 식물성 먹이를 먹을 수 있어 양식 비용을 절감할 수 있다. 또한, 먹이가 되는 다른 어종에 대한 남획 압력을 줄이고, 먹이 사슬 상위 단계에서 축적될 수 있는 독소 농축 문제도 피할 수 있다. 이러한 장점 때문에 틸라피아는 "수중 닭고기"라는 별명으로 불리며 선호된다.[105]
많은 대형 시클리드는 인기 있는 낚시 대상어이기도 하다. 남아메리카의 피콕 베스(''Cichla'' 속)는 가장 인기 있는 스포츠 낚시 대상어 중 하나로, 전 세계 여러 수역에 도입되었다. 미국 플로리다주에서는 피콕 베스가 연간 800만달러 이상의 어업 및 스포츠 낚시 관련 수익과 수백만 시간의 낚시 활동을 창출하는 것으로 알려져 있다.[107] 낚시꾼들이 선호하는 다른 시클리드로는 오스카, 마야 시클리드( ''Cichlasoma urophthalmus''), 그리고 재규어 시클리드( ''Parachromis managuensis'') 등이 있다.[107]
8. 3. 잡종 및 품종 개량

일부 시클리드는 야생과 인공 조건 모두에서 관련 종과 쉽게 잡종화된다.[113] 이는 유럽 잉어과 어류와 같은 다른 어류 그룹에서도 나타나는 현상이다.[114] 특이하게도 시클리드 잡종은 특히 양식과 수족관 거래에서 광범위하게 상업적으로 이용되어 왔다.[7][115] 예를 들어, 틸라피아의 붉은색 잡종 품종은 빠른 성장 속도 때문에 양식에서 종종 선호된다. 또한, 틸라피아 잡종화는 연못에서의 개체 수 밀도를 조절하거나 번식을 막기 위해 수컷만으로 이루어진 개체군을 생산하는 데 이용되기도 한다.[7]
수족관에서 가장 흔하게 볼 수 있는 잡종 중 하나는 블러드 파라웃 시클리드(blood parrot cichlid)이다. 이 물고기는 여러 종, 특히 ''암필로푸스속''(Amphilophus)에 속하는 종들의 교배종으로 여겨진다. 가장 유력한 조합은 ''암필로푸스 라비아투스''(Amphilophus labiatus)와 ''비에하 신스필루스''(Vieja synspillus)의 교배이다. 블러드 파라웃 시클리드는 삼각형 모양의 입, 비정상적인 척추, 그리고 때때로 꼬리지느러미가 없는('러브 하트' 형태로 알려짐) 독특한 외형 때문에 수족관 애호가들 사이에서 논란이 되기도 한다. 일부는 이 물고기를 '물고기 세계의 프랑켄슈타인 괴물'이라고 부르기도 했다.[116] 또 다른 주목할 만한 잡종으로는 플라워혼 시클리드(flowerhorn cichlid)가 있다. 이 시클리드는 2001년부터 2003년 후반까지 아시아 일부 지역에서 큰 인기를 끌었으며, 소유자에게 행운을 가져다준다고 믿어졌다.[117] 그러나 2004년 이후 인기가 감소하면서[118] 일부 소유주들이 많은 개체를 말레이시아와 싱가포르의 강과 운하에 방류했고, 이로 인해 토착 어종의 생존을 위협하는 문제가 발생하고 있다.[119]

수많은 시클리드 종들은 관상용 수족관 품종을 개발하기 위해 선택적으로 사육되어 왔다. 특히 엔젤피시와 디스커스는 집중적인 품종 개량 프로그램을 통해 색상과 지느러미 형태에 영향을 미치는 많은 돌연변이 품종이 만들어졌다.[14][120][121] 다른 시클리드 종들, 예를 들어 오스카, 콘빅트 시클리드, ''펠비카크로미스 풀케르'' 등은 알비노, 백색증, 황색증과 같은 색소 돌연변이를 위해 사육되었다.[14][108] 이러한 색소 돌연변이는 우성 또는 열성 유전 형질로 나타날 수 있다.[122] 예를 들어, 콘빅트 시클리드의 백색증은 열성 유전 형질이지만,[123] ''오레오크로미스 닐로티쿠스 닐로티쿠스''의 붉은색은 우성 유전 돌연변이에 의해 나타난다.[124]
그러나 이러한 선택적 번식은 의도하지 않은 결과를 낳기도 한다. 예를 들어, ''미크로게오파구스 라미레지''의 잡종 품종은 건강 문제나 번식 능력 저하 문제를 겪는 경우가 있다.[125] 마찬가지로, 의도적인 근친교배는 엔젤피시에서 '노치'(notched) 표현형과 같은 신체적 이상을 유발할 수도 있다.[126]
참조
[1]
논문
Biogeography of the Mesoamerican Cichlidae (Teleostei: Heroini): Colonization through the GAARlandia land bridge and early diversification
https://www.research[...]
