야이켈롭테루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
야이켈롭테루스는 프테리고투스과에 속하는 멸종된 유절지동물로, 다른 유르프테리드와 유사하게 확대된 협각과 납작한 미부를 가지고 있다. 독일에서 처음 발견되어 연구가 시작되었으며, 이후 속명과 종명이 변경되는 과정을 거쳤다. 야이켈롭테루스는 크기가 매우 컸으며, 2.5미터 이상으로 성장했을 것으로 추정된다. 이들은 강력한 포식자였으며, 겹눈을 통해 높은 시력을 확보하여 먹이를 사냥했을 것으로 보인다.
더 읽어볼만한 페이지
- 데본기의 동물 - 이크티오스테가
이크티오스테가는 데본기 후기에 살았던 사지동물로, 어류와 육상 동물의 특징을 모두 가지며, 튼튼한 사지와 척추를 가지고 물가에서 육상으로의 진출을 시도했던 초기 사지동물의 생활 방식을 보여준다. - 데본기의 동물 - 프테리고투스
프테리고투스는 실루리아기부터 데본기까지 존재했던 최대 1.75m의 대형 바다전갈로, 굽은 발톱과 강력한 협각을 이용해 먹이를 사냥하는 포식자였으며, 얕은 바다 등 다양한 환경에서 서식했다. - 광익목 - 바다전갈
바다전갈은 고생대 오르도비스기부터 페름기까지 살았던 멸종된 절지동물로, 머리가슴, 협각, 다리, 후체로 나뉘며 서새와 "Kiemenplatten"이라는 호흡기를 가졌고, 일부는 2.5m에 달하는 큰 크기로 바다와 담수에서 서식했다. - 광익목 - 메가라크네
메가라크네는 아르헨티나에서 발견된 멸종된 유절지동물로, 처음에는 거대한 거미로 오인되었으나, 2005년 바다 전갈로 재분류되었다.
| 야이켈롭테루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Jaekelopterus |
| 명명자 | Waterston, 1964 |
| 속 | 야이켈롭테루스속 |
| 타입 종 | †Jaekelopterus rhenaniae |
| 타입 종 명명자 | (Jaekel, 1914) |
| 이명 | Pterygotus rhenaniae Jaekel, 1914 Pterygotus howelli Kjellesvig-Waering & Størmer, 1952 |
| 시대 | 프라기안-초기 엠시안, |
| 하위 분류 | |
| 종 | †J. rhenaniae (Jaekel, 1914) †J. howelli (Kjellesvig-Waering & Størmer, 1952) |
2. 발견 및 연구 역사
오토 예켈은 1914년 독일 라인란트의 하부 데본기 퇴적층인 알켄에서 발견된 화석 전텔슨(텔슨 바로 앞의 분절)을 모식표본으로 하여 ''야이켈롭테루스''를 ''프테리고투스''의 한 종인 ''P. rhenaniae''로 처음 기술했다.[5] 예켈은 이 전텔슨이 ''프테리고투스''의 특징이라고 생각했으며, 발견된 다른 요소들은 이전에 알려진 ''P. buffaloensis''와 같은 종과 거의 다르지 않다고 보았다. 그는 당시 생물의 길이를 약 1미터(협각을 포함하면 1.5미터)로 추정했다.[5]
1936년 노르웨이 고생물학자 레이프 스퇴르메르는 독일 오버라트 근처에서 발견된 생식 부속기, 협각 및 중구개(복부의 일부인 큰 판) 및 텔슨 조각을 포함한 더 포괄적인 자료를 바탕으로 이 종에 대한 더 자세하고 포괄적인 설명을 제공했다.[6] 스퇴르메르는 생식 부속기가 다른 ''프테리고투스'' 종과 구별되는 분절형이라고 해석했다.[7]