2024-03-27
[2]
논문
The evolution of pharyngognathy: A phylogenetic and functional appraisal of the pharyngeal jaw key innovation in labroid fishes and beyond
[3]
논문
On the Eocene cichlids from the Lumbrera Formation: additions and implications for the Neotropical ichthyofauna
[4]
서적
Fishes of the World
https://web.archive.[...]
Wiley
2018-11-05
[5]
웹사이트
List of nominal species of Cichlidae
http://www.fishbase.[...]
2012-02-00
[6]
서적
The Fresh and Brackish Water Fishes of Lower Guinea, West-Central Africa
Musée Royal de l'Afrique Centrale
[7]
논문
Ways of speciation in fishes
1963-06-00
[8]
논문
Captive breeding for the conservation of cichlid fishes
1990-12-00
[9]
논문
Out of Tanganyika: Genesis, explosive speciation, key-innovations and phylogeography of the haplochromine cichlid fishes
[10]
서적
The Cichlid Diversity of Lake Malawi / Nyasa / Niassa: Identification, distribution, and taxonomy
Cichlid Press
[11]
논문
African Cichlid Fishes: Model Systems for Evolutionary Biology
2000-11-00
[12]
웹사이트
Fact sheet for ''Oreochromis mossambicus'' {{small|(Peters, 1852)}}
https://web.archive.[...]
Gulf States Marine Fisheries Commission
2006-10-20
[13]
논문
Labroid intrarelationships revisited: morphological complexity, key innovations, and the study of comparative diversity
[14]
서적
The Cichlid Aquarium
Tetra Press
[15]
서적
The Diversity of Fishes
Blackwell Publishing, Inc.
[16]
FishBase
2012-02-00
[17]
서적
Phylogeny and classification of neotropical fishes
EDIPUCRS
[18]
논문
Phylogeny and biogeography of cichlid fishes (Teleostei: Perciformes: Cichlidae)
[19]
서적
Fishes of the World
John Wiley & Sons, Inc.
[20]
웹사이트
Phylogeny of major groups of cichlids
https://web.archive.[...]
2007-06-10
[21]
논문
Multilocus phylogeny of cichlid fishes (Pisces: Perciformes): evolutionary comparison of microsatellite and single-copy nuclear loci.
1998-07-01
[22]
논문
The species flocks of East African cichlid fishes: Recent advances in molecular phylogenetics and population genetics
http://nbn-resolving[...]
2004-06-00
[23]
논문
Priority management actions for alien freshwater fish species in Australia
2004-08-00
[24]
서적
Guide to Cichlids
T.F.H. Publications
[25]
웹사이트
Tilapia :: Far North Queensland
https://web.archive.[...]
2007-04-19
[26]
웹사이트
''Archocentrus nigrofasciatus'', Convict cichlid
https://web.archive.[...]
FishBase
2007-03-29
[27]
서적
Hawai'i's native and exotic freshwater animals
Mutual Publishing
[28]
서적
A field guide to freshwater fishes of North America north of Mexico
https://archive.org/[...]
Houghton Mifflin Company
[29]
웹사이트
Fact Sheet for Tilapia zilli (Gervais, 1848)
https://web.archive.[...]
Gulf States Marine Fisheries Commission
2005-08-03
[30]
웹사이트
Nonindigenous Fishes of Florida – With a Focus on South Florida
https://web.archive.[...]
U.S. Department of the Interior, U.S. Geological Survey, Coastal and Marine Science Center
2002-10-11
[31]
서적
The Cichlid Aquarium
Tetra Press
[32]
FishBase
[33]
FishBase
[34]
뉴스
Rough waters: one of the world's most turbulent rivers is home to a wide array of fish species. Now, large dams are threatening their future.
https://archive.toda[...]
2009-04-20
[35]
서적
Brackish-Water Fishes
Aqualog
[36]
논문
A new species of Danakilia (Teleostei, Cichlidae) from Lake Abaeded in the Danakil Depression of Eritrea (East Africa)
https://pdfs.semanti[...]
2010-11-29
[37]
논문
Cichlid biogeography: comment and review
2004-06-01
[38]
논문
Phylogeny and Taxonomic Revision of the Endemic Malagasy Genus Ptychochromis (Teleostei: Cichlidae), with the Description of Five New Species and a Diagnosis for Katria, New Genus
https://www.biodiver[...]
2006-01-01
[39]
논문
Phylogeny of the Cichlid Subfamily Etroplinae and Taxonomic Revision of the Malagasy Cichlid Genus Paretroplus (Teleostei: Cichlidae)
2008-01-01
[40]
FishBase
2011-07-01
[41]
FishBase
2011-07-01
[42]
웹사이트
Family Cichlidae-Cichlids
https://www.fishbase[...]
2019-12-04
[43]
웹사이트
Cichlidae
https://animaldivers[...]
University of Michigan Museum of Zoology
2019-12-03
[44]
서적
The Dissection of Vertebrates
[45]
논문
Melanochromis Crabro Sp. Nov.: a Cichlid Fish From Lake Malawi Which Feeds On Ectoparasites and Catfish Eggs
1981-01-01
[46]
웹사이트
Rhamphochromis esox
http://malawicichlid[...]
malawicichlids.com: The Cichlid Fishes of Lake Malawi
2007-04-19
[47]
논문
Paedophagia among cichlid fishes of Lake Victoria and Lake Malawi/Nyasa
[48]
논문
Head ramming behaviour by three paedophagous cichlids in Lake Malawi, Africa
[49]
논문
The disputed feeding behavior of a paedophagous haplochromine cichlid (Pisces) observed and discussed
[50]
논문
Paedophagy in Malawi cichlids
[51]
논문
An example of "mimicry" in fishes
[52]
논문
A taxonomic study of the genus ''Docimodus'' Boulenger (Pisces, Cichlidae) a group of fishes with unusual feeding habits from Lake Malawi
[53]
논문
Occasional egg-eating by the scale-eater ''Plecodus straeleni'' (Cichlidae) of Lake Tanganyika
[54]
논문
Feigning death in the Central American cichlid ''Parachromis friedrichsthalii''
[55]
논문
Field observation on death feigning: a unique hunting behavior by the predatory cichlid, ''Haplochromis livingstoni'', of Lake Malawi
[56]
웹사이트
Study: Cichlids can determine their social rank by observation
http://news.stanford[...]
Stanford University
2018-12-11
[57]
논문
Visual Information Alone Changes Behavior and Physiology during Social Interactions in a Cichlid Fish (Astatotilapia burtoni)
2011-05-25
[58]
논문
Fighting cichlids size up opposition with lateral line
2015-10-01
[59]
논문
Cognitive skills and the evolution of social systems
2017-01-01
[60]
논문
Two types of dominant male cichlid fish: behavioral and hormonal characteristics
2016-08-15
[61]
논문
Lateralization of lateral displays in convict cichlids
2011-10-23
[62]
논문
Alternative male mating acttics in a cichlid, ''Pelvicachromis pulcher'': a comparison of reproductive effort and success
[63]
논문
A sensory bias has triggered the evolution of egg-spots in cichlid fishes
2011-10-18
[64]
논문
The evolution of cichlid fish egg-spots is linked with a cis-regulatory change
2014-10-09
[65]
논문
Out of Tanganyika: Genesis, explosive speciation, key-innovations and phylogeography of the haplochromine cichlid fishes
2005-01-01
[66]
논문
Evidence for selfing in a vertebrate from whole-genome sequencing.