1964년, 영국 고생물학자 찰스 워터스턴은 ''프테리고투스 레나니아에''를 수용하기 위해 ''야이켈롭테루스'' 속을 새로 설정했다. 그는 ''야이켈롭테루스''의 복부 부속기가 ''프테리고투스''의 부속기와 달리 분절되어 있다는 점을 들어 다른 ''프테리고투스'' 종과 충분히 다르다고 생각했다.[8] 워터스턴은 ''야이켈롭테루스''를 분절된 생식 부속기, 사다리꼴 전체, 좁고 긴 협각과 램에 직각에 가까운 말단 이빨, 그리고 약간 앞쪽으로 기울어진 주요 이빨과 확장된 말단 가시와 등쪽 용골이 있는 텔슨을 가진 프테리고티드라고 진단했다.[8] 속명은 오토 예켈을 기리기 위해 지어졌으며, 그리스어 단어 πτερόν (''pteron'')은 "날개"를 의미하며, 유립테루스 이름에서 흔히 사용되는 형용사이다.[8]
1974년, 스토르머는 ''Jaekelopterus''와 다른 프테리고티드의 생식 부속기의 상당한 차이점을 가정하여, 이 속을 수용하기 위해 새로운 과인 야이켈롭테리데를 세웠다.[7] 이 다른 특징은 1936년 스토르머의 오해였음이 입증되었으며, 사실 ''Jaekelopterus''의 생식 부속기는 ''프테리고투스''처럼 분절되지 않았다.[1] 따라서 야이켈롭테리데과는 이후 거부되었고 프테리고티데과와 동의어로 처리되었다.[7]
''프테리고투스''의 또 다른 종인 ''P. howelli''는 미국의 고생물학자 에릭 켈레스비그-와에링과 스토르머가 1952년에 와이오밍의 베어투스 버트 형성의 하부 데본기 퇴적층에서 발견된 화석 텔슨과 등판(몸 분절의 등쪽 부분)을 기반으로 명명했다. 종명 ''howelli''는 켈레스비그-와에링과 스토르머에게 설명에 사용된 화석 표본을 대여해 준 프린스턴 대학교의 벤자민 하웰 박사를 기리기 위해 지어졌다.[9] 이 종은 2007년 노르웨이 고생물학자 O. 에릭 테틀리에 의해 ''Jaekelopterus''로 지정되어 ''야이켈롭테루스 하웰리''가 되었다.[10]
2. 1. 오토 예켈의 초기 연구 (1914)
오토 예켈은 1914년 독일 라인란트의 하부 데본기 퇴적층인 알켄에서 발견된 화석 전텔슨(텔슨 바로 앞의 분절)을 모식표본으로 하여 ''야이켈롭테루스''를 ''프테리고투스''의 한 종인 ''P. rhenaniae''로 처음 기술했다.[5] 예켈은 이 전텔슨이 ''프테리고투스''의 특징이라고 생각했으며, 발견된 다른 요소들은 이전에 알려진 ''P. buffaloensis''와 같은 종과 거의 다르지 않다고 보았다. 그는 당시 생물의 길이를 약 1미터(협각을 포함하면 1.5미터)로 추정했다.[5]]]
2. 2. 추가 연구 및 학명 변경 (1936-2007)
1936년 노르웨이 고생물학자 레이프 스퇴르메르는 독일 오버라트 근처에서 발견된 생식 부속기, 협각 및 중구개(복부의 일부인 큰 판) 및 텔슨 조각을 포함한 더 포괄적인 자료를 바탕으로 이 종에 대한 더 자세하고 포괄적인 설명을 제공했다.[6] 스퇴르메르는 생식 부속기가 다른 ''프테리고투스'' 종과 구별되는 분절형이라고 해석했다.[7]1964년, 영국 고생물학자 찰스 워터스턴은 ''프테리고투스 레나니아에''를 수용하기 위해 ''야이켈롭테루스'' 속을 새로 설정했다. 그는 ''야이켈롭테루스''의 복부 부속기가 ''프테리고투스''의 부속기와 달리 분절되어 있다는 점을 들어 다른 ''프테리고투스'' 종과 충분히 다르다고 생각했다.[8] 워터스턴은 ''야이켈롭테루스''를 분절된 생식 부속기, 사다리꼴 전체, 좁고 긴 협각과 램에 직각에 가까운 말단 이빨, 그리고 약간 앞쪽으로 기울어진 주요 이빨과 확장된 말단 가시와 등쪽 용골이 있는 텔슨을 가진 프테리고티드라고 진단했다.[8] 속명은 오토 예켈을 기리기 위해 지어졌으며, 그리스어 단어 πτερόν (''pteron'')은 "날개"를 의미하며, 유립테루스 이름에서 흔히 사용되는 형용사이다.[8]
]]
1974년, 스토르머는 ''Jaekelopterus''와 다른 프테리고티드의 생식 부속기의 상당한 차이점을 가정하여, 이 속을 수용하기 위해 새로운 과인 야이켈롭테리데를 세웠다.[7] 이 다른 특징은 1936년 스토르머의 오해였음이 입증되었으며, 사실 ''Jaekelopterus''의 생식 부속기는 ''프테리고투스''처럼 분절되지 않았다.[1] 따라서 야이켈롭테리데과는 이후 거부되었고 프테리고티데과와 동의어로 처리되었다.[7]
''프테리고투스''의 또 다른 종인 ''P. howelli''는 미국의 고생물학자 에릭 켈레스비그-와에링과 스토르머가 1952년에 와이오밍의 베어투스 버트 형성의 하부 데본기 퇴적층에서 발견된 화석 텔슨과 등판(몸 분절의 등쪽 부분)을 기반으로 명명했다. 종명 ''howelli''는 켈레스비그-와에링과 스토르머에게 설명에 사용된 화석 표본을 대여해 준 프린스턴 대학교의 벤자민 하웰 박사를 기리기 위해 지어졌다.[9] 이 종은 2007년 노르웨이 고생물학자 O. 에릭 테틀리에 의해 ''Jaekelopterus''로 지정되어 ''야이켈롭테루스 하웰리''가 되었다.[10]
3. 특징
야이켈롭테루스는 프테리고투스과(Pterygotidae)에 속하며, 다른 프테리고투스과 유립테리드와 유사하게 확대되고 강력한 협각과 납작하고 확장된 미부(telson)를 가진다.[10][3] 협각은 먹이를 잡고 찢는 데 특화되어 있으며,[10] 미부는 수영할 때 방향타 역할을 했을 것으로 추정된다.[3]

3. 1. 크기

프테리고투스류 유르프테리드는 ''프테리고투스''(Pterygotus)와 ''아쿠티라무스''(Acutiramus)와 같이 가장 큰 유르프테리드 중 다수를 포함한다. ''야이켈롭테루스''(Jaekelopterus)와 그 친척 및 다른 거대한 고생대 무척추동물의 전례 없는 큰 크기에 기여했을 가능성이 있는 몇 가지 요인으로는 포식, 구애 행동, 경쟁 및 환경 자원이 제시되었다.[14]
호흡, 탈피의 에너지 비용, 이동 및 외골격의 실제 특성과 같은 요인은 절지동물의 크기를 제한한다.[1] 튼튼하고 단단하게 경화된 발톱 외에 프테리고투스류의 보존된 대부분의 큰 몸체 분절은 얇고 무광물화되어 있다. 심지어 등판(tergite)과 복판(sternite)(복부 분절의 표면을 형성하는 판)도 일반적으로 종이처럼 얇게 압착되어 보존되어 프테리고투스류는 매우 가벼운 구조였음을 시사한다.[1] 비슷한 경량화 적응은 거대한 지네류와 유사한 ''아르트로플레우라''(Arthropleura)와 같은 다른 고생대 거대 절지동물에서도 관찰할 수 있으며, 거대 절지동물 크기의 진화에 필수적이라고 제안되었다.[15] 가벼운 구조는 신체 크기를 제한하는 요인의 영향을 줄인다.[1]
''야이켈롭테루스'' 및 다른 거대 프테리고투스류 유르프테리드는 가장 큰 절지동물이지만, 경량 구조는 무게가 가장 무겁지 않았을 가능성을 의미했다. 다른 거대한 유르프테리드, 특히 2미터에 가까운 ''히베르토프테루스''(Hibbertopterus)와 같은 히베르토프테리데(Hibbertopteridae)의 몸이 깊은 보행 형태는 무게 면에서 프테리고투스류 및 다른 거대 절지동물과 경쟁하거나 능가했을 수 있다.[16]
'야이켈롭테루스 레나니아에'는 협각을 제외하고 약 2.33~2.59미터, 협각을 포함하면 최대 3.59미터까지 성장했을 것으로 추정된다. '야이켈롭테루스 하웰리'는 '야이켈롭테루스 레나니아에'보다 작지만, 최대 80센티미터까지 성장했을 것으로 추정된다.