2023-12-27
[67]
논문
Social and reproductive physiology and behavior of the Neotropical cichlid fish Cichlasoma dimerus under laboratory conditions
2011-07-22
[68]
논문
Male size, spawning pit size and female mate choice in a lekking cichlid fish
1995-01-01
[69]
논문
Parallel life history evolution in mouthbrooding cichlids from the African Great Lakes
2008-10-07
[70]
논문
Reproductive Behaviour of the Rainbow Cichlid, Herotilapia Multispinosa (Pisces, Cichlidae)
1978-01-01
[71]
논문
Convergent Evolution within an Adaptive Radiation of Cichlid Fishes
2012-12-00
[72]
논문
Fin digging and leaf lifting by the convict cichlid, Cichlasoma nigrofasciatum: examples of parental food provisioning
1995-03-00
[73]
논문
Maternal Food Provisioning in a Substrate-Brooding African Cichlid
2014-06-09
[74]
논문
Mating and parental care in Lake Tanganyika's cichlids
2011-07-21
[75]
논문
Evolution of bower building in Lake Malawi cichlid fish: phylogeny, morphology, and behavior
2015-02-27
[76]
논문
Evolutionary transitions in parental care in cichlid fish
1998-12-07
[77]
논문
Temporal diversification of Central American cichlids
2010-12-00
[78]
논문
Evolutionary conservation of microsatellite flanking regions and their use in resolving the phylogeny of cichlid fishes (Pisces: Perciformes)
http://nbn-resolving[...]
1996-11-22
[79]
논문
Family Structure in the Dwarf Cichlid Apistogramma trifasciatum Eigenmann and Kennedy)
2010-04-26
[80]
논문
Reproductive parasitism of broodcare helpers in a cooperatively breeding fish
1999-09-01
[81]
서적
Cichlid Research: State of the Art
2001
[82]
논문
Communal Care and Kidnapping of Young by Parental Cichlids
1977
[83]
논문
Ethology and ecology of cichlid fishes of the genus ''Etroplus'' in Sri Lanka: Preliminary findings
1977
[84]
논문
Nest-building and communal care of young by ''Tilapia rendalli dumeril'' (pisces, cichlidae) in Lake Malawi
1981
[85]
웹사이트
Neolamprologus brichardi
http://www.africanci[...]
2008-04-08
[86]
논문
Reproductive guilds of fishes: A proposal and definition
1975
[87]
서적
Cichlid Fishes: Behaviour, ecology and evolution
Chapman and Hall
1991
[88]
논문
Paternal mouthbrooding in the black-chinned tilapia, ''Sarotherodon melanotheron'' (Pisces: Cichlidae): Changes in gonadal steroids and potential for vitellogenin transfer to larvae
2000-02-00
[89]
간행물
Mysterious mouthbrooders
1999-01-00
[90]
잡지
The extraordinary evolution of cichlid fishes
https://www.scientif[...]
2023-04-29
[91]
논문
Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa
1999
[92]
논문
Adaptive phenotypic plasticity in the Midas cichlid fish pharyngeal jaw and its relevance in adaptive radiation
2011
[93]
논문
Sympatric speciation in Nicaraguan crater lake cichlid fish
https://kops.uni-kon[...]
2006
[94]
논문
Ancient hybridization fuels rapid cichlid fish adaptive radiations
2017-02-10
[95]
보고서
IUCN Red List of Threatened Species
http://www.iucnredli[...]
IUCN
2011-04-26
[96]
IUCN
''Haplochromis thereuterion''
2010
[97]
뉴스
Dark secret of the lake
https://www.newscien[...]