미국 고생물학자 알렉산더 카이저(Alexander Kaiser)와 남아프리카 고생물학자 자코 클록(Jaco Klok)은 2008년에 ''야이켈롭테루스''의 거대한 크기 추정치가 과장되었다고 제안했다. 그들은 크기 추정치는 협각(chelicerae)과 몸길이 사이의 상대적 비율이 동물이 성숙함에 따라 동일하게 유지된다는 것을 가정한다고 지적했다. 치아(발톱의 톱니)는 양성 생장 이형성(더 큰 표본에서 비례적으로 더 큼)을 보이는 것으로 관찰되었으며, 카이저와 클록은 이것이 협각 전체에서 발생했을 수 있다고 제안한다. 또한, 너비가 27cm인 동일 종의 가장 큰 기절(다리 분절)은 총 최대 몸길이가 1.8미터에 불과함을 시사한다.[17] 양성 생장 이형성은 ''야이켈롭테루스''의 가장 가까운 친척을 포함하여 다른 유르프테리드 속의 유르프테리드 협각 전체에서 입증되지 않았다. 또한 원래 발견과 같은 지층에서 발견된 또 다른 발톱을 포함하여, 큰 협각과 비율이 유사한 ''J. rhenaniae''의 묘사되지 않은 표본이 몇 개 있다. 카이저와 클록에게 같은 해에 답변한 브래디, 포쉬만, 테틀리의 의견으로는 2.5미터 주변의 크기 추정치가 이 종의 최대 크기에 대한 가장 정확한 추정치로 남아 있다.[18]
3. 2. 협각과 미부
3. 3. 시각
야이켈롭테루스는 겹눈을 가지고 있었으며, 홑눈 사이 각도(IOA)가 낮아 높은 시력을 가졌을 것으로 추정된다.[21][22] 겹눈의 렌즈 수와 홑눈 간 각도(IOA)는 절지동물의 시력을 결정하는 주요 요소이며, 특히 낮은 IOA는 활동적인 포식자에게서 나타나는 특징이다.[22]
야이켈롭테루스의 겹눈에 대한 연구 결과, 평균 IOA(0.87°)는 현대 포식성 절지동물과 비슷하며, 나이가 들면서 시력이 증가하는 양상을 보였다.[19] 이는 성체 야이켈롭테루스가 더 깊은 물과 같은 어두운 환경에서 서식했음을 시사하며, 유해류가 짝짓기와 산란을 위해 연안 환경으로 이동했다는 흔적 화석 증거와도 일치한다.[19]
야이켈롭테루스는 포식 동물에게 전형적인 전면 중첩 시야, 즉 입체 시야를 가지고 있었다. 구조적으로 야이켈롭테루스의 눈은 말굽게의 눈과 거의 동일하며, 겹눈의 수용체 세포 패턴 역시 유사하지만 동일하지는 않아 특화된 시각 시스템을 가졌음을 알 수 있다.[20] 야이켈롭테루스의 광수용체는 약 70μm로, 사람(1-2μm)이나 대부분의 절지동물(1-2μm)보다 훨씬 크지만, 크기 면에서는 현대 말굽게의 광수용체와 일치한다.[20]
현대 말굽게의 눈은 가장자리 인식을 높이고 대비를 향상시키는 특징을 가지는데, 이는 낮고 흩어진 빛 조건에서 중요하다.[20] 야이켈롭테루스의 눈도 매우 유사하여, 어두운 환경에서 대비를 높여 먹이를 효과적으로 감지하는 데 유리했을 것으로 보인다.[20]
4. 분류


''야이켈롭테루스''는 프테리고투스상과 프테리고투스과에 속한다.[2][1][11] ''야이켈롭테루스''는 프테리고투스와 유사하며, 생식 부속지와 텔슨(미부)의 특징에서만 구별된다. 이러한 유사성은 일부 연구자들로 하여금 프테리고투스과가 속 수준에서 과도하게 분할되었는지에 대한 의문을 제기하게 했다. 2010년 미국의 고생물학자 제임스 C. 람스델과 데이비드 A. 레그는 ''야이켈롭테루스'', ''프테리고투스'', ''아쿠티라무스''가 생식 부속지의 유사성을 기반으로 서로 동의어일 수 있다고 제안했다.[12]
협각에서 차이점이 관찰되었지만, 1964년 찰스 D. 워터스턴은 유립테리드에서 속을 구분하는 근거로 협각의 형태가 생활 방식에 따라 달라지고 개체 발생 전반에 걸쳐 변화하기 때문에 의문을 제기했다. 람스델과 레그는 텔슨 형태가 유립테리드에서 속을 구별하는 데 사용될 수 있지만, ''야이켈롭테루스''의 삼각형 텔슨이 ''프테리고투스''와 ''아쿠티라무스''에 존재하는 노 모양 텔슨의 형태학적 범위 내에 있을 수 있다고 언급했다.[12]
2007년 영국 고생물학자 사이먼 J. 브래디, 독일 고생물학자 마르쿠스 포쉬만, O. 에릭 테틀리가 실시한 계통 발생 분석[1] 결과 ''야이켈롭테루스''는 프테리고투스과 내에서 가장 진화된 분류군 중 하나로 밝혀졌다.[1] 또한, 이들은 코프의 법칙에 따라 프테리고투스과가 시간에 따라 몸 크기가 커지는 경향을 보였을 가능성을 제시했다.[1][13]
아래 분기도는 가장 잘 알려진 9개의 프테리고투스과 종과 2개의 외군 분류군 (''슬리모니아 아큐미나타''와 ''휴밀러리아 소셜리스'')을 기반으로 한다.
| 프테리고투스상과 | |
|---|---|
| 휴밀러리아 소셜리스 (20 cm) | |
| 슬리모니아 아큐미나타 (100 cm) | |
| 프테리고투스과 | |
| 키우르콥테루스 벤트리코수스 (70 cm) | |
| 에레토프테루스 | 에레토프테루스 웨이랜드스미시 (60 cm) |
| 에레토프테루스 오실리엔시스 (90 cm) | |
| 에레토프테루스 세리카우다투스 (60 cm) | |
| 에레토프테루스 빌로부스 (70 cm) | |
| 프테리고투스 앵글리쿠스 (160 cm) | |
| '야이켈롭테루스 레나니아에' (250 cm) | |
| 아쿠티라무스 | 아쿠티라무스 마크로프탈무스 (200 cm) |
| 아쿠티라무스 보헤미쿠스 (210 cm) | |
4. 1. 계통 발생
''야이켈롭테루스''는 프테리고토이데아 상과의 프테리고티다에 과에 속한다.[2][1][11] ''야이켈롭테루스''는 프테리고투스와 유사하여 생식 부속지와 텔슨(미부)의 특징에서만 구별된다. 이러한 유사성은 일부 연구자들로 하여금 프테리고티드가 속 수준에서 과도하게 분할되었는지에 대한 의문을 제기하게 했다. 2010년, 미국의 고생물학자 제임스 C. 람스델과 데이비드 A. 레그는 ''야이켈롭테루스'', ''프테리고투스'', ''아쿠티라무스''가 생식 부속지의 유사성을 기반으로 서로 동의어일 수 있다고 제안했다.[12]
협각에서 차이점이 관찰되었지만, 1964년 찰스 D. 워터스턴은 유라프테리드에서 속을 구분하는 근거로 협각의 형태가 생활 방식에 따라 달라지고 개체 발생 전반에 걸쳐 변화하기 때문에 의문을 제기했다. 람스델과 레그는 텔슨 형태가 유라프테리드에서 속을 구별하는 데 사용될 수 있지만, ''야이켈롭테루스''의 삼각형 텔슨이 ''프테리고투스''와 ''아쿠티라무스''에 존재하는 노 모양 텔슨의 형태학적 범위 내에 있을 수 있다고 언급했다.[12]
2007년 영국 고생물학자 사이먼 J. 브래디, 독일 고생물학자 마르쿠스 포쉬만, O. 에릭 테틀리가 실시한 계통 발생 분석[1] 결과 ''야이켈롭테루스''는 프테리고티드 내에서 가장 진화된 분류군 중 하나로 밝혀졌다.[1] 또한, 이들은 코프의 법칙에 따라 프테리고투스과가 시간에 따라 몸 크기가 커지는 경향을 보였을 가능성을 제시했다.[1][13]
아래 분기도는 가장 잘 알려진 9개의 프테리고티드 종과 2개의 외군 분류군 (''슬리모니아 아큐미나타''와 ''휴밀러리아 소셜리스'')을 기반으로 한다.
| 프테리고티오이데아 | |
|---|---|
| 휴밀러리아 소셜리스 (20 cm) | |
| 슬리모니아 아큐미나타 (100 cm) | |
| 프테리고티다에 | |
| 키우르콥테루스 벤트리코수스 (70 cm) | |
| 에레토프테루스 | 에레토프테루스 웨이랜드스미시 (60 cm) |
| 에레토프테루스 오실리엔시스 (90 cm) | |
| 에레토프테루스 세리카우다투스 (60 cm) | |
| 에레토프테루스 빌로부스 (70 cm) | |
| 프테리고투스 앵글리쿠스 (160 cm) | |
| '야이켈롭테루스 레나니아에' (250 cm) | |
| 아쿠티라무스 | 아쿠티라무스 마크로프탈무스 (200 cm) |
| 아쿠티라무스 보헤미쿠스 (210 cm) | |
5. 고생물학

프테리고티과에 속하는 ''야이켈롭테루스''와 다른 유르프테리드의 형태와 신체 구조는 이들이 완전히 수생 생활에 적응했음을 시사한다. 브래디, 포쉬만, 테틀리는 2007년 연구에서 ''야이켈롭테루스''와 같은 크기와 구조를 가진 절지동물이 육지에서 걸을 수 있을 가능성은 매우 낮다고 보았다.[1] ''야이켈롭테루스''와 같은 유르프테리드는 종종 "바다 전갈"이라고 불리지만, ''야이켈롭테루스'' 화석이 발견된 퇴적층은 이들이 비해양 수생 환경에서 살았음을 시사한다. ''J. howelli'' 화석이 발견된 와이오밍주의 베어투스 버트 형성은 조용하고 얕은 기수역 환경으로 해석된다. 이 종은 다른 두 유르프테리드 종인 ''도르프테루스 앙구스티콜리스''와 ''스트로빌롭테루스 프린스토니''와 함께 발견되었다.[2] 라인란트에서 ''J. rhenaniae''를 산출하는 화석 유적지도 기수에서 담수에 이르는 얕은 수생 환경의 일부였던 것으로 해석된다.[7]