2004-02-28
[98]
논문
Fisheries and Cichlid Evolution in the African Great Lakes: Progress and Problems
2009-12-00
[99]
논문
Adaptive responses in resurgent Lake Victoria cichlids over the past 30 years
2013-03-00
[100]
서적
Conservation Biology: For the coming decade
Springer
1998
[101]
논문
Cascading Effects of the Introduced Nile Perch on the Detritivorous/Phytoplanktivorous Species in the Sublittoral Areas of Lake Victoria
https://web.archive.[...]
1993-09-00
[102]
웹사이트
Geographic Patterns
http://www.iucnredli[...]
2017-03-25
[103]
논문
Wetland ecotones as refugia for endangered fishes
1996-12-00
[104]
논문
A pharyngeal jaw evolutionary innovation facilitated extinction in Lake Victoria cichlids
2015-11-27
[105]
서적
The Cichlid Fishes
https://archive.org/[...]
Cambridge, MA: Perseus Publishing
2000
[106]
서적
Tilapias as Alien Aquatics in Asia and the Pacific: A Review
Food & Agriculture Org.
2004
[107]
웹사이트
Fact Exotic Freshwater Fishes
http://floridafisher[...]
2007-03-18
[108]
서적
Aquarium Atlas
Tetra Press
[109]
서적
A Fishkeepers Guide to Central American Cichlids
Tetra Press
[110]
서적
Aquarium Fish
Harper Collins
[111]
서적
Back to Nature Guide to Malawi Cichlids
Druckhaus Beltz
[112]
서적
A Fishkeepers Guide to South American Cichlids
Tetra Press
[113]
학술지
Hybrid origin of a cichlid population in Lake Malawi: Implications for genetic variation and species diversity
2003-07
[114]
학술지
Fertility of roach × bream hybrids, ''Rutilus rutilus'' (L.) × ''Abramis brama'' (L.), and their identification
1987-03
[115]
웹사이트
Frequently asked questions on parrot cichlids
http://www.practical[...]
2006-10-20
[116]
웹사이트
It's The Frankenstein Monster of the Fish World: The Blood Parrot!
http://www.aquafrien[...]
AquaFriend.com
2002-10-27
[117]
뉴스
Singapore's 'lucky' pet Luohan can outnumber people in homes
2003-07-01
[118]
뉴스
Crayfish the latest fad among pet lovers
2004-09-03
[119]
뉴스
Flower horn: Joy in homes, a pest in rivers
2004-07-14
[120]
잡지
Angelfish genetics
[121]
학술지
Genetic diversity among wild forms and cultivated varieties of Discus (''Symphysodon'' spp.) as revealed by random amplified polymorphic DNA (RAPD) fingerprinting
1999-03-30
[122]
서적
Cichlid Fishes: Behaviour, ecology and evolution
Chapman and Hall
[123]
학술지
Inheritance of pink body colouration in ''Cichlasoma nigrofasciatum'' Günther (Pisces, Cichlidae)
[124]
학술지
The genetics and history of red, blond, and associated color variants in ''Oreochromis niloticus''
[125]
서적
American cichlids I: Dwarf Cichlids. A handbook for their identification, care and breeding.
Tetra Press
[126]
학술지
Notched – An Angelfish Deformity
[127]
학술지
Molecular phylogeny and revised classification of the haplotilapiine cichlid fishes formerly referred to as ''"Tilapia"''
[128]
학술지
Eocene cichlid fishes from Tanzania, east Africa
2001-01-19
[129]
학술지
Phylogeographic analysis of genus ''Herichthys'' (Perciformes: Cichlidae), with descriptions of ''Nosferatu'' new genus and ''H. tepehua'' n. sp.
[130]
서적
魚の名前
東京書籍
2006
[131]
웹인용
Phylogenetic Classification of Bony Fishes - Based mostly on Molecular Data — Version 3
http://www.deepfin.o[...]
2015-09-30
[132]
간행물
Phylogenetic classification of bony fishes
2017-07
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com