''야이켈롭테루스''의 협각은 확대되고 튼튼하며, 굽은 자유 분지(ramus)와 서로 다른 길이와 크기의 치상돌기를 가지고 있는데, 이는 현존하는 전갈과 갑각류에서 강력한 천공 및 파지 능력에 해당하는 적응이다. 유타주의 데본기에서 발견된 포라스피디과 무악어류 어류 ''레크리아스피스 파툴라'' 화석의 일부 천공 상처는 ''야이켈롭테루스 하웰리''에 의해 발생했을 가능성이 있다.[21] 최신 연구에 따르면 ''야이켈롭테루스''는 활동적이고 시각적인 포식자였다.[22] 다 자란 ''야이켈롭테루스''는 그들의 환경에서 최상위 포식자였으며, 더 작은 절지동물(식인 행위 포함)과 초기 척추동물을 잡아먹었을 것이다.[1]
강력하고 활동적인 포식자인 ''야이켈롭테루스''는 매우 민첩하고 기동성이 뛰어났을 것이다. ''야이켈롭테루스''와 다른 프테리고티드의 수영 노와 텔슨의 유체역학은 이 그룹의 모든 구성원이 공중 부양, 전진 운동, 빠른 회전을 할 수 있었음을 시사한다. 그들이 반드시 빠르게 헤엄치는 동물은 아니었지만, 석호와 강어귀와 같은 서식지에서 먹이를 쫓아갈 수 있었을 것이다.[3][19]
5. 1. 거대화
''야이켈롭테루스''를 포함한 프테리고투스류 유르프테리드는 프테리고투스(Pterygotus) 및 아쿠티라무스(Acutiramus)와 같이 가장 큰 유르프테리드 중 다수를 포함한다.[14] 이들의 거대한 크기에 영향을 미친 요인으로는 포식, 구애 행동, 경쟁, 환경 자원 등이 제시되었다.[14]
절지동물의 크기를 제한하는 요인으로는 호흡, 탈피의 에너지 비용, 이동, 외골격의 특성 등이 있다.[1] 그러나 프테리고투스류는 대부분 얇고 무광물화된 몸체 분절을 가지고 있으며, 등판(tergite)과 복판(sternite)도 종이처럼 얇게 보존되어 매우 가벼운 구조였음을 알 수 있다.[1] 이러한 경량화 적응은 거대한 지네류와 유사한 ''아르트로플레우라(Arthropleura)''와 같은 다른 고생대 거대 절지동물에서도 관찰되며, 거대 절지동물 크기 진화에 필수적인 요소로 여겨진다.[15] 가벼운 구조는 신체 크기를 제한하는 요인의 영향을 줄여 거대화를 가능하게 했을 것이다.[1]
''야이켈롭테루스'' 및 다른 거대 프테리고투스류 유르프테리드는 가장 큰 절지동물이지만, 경량 구조 때문에 무게가 가장 무겁지는 않았을 가능성이 있다.[16] 특히 2미터에 가까운 ''히베르토프테루스(Hibbertopterus)''와 같은 히베르토프테리데(Hibbertopteridae)의 몸이 깊은 보행 형태는 무게 면에서 이들을 능가했을 수도 있다.[16]
5. 2. 발생 과정
유해류는 다른 모든 절지동물과 마찬가지로 "령기"라고 불리는 일련의 단계를 거쳐 성숙하며, 이는 빠른 성장을 동반한 탈피 기간으로 구성된다.[2] 협각류(유해류, 말발굽게, 바다거미, 거미류와 같은 다른 유기체와 함께 ''Jaekelopterus''가 속한 그룹)는 일반적으로 직접 발달을 하며, 이는 부화 후에 극심한 형태적 변화가 없음을 의미한다.[2] 유해류의 발생 과정은 현대의 말발굽게와 대체로 유사하지만, 유해류는 진정한 직접 발달을 하며, 성체와 동일한 수의 부속지와 분절을 가지고 부화한다.[2]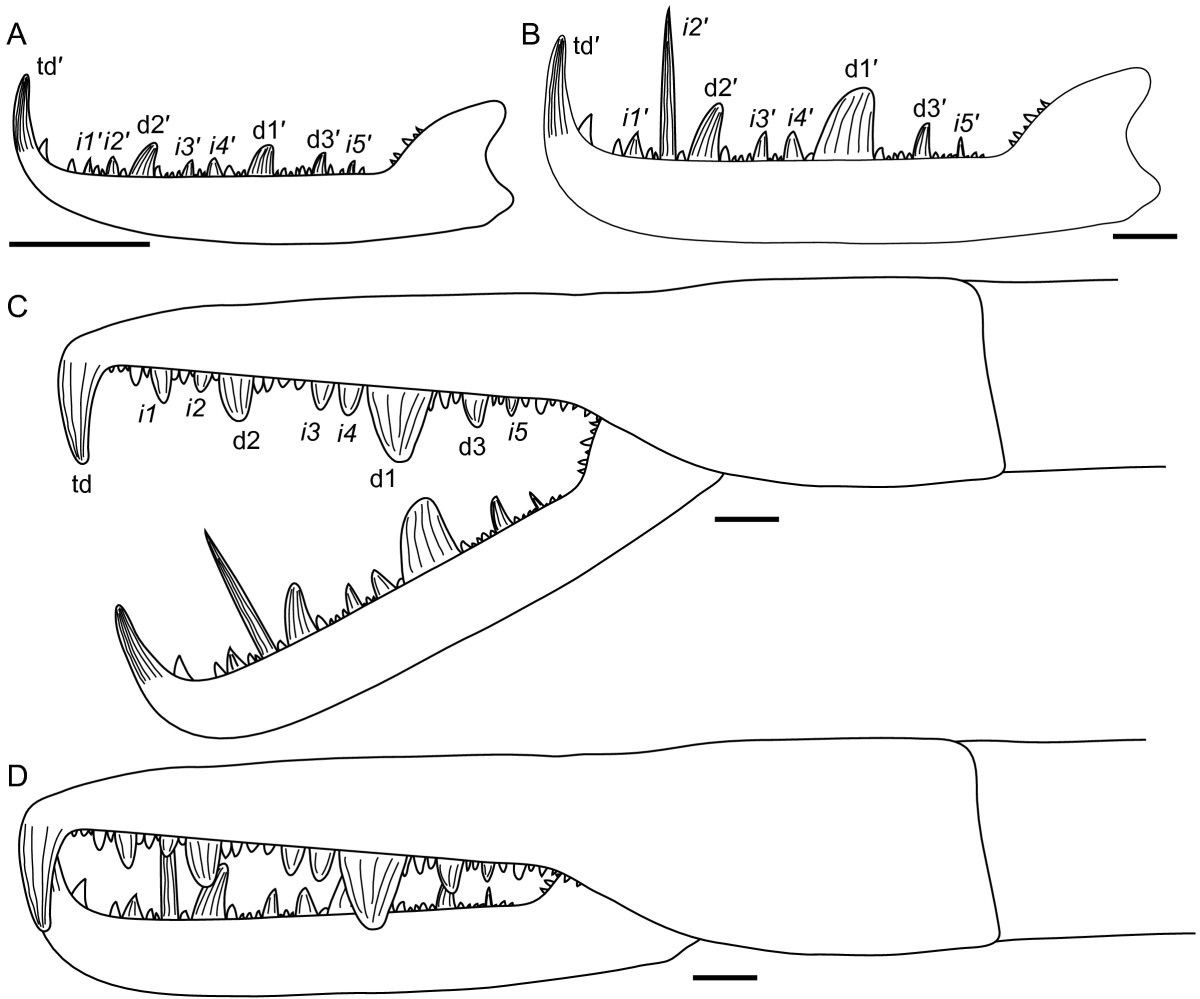
''Jaekelopterus howelli''의 여러 화석령기가 알려져 있지만, 표본의 단편적이고 불완전한 상태로 인해 그 발생 과정을 자세히 연구하기 어렵다.[2] 협각의 형태는 모든 연령대에서 치아의 배열과 수가 동일하여 유사하지만, 주요 치아는 중간 치아에 비해 크기가 커졌는데, 어린 개체에서는 중간 치아의 1.5배, 성체에서는 중간 치아의 최대 3.5배에 달했다.[2] 말단 치아는 어린 표본보다 성체 표본에서 훨씬 크고 견고했다.[2] 두 번째 중간 치아는 어린 개체에서 다른 중간 치아와 크기가 다르지 않지만, 성체에서는 엄청나게 길어져서 어떤 주요 치아보다 두 배 이상 길다.[2] 이러한 성장은 특히 ''Jaekelopterus'', 특히 ''J. howelli''에게 고유하다.[2]
''Jaekelopterus''의 후구개 또한 동물이 성숙함에 따라 크기가 변했다.[2] ''J. rhenaniae''에서 후구개의 상대적 너비는 발생 과정을 통해 감소했다.[2] ''J. howelli''의 후구개 또한 어린 개체보다 성체에서 더 넓지만, 어린 개체와 성체에서 측정된 길이-너비 비율은 큰 차이가 없었고, 어린 개체는 1.43, 성체는 1.46이었다.[2] 이러한 후구개 크기의 변화는 다른 유해류 속에서도 관찰되었다.[2]
5. 3. 식성
야이켈롭테루스의 협각은 확대되고 튼튼하며, 굽은 자유 분지(ramus)와 서로 다른 길이와 크기의 치상돌기를 가지고 있는데, 이는 현존하는 전갈과 갑각류에서 강력한 천공 및 파지 능력에 해당한다.[21] 유타주의 데본기에서 발견된 포라스피디과 무악어류 어류 ''레크리아스피스 파툴라'' 화석의 일부 천공 상처는 ''야이켈롭테루스 하웰리''에 의해 발생했을 가능성이 있다.[21] 최신 연구에 따르면 ''야이켈롭테루스''는 활동적이고 시각적인 포식자였다.[22] 다 자란 ''야이켈롭테루스''는 그들의 환경에서 최상위 포식자였으며, 더 작은 절지동물(식인 행위 포함)과 초기 척추동물을 잡아먹었을 것이다.[1]강력하고 활동적인 포식자인 ''야이켈롭테루스''는 매우 민첩하고 기동성이 뛰어났을 것이다. ''야이켈롭테루스''와 다른 프테리고티드의 수영 노와 텔슨의 유체역학은 이 그룹의 모든 구성원이 공중 부양, 전진 운동, 빠른 회전을 할 수 있었음을 시사한다.[3][19] 그들이 반드시 빠르게 헤엄치는 동물은 아니었지만, 석호와 강어귀와 같은 서식지에서 먹이를 쫓아갈 수 있었을 것이다.[3][19]
6. 한국과의 관계
참조
[1]
논문
Giant claw reveals the largest ever arthropod
[2]
논문
Babes in the wood – a unique window into sea scorpion ontogeny
[3]
논문
The pterygotid telson as a biological rudder
1988-01-01
[4]
논문
Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
[5]
논문
Ein grosser ''Pterygotus'' aus dem rheinischen Unterdevon
https://de.wikisourc[...]
2018-01-12
[6]
논문
Eurypteriden aus dem Rheinischen Unterdevon
https://books.google[...]
1936
[7]
논문
On the Emsian (Lower Devonian) arthropods of the Rhenish Slate Mountains: 5. Rare and poorly known eurypterids from Willwerath, Germany
2006-12-01
[8]
논문
II. Observations on Pterygotid Eurypterids
https://www.research[...]
1964-01-01
[9]
논문
A lower Devonian ''Pterygotus'' from Wyoming
1952
[10]
논문
Distribution and dispersal history of Eurypterida (Chelicerata)
http://fossilinsects[...]
[11]
논문
A summary list of fossil spiders and their relatives (version 16.0)
http://www.wsc.nmbe.[...]
2018-12-07
[12]
논문
An isolated pterygotid ramus (Chelicerata: Eurypterida) from the Devonian Beartooth Butte Formation, Wyoming
https://www.cambridg[...]
[13]
논문
Chapter 17: Gigantism, dwarfism, and Cope's rule: "nothing in evolution makes sense without a phylogeny"
2004-06-01
[14]
논문
Gigantism in Palaeozoic arthropods
1985
[15]
웹인용
Fossil giants and surviving dwarfs. Arthropleurida and Pselaphognatha (Atelocerata, Diplopoda): characters, phylogenetic relationships and construction
https://eurekamag.co[...]
Kraus, O., Brauckmann, C.
2003-08-26
[16]
논문
Hallipterus excelsior, a Stylonurid (Chelicerata: Eurypterida) from the Late Devonian Catskill Delta Complex, and its phylogenetic position in the Hardieopteridae
[17]
논문
Do giant claws mean giant bodies? An alternative view on exaggerated scaling relationships
2008-06-23
[18]
논문
Reply: giant claws and big bodies
2008-06-23
[19]
논문
Telltale eyes: the lateral visual systems of Rhenish Lower Devonian eurypterids (Arthropoda, Chelicerata) and their palaeobiological implications
2016-03-01
[20]
논문
Insights into the 400 million-year-old eyes of giant sea scorpions (Eurypterida) suggest the structure of Palaeozoic compound eyes
2019-11-28
[21]
논문
New poraspids (Agnatha, Heterostraci) from the Early Devonian of the western United States
2011
[22]
논문
All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids
2015-08-01
[23]
뉴스
Biology Letters abstract
http://www.journals.[...]
[24]
뉴스
BBC News, "Man-sized sea scorpion claw found"
http://news.bbc.co.u[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com